- Обмен нуклеотидов. Матричные биосинтезы

Содержание
- 2. Нуклеиновые кислоты (ДНК и РНК) – это биополимеры (полинуклеотиды), состоящие из мононуклеотидов, соединённых фосфодиэфирными связями. ДНК
- 3. Функции РНК: м-РНК является копией гена и матрицей (планом) для синтеза белка т-РНК осуществляет транспорт аминокислот
- 4. Строение мононуклеотида азотистое основание пентоза остаток фосфорной кислоты нуклеозид НУКЛЕОТИД
- 5. Пуриновые основания нуклеиновых кислот
- 6. Пиримидиновые основания нуклеиновых кислот
- 7. Углеводы нуклеотидов (пентозы) (РНК) (ДНК) рибоза дезоксирибоза
- 8. Гидролиз нуклеопротеинов
- 9. Катаболизм пуриновых оснований В норме содержание мочевой кислоты в крови 0,14-0,5 ммоль/л
- 10. Нарушения обмена пуринов
- 11. Ингибитор ксантиноксидазы
- 12. Распад пиримидиновых оснований
- 13. Особенности синтеза нуклеотидов Синтез идёт из обычных простых предшественников (ак, углекислого газа и т.п.) Синтезируются не
- 14. (фосфорибозил-пирофосфат) Синтез пуриновых нуклеотидов
- 15. Происхождение атомов пуринового кольца
- 16. Синтез пуриновых нуклеотидов
- 17. Синтез пиримидиновых нуклеотидов оротидинмонофосфат (ОМФ) уридинмонофосфат (УМФ) глу-NH2
- 18. Синтез пиримидиновых нуклеотидов
- 19. Синтез дезоксирибонуклеотидов
- 20. Ингибиторы синтеза нуклеиновых кислот
- 21. Нарушения обмена нуклеотидов Нарушение синтеза пиримидинов – оротацидурия – дефект ОМФ-декарбоксилазы (недостаток синтеза пиримидиновых нуклеотидов, снижение
- 22. основание фосфодиэфирная связь Первичная структура нуклеиновых кислот основание основание
- 23. Направление роста цепи 5'→3' Поступающий нуклеотид он х х х Растущая цепь НК Матрица – цепь
- 24. Биосинтез ДНК (репликация) является: матричным (матрица – обе нити ДНК) комплиментарным фрагментарным (нити ДНК синтезируются в
- 25. Полуконсервативность биосинтеза ДНК
- 27. Особенности репликации ДНК-полимеразы δ и ε не могут соединять между собой два мононуклеотида, а только достраивают
- 28. Этапы репликации 1. Инициация: Топоизомераза находит точку начала репликации, гидролизует одну фосфодиэфирную связь и даёт возможность
- 29. Этапы репликации ДНК-связывающие белки (SSB-белки) стабилизируют репликативную вилку, не давая восстанавливаться водородным связям между комплиментарными нуклеотидами
- 30. Инициация репликации ДНК-полимераза α ДНК-полимераза δ ДНК-полимераза ε
- 31. Ориджин репликации репликон Инициация репликации
- 32. 2. Элонгация ДНК-полимераза δ продолжает удлинять нить из дезоксирибонуклеотидов на лидирующей нити, а ДНК-полимераза ε –
- 33. Элонгация репликации ДНК-полимераза α (праймаза) ДНК-полимераза δ ДНК-полимераза ε
- 34. Элонгация репликации
- 35. Терминация ДНК-полимераза β (фермент репарации) удаляет праймеры и достраивает фрагменты ДНК ДНК-лигаза соединяет фрагменты между собой
- 36. Функции ДНК-полимеразы β
- 37. Репарация ДНК
- 38. Вторичная структура ДНК
- 39. Строение нуклеосомы
- 40. Для биосинтеза РНК (транскрипции) необходимы: МАТРИЦА – участок одной из нитей ДНК – (транскриптон) СТРОИТЕЛЬНЫЙ МАТЕРИАЛ:
- 41. Биосинтез РНК
- 42. Транскрибируемая нить Нетранскрибируемая нить РНК-полимераза ДНК пре-РНК (первичный транскрипт) Биосинтез РНК мононуклеотиды Сайт терминации
- 43. Процессинг РНК (1. сплайсинг)
- 44. м-РНК 7-метилгуанозин полиаденилат 5' 3' Процессинг (2. модификация концов м-РНК)
- 45. Состав зрелой м-РНК
- 46. Вторичная структура р-РНК
- 47. Вторичная структура т-РНК антикодон
- 48. Общая схема биосинтеза белка
- 49. Компоненты белоксинтезирующей системы мРНК 20 Аминокислот 20 Аминоацил-тРНК синтетаз (АРС-аз) Изоакцепторные тРНК Рибосомы в виде полисом
- 50. Ядерные стадии биосинтеза белка Биосинтез мРНК (транскрипция) Созревание мРНК (посттранскрипционный процессинг)
- 51. Функции т-РНК Акцепторная: связывает аминокислоту и транспортирует её к месту синтеза белка Адаптерная: встраивает аминокислоту на
- 52. Строение т-РНК антикодон
- 53. Таблица генетического кода
- 54. Характеристика генетического кода Триплетность (1 аминокислота кодируется 3 нуклеотидами) Специфичность (каждому кодону соответствует только 1 аминокислота)
- 55. Колинеарность генетического кода Глу-тРНК
- 56. Цитоплазматические стадии биосинтеза белка Активация аминокислот, или образование аминоацил-тРНК Инициация Элонгация Терминация Посттрансляционная модификация
- 57. Синтез аминоацил-тРНК
- 58. Рибосома эукариотов
- 59. Функции активной рибосомы Связывание и удерживание белоксинтезирующей системы Транслокация – перемещение м-РНК через рибосому каждый раз
- 60. Функционирующая рибосома
- 61. Образование инициирующего комплекса
- 62. Инициация: К малой субъединице рибосомы прикрепляется м-РНК К инициирующему кодону прикрепляется Мет-т-РНК К образовавшемуся комплексу присоединяется
- 63. Инициация трансляции
- 64. Элонгация: В пептидильном центре находится Мет-т-РНК, а в аминоацильном – аа-тРНК, соответствующая второму кодону. Мет отрывается
- 65. ГТФ ГДФ+НР
- 66. Строение полирибосомы
- 67. Посттрансляционный процессинг Модификация N-конца полипептидной цепи Фолдинг (формирование пространственной структуры) Химическая модификация (гидроксилирование, гликозилирование и др.)
- 68. Регуляция биосинтеза 1: РНК полимераза, 2: Репрессор, 3: Промотор, 4: Оператор, 5: Лактоза, 6, 7, 8:
- 69. Действие регуляторных белков
- 70. Регуляция биосинтеза Энхансеры – участки ДНК, присоединение к которым регуляторных белков усиливает транскрипцию Сайленсеры – участки
- 71. Амплификация
- 72. Типы генов в геноме Структурные гены (кодируют белки) Регуляторные гены: Гены-регуляторы (регулируют работу структурных генов) Процессинг-гены
- 73. Клеточная дифференцировка
- 74. Классификация мутаций
- 75. Типы мутаций
- 76. Типы генных мутаций
- 77. Проявления мутаций
- 79. Механизмы увеличения числа и разнообразия генов в геноме
- 80. Механизмы увеличения числа и разнообразия генов в геноме (кроссинговер)
- 81. Механизмы увеличения числа и разнообразия генов в геноме (амплификация)
- 82. Полиморфизм белков – изменение первичной структуры белка в пределах одного вида без изменения функций. В его
- 83. Схема возникновения наследственных болезней
- 84. Биохимические проявления наследственных болезней
- 86. Скачать презентацию

Нуклеиновые кислоты (ДНК и РНК)
– это биополимеры (полинуклеотиды), состоящие из мононуклеотидов,
Нуклеиновые кислоты (ДНК и РНК)
– это биополимеры (полинуклеотиды), состоящие из мононуклеотидов,

Функции РНК:
м-РНК является копией гена и матрицей (планом) для синтеза белка
т-РНК
Функции РНК:
м-РНК является копией гена и матрицей (планом) для синтеза белка
т-РНК

Строение мононуклеотида
азотистое основание
пентоза
остаток фосфорной кислоты
нуклеозид
НУКЛЕОТИД
Строение мононуклеотида
азотистое основание
пентоза
остаток фосфорной кислоты
нуклеозид
НУКЛЕОТИД
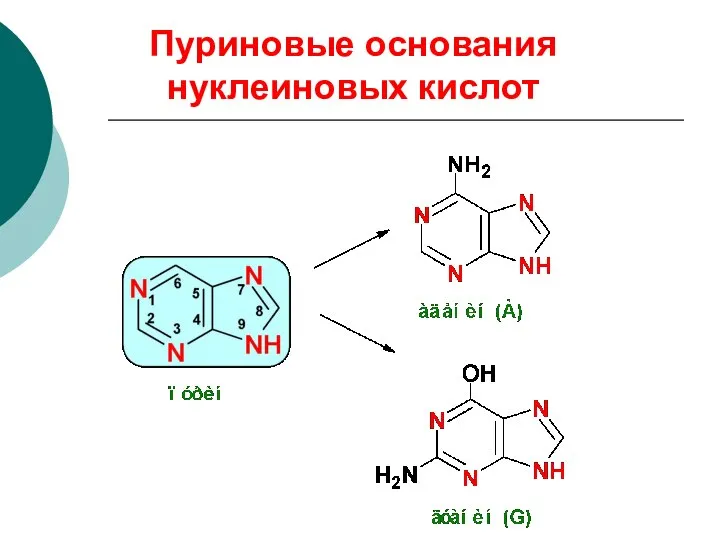
Пуриновые основания нуклеиновых кислот
Пуриновые основания нуклеиновых кислот

Пиримидиновые основания нуклеиновых кислот
Пиримидиновые основания нуклеиновых кислот

Углеводы нуклеотидов (пентозы)
(РНК) (ДНК)
рибоза дезоксирибоза
Углеводы нуклеотидов (пентозы)
(РНК) (ДНК)
рибоза дезоксирибоза
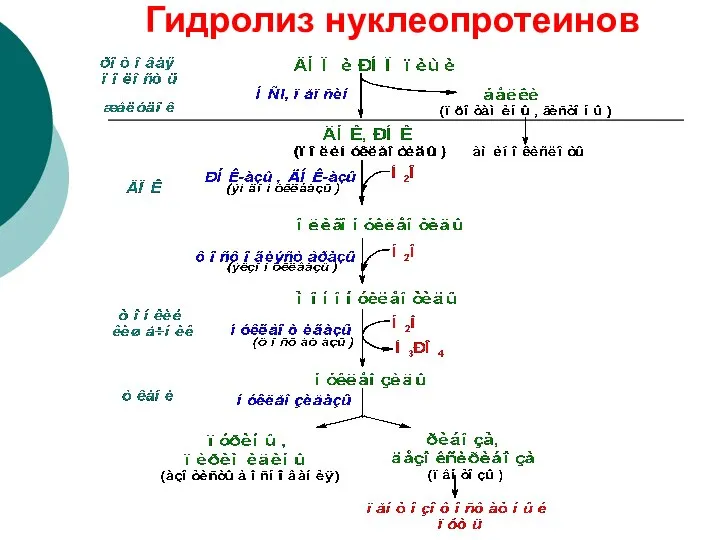
Гидролиз нуклеопротеинов
Гидролиз нуклеопротеинов

Катаболизм пуриновых оснований
В норме содержание мочевой кислоты в крови 0,14-0,5 ммоль/л
Катаболизм пуриновых оснований
В норме содержание мочевой кислоты в крови 0,14-0,5 ммоль/л

Нарушения обмена пуринов
Нарушения обмена пуринов

Ингибитор ксантиноксидазы
Ингибитор ксантиноксидазы

Распад пиримидиновых оснований
Распад пиримидиновых оснований

Особенности синтеза нуклеотидов
Синтез идёт из обычных простых предшественников (ак, углекислого газа
Особенности синтеза нуклеотидов
Синтез идёт из обычных простых предшественников (ак, углекислого газа
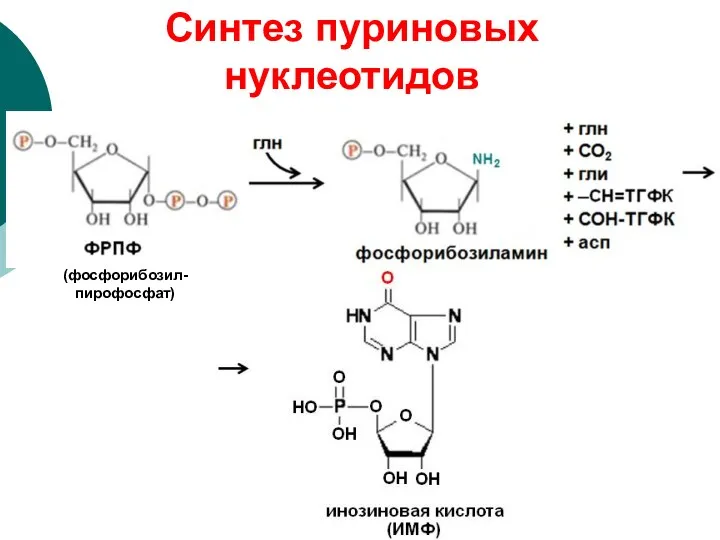
(фосфорибозил-пирофосфат)
Синтез пуриновых нуклеотидов
(фосфорибозил-пирофосфат)
Синтез пуриновых нуклеотидов

Происхождение атомов пуринового кольца
Происхождение атомов пуринового кольца
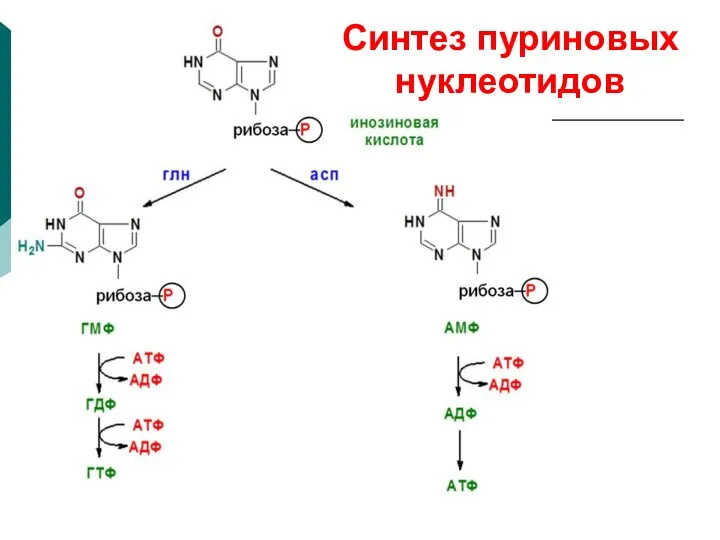
Синтез пуриновых нуклеотидов
Синтез пуриновых нуклеотидов
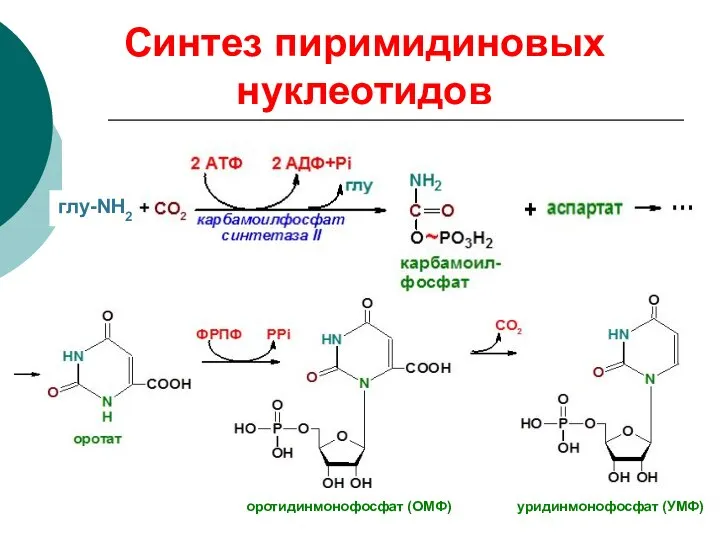
Синтез пиримидиновых нуклеотидов
оротидинмонофосфат (ОМФ) уридинмонофосфат (УМФ)
глу-NH2
Синтез пиримидиновых нуклеотидов
оротидинмонофосфат (ОМФ) уридинмонофосфат (УМФ)
глу-NH2
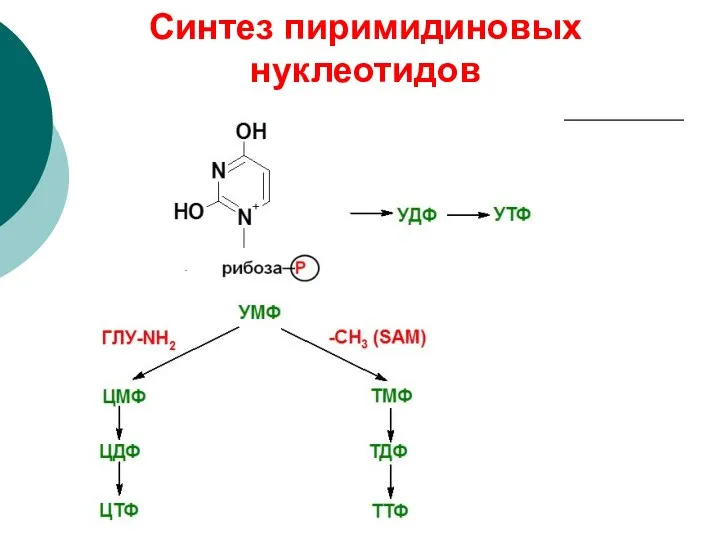
Синтез пиримидиновых нуклеотидов
Синтез пиримидиновых нуклеотидов
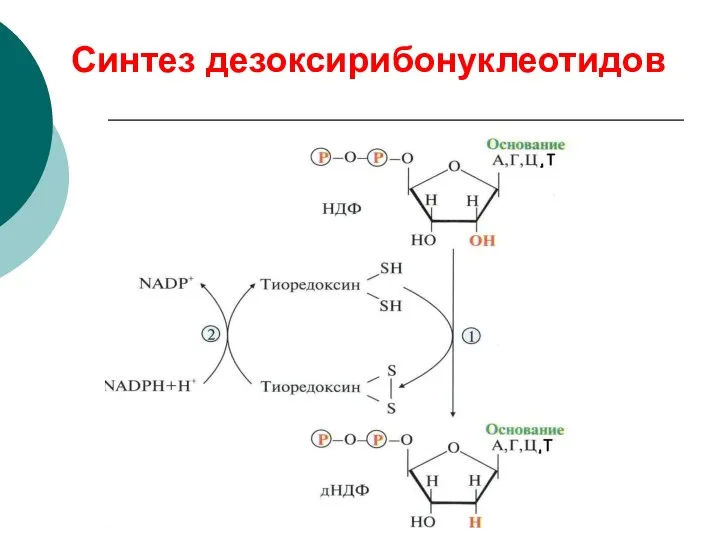
Синтез дезоксирибонуклеотидов
Синтез дезоксирибонуклеотидов
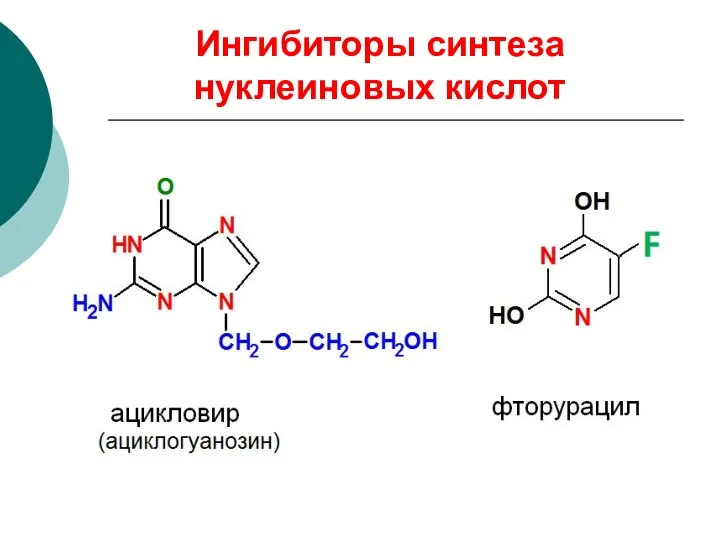
Ингибиторы синтеза нуклеиновых кислот
Ингибиторы синтеза нуклеиновых кислот

Нарушения обмена нуклеотидов
Нарушение синтеза пиримидинов – оротацидурия – дефект ОМФ-декарбоксилазы (недостаток
Нарушения обмена нуклеотидов
Нарушение синтеза пиримидинов – оротацидурия – дефект ОМФ-декарбоксилазы (недостаток

основание
фосфодиэфирная связь
Первичная структура нуклеиновых кислот
основание
основание
основание
фосфодиэфирная связь
Первичная структура нуклеиновых кислот
основание
основание

Направление роста цепи
5'→3'
Поступающий нуклеотид
он х
х
х
Растущая цепь НК
Матрица – цепь ДНК
5'
5'
3'
3'
Принцип биосинтеза
НК
Направление роста цепи
5'→3'
Поступающий нуклеотид
он х
х
х
Растущая цепь НК
Матрица – цепь ДНК
5'
5'
3'
3'
Принцип биосинтеза
НК

Биосинтез ДНК (репликация) является:
матричным (матрица – обе нити ДНК)
комплиментарным
фрагментарным (нити ДНК
Биосинтез ДНК (репликация) является:
матричным (матрица – обе нити ДНК)
комплиментарным
фрагментарным (нити ДНК
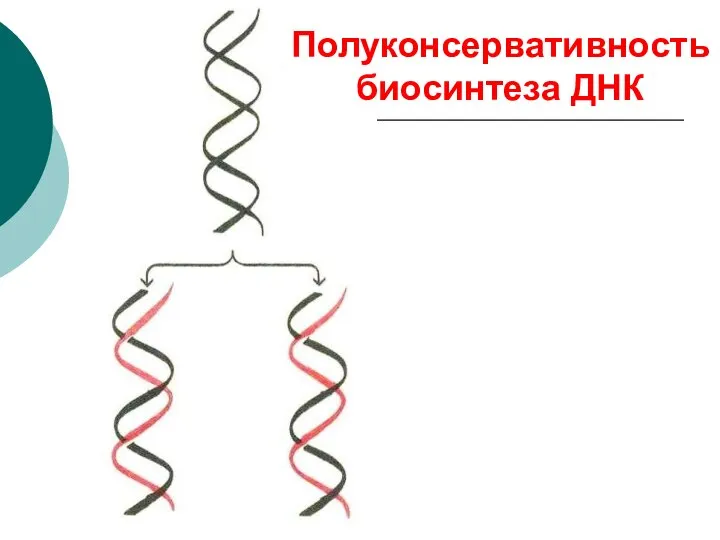
Полуконсервативность биосинтеза ДНК
Полуконсервативность биосинтеза ДНК


Особенности репликации
ДНК-полимеразы δ и ε не могут соединять между собой два
Особенности репликации
ДНК-полимеразы δ и ε не могут соединять между собой два

Этапы репликации
1. Инициация:
Топоизомераза находит точку начала репликации, гидролизует одну фосфодиэфирную связь
Этапы репликации
1. Инициация:
Топоизомераза находит точку начала репликации, гидролизует одну фосфодиэфирную связь

Этапы репликации
ДНК-связывающие белки (SSB-белки) стабилизируют репликативную вилку, не давая восстанавливаться водородным
Этапы репликации
ДНК-связывающие белки (SSB-белки) стабилизируют репликативную вилку, не давая восстанавливаться водородным

Инициация репликации
ДНК-полимераза α
ДНК-полимераза δ
ДНК-полимераза ε
Инициация репликации
ДНК-полимераза α
ДНК-полимераза δ
ДНК-полимераза ε
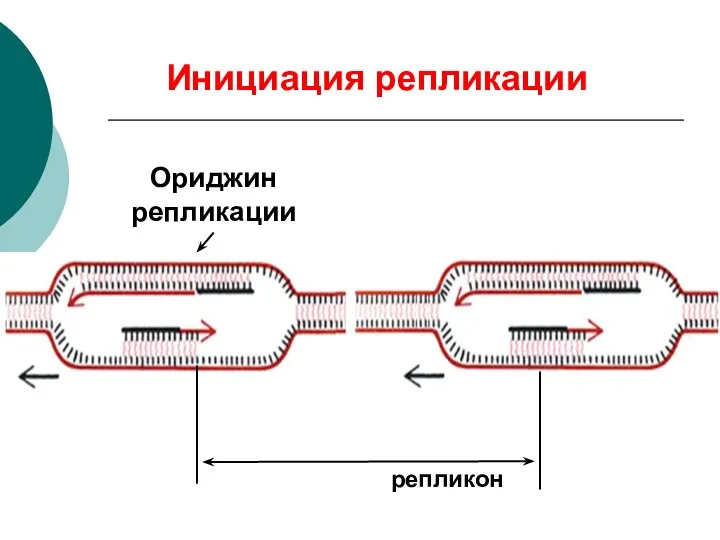
Ориджин репликации
репликон
Инициация репликации
Ориджин репликации
репликон
Инициация репликации

2. Элонгация
ДНК-полимераза δ продолжает удлинять нить из дезоксирибонуклеотидов на лидирующей нити,
2. Элонгация
ДНК-полимераза δ продолжает удлинять нить из дезоксирибонуклеотидов на лидирующей нити,

Элонгация репликации
ДНК-полимераза α (праймаза)
ДНК-полимераза δ
ДНК-полимераза ε
Элонгация репликации
ДНК-полимераза α (праймаза)
ДНК-полимераза δ
ДНК-полимераза ε

Элонгация репликации
Элонгация репликации

Терминация
ДНК-полимераза β (фермент репарации) удаляет праймеры и достраивает фрагменты ДНК
ДНК-лигаза
Терминация
ДНК-полимераза β (фермент репарации) удаляет праймеры и достраивает фрагменты ДНК
ДНК-лигаза
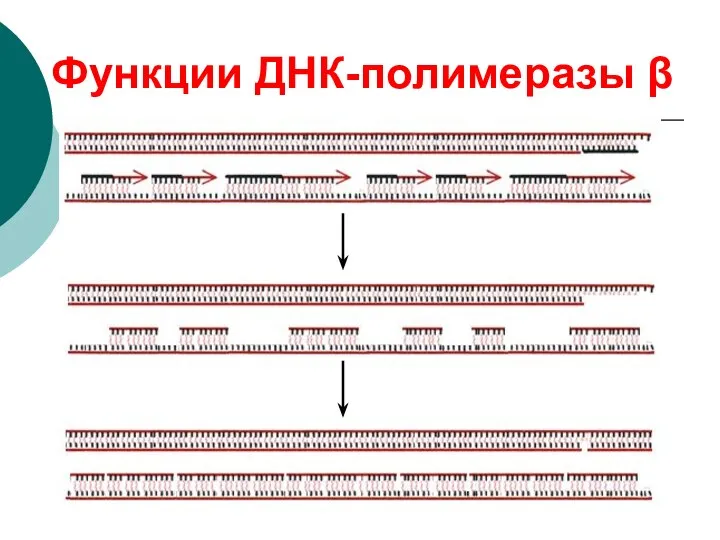
Функции ДНК-полимеразы β
Функции ДНК-полимеразы β

Репарация ДНК
Репарация ДНК

Вторичная структура ДНК
Вторичная структура ДНК
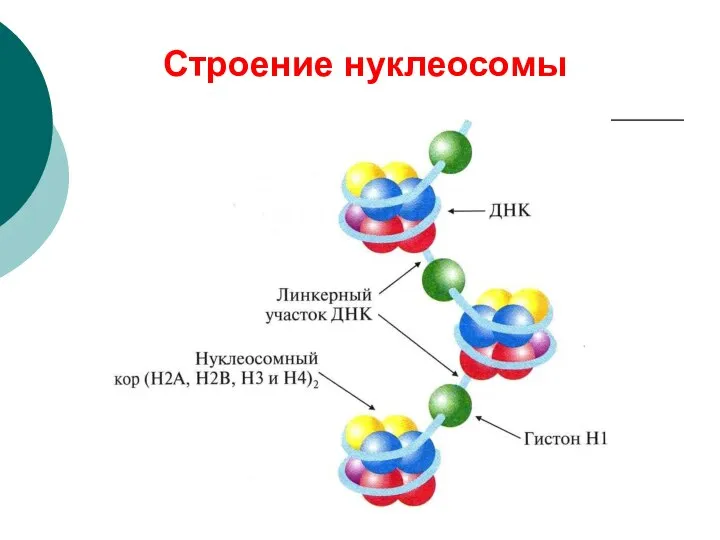
Строение нуклеосомы
Строение нуклеосомы

Для биосинтеза РНК (транскрипции) необходимы:
МАТРИЦА – участок одной из нитей ДНК
Для биосинтеза РНК (транскрипции) необходимы:
МАТРИЦА – участок одной из нитей ДНК
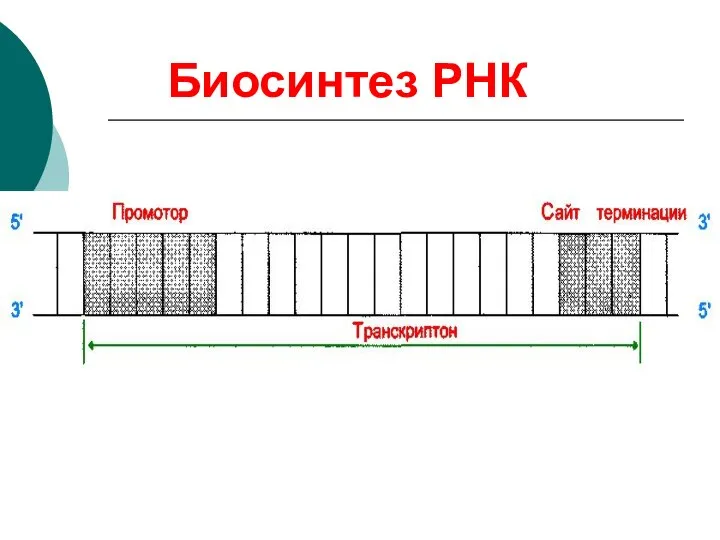
Биосинтез РНК
Биосинтез РНК

Транскрибируемая нить
Нетранскрибируемая нить
РНК-полимераза
ДНК
пре-РНК (первичный транскрипт)
Биосинтез РНК
мононуклеотиды
Сайт терминации
Транскрибируемая нить
Нетранскрибируемая нить
РНК-полимераза
ДНК
пре-РНК (первичный транскрипт)
Биосинтез РНК
мононуклеотиды
Сайт терминации
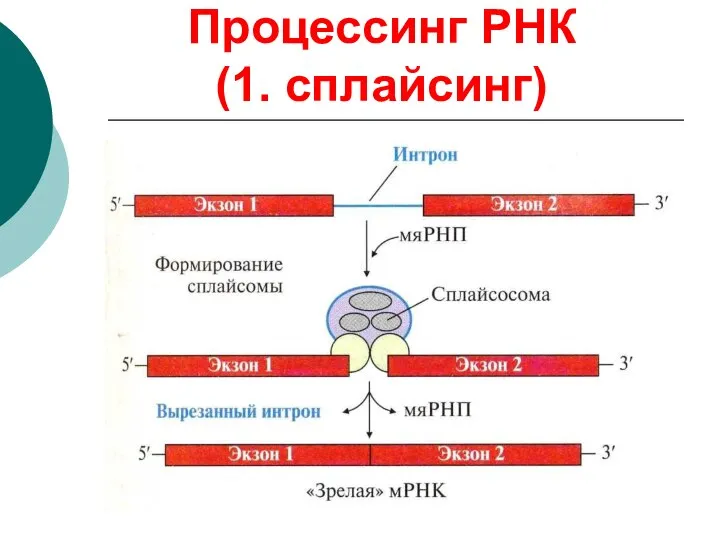
Процессинг РНК
(1. сплайсинг)
Процессинг РНК
(1. сплайсинг)

м-РНК
7-метилгуанозин
полиаденилат
5'
3'
Процессинг
(2. модификация концов м-РНК)
м-РНК
7-метилгуанозин
полиаденилат
5'
3'
Процессинг
(2. модификация концов м-РНК)

Состав зрелой м-РНК
Состав зрелой м-РНК
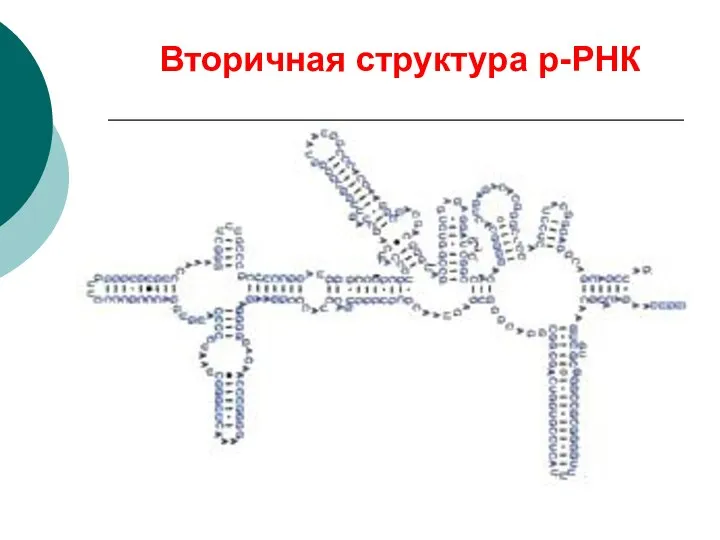
Вторичная структура р-РНК
Вторичная структура р-РНК

Вторичная структура т-РНК
антикодон
Вторичная структура т-РНК
антикодон

Общая схема биосинтеза белка
Общая схема биосинтеза белка

Компоненты белоксинтезирующей системы
мРНК
20 Аминокислот
20 Аминоацил-тРНК синтетаз (АРС-аз)
Изоакцепторные тРНК
Рибосомы в
Компоненты белоксинтезирующей системы
мРНК
20 Аминокислот
20 Аминоацил-тРНК синтетаз (АРС-аз)
Изоакцепторные тРНК
Рибосомы в

Ядерные стадии биосинтеза белка
Биосинтез мРНК (транскрипция)
Созревание мРНК (посттранскрипционный процессинг)
Ядерные стадии биосинтеза белка
Биосинтез мРНК (транскрипция)
Созревание мРНК (посттранскрипционный процессинг)

Функции т-РНК
Акцепторная: связывает аминокислоту и транспортирует её к месту синтеза белка
Адаптерная:
Функции т-РНК
Акцепторная: связывает аминокислоту и транспортирует её к месту синтеза белка
Адаптерная:

Строение т-РНК
антикодон
Строение т-РНК
антикодон

Таблица генетического кода
Таблица генетического кода

Характеристика генетического кода
Триплетность (1 аминокислота кодируется 3 нуклеотидами)
Специфичность (каждому кодону соответствует
Характеристика генетического кода
Триплетность (1 аминокислота кодируется 3 нуклеотидами)
Специфичность (каждому кодону соответствует

Колинеарность генетического
кода
Глу-тРНК
Колинеарность генетического
кода
Глу-тРНК

Цитоплазматические стадии биосинтеза белка
Активация аминокислот, или образование аминоацил-тРНК
Инициация
Элонгация
Терминация
Посттрансляционная модификация
Цитоплазматические стадии биосинтеза белка
Активация аминокислот, или образование аминоацил-тРНК
Инициация
Элонгация
Терминация
Посттрансляционная модификация

Синтез аминоацил-тРНК
Синтез аминоацил-тРНК

Рибосома эукариотов
Рибосома эукариотов

Функции активной рибосомы
Связывание и удерживание белоксинтезирующей системы
Транслокация – перемещение м-РНК через
Функции активной рибосомы
Связывание и удерживание белоксинтезирующей системы
Транслокация – перемещение м-РНК через
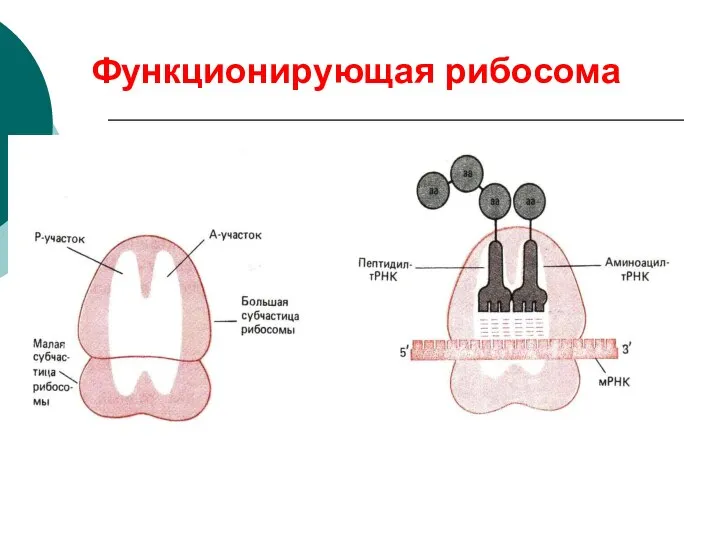
Функционирующая рибосома
Функционирующая рибосома

Образование инициирующего комплекса
Образование инициирующего комплекса

Инициация:
К малой субъединице рибосомы прикрепляется м-РНК
К инициирующему кодону прикрепляется Мет-т-РНК
К образовавшемуся
Инициация:
К малой субъединице рибосомы прикрепляется м-РНК
К инициирующему кодону прикрепляется Мет-т-РНК
К образовавшемуся
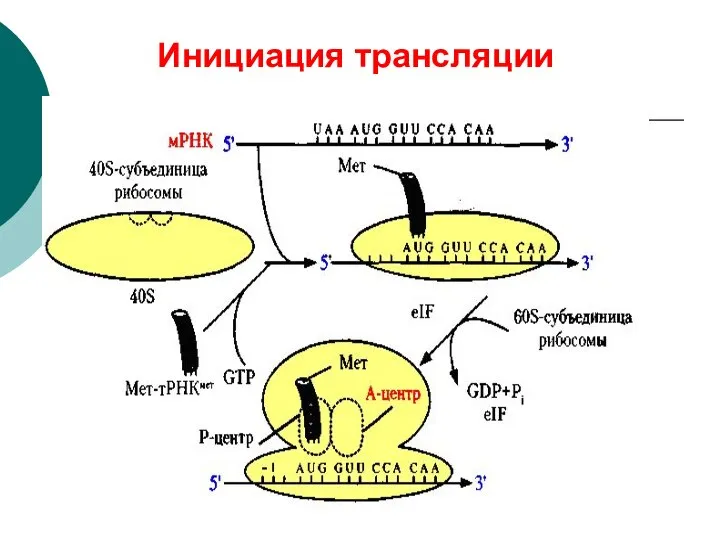
Инициация трансляции
Инициация трансляции

Элонгация:
В пептидильном центре находится Мет-т-РНК, а в аминоацильном – аа-тРНК, соответствующая
Элонгация:
В пептидильном центре находится Мет-т-РНК, а в аминоацильном – аа-тРНК, соответствующая
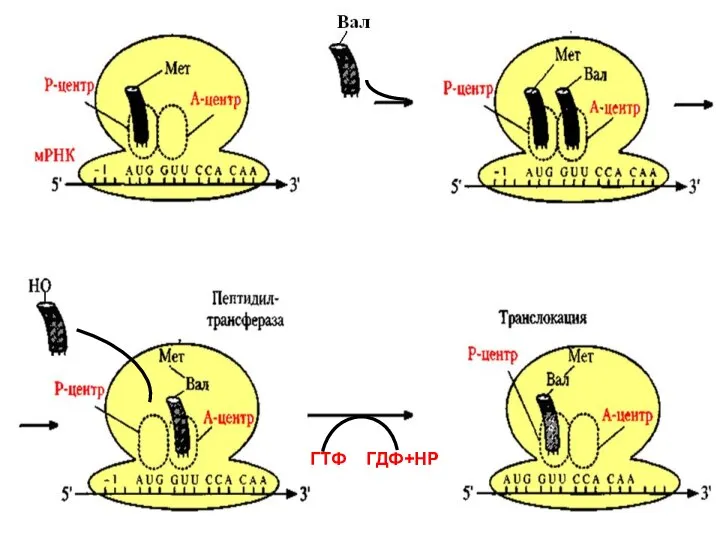
ГТФ ГДФ+НР
ГТФ ГДФ+НР
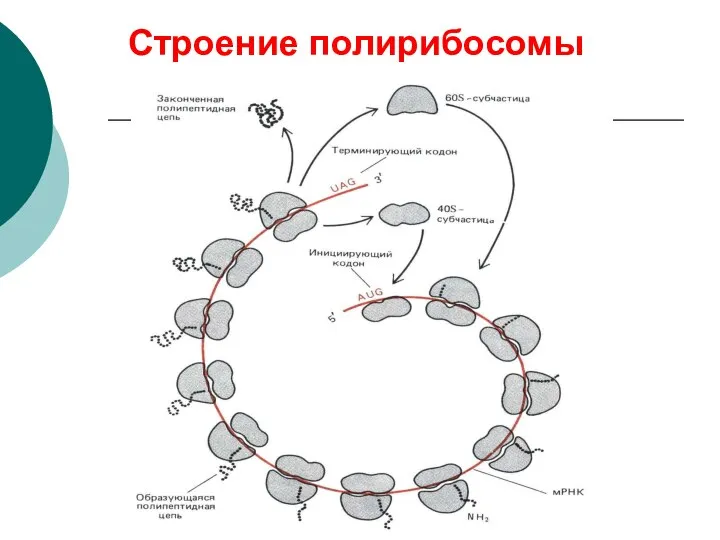
Строение полирибосомы
Строение полирибосомы

Посттрансляционный процессинг
Модификация N-конца полипептидной цепи
Фолдинг (формирование пространственной структуры)
Химическая модификация (гидроксилирование, гликозилирование
Посттрансляционный процессинг
Модификация N-конца полипептидной цепи
Фолдинг (формирование пространственной структуры)
Химическая модификация (гидроксилирование, гликозилирование

Регуляция биосинтеза
1: РНК полимераза, 2: Репрессор, 3: Промотор, 4: Оператор, 5:
Регуляция биосинтеза
1: РНК полимераза, 2: Репрессор, 3: Промотор, 4: Оператор, 5:
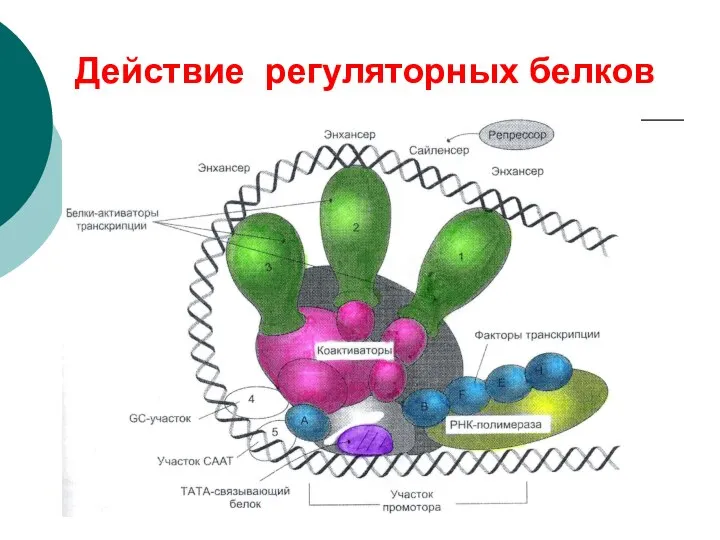
Действие регуляторных белков
Действие регуляторных белков

Регуляция биосинтеза
Энхансеры – участки ДНК, присоединение к которым регуляторных белков усиливает
Регуляция биосинтеза
Энхансеры – участки ДНК, присоединение к которым регуляторных белков усиливает
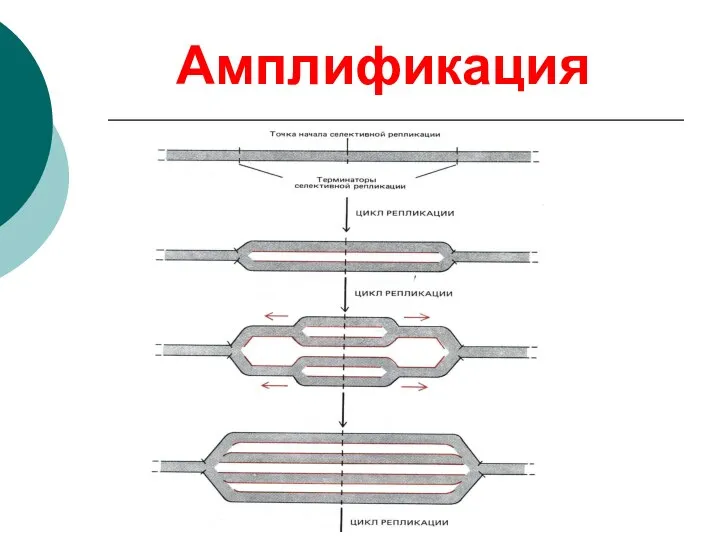
Амплификация
Амплификация

Типы генов в геноме
Структурные гены (кодируют белки)
Регуляторные гены:
Гены-регуляторы (регулируют работу
Типы генов в геноме
Структурные гены (кодируют белки)
Регуляторные гены:
Гены-регуляторы (регулируют работу

Клеточная дифференцировка
Клеточная дифференцировка

Классификация мутаций
Классификация мутаций

Типы мутаций
Типы мутаций

Типы генных мутаций
Типы генных мутаций

Проявления мутаций
Проявления мутаций

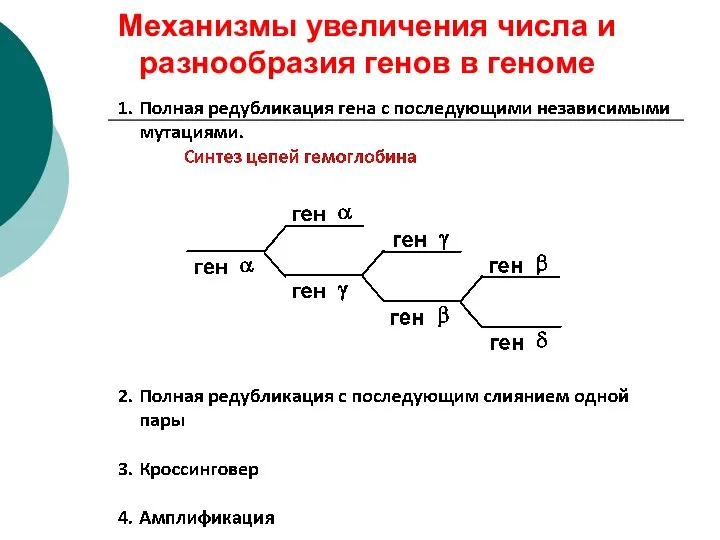
Механизмы увеличения числа и разнообразия генов в геноме
Механизмы увеличения числа и разнообразия генов в геноме

Механизмы увеличения числа и разнообразия генов в геноме
(кроссинговер)
Механизмы увеличения числа и разнообразия генов в геноме
(кроссинговер)

Механизмы увеличения числа и разнообразия генов в геноме
(амплификация)
Механизмы увеличения числа и разнообразия генов в геноме
(амплификация)

Полиморфизм белков –
изменение первичной структуры белка в пределах одного
Полиморфизм белков –
изменение первичной структуры белка в пределах одного
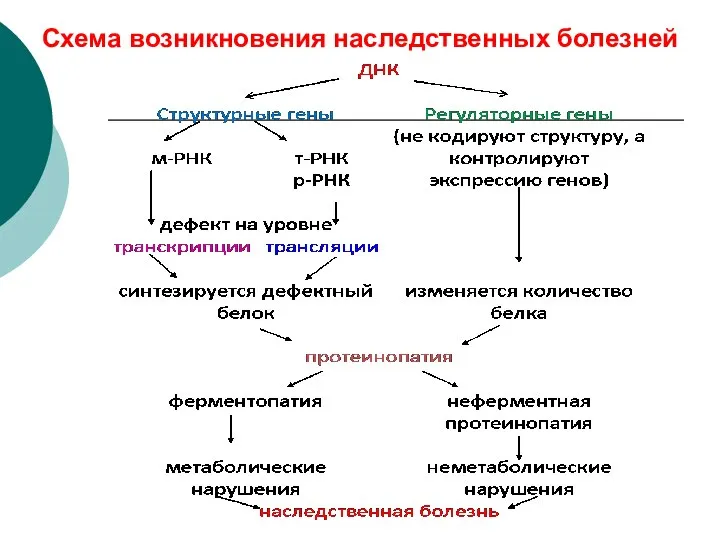
Схема возникновения наследственных болезней
Схема возникновения наследственных болезней

Биохимические проявления наследственных болезней
Биохимические проявления наследственных болезней
 Реакционная способность углеводородов
Реакционная способность углеводородов Неметаллы. Общая характеристика
Неметаллы. Общая характеристика Жесткость воды и способы ее устранения
Жесткость воды и способы ее устранения Основания LiOH - гидроксид лития
Основания LiOH - гидроксид лития Розв’язування задач і вправ з теми Алкани. 9 клас
Розв’язування задач і вправ з теми Алкани. 9 клас Аминдердің химиялық қасиеттері
Аминдердің химиялық қасиеттері Исследование соединений со структурой слоистого перовскита
Исследование соединений со структурой слоистого перовскита Химические реакции
Химические реакции Буферное кислотно - основное взаимодействие
Буферное кислотно - основное взаимодействие Неметаллы VIА-подгруппы
Неметаллы VIА-подгруппы Презентация по Химии "«Кислоты" - скачать смотреть бесплатно
Презентация по Химии "«Кислоты" - скачать смотреть бесплатно Строение и свойства циклоалканов
Строение и свойства циклоалканов Химия в повседневной жизни человека
Химия в повседневной жизни человека Жидкие системы
Жидкие системы Введение в химию. 8 класс
Введение в химию. 8 класс Простые вещества металлы
Простые вещества металлы Закон сохранения массы веществ
Закон сохранения массы веществ Аттестационная работа. Разработка по выполнению исследовательской работы. Выращивание кристаллов в домашних условиях
Аттестационная работа. Разработка по выполнению исследовательской работы. Выращивание кристаллов в домашних условиях Гормоны гипофиза. (Лекция 12)
Гормоны гипофиза. (Лекция 12) Алюминиевые сплавы
Алюминиевые сплавы Конструкционные функциональные волокнистые композиты
Конструкционные функциональные волокнистые композиты Organic compounds: nomenclature
Organic compounds: nomenclature Состав, строение, классификация кислот
Состав, строение, классификация кислот Основные понятия химии. Законы стехиометрии
Основные понятия химии. Законы стехиометрии Взаимодействие элементов-неметаллов между собой. 8 класс
Взаимодействие элементов-неметаллов между собой. 8 класс Изотопы, их свойства и применение
Изотопы, их свойства и применение Олиго- и гомополисахариды
Олиго- и гомополисахариды Геохимия природных процессов. Редкие элементы, как индикаторы геодинамических обстановок формирования комплексов. (Лекция 5)
Геохимия природных процессов. Редкие элементы, как индикаторы геодинамических обстановок формирования комплексов. (Лекция 5)