- БИОТЕХНОЛОГИЯ Гормональных препаратов

Содержание
- 4. Всего в организме вырабатывается более 100 гормонов и каждый из них играет очень важную роль в
- 6. ЛИПОФИЛЬНЫЕ ГОРМОНЫ КЛЕТКИ
- 7. ГИДРОФИЛЬНЫЕ ГОРМОНЫ
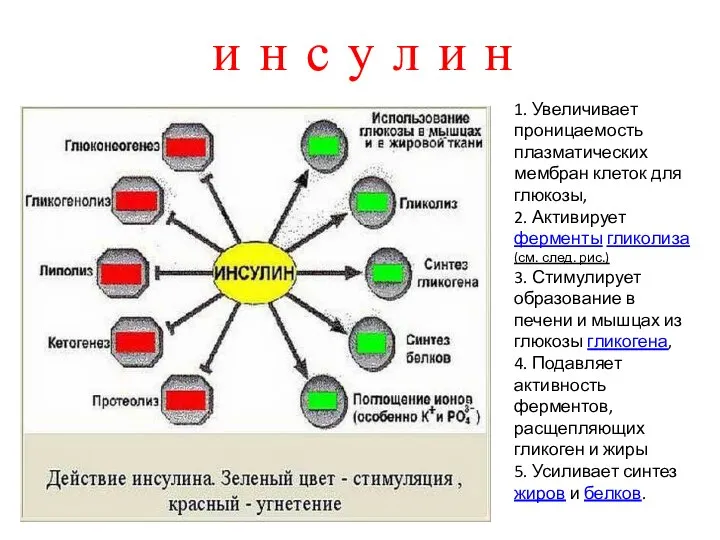
- 8. и н с у л и н 1. Увеличивает проницаемость плазматических мембран клеток для глюкозы, 2.
- 9. Ферменты гликолиза: 1 — Гексокиназа 2 — Глюкозо-6-фосфатизомераза 3 — 6-Фосфофруктокиназа 4 — Альдолаза 5 —
- 10. Инсулин вырабатывается в бета-клетках островков Лангерганса поджелудочной железы. 70% матричной РНК, выделенной из этих клеток, кодируют
- 11. ПОСЛЕДСТВИЯ ДЕФИЦИТА ИНСУЛИНА
- 12. Человеческий инсулин Инсулин это полипептид с м.м. 5808; - 51 аминокислота - 2 полипептидные цепи, соединенные
- 13. как происходит биосинтез инсулина в организме, в β-клетках островков Лангерганса
- 14. СИНТЕЗ ИНСУЛИНА В КЛЕТКАХ ПОДЖЕЛУДОЧНОЙ ЖЕЛЕЗЫ
- 15. 1 - синтез полипептидной цепи препроинсулина; 2 - синтез происходит на полирибосомах, прикрепленных к наружной поверхности
- 16. Биосинтез инсулина в β-клетках островков Лангерганса L-пептид β-пептид C-пептид ɑ-пептид 20 аминокислот 33 аминокислоты эндопептидаза эндопептидаза
- 17. МОНОМЕРЫ инсулина Структура кристалла инсулина, депонируемого в β-клетках островков ЛАНГЕРГАНСА. Image of six insulin molecules assembled
- 18. БИОСИНТЕЗ ЧЕЛОВЕЧЕСКОГО ИНСУЛИНА Ι. Замещение 1 аминокислоты в молекуле свиного инсулина ΙΙ. Синтез ɑ- и β-цепей
- 19. Преобразование инсулина свиней в инсулин человека (ала – у свиней) Моноинсулин Ферментативное замещение 30-й аминокислоты в
- 20. Микробиологический синтез инсулина: Химическим путем создают синтетический ген проинсулина. На N-конце гена - кодон ТAС (метионин),
- 21. Очистка синтетического инсулина Важно очистить инсулин проинсулина, который индуцируют выработку антиинсулиновых антител. Используется хроматографическая очистка: Ионообменная,
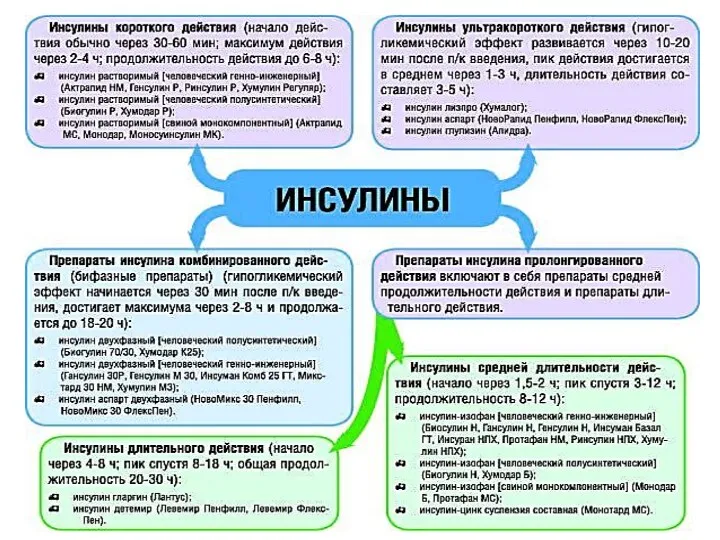
- 22. Стандартизация инсулина По чистоте классифицируют препараты инсулина на 4 группы, в зависимости от содержания проинсулина: обычные,
- 24. Соматотропный гормон
- 25. Обратитмся к литературе. И на сей раз к приключенческой. Но вовсе не замысловатый сюжет повести Еремея
- 26. «Открытие» соматотропина произошло задолго до XX столетия. Правда, врачи и алхимики средневековья не предполагали, какое именно
- 27. Ну зачем же так категорично ставить вопрос, возможно, поправит меня читатель, ведь никто не запрещает специалистам
- 28. Рукотворный синтезатор белков (метод Меррифилда — твердофазного синтеза белков. Вспомните, ученый «пришивал» к поверхности твердого носителя
- 29. Структура гормона передней доли гипофиза (СТГ),
- 30. СОМАТОТРОПНЫЙ ГОРМОН (СТГ) Состоит из 191 амк. Регулирует рост человека, применяется и для лечения ожогов, переломов,
- 31. Схема экспрессии чужеродных генов довольно проста, но на практике возникают следующие проблемы. Регуляторные сигналы эукариот сильно
- 32. Получение гонадотропного гормона в клетках Е. Coli разработано К. Итакурой и Г. Бойером. Аминокислотная последовательность этого
- 33. Получение гонадотропного гормона соматостатина через гибридный белок - бета-галактозидазу В образующемся химерном белке соматостатин отделен от
- 34. Эритропоэтин
- 35. Структура молекулы эритропоэтина (схема)
- 37. Скачать презентацию

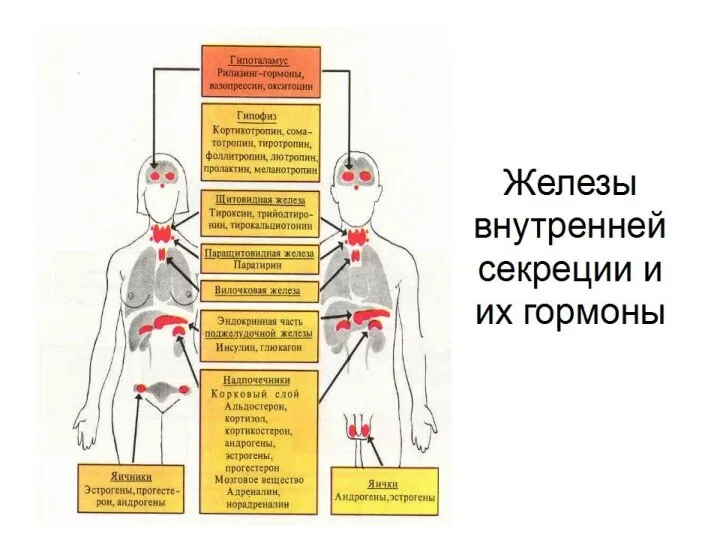
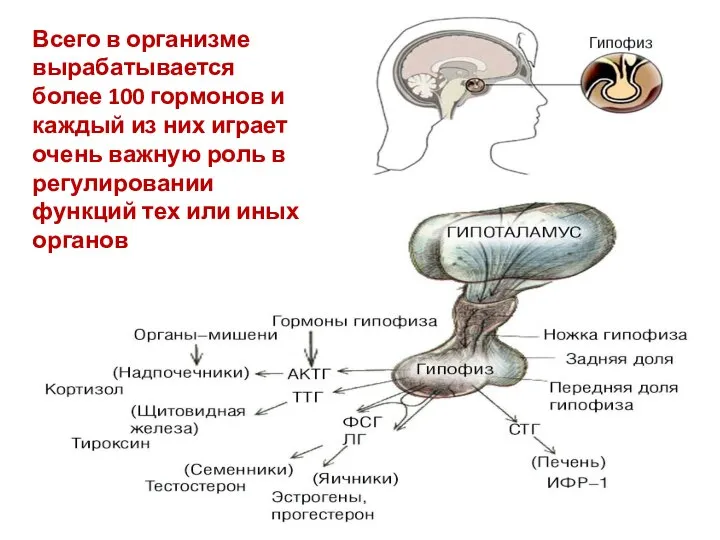
Всего в организме вырабатывается более 100 гормонов и каждый из них
Всего в организме вырабатывается более 100 гормонов и каждый из них

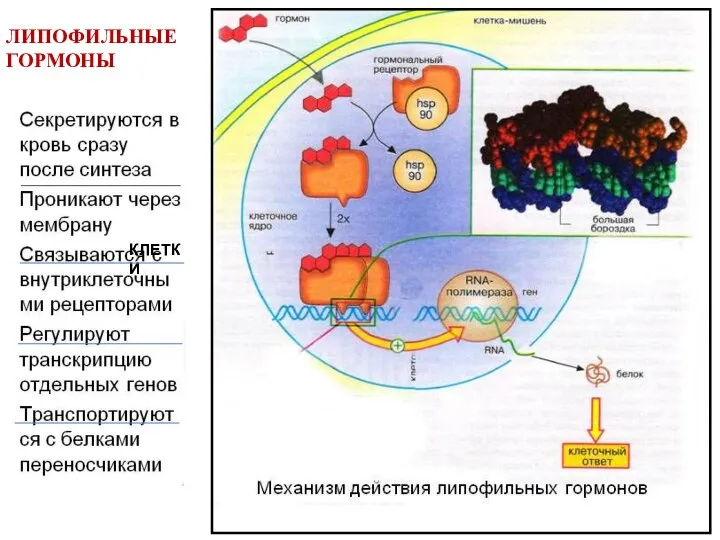
ЛИПОФИЛЬНЫЕ
ГОРМОНЫ
КЛЕТКИ
ЛИПОФИЛЬНЫЕ
ГОРМОНЫ
КЛЕТКИ
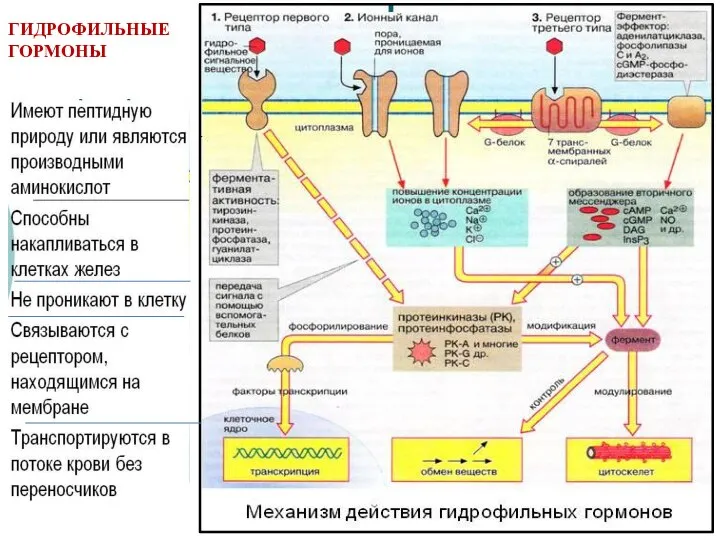
ГИДРОФИЛЬНЫЕ ГОРМОНЫ
ГИДРОФИЛЬНЫЕ ГОРМОНЫ
и н с у л и н
1. Увеличивает проницаемость плазматических мембран
и н с у л и н
1. Увеличивает проницаемость плазматических мембран
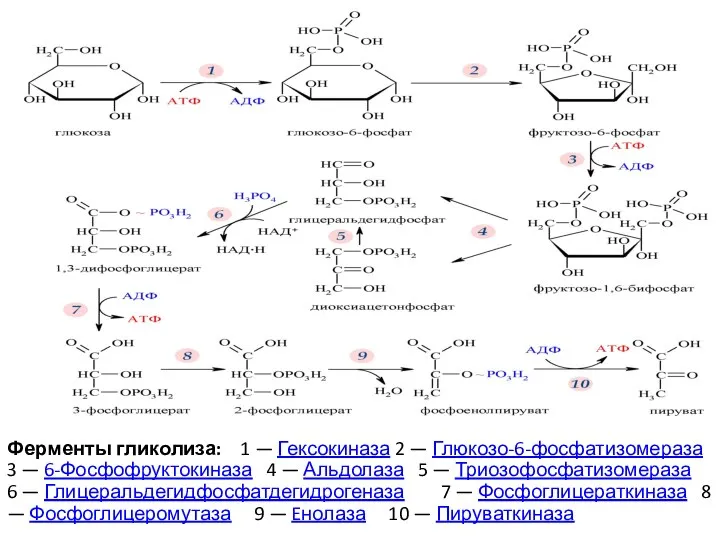
Ферменты гликолиза: 1 — Гексокиназа 2 — Глюкозо-6-фосфатизомераза 3 — 6-Фосфофруктокиназа
Ферменты гликолиза: 1 — Гексокиназа 2 — Глюкозо-6-фосфатизомераза 3 — 6-Фосфофруктокиназа
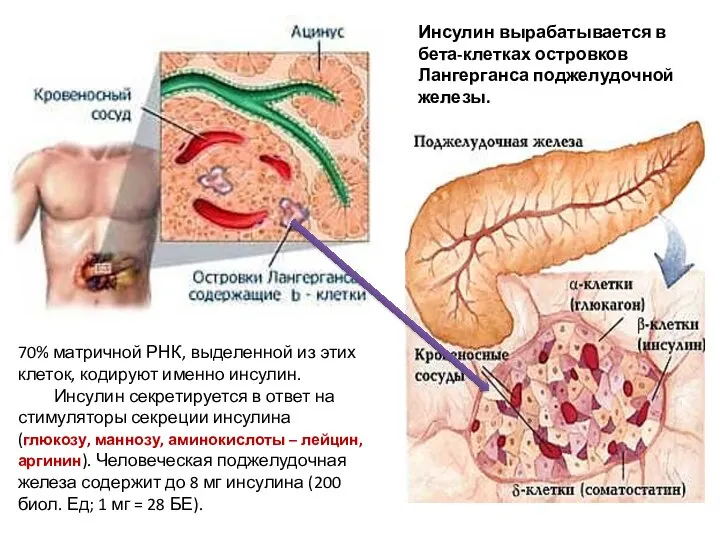
Инсулин вырабатывается в бета-клетках островков Лангерганса поджелудочной железы.
70% матричной РНК,
Инсулин вырабатывается в бета-клетках островков Лангерганса поджелудочной железы.
70% матричной РНК,
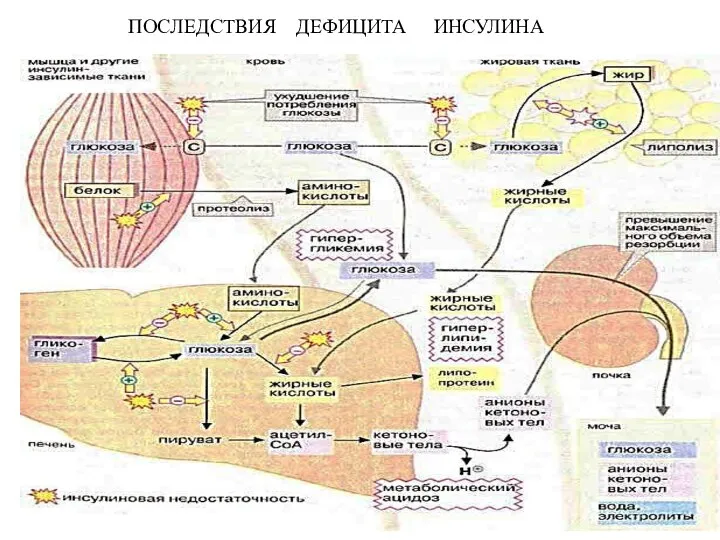
ПОСЛЕДСТВИЯ ДЕФИЦИТА ИНСУЛИНА
ПОСЛЕДСТВИЯ ДЕФИЦИТА ИНСУЛИНА
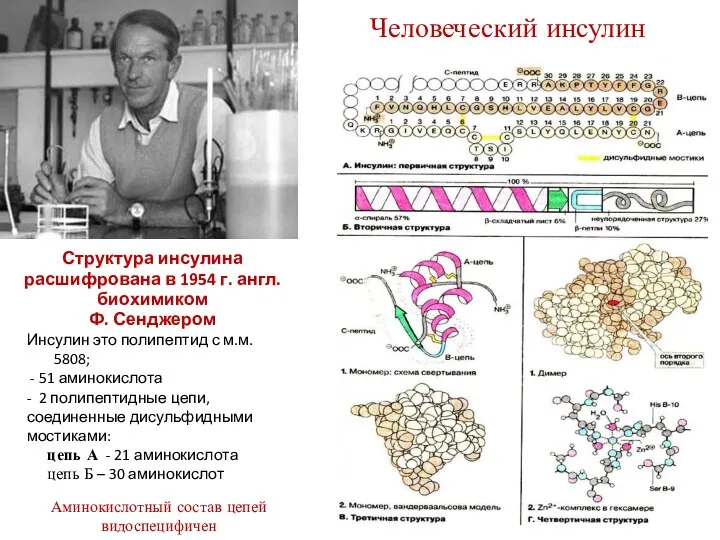
Человеческий инсулин
Инсулин это полипептид с м.м. 5808;
- 51 аминокислота
-
Человеческий инсулин
Инсулин это полипептид с м.м. 5808;
- 51 аминокислота
-
как происходит биосинтез инсулина в организме, в β-клетках островков Лангерганса
как происходит биосинтез инсулина в организме, в β-клетках островков Лангерганса

СИНТЕЗ ИНСУЛИНА В КЛЕТКАХ ПОДЖЕЛУДОЧНОЙ ЖЕЛЕЗЫ
СИНТЕЗ ИНСУЛИНА В КЛЕТКАХ ПОДЖЕЛУДОЧНОЙ ЖЕЛЕЗЫ
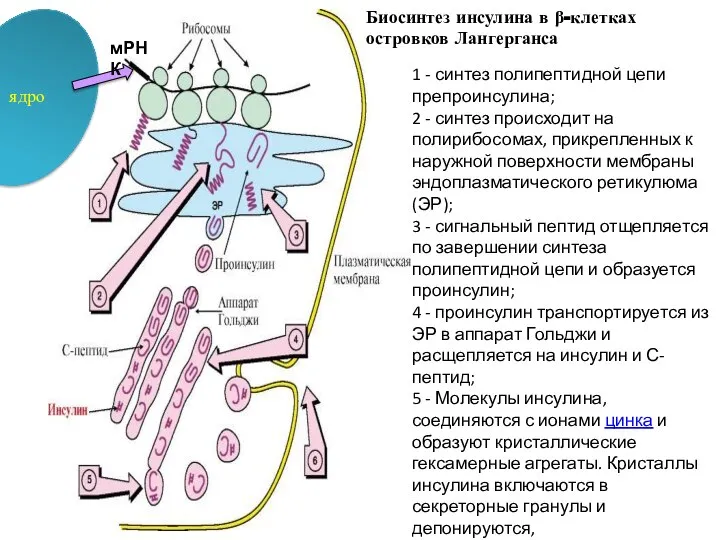
1 - синтез полипептидной цепи препроинсулина;
2 - синтез происходит на
1 - синтез полипептидной цепи препроинсулина;
2 - синтез происходит на
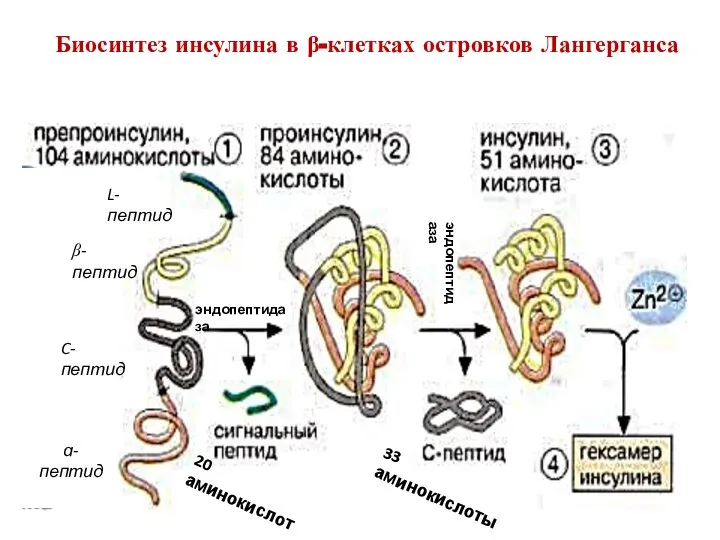
Биосинтез инсулина в β-клетках островков Лангерганса
L-пептид
β-пептид
C-пептид
ɑ-пептид
20 аминокислот
33 аминокислоты
эндопептидаза
эндопептидаза
Биосинтез инсулина в β-клетках островков Лангерганса
L-пептид
β-пептид
C-пептид
ɑ-пептид
20 аминокислот
33 аминокислоты
эндопептидаза
эндопептидаза
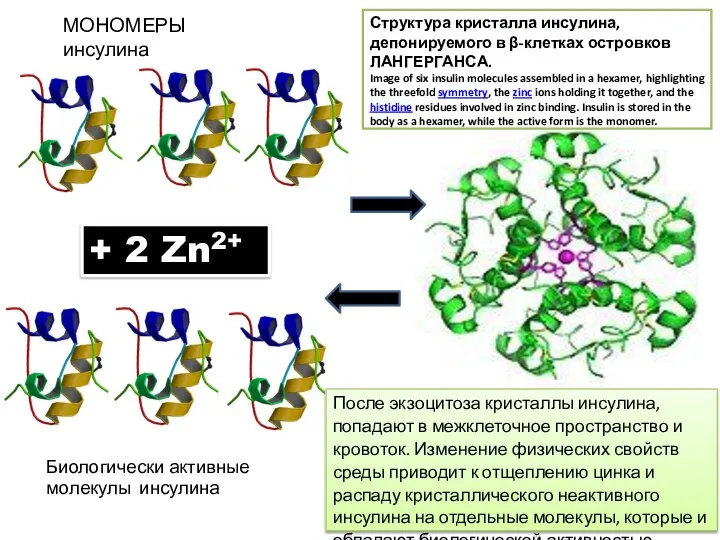
МОНОМЕРЫ инсулина
Структура кристалла инсулина, депонируемого в β-клетках островков ЛАНГЕРГАНСА.
Image of
МОНОМЕРЫ инсулина
Структура кристалла инсулина, депонируемого в β-клетках островков ЛАНГЕРГАНСА.
Image of

БИОСИНТЕЗ ЧЕЛОВЕЧЕСКОГО ИНСУЛИНА
Ι. Замещение 1 аминокислоты в молекуле свиного инсулина
ΙΙ. Синтез
БИОСИНТЕЗ ЧЕЛОВЕЧЕСКОГО ИНСУЛИНА
Ι. Замещение 1 аминокислоты в молекуле свиного инсулина
ΙΙ. Синтез

Преобразование инсулина свиней в инсулин человека
(ала – у свиней)
Моноинсулин
Ферментативное
Преобразование инсулина свиней в инсулин человека
(ала – у свиней)
Моноинсулин
Ферментативное
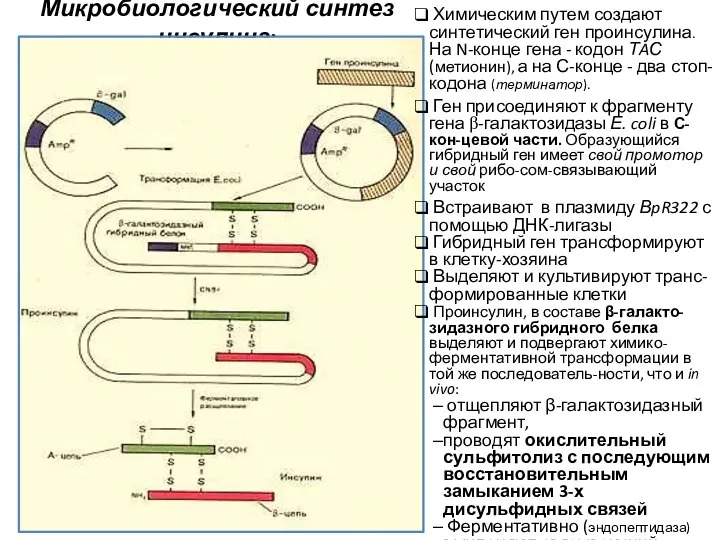
Микробиологический синтез инсулина:
Химическим путем создают синтетический ген проинсулина. На N-конце
Микробиологический синтез инсулина:
Химическим путем создают синтетический ген проинсулина. На N-конце

Очистка синтетического инсулина
Важно очистить инсулин проинсулина, который индуцируют выработку антиинсулиновых антител.
Очистка синтетического инсулина
Важно очистить инсулин проинсулина, который индуцируют выработку антиинсулиновых антител.

Стандартизация инсулина
По чистоте классифицируют препараты инсулина на 4 группы, в зависимости
Стандартизация инсулина
По чистоте классифицируют препараты инсулина на 4 группы, в зависимости

Соматотропный гормон
Соматотропный гормон

Обратитмся к литературе. И на сей раз к приключенческой. Но вовсе
Обратитмся к литературе. И на сей раз к приключенческой. Но вовсе

«Открытие» соматотропина произошло задолго до XX столетия. Правда, врачи и алхимики
«Открытие» соматотропина произошло задолго до XX столетия. Правда, врачи и алхимики

Ну зачем же так категорично ставить вопрос, возможно, поправит меня читатель,
Ну зачем же так категорично ставить вопрос, возможно, поправит меня читатель,

Рукотворный синтезатор белков (метод Меррифилда — твердофазного синтеза белков. Вспомните, ученый
Рукотворный синтезатор белков (метод Меррифилда — твердофазного синтеза белков. Вспомните, ученый

Структура гормона передней доли гипофиза (СТГ),
Структура гормона передней доли гипофиза (СТГ),

СОМАТОТРОПНЫЙ ГОРМОН (СТГ)
Состоит из 191 амк. Регулирует рост человека, применяется и
СОМАТОТРОПНЫЙ ГОРМОН (СТГ)
Состоит из 191 амк. Регулирует рост человека, применяется и

Схема экспрессии чужеродных генов довольно проста, но на практике возникают следующие
Схема экспрессии чужеродных генов довольно проста, но на практике возникают следующие

Получение гонадотропного гормона в клетках Е. Coli разработано К. Итакурой и
Получение гонадотропного гормона в клетках Е. Coli разработано К. Итакурой и
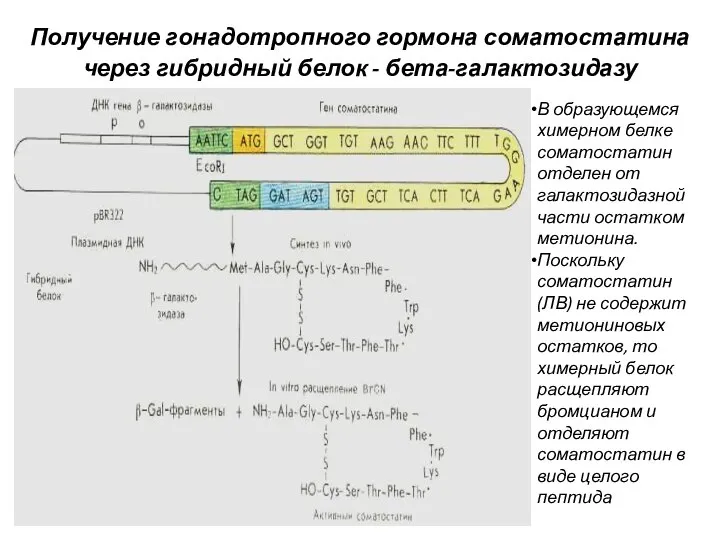
Получение гонадотропного гормона соматостатина через гибридный белок - бета-галактозидазу
В образующемся химерном
Получение гонадотропного гормона соматостатина через гибридный белок - бета-галактозидазу
В образующемся химерном

Эритропоэтин
Эритропоэтин
Структура молекулы эритропоэтина (схема)
Структура молекулы эритропоэтина (схема)
 Курсовая работа «Религиозный радикализм: государственно-общественное регулирование»
Курсовая работа «Религиозный радикализм: государственно-общественное регулирование» Компьютерная графика
Компьютерная графика Органы мочевой системы
Органы мочевой системы  Benjamin Franklin (1706-1790)
Benjamin Franklin (1706-1790) Аудитория Уанета Январь 2011 г. gemiusAudience
Аудитория Уанета Январь 2011 г. gemiusAudience An american summer
An american summer Презентация "Театр и музыка Древней Греции (10 класс)" - скачать презентации по МХК
Презентация "Театр и музыка Древней Греции (10 класс)" - скачать презентации по МХК Введение в направление подготовки: местное самоуправление
Введение в направление подготовки: местное самоуправление Изобразительное искусство романтизма
Изобразительное искусство романтизма Основные особенности управленческой деятельности Планирование воспитательной деятельности в образовательном учреждении
Основные особенности управленческой деятельности Планирование воспитательной деятельности в образовательном учреждении Техническое обслуживание и ремонт газораспределительного механизма двигателя А-41
Техническое обслуживание и ремонт газораспределительного механизма двигателя А-41 150 культур Дона
150 культур Дона Религия. Иудаизм
Религия. Иудаизм Звіт про роботу в конкурсах - презентация для начальной школы_
Звіт про роботу в конкурсах - презентация для начальной школы_ запросы в БД
запросы в БД  Гимнастика. Виды гимнастики
Гимнастика. Виды гимнастики Понятие и признаки правоотношений
Понятие и признаки правоотношений Государственные символы России. Герб. Флаг. Гимн России
Государственные символы России. Герб. Флаг. Гимн России Экспериментальный сад «Лоулайн Лэб» (Lowline Lab)
Экспериментальный сад «Лоулайн Лэб» (Lowline Lab) Гимнастика, часть физической культуры
Гимнастика, часть физической культуры Негізгі және қосымша шағымдарын анықтау
Негізгі және қосымша шағымдарын анықтау Презентация по алгебре Линейные уравнения с одной переменной 7 класс
Презентация по алгебре Линейные уравнения с одной переменной 7 класс Dativ
Dativ Basics of software development O. Fedorova, associate professor of Department PMI
Basics of software development O. Fedorova, associate professor of Department PMI ПРЕЗЕНТАЦИЯ НА ТЕМУ: «СИСТЕМА ВЫСШЕГО ОБРАЗОВАНИЯ ВЕЛИКОБРИТАНИИ».
ПРЕЗЕНТАЦИЯ НА ТЕМУ: «СИСТЕМА ВЫСШЕГО ОБРАЗОВАНИЯ ВЕЛИКОБРИТАНИИ». Зачем нужен менеджер продукта при живом проектировщике интерфейсов? Денис Бесков / http://beskov.ru World Usability Day, Москва, 2011
Зачем нужен менеджер продукта при живом проектировщике интерфейсов? Денис Бесков / http://beskov.ru World Usability Day, Москва, 2011 Канализация электроэнергии на напряжении выше 1 кВ
Канализация электроэнергии на напряжении выше 1 кВ Презентация "Рембрандт ван Рейн (1606-1669)" - скачать презентации по МХК
Презентация "Рембрандт ван Рейн (1606-1669)" - скачать презентации по МХК