- Биохимия мышечной ткани

Содержание
- 3. Сократительная система
- 4. Организация скелетных мышц позвоночных Сократительные элементы, саркомеры, состоят из двух типов параллельных нитей, толстых филаментов миозина
- 6. В количественном отношении наиболее важным белком миофибрилл является миозин (~65% мышечного белка). Молекула миозина построена из
- 7. Главным белком тонких нитей является актин (42 кДа, ~20-25% мышечного белка). Фибриллярный F-актин является важным структурным
- 9. Механизм сокращения мышечных волокон Сокращение мышечных волокон обусловлено продольным скольжением толстых миозиновых и тонких актиновых филаментов
- 10. В отсутствие АТФ (АТР), т. е. в исходном состоянии, головки молекул миозина прочно связаны с актиновыми
- 11. Регуляция сокращения мышечных волокон
- 13. Электромеханическое сопряжение Сокращением мышечного волокна управляют двигательные нейроны, которые выделяют нейромедиатор ацетилхолин в нервно-мышечные соединении (синапсы).
- 15. Саркоплазматический ретикулум Саркоплазматический ретикулум [СР (SR)] – разветвленная подобная эндоплазматическому ретикулуму органелла, окружающая индивидуальные миофибриллы подобно
- 16. Переносу потенциала действия на СР индивидуальной миофибриллы способствуют поперечные трубочки Т-системы, представляющие трубчатые впячивания клеточной мембраны
- 18. В расслабленной скелетной мышце комплекс тропонина (субъединицы = Т, С, I) с тропомиозином препятствует взаимодействию миозиновых
- 19. Таким образом, при сокращении мышечного волокна скелетных мышц позвоночных происходит следующая последовательность событий. При поступлении сигнала
- 20. Источники энергии
- 21. Энергетический обмен в мышечной ткани Важнейшей функцией мышечного волокна является сократительная. Процесс сокращения и расслабления связан
- 22. Потребности работающей мышцы в АТФ удовлетворяются за счет следующих ферментативных реакций: 1. Резерв в виде креатинфосфата.
- 23. 2 Анаэробный гликолиз. В мышечной ткани наиболее важным долгосрочным энергетическим резервом является гликоген. В покоящейся ткани
- 24. 4. Образование инозинмонофосфата [ИМФ (IMP)]. Другим источником быстрого восстановления уровня АТФ является конверсия АДФ в АТФ
- 25. Метаболическая регуляция мышечного сокращения
- 27. В клетках, не содержащих митохондрий (например, в эритроцитах), или в тканях при недостаточном снабжении кислородом (например,
- 28. Обратная связь, подобная циклу Кори, существует в цикле аланина, в котором также участвует пируват. Цикл аланина
- 30. Метаболизм белков и аминокислот Скелетные мышцы активно участвуют в метаболизме аминокислот. Это наиболее важный участок деградации
- 31. Синтез и расщепление мышечных белков контролируются гормонами. Тестостерон и синтетические анаболики стимулируют биосинтез белка; напротив, кортизол
- 32. Белки мышц Белки саркоплазмы миоглобин, белки–ферменты Белки миофибрилл Сократительные белки: миозин, актин, актомиозин Регуляторные белки: тропомиозин,
- 33. Миоглобин Дыхательный белок мышц Гемопротеин Содержит 153 аминокислотных остатка Основная функция – перенос кислорода в мышцах
- 35. Миозин Составляет 50 – 55% от массы миофибрилл Фибриллярный белок Период полураспада – 20 дней Состоит
- 36. Актин Составляет 20% от массы миофибрилл Небольшой глобулярный белок Состоит из 1 полипептидной цепи (374 аминокислотных
- 37. актиновая нить
- 38. Тропомиозин фибриллярный белок состоит из 2 α-спиралей на 1 молекулу тропомиозина приходится 7 молекул актина молекула
- 39. Тропонин Глобулярный белок В состав входят 3 субъединицы: Тн-Т – тропомиозинсвязывающая субъединица – отвечает за связь
- 40. В мышцах присутствует фетальная форма миозина, обладающая измененной структурой, сниженной АТФ-азной активностью С возрастом в мышцах
- 41. Биосинтез и распад креатина Креатин и креатининфосфат – важные азотистые вещества мышцы. Находится креатин в мышцах,
- 42. Синтез креатина Первая стадия синтеза креатина протекает в почках под действием глицин-амидинотрансферазы. Арг Гли Гуанидинуксусная кислота
- 43. Вторая стадия – метилирование- протекает в печени. Гуанидинуксусная кислота S-аденозил метионин S-аденозил гомоцистеин Гуанидинацетат метилтрансфераза Креатин
- 44. В мышцах имеется высокоэнергетическое вещество – креатинфосфат. Креатин КФК + АТФ + АДФ Креатинфосфат
- 45. Креатинин образуется в результате неферментативного дефосфорилирования креатинфосфата. Фн Н20 Креатинфосфат Креатинин
- 46. Содержание в плазме крови В плазме крови в небольших количествах содержатся креатин и креатинин. Содержание креатинина
- 47. Диагностическое значение уровень креатинина в сыворотке – чувствительный показатель состояния функции почек, снижение выделение креатинина с
- 48. Физиологическая креатинурия наблюдается у новорожденных, так как преобладает скорость синтеза креатина, у пожилых вследствие атрофии мышц,
- 50. Скачать презентацию

Сократительная система
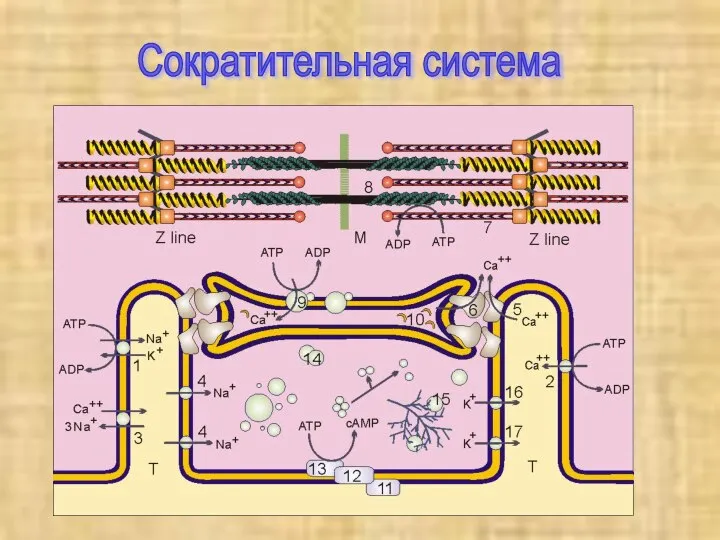
Сократительная система
Организация скелетных мышц позвоночных
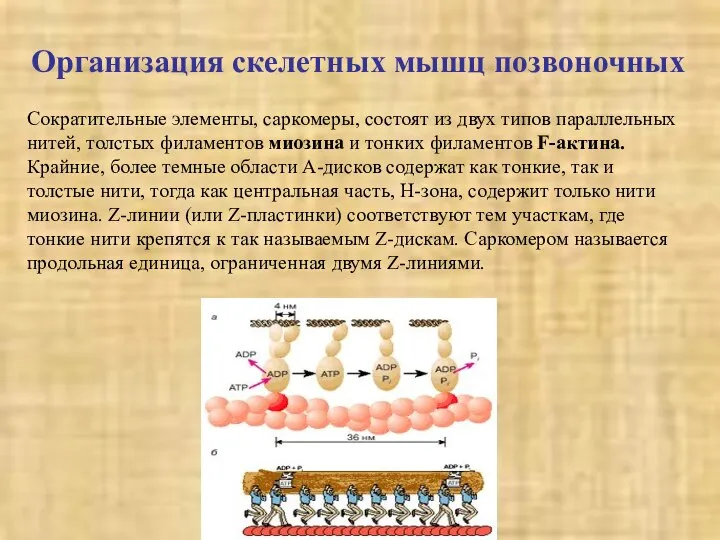
Сократительные элементы, саркомеры, состоят из двух типов параллельных
Организация скелетных мышц позвоночных
Сократительные элементы, саркомеры, состоят из двух типов параллельных


В количественном отношении наиболее важным белком миофибрилл является миозин (~65% мышечного
В количественном отношении наиболее важным белком миофибрилл является миозин (~65% мышечного
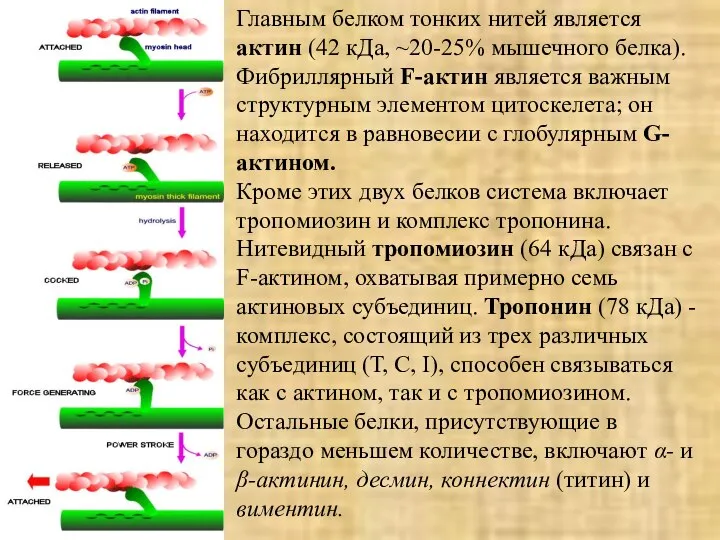
Главным белком тонких нитей является актин (42 кДа, ~20-25% мышечного белка).
Главным белком тонких нитей является актин (42 кДа, ~20-25% мышечного белка).

Механизм сокращения мышечных волокон
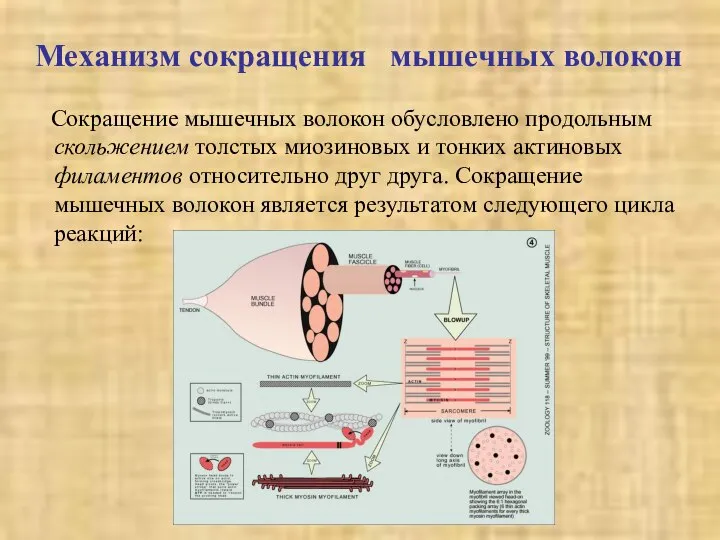
Сокращение мышечных волокон обусловлено продольным скольжением толстых миозиновых
Механизм сокращения мышечных волокон
Сокращение мышечных волокон обусловлено продольным скольжением толстых миозиновых

В отсутствие АТФ (АТР), т. е. в исходном состоянии, головки молекул
В отсутствие АТФ (АТР), т. е. в исходном состоянии, головки молекул
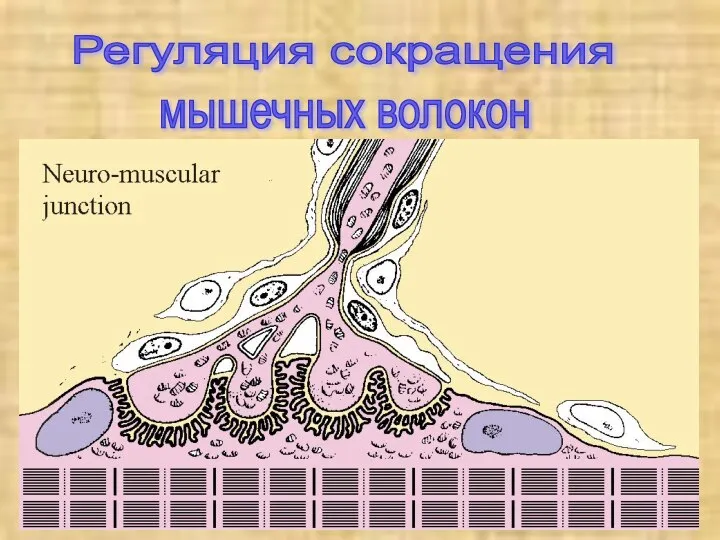
Регуляция сокращения
мышечных волокон
Регуляция сокращения
мышечных волокон


Электромеханическое сопряжение
Сокращением мышечного волокна управляют двигательные нейроны, которые выделяют
Электромеханическое сопряжение
Сокращением мышечного волокна управляют двигательные нейроны, которые выделяют

![Саркоплазматический ретикулум Саркоплазматический ретикулум [СР (SR)] – разветвленная подобная эндоплазматическому ретикулуму](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1299556/slide-14.jpg)
Саркоплазматический ретикулум
Саркоплазматический ретикулум [СР (SR)] – разветвленная подобная эндоплазматическому ретикулуму
Саркоплазматический ретикулум
Саркоплазматический ретикулум [СР (SR)] – разветвленная подобная эндоплазматическому ретикулуму
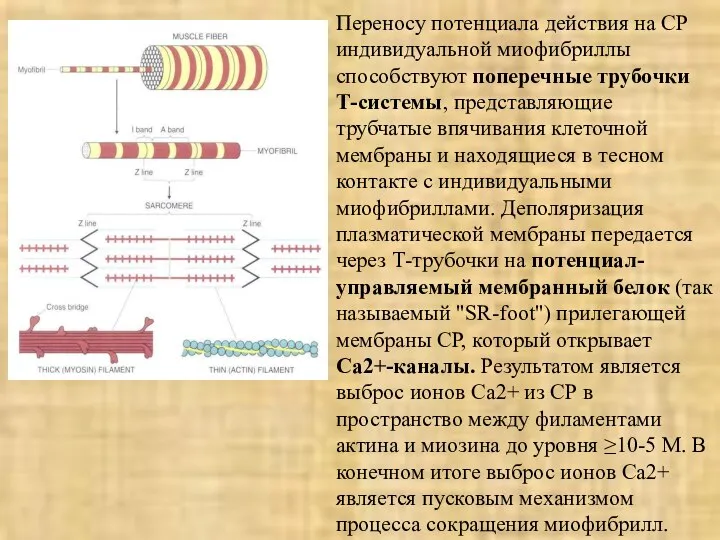
Переносу потенциала действия на СР индивидуальной миофибриллы способствуют поперечные трубочки Т-системы,
Переносу потенциала действия на СР индивидуальной миофибриллы способствуют поперечные трубочки Т-системы,


В расслабленной скелетной мышце комплекс
тропонина (субъединицы = Т, С, I)
тропонина (субъединицы = Т, С, I)
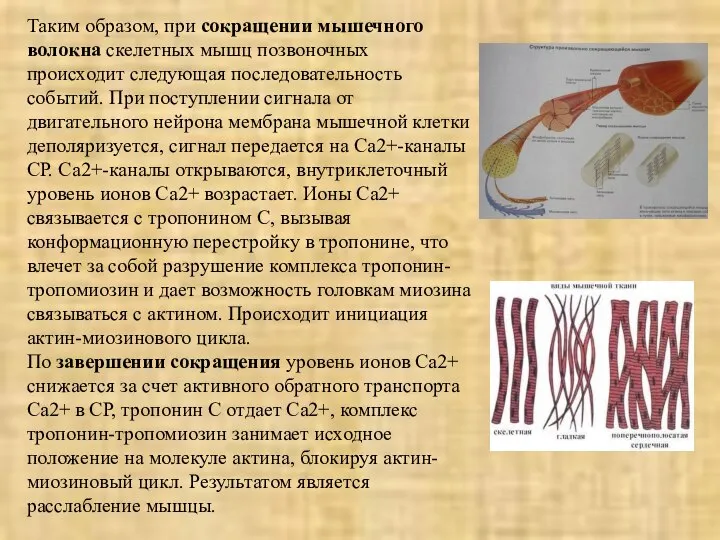
Таким образом, при сокращении мышечного волокна скелетных мышц позвоночных происходит следующая
Таким образом, при сокращении мышечного волокна скелетных мышц позвоночных происходит следующая
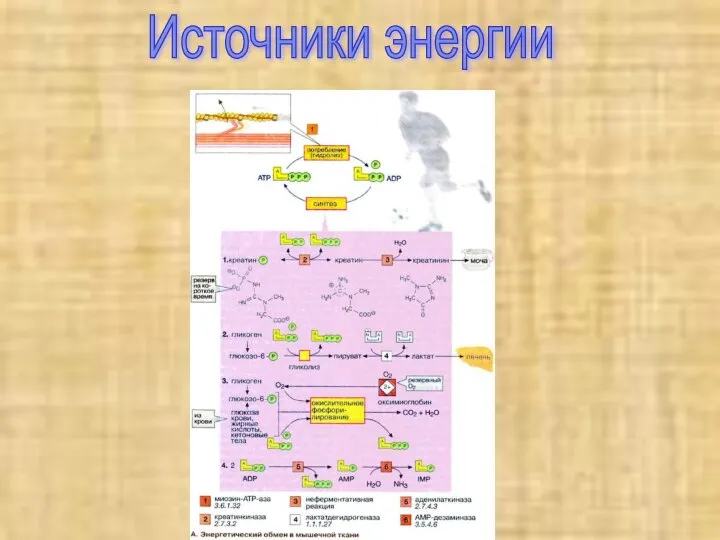
Источники энергии
Источники энергии
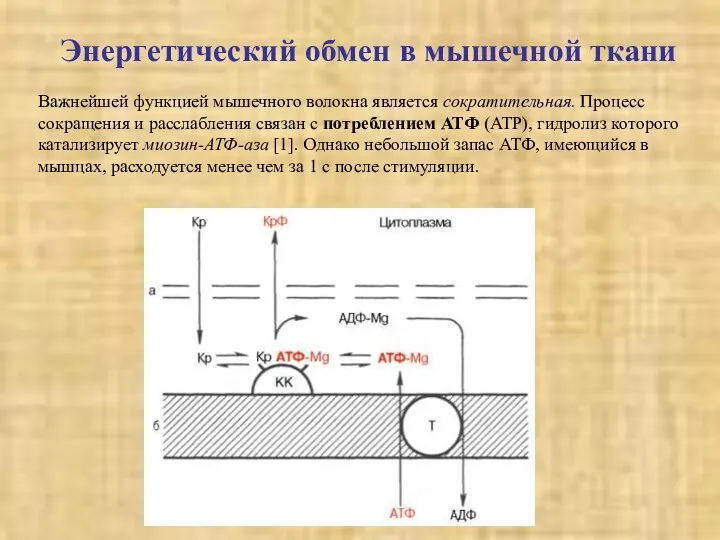
Энергетический обмен в мышечной ткани
Важнейшей функцией мышечного волокна является сократительная. Процесс
Энергетический обмен в мышечной ткани
Важнейшей функцией мышечного волокна является сократительная. Процесс

Потребности работающей мышцы в АТФ удовлетворяются за счет следующих ферментативных реакций:
1.
Потребности работающей мышцы в АТФ удовлетворяются за счет следующих ферментативных реакций:
1.

2 Анаэробный гликолиз. В мышечной ткани наиболее важным долгосрочным энергетическим резервом
2 Анаэробный гликолиз. В мышечной ткани наиболее важным долгосрочным энергетическим резервом
![4. Образование инозинмонофосфата [ИМФ (IMP)]. Другим источником быстрого восстановления уровня АТФ](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1299556/slide-23.jpg)
4. Образование инозинмонофосфата [ИМФ (IMP)]. Другим источником быстрого восстановления уровня АТФ
4. Образование инозинмонофосфата [ИМФ (IMP)]. Другим источником быстрого восстановления уровня АТФ

Метаболическая регуляция
мышечного сокращения
Метаболическая регуляция
мышечного сокращения


В клетках, не содержащих митохондрий (например, в эритроцитах), или в тканях
В клетках, не содержащих митохондрий (например, в эритроцитах), или в тканях

Обратная связь, подобная циклу Кори, существует в цикле аланина, в котором
Обратная связь, подобная циклу Кори, существует в цикле аланина, в котором


Метаболизм белков и аминокислот
Скелетные мышцы активно участвуют в метаболизме аминокислот. Это
Метаболизм белков и аминокислот
Скелетные мышцы активно участвуют в метаболизме аминокислот. Это
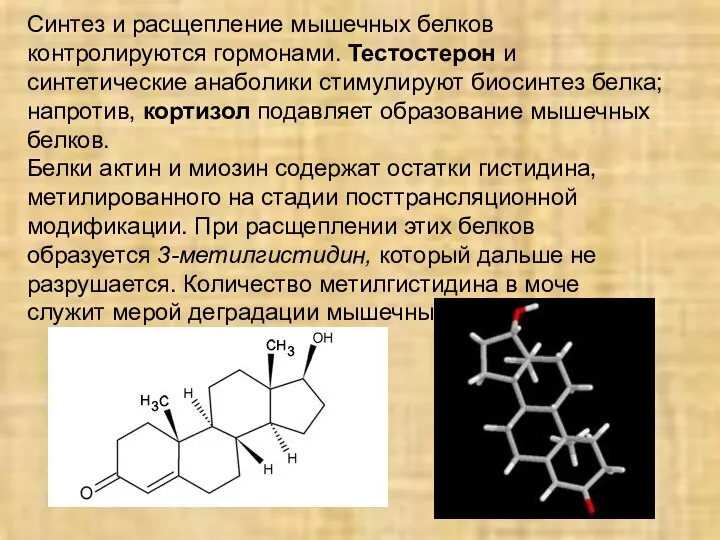
Синтез и расщепление мышечных белков контролируются гормонами. Тестостерон и синтетические анаболики
Синтез и расщепление мышечных белков контролируются гормонами. Тестостерон и синтетические анаболики
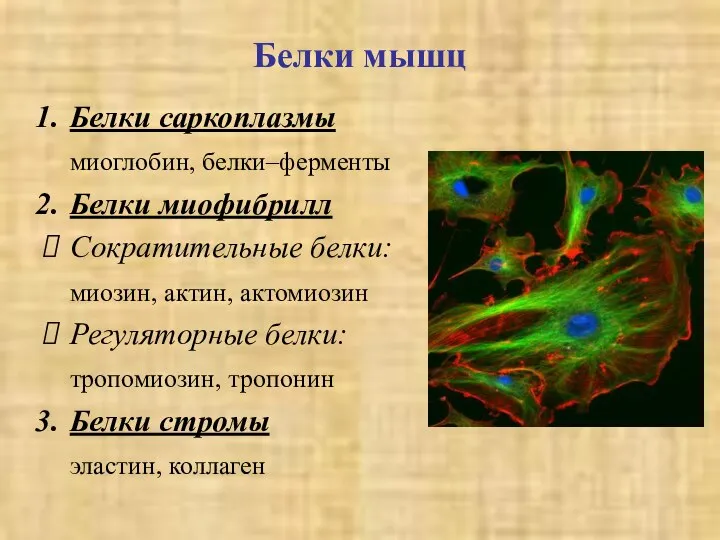
Белки мышц
Белки саркоплазмы
миоглобин, белки–ферменты
Белки миофибрилл
Сократительные белки:
миозин, актин, актомиозин
Регуляторные белки:
тропомиозин, тропонин
Белки стромы
эластин,
Белки мышц
Белки саркоплазмы
миоглобин, белки–ферменты
Белки миофибрилл
Сократительные белки:
миозин, актин, актомиозин
Регуляторные белки:
тропомиозин, тропонин
Белки стромы
эластин,

Миоглобин
Дыхательный белок мышц
Гемопротеин
Содержит 153 аминокислотных остатка
Основная функция – перенос кислорода в
Миоглобин
Дыхательный белок мышц
Гемопротеин
Содержит 153 аминокислотных остатка
Основная функция – перенос кислорода в

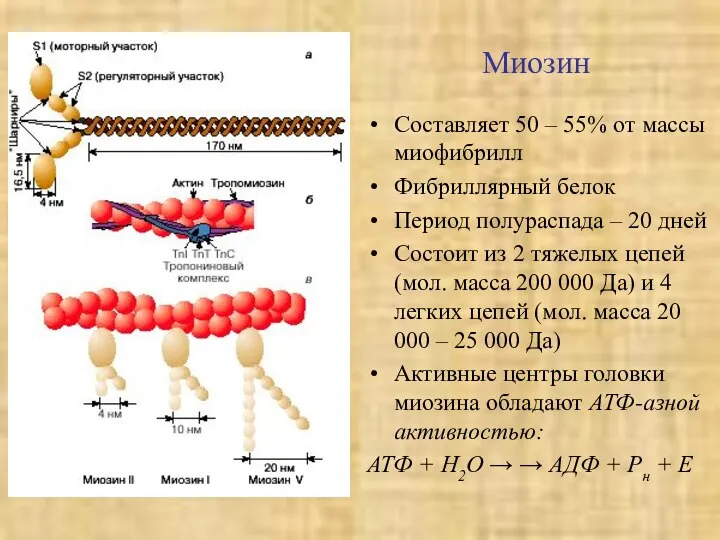
Миозин
Составляет 50 – 55% от массы миофибрилл
Фибриллярный белок
Период полураспада – 20
Миозин
Составляет 50 – 55% от массы миофибрилл
Фибриллярный белок
Период полураспада – 20
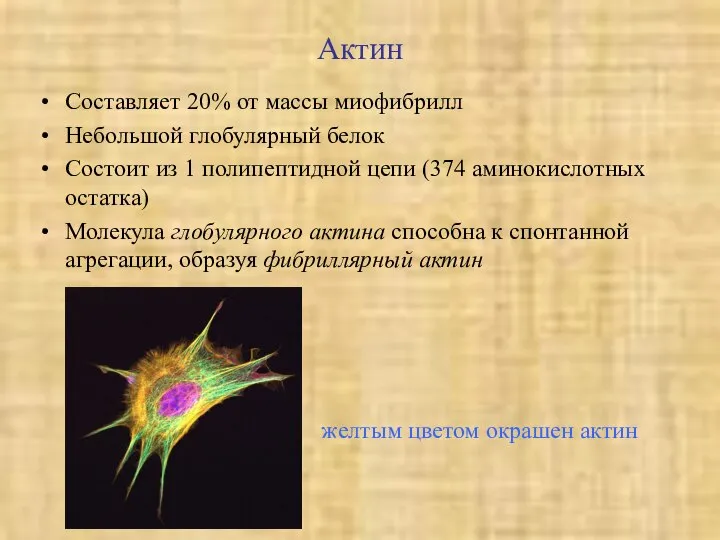
Актин
Составляет 20% от массы миофибрилл
Небольшой глобулярный белок
Состоит из 1 полипептидной цепи
Актин
Составляет 20% от массы миофибрилл
Небольшой глобулярный белок
Состоит из 1 полипептидной цепи

актиновая нить
актиновая нить
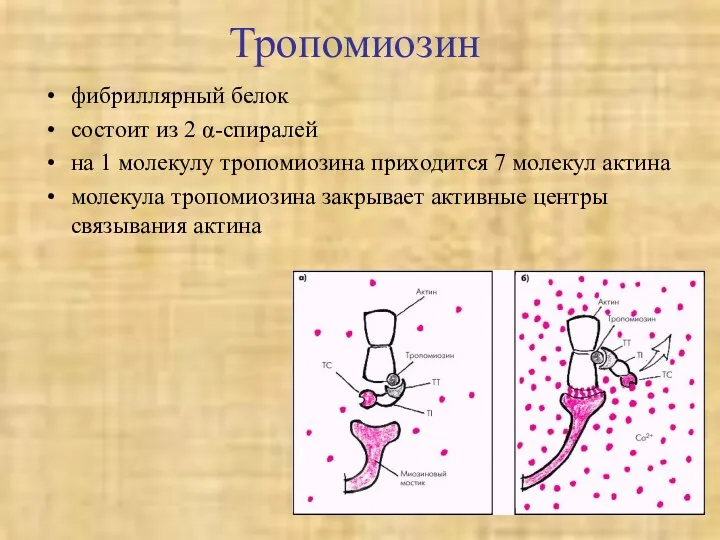
Тропомиозин
фибриллярный белок
состоит из 2 α-спиралей
на 1 молекулу тропомиозина приходится 7 молекул
Тропомиозин
фибриллярный белок
состоит из 2 α-спиралей
на 1 молекулу тропомиозина приходится 7 молекул
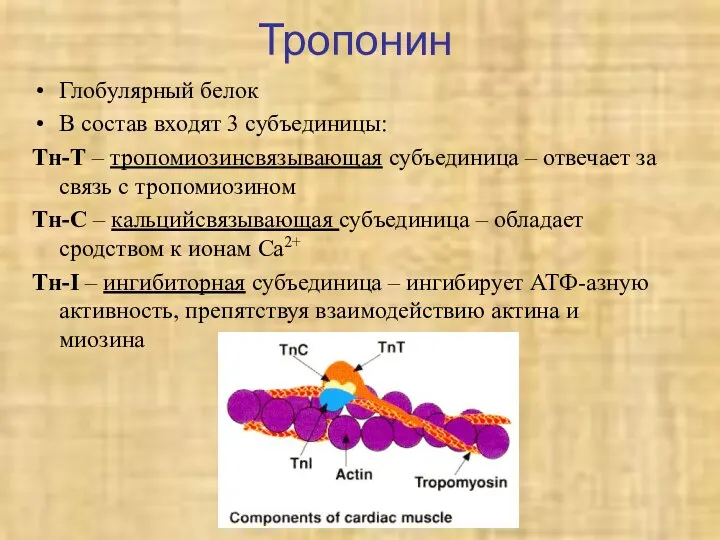
Тропонин
Глобулярный белок
В состав входят 3 субъединицы:
Тн-Т – тропомиозинсвязывающая субъединица – отвечает
Тропонин
Глобулярный белок
В состав входят 3 субъединицы:
Тн-Т – тропомиозинсвязывающая субъединица – отвечает
В мышцах присутствует фетальная форма миозина, обладающая измененной структурой, сниженной АТФ-азной
В мышцах присутствует фетальная форма миозина, обладающая измененной структурой, сниженной АТФ-азной

Биосинтез и распад
креатина
Креатин и креатининфосфат – важные азотистые вещества мышцы.
Находится
Биосинтез и распад
креатина
Креатин и креатининфосфат – важные азотистые вещества мышцы.
Находится

Синтез креатина
Первая стадия синтеза креатина протекает в почках под действием глицин-амидинотрансферазы.
Арг
Гли
Синтез креатина
Первая стадия синтеза креатина протекает в почках под действием глицин-амидинотрансферазы.
Арг
Гли
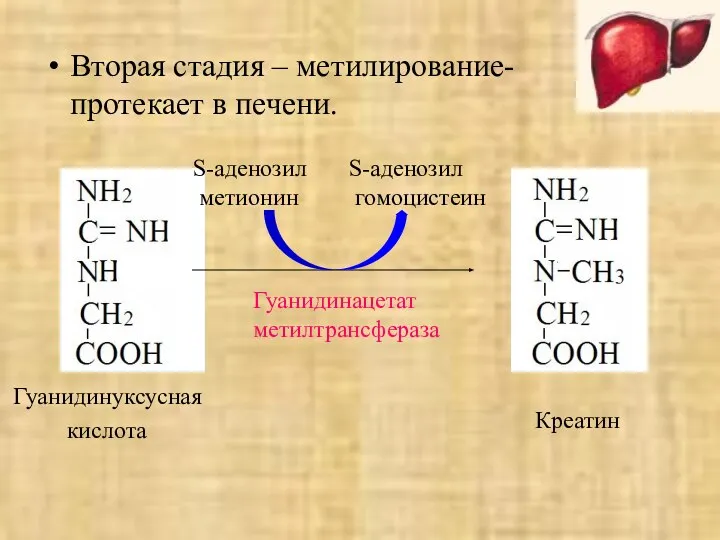
Вторая стадия – метилирование- протекает в печени.
Гуанидинуксусная
кислота
S-аденозил
Вторая стадия – метилирование- протекает в печени.
Гуанидинуксусная
кислота
S-аденозил
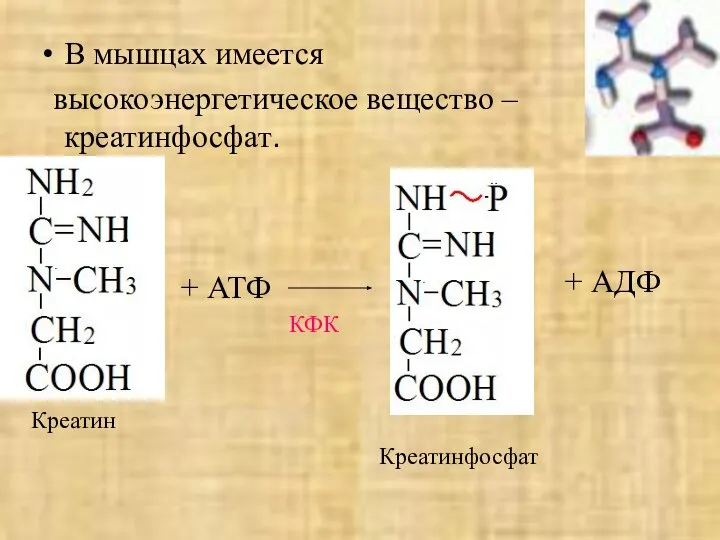
В мышцах имеется
высокоэнергетическое вещество – креатинфосфат.
Креатин
КФК
+ АТФ
+ АДФ
Креатинфосфат
В мышцах имеется
высокоэнергетическое вещество – креатинфосфат.
Креатин
КФК
+ АТФ
+ АДФ
Креатинфосфат
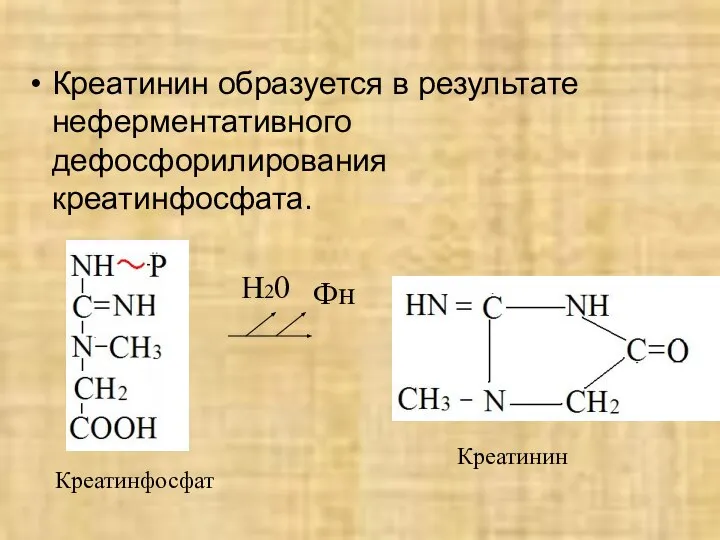
Креатинин образуется в результате неферментативного дефосфорилирования креатинфосфата.
Фн
Н20
Креатинфосфат
Креатинин
Креатинин образуется в результате неферментативного дефосфорилирования креатинфосфата.
Фн
Н20
Креатинфосфат
Креатинин

Содержание в плазме крови
В плазме крови в небольших количествах содержатся креатин
Содержание в плазме крови
В плазме крови в небольших количествах содержатся креатин

Диагностическое значение
уровень креатинина в сыворотке – чувствительный показатель состояния функции
Диагностическое значение
уровень креатинина в сыворотке – чувствительный показатель состояния функции

Физиологическая креатинурия наблюдается
у новорожденных, так как преобладает скорость синтеза
Физиологическая креатинурия наблюдается
у новорожденных, так как преобладает скорость синтеза
 Мікропроцесорний пристрій автоматизованого контролю параметрів в артезіанській свердловині
Мікропроцесорний пристрій автоматизованого контролю параметрів в артезіанській свердловині ß-адреноблокаторы – величайший прорыв в лечении сердечно-сосудистых заболеваний
ß-адреноблокаторы – величайший прорыв в лечении сердечно-сосудистых заболеваний Кинематика
Кинематика уральский рекреационный район
уральский рекреационный район Машинно-ориентированные языки программирования
Машинно-ориентированные языки программирования Презентация Запасы марганцевых и хромовых руд
Презентация Запасы марганцевых и хромовых руд  Классификация опасностей
Классификация опасностей 4 занятие.ppt
4 занятие.ppt La cigale et la fourmi
La cigale et la fourmi Презентация на тему "Эстафета олимпийского огня в Гатчине" - скачать презентации по Педагогике
Презентация на тему "Эстафета олимпийского огня в Гатчине" - скачать презентации по Педагогике Народные зимние праздники (интеллектуальная игра для 5-6 классов)
Народные зимние праздники (интеллектуальная игра для 5-6 классов) Женские пропорции
Женские пропорции Хоккейная площадка ВДНХ
Хоккейная площадка ВДНХ Судебная практика. Качели. Возмещение вреда
Судебная практика. Качели. Возмещение вреда Цветные революции
Цветные революции Проектирование тепловой защиты зданий
Проектирование тепловой защиты зданий Брак коренных народов севера
Брак коренных народов севера Домашние опасности - презентация для начальной школы_
Домашние опасности - презентация для начальной школы_ Таможенные органы РФ
Таможенные органы РФ  Индивидуальный проект. Калькулятор обоев
Индивидуальный проект. Калькулятор обоев Презентация "Гжель" - скачать презентации по МХК
Презентация "Гжель" - скачать презентации по МХК Комбинаторные задачи. №1. Из цифр 1, 5, 9 составить все трёхзначные числа без повторяющихся цифр. №2. ИЗ цифр 2, 4, 7 составить трёхзнач
Комбинаторные задачи. №1. Из цифр 1, 5, 9 составить все трёхзначные числа без повторяющихся цифр. №2. ИЗ цифр 2, 4, 7 составить трёхзнач The political system of Great Britain
The political system of Great Britain Электрические измерения и приборы
Электрические измерения и приборы Политика и финансы
Политика и финансы мышцы шеи1 лекц 8
мышцы шеи1 лекц 8 ИНФЛЯЦИЯ Экономика 11 класс
ИНФЛЯЦИЯ Экономика 11 класс  Механика грунтов
Механика грунтов