- БИОХИМИЯ МЫШЕЧНОЙ ТКАНИ

Содержание
- 2. Мышечная ткань –поперечно-полосатая (скелетная и сердечная) и гладкая. Обладает возбудимостью и сократимостью. Поперечно-полосатая состоит из огромных
- 3. Сердечная мышца – содержит 3 типа клеток: 1.типичные (рабочие) миокардиоциты (одноядерные, с миофибриллами, как в скелетных
- 5. Гладкая мышечная ткань – образована одноядерными клетками (много митохондрий, ЭПС развит слабо), миозиновые филламенты вне сокращения
- 6. Саркомер – регулярно повторяющаяся структура (1,8 мкм), до 1000-1200 по длине миофибриллы. Состоит из упорядоченно расположенных
- 7. Мышечные белки Миофибриллярные (миозин, актин, тропомиозин, тропонин). Растворяются в растворах с высокой ионной силой. Больше всего
- 8. Фибриллярные белки: Миозин образует темные диски (А) саркомеров, М.м 470 000 Да. 2 полипептидные (тяжелые) цепи
- 13. Этапы мышечного сокращения и расслабления 1.Гидролиз АТФ миозиновой головкой 2. Связывание комплекса миозин+АДФ+Ф с актином 3.
- 14. Особенности сокращения сердечной мышцы Источником Ca2+является внеклеточная жидкость. Поэтому, при снижении концентрации кальция во внеклеточной жидкости
- 15. Особенность сокращения гладкой мышцы Двигательный импульс изменяет проницаемость мембраны не для Na+, а для Са2+,который поступает
- 16. Роль Са2+в метаболизме мышечной ткани. Связывание Са2+с тропонином С изменяет конформацию тропонин-тропомиозинового комплекса, тем самым освобождает
- 17. АТФ – зависимые процессы при мышечном сокращении 1. Работа Na +- насоса клеточной мембраны по перемещению
- 18. Метаболические процессы, поддерживающие необходимую концентрацию АТФ при мышечной работе Креатинфосфокиназная реакция Гликогенолиз, гликолиз Аэробное окисление глюкозы,
- 20. При интенсивной мышечной работе запас АТФ истощается за 1 секунду! 1.Еще несколько секунд запас АТФ поддерживает
- 21. В мышцах функционируют разные моторные единицы в составе которых разные мышечные волокна, отличающиеся формами миозина, АТФ-азной
- 25. Особенности углеводного обмена Поступление глюкозы усиливается при мышечной работе или под влиянием инсулина. В покое –
- 26. Углеводный обмен Активность ключевых ферментов гликолиза в 20 раз выше, чем в печени. Гликолиз практически необратим,
- 27. Углеводный обмен После приема углеводной пищи, в покое создается запас гликогена (1-2% от массы ткани). 15
- 28. Липидный обмен Преобладают реакции катаболизма. Жировые депо могут формироваться в прилежащих к мышцах тканях. В миоцитах
- 29. L-КАРНИТИН(N-гуанидин-N-метилглицин) Содержание карнитина в мышцах много >>, чем в печени или др. органах. Синтезируется в печени
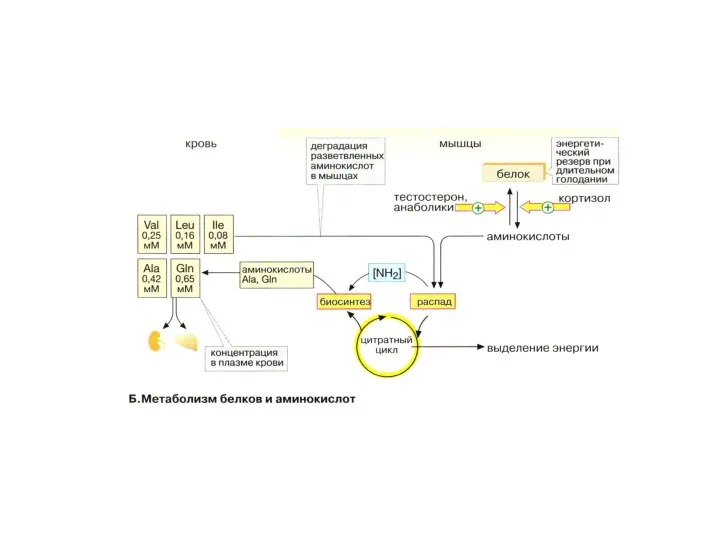
- 30. Аминокислотный и белковый обмен Тестостерон активирует синтез белков, кортизол – подавляет. При голодании организм использует мышечные
- 31. аминокислоты Скелетные мышцы – основной компартмент деградации аминокислот, особенно с разветвленной цепью. В кровь активно поступают
- 32. Гормонально-метаболическая ситуация в организме при физической нагрузке Скелетные мышцы, имея большую массу (до 50% веса тела)
- 33. Литература: Трудные вопросы биохимии. Избранные лекции: учебное пособие/ под редакцией Т.С.Федоровой, В.Ю.Сереброва. – Томск: СибГМУ, 2006.-318с.
- 35. Скачать презентацию

Мышечная ткань –поперечно-полосатая (скелетная и сердечная) и гладкая.
Обладает возбудимостью и
Мышечная ткань –поперечно-полосатая (скелетная и сердечная) и гладкая. Обладает возбудимостью и

Сердечная мышца – содержит 3 типа клеток:
1.типичные (рабочие) миокардиоциты
(одноядерные, с
Сердечная мышца – содержит 3 типа клеток: 1.типичные (рабочие) миокардиоциты (одноядерные, с

Гладкая мышечная ткань – образована одноядерными клетками (много митохондрий, ЭПС развит
Гладкая мышечная ткань – образована одноядерными клетками (много митохондрий, ЭПС развит

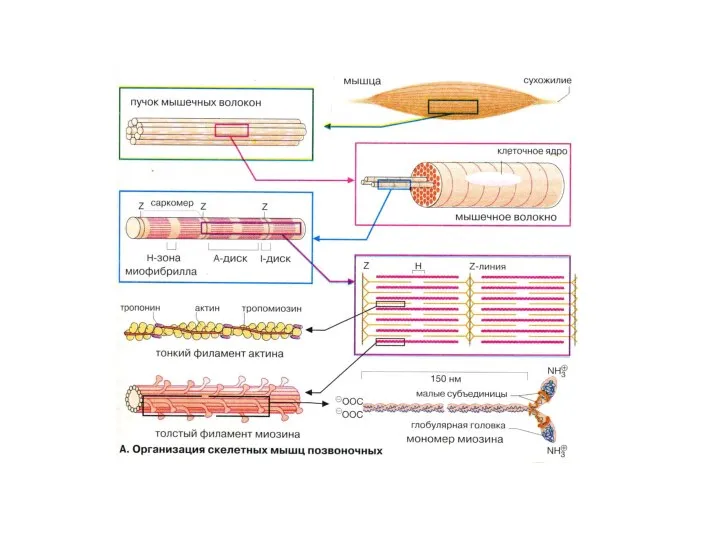
Саркомер – регулярно повторяющаяся структура (1,8 мкм), до 1000-1200 по длине
Саркомер – регулярно повторяющаяся структура (1,8 мкм), до 1000-1200 по длине

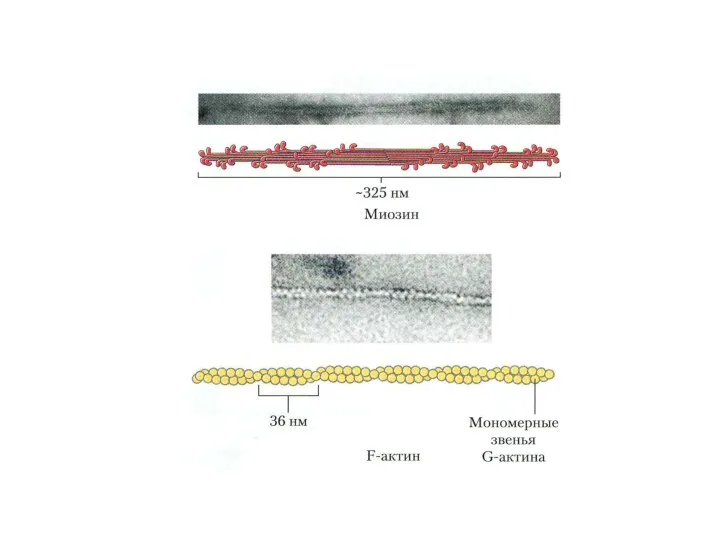
Мышечные белки
Миофибриллярные (миозин, актин, тропомиозин, тропонин). Растворяются в растворах с высокой
Мышечные белки
Миофибриллярные (миозин, актин, тропомиозин, тропонин). Растворяются в растворах с высокой

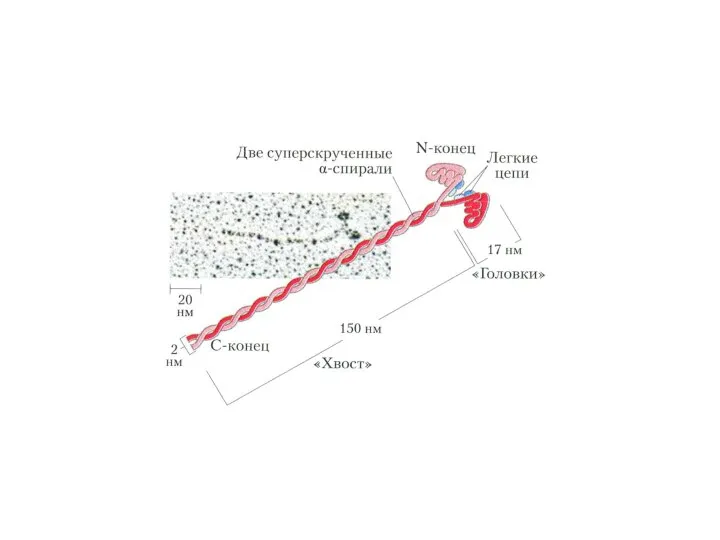
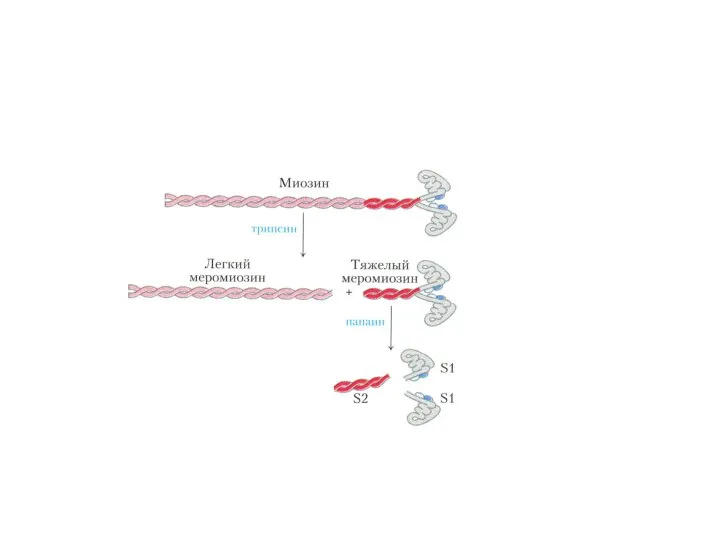
Фибриллярные белки:
Миозин образует темные диски (А) саркомеров, М.м 470 000 Да.
Фибриллярные белки: Миозин образует темные диски (А) саркомеров, М.м 470 000 Да.
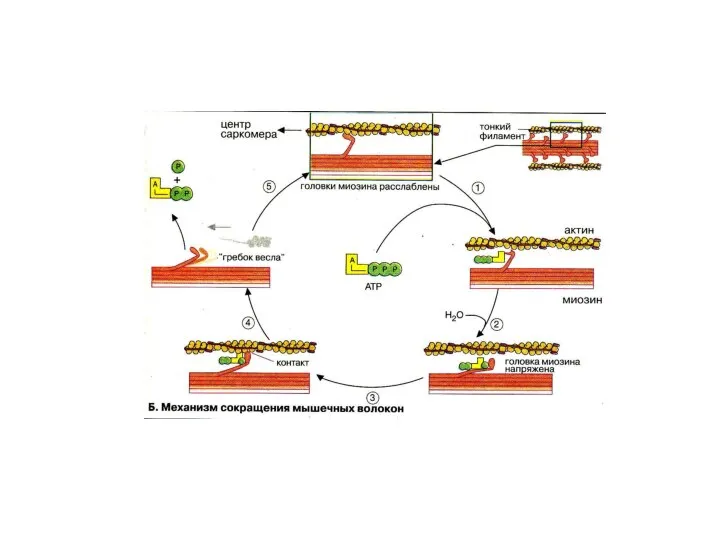

Этапы мышечного сокращения и расслабления
1.Гидролиз АТФ миозиновой головкой
2. Связывание комплекса миозин+АДФ+Ф
Этапы мышечного сокращения и расслабления
1.Гидролиз АТФ миозиновой головкой
2. Связывание комплекса миозин+АДФ+Ф

Особенности сокращения сердечной мышцы
Источником Ca2+является внеклеточная
жидкость. Поэтому, при снижении
концентрации кальция
Особенности сокращения сердечной мышцы
Источником Ca2+является внеклеточная
жидкость. Поэтому, при снижении
концентрации кальция

Особенность сокращения гладкой мышцы
Двигательный импульс изменяет проницаемость мембраны не для Na+,
Особенность сокращения гладкой мышцы
Двигательный импульс изменяет проницаемость мембраны не для Na+,

Роль Са2+в метаболизме мышечной ткани.
Связывание Са2+с тропонином С изменяет конформацию тропонин-тропомиозинового
Роль Са2+в метаболизме мышечной ткани.
Связывание Са2+с тропонином С изменяет конформацию тропонин-тропомиозинового

АТФ – зависимые процессы при мышечном сокращении
1. Работа Na +- насоса
АТФ – зависимые процессы при мышечном сокращении
1. Работа Na +- насоса

Метаболические процессы, поддерживающие необходимую концентрацию АТФ при мышечной работе
Креатинфосфокиназная реакция
Гликогенолиз,
Метаболические процессы, поддерживающие необходимую концентрацию АТФ при мышечной работе
Креатинфосфокиназная реакция
Гликогенолиз,
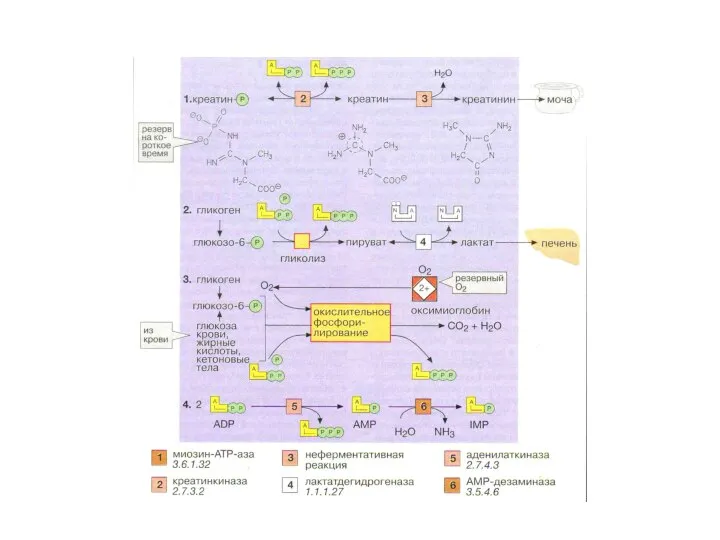

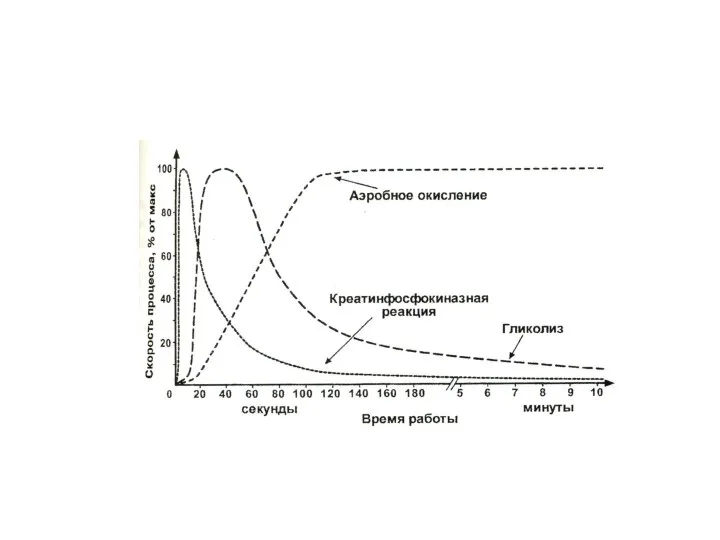
При интенсивной мышечной работе запас АТФ истощается за 1 секунду!
1.Еще несколько
При интенсивной мышечной работе запас АТФ истощается за 1 секунду!
1.Еще несколько

В мышцах функционируют разные моторные единицы в составе которых разные мышечные
В мышцах функционируют разные моторные единицы в составе которых разные мышечные
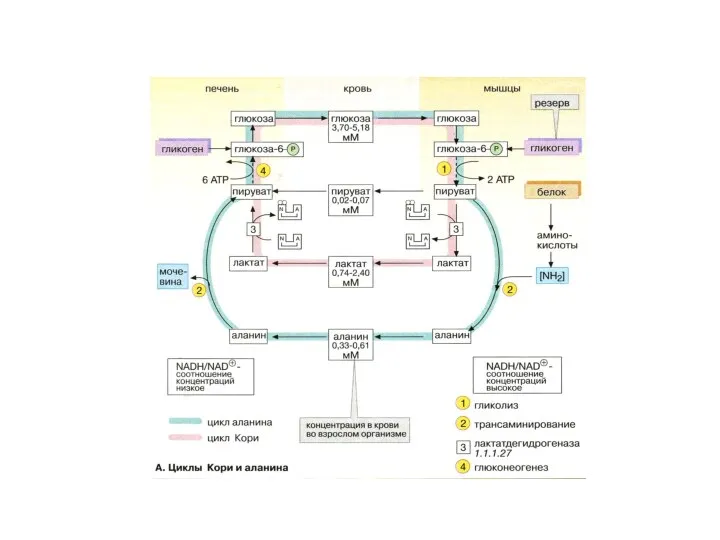

Особенности углеводного обмена
Поступление глюкозы усиливается при мышечной работе или под
Особенности углеводного обмена
Поступление глюкозы усиливается при мышечной работе или под

Углеводный обмен
Активность ключевых ферментов гликолиза в 20 раз выше, чем в
Углеводный обмен
Активность ключевых ферментов гликолиза в 20 раз выше, чем в

Углеводный обмен
После приема углеводной пищи, в покое создается запас гликогена (1-2%
Углеводный обмен
После приема углеводной пищи, в покое создается запас гликогена (1-2%

Липидный обмен
Преобладают реакции катаболизма.
Жировые депо могут формироваться в прилежащих к мышцах
Липидный обмен
Преобладают реакции катаболизма.
Жировые депо могут формироваться в прилежащих к мышцах

L-КАРНИТИН(N-гуанидин-N-метилглицин)
Содержание карнитина в мышцах много >>, чем в печени или др.
L-КАРНИТИН(N-гуанидин-N-метилглицин)
Содержание карнитина в мышцах много >>, чем в печени или др.

Аминокислотный и белковый обмен
Тестостерон активирует синтез белков, кортизол – подавляет.
При голодании
Аминокислотный и белковый обмен
Тестостерон активирует синтез белков, кортизол – подавляет.
При голодании

аминокислоты
Скелетные мышцы – основной компартмент деградации аминокислот, особенно с разветвленной цепью.
аминокислоты
Скелетные мышцы – основной компартмент деградации аминокислот, особенно с разветвленной цепью.

Гормонально-метаболическая ситуация в организме при физической нагрузке
Скелетные мышцы, имея большую массу
Гормонально-метаболическая ситуация в организме при физической нагрузке
Скелетные мышцы, имея большую массу

Литература:
Трудные вопросы биохимии. Избранные лекции: учебное пособие/ под редакцией Т.С.Федоровой, В.Ю.Сереброва.
Литература:
Трудные вопросы биохимии. Избранные лекции: учебное пособие/ под редакцией Т.С.Федоровой, В.Ю.Сереброва.
 Христианское образование
Христианское образование Освоение культурного наследия Многоканальная модель освоения культурного наследия
Освоение культурного наследия Многоканальная модель освоения культурного наследия Введение в XML
Введение в XML Режим дня младшего школьника
Режим дня младшего школьника  Whatsapp-лендинг. Очередь клиентов в любой бизнес без сайта и с конверсией до 80%
Whatsapp-лендинг. Очередь клиентов в любой бизнес без сайта и с конверсией до 80% Вкусовой и обонятельный анализатор
Вкусовой и обонятельный анализатор профессиональный стандарт педагога (ПСП)
профессиональный стандарт педагога (ПСП) Каменная кладка. Четыре стадии работы кладки под нагрузкой при сжатии
Каменная кладка. Четыре стадии работы кладки под нагрузкой при сжатии Техническая информация Thermo Top C, Thermo Top V
Техническая информация Thermo Top C, Thermo Top V Народные гуляния как традиция, исторически сложившаяся в советскую эпоху
Народные гуляния как традиция, исторически сложившаяся в советскую эпоху Презентация_modif
Презентация_modif  Құқықтық сана және құқықтық тәртіп
Құқықтық сана және құқықтық тәртіп Татарские народные праздники и обычаи
Татарские народные праздники и обычаи МЕТОДИЧЕСКАЯ ТЕМА: «АКТИВИЗАЦИЯ ПОЗНАВАТЕЛЬНОЙ ДЕЯТЕЛЬНОСТИ УЧАЩИХСЯ НА УРОКАХ ПУТЕМ ИСПОЛЬЗОВАНИЯ НОВЫХ ПЕДАГОГИЧЕСКИХ ТЕХН
МЕТОДИЧЕСКАЯ ТЕМА: «АКТИВИЗАЦИЯ ПОЗНАВАТЕЛЬНОЙ ДЕЯТЕЛЬНОСТИ УЧАЩИХСЯ НА УРОКАХ ПУТЕМ ИСПОЛЬЗОВАНИЯ НОВЫХ ПЕДАГОГИЧЕСКИХ ТЕХН План строительства пятиэтажного здания
План строительства пятиэтажного здания Студенческие арт-клубы УГНТУ
Студенческие арт-клубы УГНТУ Адаптация учащихся в предметной учебной деятельности формированием понятийного аппарата, или Как овладеть языком предмета - през
Адаптация учащихся в предметной учебной деятельности формированием понятийного аппарата, или Как овладеть языком предмета - през Общая характеристика Зарубежной Европы
Общая характеристика Зарубежной Европы Карловы Вары
Карловы Вары Формула Тейлора
Формула Тейлора  Метод комплексных амплитуд
Метод комплексных амплитуд Социальные проблемы и безработица Выполнили ученицы 11Б класса СОШ№2: Коптева Анастасия Грачева Наталья
Социальные проблемы и безработица Выполнили ученицы 11Б класса СОШ№2: Коптева Анастасия Грачева Наталья  Boże Narodzenie
Boże Narodzenie Выполнила учитель начальных классов МОУ – Сукроменская СОШ Банникова Наталья Олеговна
Выполнила учитель начальных классов МОУ – Сукроменская СОШ Банникова Наталья Олеговна Презентация "Путешествие от барокко к классицизму 4" - скачать презентации по МХК
Презентация "Путешествие от барокко к классицизму 4" - скачать презентации по МХК Современная Front-End разработка. Объектно-ориентированное программирование в JavaScript. (Лекция 4)
Современная Front-End разработка. Объектно-ориентированное программирование в JavaScript. (Лекция 4) Електроємність. Конденсатори. (Лекция 6)
Електроємність. Конденсатори. (Лекция 6) Инклюзивный ПрофГид. Содействие профориентации и трудоустройству лиц с ОВЗ и инвалидностью
Инклюзивный ПрофГид. Содействие профориентации и трудоустройству лиц с ОВЗ и инвалидностью