- Принципы интеграции метаболических путей

Содержание
- 2. Интегра́ция (от лат. integratio — «соедине-ние») объединение частей в целое. В биологии понятие интеграции ввёл английский
- 3. Мы будем рассматривать молекулярные механизмы интеграции обмена в рамках физи-ологических ответов организма: в условиях сравнительно слабых
- 4. Живые системы – открытые системы: обме-ниваются с окружающей их средой вещест-вом, энергией и информацией. Медико-биологическая наука
- 5. Биологической эволюции потребовалось около 1 млрд. лет от момента появления одно-клеточных организмов для создания первых многоклеточных
- 6. Метаболические пути состоят из последова-тельных реакций: продукт предыдущей ста-новится субстратом следующей реакции. Необратимость - общее свойство
- 7. Метаболизм = катаболические пути (рас-щепление сложных биомолекул на более простые - конечные продукты с выделе-нием свободной
- 8. Катаболические пути сходятся – образуя че-рез промежуточный продукт пируват, по сути, один конечный продукт: ацетил-СоА. Анаболические
- 10. Процессы анаболизма и катаболизма находятся в организме в состоянии динами-ческого равновесия или преобладания одного из них
- 11. В различных точках метаболических путей находятся ключевые метаболиты. Они обес-печивают переключение метаболизма с одно-го метаболического пути
- 12. 3. Ацетил-КоА. Включает углеводы, липиды и белки в первую реакцию ЦТК (цитрат лиаза) и является субстратом
- 13. Благодаря необратимости ПВК-ДГ и цитрат синтазы обратное превращение ацетил-КоА в пируват невозможно. То есть, в организме
- 14. Извлеченная в реакция катаболизма энергия, аккумулируется в трёх формах: 1. Макроэргические связи АТФ. АТФ посто-янно образуется
- 15. 2. Пиридиновые и флавиновые нуклеотиды: НАДН, ФАДН2 и НАДФН. Это переносчики атомов Н, которые в катаболических
- 16. б). НАДФН – главный донор богатых энергией атомов Н для биосинтетических путей. Молекулы субстратов и интермедиатов
- 17. Пять стратегий регуляции метаболизма I. Быстрое изменение каталитической актив-ности ключевых ферментов под влиянием аллостерических регуляторов. II.
- 18. IV. Компартментализация: - матрикс митохондрий: ЦТК, синтез кетоно-вых тел и β-окисление жирных кислот; - цитоплазма: гликолиз,
- 19. В чём и посредством чего проявляется интеграция обмена углеводов, липидов и белков? 1. Общие источники свободной
- 20. Ключевая роль печени в интеграции обмена углеводов, липидов и белков, а также в стабилизации метаболического гомеостаза
- 21. Эндокринная система в интеграции обмена углеводов, липидов и белков На уровне целостного организма интеграция углеводного, липидного
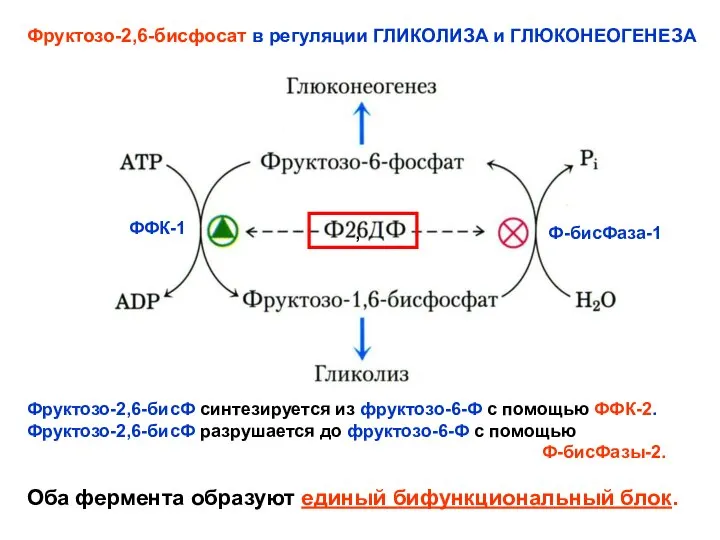
- 24. , Фруктозо-2,6-бисфосат в регуляции ГЛИКОЛИЗА и ГЛЮКОНЕОГЕНЕЗА ФФК-1 Ф-бисФаза-1 Фруктозо-2,6-бисФ синтезируется из фруктозо-6-Ф с помощью ФФК-2.
- 25. Роль гормонов в регуляции концентрации фруктозо-2,6-бифосфата
- 29. Скачать презентацию

Интегра́ция (от лат. integratio — «соедине-ние») объединение частей в целое.
В биологии понятие интеграции
Интегра́ция (от лат. integratio — «соедине-ние») объединение частей в целое.
В биологии понятие интеграции

Мы будем рассматривать молекулярные механизмы интеграции обмена в рамках физи-ологических
Мы будем рассматривать молекулярные механизмы интеграции обмена в рамках физи-ологических

Живые системы – открытые системы: обме-ниваются с окружающей их средой
Живые системы – открытые системы: обме-ниваются с окружающей их средой

Биологической эволюции потребовалось около 1 млрд. лет от момента появления
Биологической эволюции потребовалось около 1 млрд. лет от момента появления

Метаболические пути состоят из последова-тельных реакций: продукт предыдущей ста-новится субстратом
Метаболические пути состоят из последова-тельных реакций: продукт предыдущей ста-новится субстратом

Метаболизм = катаболические пути (рас-щепление сложных биомолекул на более простые -
Метаболизм = катаболические пути (рас-щепление сложных биомолекул на более простые -

Катаболические пути сходятся – образуя че-рез промежуточный продукт пируват, по
Катаболические пути сходятся – образуя че-рез промежуточный продукт пируват, по
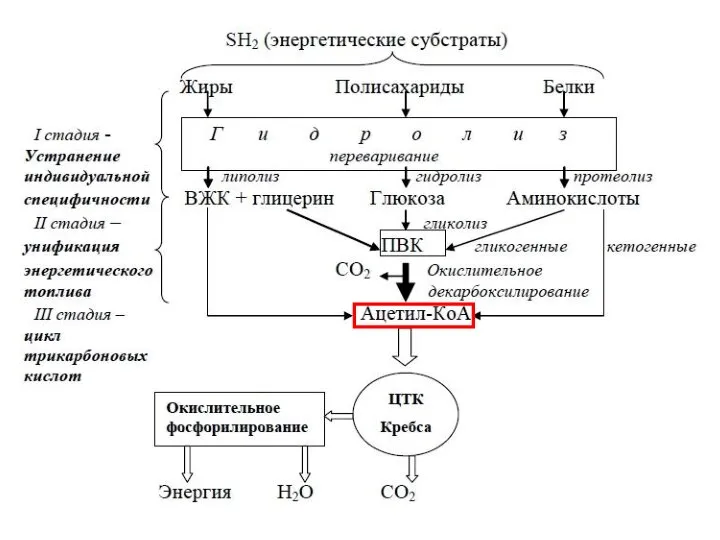

Процессы анаболизма и катаболизма находятся в организме в состоянии динами-ческого
Процессы анаболизма и катаболизма находятся в организме в состоянии динами-ческого

В различных точках метаболических путей находятся ключевые метаболиты. Они обес-печивают
В различных точках метаболических путей находятся ключевые метаболиты. Они обес-печивают

3. Ацетил-КоА. Включает углеводы, липиды и белки в первую реакцию ЦТК
3. Ацетил-КоА. Включает углеводы, липиды и белки в первую реакцию ЦТК

Благодаря необратимости ПВК-ДГ и цитрат синтазы обратное превращение ацетил-КоА в
Благодаря необратимости ПВК-ДГ и цитрат синтазы обратное превращение ацетил-КоА в

Извлеченная в реакция катаболизма энергия, аккумулируется в трёх формах:
1. Макроэргические
Извлеченная в реакция катаболизма энергия, аккумулируется в трёх формах:
1. Макроэргические

2. Пиридиновые и флавиновые нуклеотиды: НАДН, ФАДН2 и НАДФН. Это переносчики
2. Пиридиновые и флавиновые нуклеотиды: НАДН, ФАДН2 и НАДФН. Это переносчики

б). НАДФН – главный донор богатых энергией атомов Н для биосинтетических
б). НАДФН – главный донор богатых энергией атомов Н для биосинтетических

Пять стратегий регуляции метаболизма
I. Быстрое изменение каталитической актив-ности ключевых ферментов под
Пять стратегий регуляции метаболизма
I. Быстрое изменение каталитической актив-ности ключевых ферментов под

IV. Компартментализация:
- матрикс митохондрий: ЦТК, синтез кетоно-вых тел и β-окисление
IV. Компартментализация:
- матрикс митохондрий: ЦТК, синтез кетоно-вых тел и β-окисление

В чём и посредством чего проявляется интеграция обмена углеводов, липидов и
В чём и посредством чего проявляется интеграция обмена углеводов, липидов и

Ключевая роль печени в интеграции обмена углеводов, липидов и белков, а
Ключевая роль печени в интеграции обмена углеводов, липидов и белков, а

Эндокринная система в интеграции обмена углеводов, липидов и белков
На уровне целостного
Эндокринная система в интеграции обмена углеводов, липидов и белков
На уровне целостного
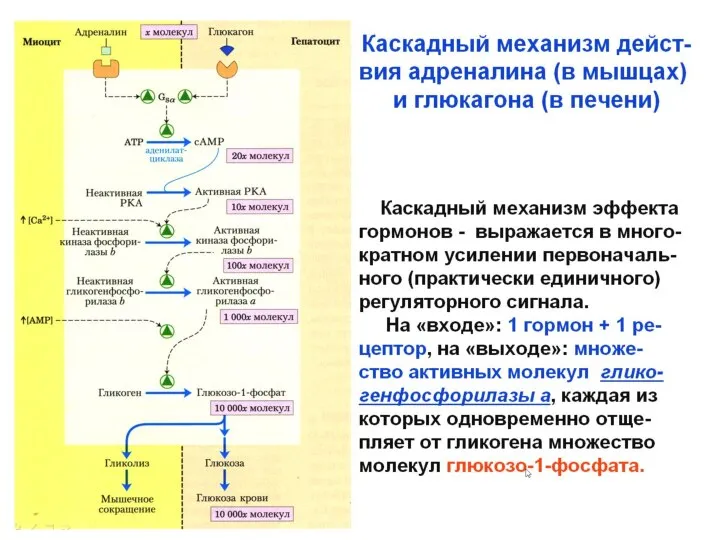
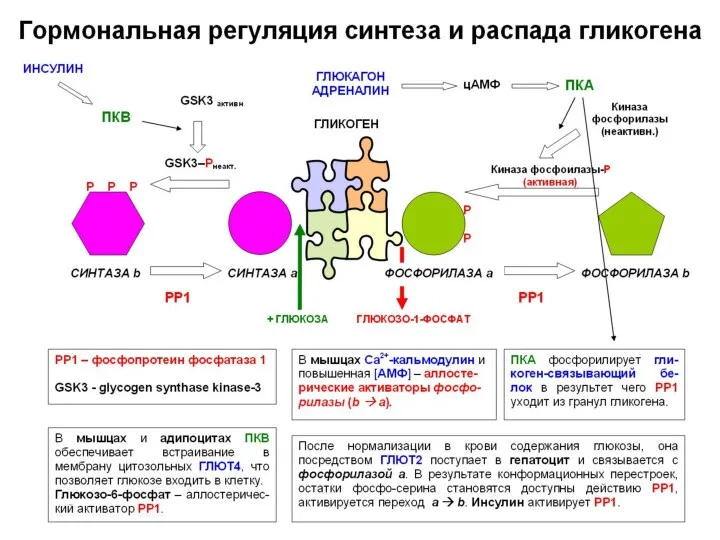
,
Фруктозо-2,6-бисфосат в регуляции ГЛИКОЛИЗА и ГЛЮКОНЕОГЕНЕЗА
ФФК-1
Ф-бисФаза-1
Фруктозо-2,6-бисФ синтезируется из фруктозо-6-Ф с помощью
,
Фруктозо-2,6-бисфосат в регуляции ГЛИКОЛИЗА и ГЛЮКОНЕОГЕНЕЗА
ФФК-1
Ф-бисФаза-1
Фруктозо-2,6-бисФ синтезируется из фруктозо-6-Ф с помощью
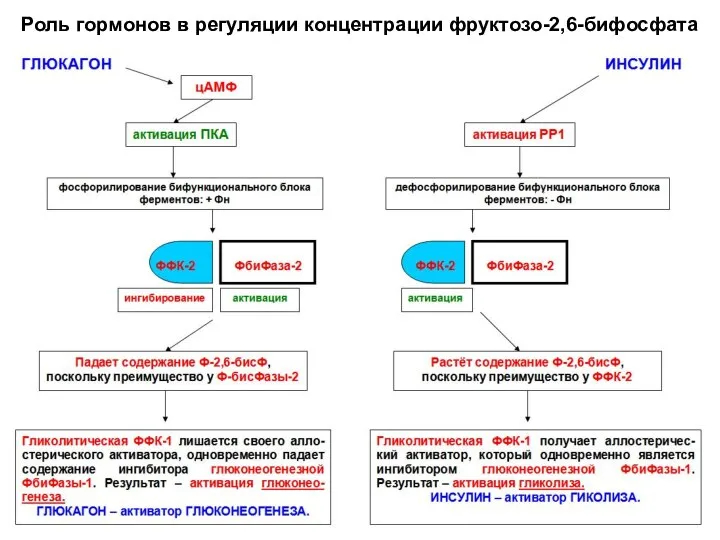
Роль гормонов в регуляции концентрации фруктозо-2,6-бифосфата
Роль гормонов в регуляции концентрации фруктозо-2,6-бифосфата
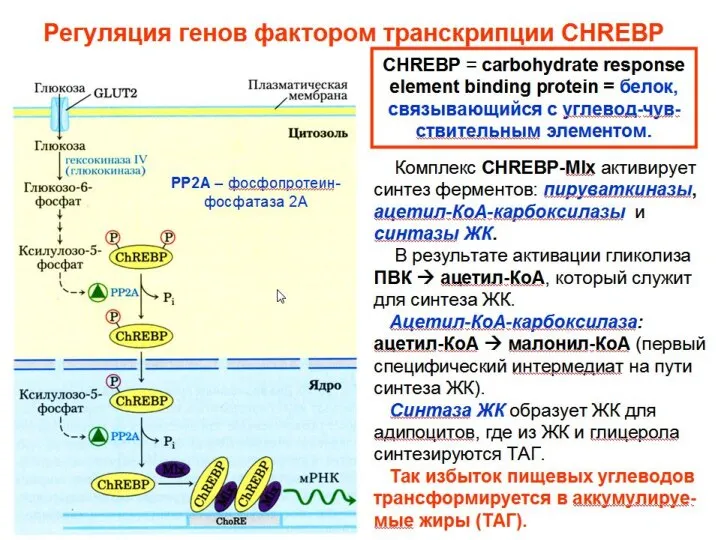
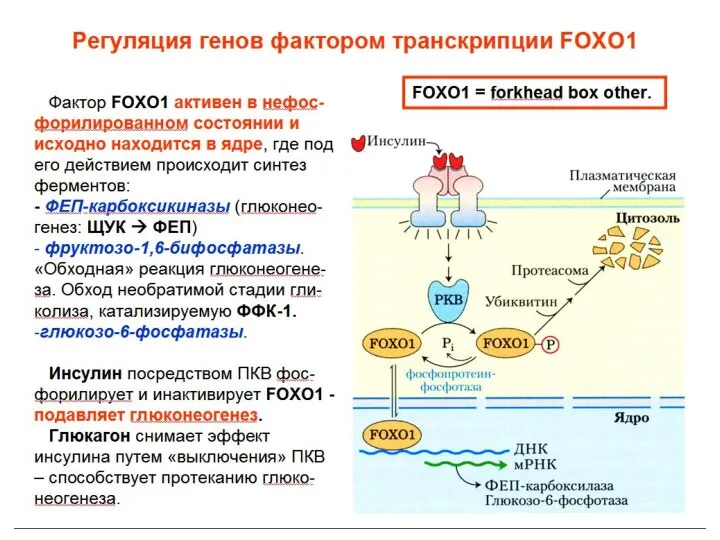
 Общеразвивающие упражнения
Общеразвивающие упражнения places-in-the-school-ppt-flashcards-fun-activities-games-picture-descriptio_53340
places-in-the-school-ppt-flashcards-fun-activities-games-picture-descriptio_53340 Модели деловой карьеры менеджера
Модели деловой карьеры менеджера Тема проповеди: Секрет 10 конвертов, или Как правильно планировать семейный бюджет
Тема проповеди: Секрет 10 конвертов, или Как правильно планировать семейный бюджет Строковые потоки и буферы
Строковые потоки и буферы Уран - самая голубая планета
Уран - самая голубая планета  Тренажёр по русскому языку 2 класс Учитель начальных классов МАОУ «СОШ №26 г. Сыктывкара» Республики Коми Иконникова Зоя Никол
Тренажёр по русскому языку 2 класс Учитель начальных классов МАОУ «СОШ №26 г. Сыктывкара» Республики Коми Иконникова Зоя Никол Конституционные правовые отношения
Конституционные правовые отношения Специальное снаряжение для водного туризма
Специальное снаряжение для водного туризма Проектирование электрооборудования станка токарно-винторезного МК6046М. Схемы
Проектирование электрооборудования станка токарно-винторезного МК6046М. Схемы Program M2 Internship at SPIN Center-PMMG
Program M2 Internship at SPIN Center-PMMG Курсовая работа по дисциплине «Основы управления персоналом» тема: «Совершенствование условий труда персонала»
Курсовая работа по дисциплине «Основы управления персоналом» тема: «Совершенствование условий труда персонала» Приключения в парке В. Маяковского. Одним субботним днем
Приключения в парке В. Маяковского. Одним субботним днем Политическая культура
Политическая культура Презентация на тему "Проект - это" - скачать презентации по Педагогике
Презентация на тему "Проект - это" - скачать презентации по Педагогике Выполнили: студенты ФФМО специальности «лечебное дело» группы 222 Буданова Анна Милинтеева Ксения.
Выполнили: студенты ФФМО специальности «лечебное дело» группы 222 Буданова Анна Милинтеева Ксения.  Геополитические отношения между Россией и Японией
Геополитические отношения между Россией и Японией Презентация "Дело в шляпе" - скачать презентации по МХК
Презентация "Дело в шляпе" - скачать презентации по МХК Noveyshaya_istoria_2
Noveyshaya_istoria_2 Основы религиозной культуры и светской этики
Основы религиозной культуры и светской этики Urok_trezvosti
Urok_trezvosti Механический мерительный инструмент с преобразователями
Механический мерительный инструмент с преобразователями Лабораторія промислових контролерів Unitronics VISION
Лабораторія промислових контролерів Unitronics VISION Парфенон
Парфенон Предел последовательности - презентация по Алгебре
Предел последовательности - презентация по Алгебре Политический процесс и политическое изменение
Политический процесс и политическое изменение Halloween. Корни этого праздника
Halloween. Корни этого праздника Презентация "Стратегический менеджмент" - скачать презентации по Экономике
Презентация "Стратегический менеджмент" - скачать презентации по Экономике