- Ферменты 2.ppt

Содержание
- 2. Выдвинутая в 1913 году Л. Михаэлисом и М. Ментен общая теория ферментативного катализа постулировала, что фермент
- 3. Структурно-функциональная организация ферментов. Схема Активные центры Якорные площадки Субстратный субстрат продукт Центры регуляции + и -
- 4. Структурно-функциональная организация ферментов. Активный (субстратный) центр - это совокупность функциональных групп, расположенных в разных участках ПП
- 5. Активный центр включает в себя: Каталитический участок или центр, непосредственно взаимодействующий с субстратом, осуществляющий катализ. Контактная,
- 6. 1 этап: постепенное «причаливание» S к «якорной» площадке F. 2 этап: напряжение и деформация: индуцированное соответствие
- 7. Химические связи, действующие при этом: 1. Силы Ван дер Ваальса 2. Электростатическое взаимодействие 3. Водородные связи
- 8. Якорный участок трипсина представлен длинным узким карманом с отрицательно заряженный Асп в глубине кармана. В этой
- 9. В гидрофобном кармане, образованном радикалами Гли, Три и Лей, химотрипсина располагается боковая цепь с ароматическим кольцом
- 10. В основе химических реакций лежит образование и разрыв химических связей По характеру разрыва ковалентных связей различают
- 11. Биологически важными нуклеофилами являются –NH2, -OH, -SH, и имидазольные группы аминокислот. Нуклеофильные формы этих групп одновременно
- 12. Электрофильные реагенты : Наиболее известными электрофилами в биохимических реакциях являются Н+, ионы Ме, углерод карбонильной группы.
- 13. По направлению реакций с учетом конечного результата можно выделить следующие типы реакций 1. Окислительно-восстановительные. Многие окислительно-
- 14. 2. Реакции кислотно- основного взаимодействия 3.Реакции замещения 4.Реакции отщепления 5.Реакции перегруппировки 6.реакции, сопровождающиеся образованием двойной связи
- 15. Факторы, определяющие активность ферментов [E], [S], [P], Km. Влияние pH, [P], tº, ионной силы на активность
- 16. Существенное влияние на активность ферментов оказывает реакция среды. Для проявления их оптимального действия чаще всего существует
- 17. В некоторых случаях сдвиг pH на единицу снижает активность фермента на 80%. Поэтому в экспериментальных условиях
- 18. Оптимум рН
- 19. Фермент pH Липаза (подж.железа) 8.0 Липаза (желудок) 4.0-5.0 Липаза(касторовое масло) 4.7 Пепсин 1.5-1.6 Трипсин 7- 8
- 20. Влияние температуры
- 21. Так как все ферменты являются белками, а белки при температуре выше 40-500 С в большинстве своем
- 22. Активность фермента повышается при повышении температуры. Начиная с определенной температуры, совпадающей с началом денатурации белка, активность
- 23. Специфичность ферментов Специфичность у разных ферментов может проявляться по-разному. Ферменты как белки, построены из L-аминокислот и
- 24. Правда, лишь небольшая часть ферментов обладает абсолютной специфичностью, т.е. катализирует превращение только одного субстрата. Чаще всего
- 25. Регуляция активности ферментов Регуляция активности ферментов бывает пассивная (с помощью изменения условий среды) т. е. есть
- 26. Активная регуляция: изостерическая;( изос- равный) регуляция с помощью субстрата (S)и продукта (Р) реакции. аллостерическая регуляция( allos-другой)
- 27. Регуляция путем изменения количества фермента. У бактерий хорошо изучен феномен индуцированного синтеза ферментов при выращивании на
- 28. Замена глюкозы на лактозу приводит к индуцированному синтезу фермента галактозидазы, расщепляющей лактозу на глюкозу и галактозу.
- 29. В животных тканях подобный быстрый синтез ферментов наблюдается реже, однако при поступлении в организм некоторых ядов,
- 30. С другой стороны, иногда под действием этих гидроксилаз чужеродные вещества превращаются в более токсичные продукты (летальный
- 31. Регуляция активности по принципу обратной связи. Допустим в клетке есть многоступенчатый биосинтетический процесс, каждая стадия которого
- 32. Аллостерическая регуляция. Аллостерические ферменты - это ферменты, располагающиеся в начале метаболического потока или на его узловых
- 33. Свойства аллостерических ферментов: 1. Являются олигомерами состоящими из протомеров. 2. Имеют как минимум два центра: активный
- 34. Существует 2 вида веществ (эффекторы), которые оказывают на фермент двоякое действие: 1)активаторы; 2) ингибиторы. Аллостерический фермент
- 35. При взаимодействии аллостерического фермента с аллостерическим активатором резко возрастает степень сродства фермента к субстрату, точнее возрастает
- 36. Кинетика ферментативных реакций
- 37. Имеется реакция: S → P + Q Представим эту реакцию в виде отдельных новых стадий: S
- 38. Основы термодинамики катализа Д. Кошланд предположил, что с термодинамической точки зрения ферменты ускоряют химические реакции за
- 39. Энергия активации Энергия активации - энергия, необходимая для перевода всех молекул моля вещества в активное состояние
- 40. Термодинамика ферментативных реакций Энергетический барьер G2 G1 ΔG = G1 – G2 GA1 GA2 Энергия активации
- 41. Термодинамика ферментативных реакций Энергетический барьер G2 G1 ΔG = G2 – G1 GA1 GA2 Энергия активации
- 42. Кинетика ферментативных реакций. Концентрация фермента. Чем выше концентрация E, тем выше скорость реакции. [E] V
- 43. Кинетика ферментативных реакций. pH Для каждого фермента существует оптимальная область pH (6,9 – 7,0 для большинства
- 44. Кинетика ферментативных реакций. Температура С увеличением температуры на 10°C скорость реакции возрастает в 2 раза (правило
- 45. Активность фермента зависит от концентрации субстратов. Исследование зависимости скорости ферментативных реакций от концентрации реагирующих веществ стало
- 46. Кинетика ферментативных реакций
- 47. Если концентрация субстрата [S] очень низкая, ограничивающим скорость реакции становится этап образования комплекса ES (связывание) и
- 48. Если концентрация субстрата [S] высокая, количество образующегося комплекса ES зависит от количества доступного фермента, и скорость
- 49. Ограничивающим скорость реакции становится этап образования продукта (катализ). В реакции, катализируемой ферментом имеется верхний предел скорости
- 50. Кинетика ферментативных реакций. Концентрация субстрата Для простых ферментов график имеет вид гиперболы и описывается уравнением Михаэлиса-Ментен.
- 51. [ES]max = [Et]общ Общее количество фермента в системе, равно сумме [E] (концентрация свободного фермента), и [ES]
- 52. Важным для правильной оценки результатов исследования зависимости скорости реакции от концентрации является измерение начальной скорости реакции.
- 53. При взаимодействии фермента и субстрата очень быстро наступает равновесие между скоростью образования и скоростью распада фермент
- 54. В реакции, катализируемой ферментом можно выделить четыре реакции, каждая из которых характеризуется собственной константой скорости. Однако,
- 55. Важной качественной характеристикой фермента является константа Михаэлиса Воспользовавшись предположениями, высказанными Генри, Михаэлисом и Ментен, а также
- 56. Основная гипотеза: этапом, ограничивающим скорость ферментативной реакции является ( ES → E + P ) отсюда
- 57. тогда, количество свободного фермента: [Et] - [ES] так как [S] >> [Et] , [S]связ [этап 1]
- 58. [этап 2] гипотеза: Образование фермент-субстратного комплекса самая быстрая реакция, результатом которой является возникновение динамического равновесия между
- 59. [Этап 3] k1 [Et] [S] - k1 [ES] [S] = [ES] (k2 + k3) k1 [Et]
- 60. отсюда v0 = k3 [ES] = k3 {[Et] [S] / { [S] + Km } =
- 61. Уравнение Михаэлиса и Ментен графически – прямоугольная гипербола Если мы простроим график зависимости скорости реакции V
- 63. Каково физическое значение Km? Уравнение Михаэлиса-Ментен можно преобразовать к такому виду
- 64. Из этого уравнения легко показать, что при [S] =10 Kmv/Vmax = 0.91при [S] = Kmv/ Vmax
- 65. При условии, что k3
- 66. Значение Km дает также некоторые представления относительно эффективности катализа и регуляции. Если [S]>> 10 Km, реакция
- 67. Если [S]
- 68. Наиболее удобное сочетание эффективности и контроля соблюдается при условии, если концентрация субстрата одного порядка со значениями
- 69. Знание Км позволит Вам оценивать концентрацию субстрата, необходимую для гарантии насыщения. Эта концентрация должна быть равна
- 70. В физиологических условиях, для эффективной работы, концентрация субстрата должна быть на уровне Км этого фермента, но
- 71. Практически рассчитать значения Км и Vmax, пользуясь кривой, описываемой уравнением Михаэлис и Ментен сложно. Более удобно
- 72. Такой способ выражения позволяет более точно рассчитать значения Км и V. Пересечение линии с осью 1/[S]
- 74. Примеры использования данных кинетических исследований ферментов в медицине Некоторые люди обладают повышенной чувствительностью к этиловому спирту.
- 75. Альдегиддегидрогеназа обычно существует в двух формах, с высоким сродством (низкие значения Км) к альдегиду и с
- 76. Семейная пара с генетической предрасположенностью к болезни Неймана-Пика ожидает ребенка. Им известно, что их будущий малыш
- 77. Если сфингомиэлин не распадается должным образом, нарушается нейронная передача. Физиологические последствия болезни Наймана-Пика - олигофрения и
- 78. Семейная пара с генетической предрасположенностью к болезни Неймана-Пика ожидает ребенка. Им известно, что их будущий малыш
- 79. У беременной были получены клетки плода (путем амниоцентеза) и размножены методом тканевой культуры. Экстракт клеток был
- 81. При знакомстве с материалом по кинетике ферментов могло сложиться впечатление, что кинетика всех ферментов основана на
- 82. У ферментов, подчиняющимся принципам кинетики Михаэлиса и Ментен с позиций процессов, протекающих в клетке, имеется ряд
- 83. Таким образом, природа вынуждена обратиться к "кооперативным" системам, в которых маленькие изменения в одном параметре, например
- 85. Аллостерический белок определяется как белок, содержащий два или больше топологически различающихся центра связывания лигандов (субстраты, ингибиторы
- 86. Кооперативность - модификация константы связывания лиганда белком предшествующим связыванием другого лиганда. Константы связывания - подобны Ks
- 87. . Км обычно принимается как константа связывания субстрата, поскольку ее проще измерять, чем Ks. Понятие кооперативность
- 88. Значения Vmax и Km определяются при экстраполяции линий до пересечения с осью абсцисс и ординат. Как
- 89. Два лиганда один из которых влияет на связывание другого, могут быть химически идентичны, например, одна молекула
- 90. У аллостерических ферментов особые свойства Полимерная структура Сигмоидная ( в отличие от гиперболической для не аллостерических)
- 91. Аллостерический фермент содержит ряд активных центров, в самом простом случае по одному на субъединицу, каждый из
- 92. которое увеличит способность других активных центров связывать лиганды (положительная кооперативность. K2
- 94. Центральная линия графика - типичная сигмовидная кривая в отсутствии любого эффектора. В присутствии активатора (А) скорость
- 95. Денатурация – нарушение пространственной структуры фермента с последующей потерей активности фермента. Денатурация вызывается рядом факторов, включая
- 96. Ингибиторы бывают разные: обратимые и необратимые Вещества со свойствами ингибиторов ферментов можно грубо разделить на обратимые
- 97. Необратимые ингибиторы известны также как инактиваторы фермента. Они связываются с ферментом, формируя прочные, обычно ковалентные связи:
- 98. Среди примеров необратимо действующих ингибиторов можно назвать диизопропилфторфосфат (ДИПФФ). Это соединение вошло в историю энзимологии как
- 99. Ацетилсалициловая кислота (известный всем аспирин) является необратимым ингибитором циклооксигеназы- фермента участвующего в синтезе простагландинов.
- 100. Ингибитором синтеза протеогликанов стенки бактерий является пенилиллин, структура которого напоминает D-аланин, встраиваемый в структуру протеогликанов. Связываясь
- 101. Предшествующее связывание субстрата к активному центру в свою очередь, вызывает изменения конформации центра связывания ингибитора, которое
- 102. Различают два механизма конкурентного торможения. 1.Конкурентное торможение путем связывания активного центра. 2. Конкурентное торможение путем изменения
- 103. Конкурентные ингибиторы не влияют на Vmax, они понижают Км. Оказывая эффект на: Km Эффект на Vmax
- 104. Малоновая кислота тормозит активность дегидрогеназы янтарной кислоты, занимая активный центр на ферменте. Учитывая обратимость реакции, избыток
- 105. Конкурентное торможение путем изменения конформации фермента. В отличие от классического варианта, ингибитор связывается не с активным
- 106. Регуляция активности ферментов путем химической модификации: Реакции ограниченного протеолиза, аденилирования, рибозилирования, ацетилирования, фосфорилирования (роль гормонов, АЦ-комплекса,
- 107. Активность фермента можно изменить путем ковалентной модификации его структуры. Ковалентная модификация структуры ферментов может быть обратимой
- 108. Регуляция активности ферментов Гормональная регуляция осуществляется на генетическом уровне путем обратного фосфорилирования. Например под действием адреналина
- 109. Гормон, циркулирующий в крови, попадает в межклеточную жидкость и контактирует с поверхностью клетки, где расположены рецепторы
- 110. Уровень, образовавшийся под действием АЦ, цАМФ определяется не только активностью АЦ, но и активностью фосфодиэстераз, которые
- 111. цГМФ оказывает эффекты противоположные цАМФ. цАМФ находится в тесном контакте с ионами Ca2+: высокая концентрация цАМФ
- 112. Изоферменты, их природа, биологическая роль, строение ЛДГ. Изоферменты - это группа родственных ферментов, катализирующих одну и
- 113. 1) скорстью катализа; 2) направлением катализируемой реакции; 3) условиями протекания реакции; 4) чувствительностью к регуляторам, факторам
- 114. Изоферменты имеют адаптивное значение, т. е. придают специфику метаболизма. Изоферменты обеспечивают межорганную связь, например, в процессе
- 115. ЛДГ4,5 в печени: ПВК -----> лактат ЛДГ1,2 в сердце: лактат ------> ПВК ЛДГ - олигомерный фермент,
- 116. Существует 5 изоферментных форм: HHHH HHHM HHMM HMMM MMMM H4 H3M H2M2 HM3 M4 ЛДГ1, ЛДГ2,
- 117. Остальные изоферменты занимают промежуточное положение. Изоферменты ЛДГ локализованы в различных тканях: ЛДГ1,2 ----> мозг, аэробные ткани
- 118. Изменение активности ферментов в онтогенезе. Онтогенез человека развивается по определенной генетической программе, которая записана на уровне
- 119. Глюкоза утилизируется анаэробным путем (анаэробный гликолиз), т. к. ткани плода не способны к ГНГ, и идет
- 120. 3. Грудной Потребность в глюкозе резко возрастает, она начинает утилизироваться аэробно, но примерно до двух лет
- 122. Скачать презентацию

Выдвинутая в 1913 году Л. Михаэлисом и
М. Ментен общая
Выдвинутая в 1913 году Л. Михаэлисом и
М. Ментен общая
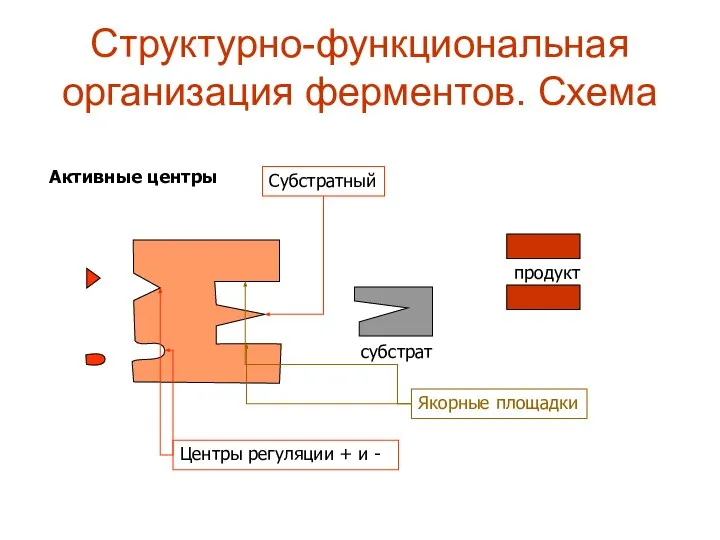
Структурно-функциональная организация ферментов. Схема
Активные центры
Якорные площадки
Субстратный
субстрат
продукт
Центры регуляции + и -
Структурно-функциональная организация ферментов. Схема
Активные центры
Якорные площадки
Субстратный
субстрат
продукт
Центры регуляции + и -

Структурно-функциональная организация ферментов.
Активный (субстратный) центр - это совокупность функциональных групп,
Структурно-функциональная организация ферментов.
Активный (субстратный) центр - это совокупность функциональных групп,

Активный центр включает в себя:
Каталитический участок или центр, непосредственно взаимодействующий с
Активный центр включает в себя:
Каталитический участок или центр, непосредственно взаимодействующий с

1 этап: постепенное «причаливание» S к «якорной» площадке F.
2 этап: напряжение
1 этап: постепенное «причаливание» S к «якорной» площадке F.
2 этап: напряжение

Химические связи, действующие при этом:
1. Силы Ван дер Ваальса
2. Электростатическое взаимодействие
3.
Химические связи, действующие при этом:
1. Силы Ван дер Ваальса
2. Электростатическое взаимодействие
3.

Якорный участок трипсина представлен длинным узким карманом с отрицательно заряженный Асп
Якорный участок трипсина представлен длинным узким карманом с отрицательно заряженный Асп
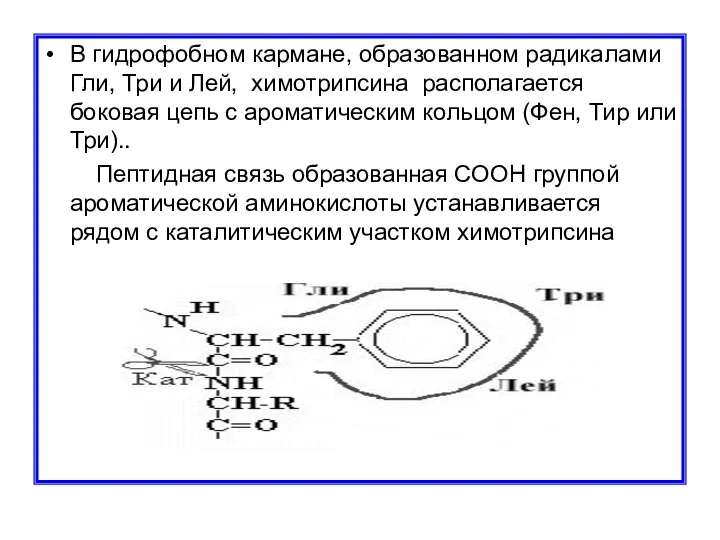
В гидрофобном кармане, образованном радикалами Гли, Три и Лей, химотрипсина располагается
В гидрофобном кармане, образованном радикалами Гли, Три и Лей, химотрипсина располагается

В основе химических реакций лежит образование и разрыв химических связей
По
В основе химических реакций лежит образование и разрыв химических связей
По

Биологически важными нуклеофилами являются –NH2, -OH, -SH, и имидазольные группы
Биологически важными нуклеофилами являются –NH2, -OH, -SH, и имидазольные группы

Электрофильные реагенты : Наиболее известными электрофилами в биохимических реакциях являются Н+,
Электрофильные реагенты : Наиболее известными электрофилами в биохимических реакциях являются Н+,

По направлению реакций с учетом конечного результата можно выделить следующие типы
По направлению реакций с учетом конечного результата можно выделить следующие типы

2. Реакции кислотно- основного взаимодействия
3.Реакции замещения
4.Реакции отщепления
5.Реакции перегруппировки
6.реакции, сопровождающиеся образованием двойной
3.Реакции замещения
4.Реакции отщепления
5.Реакции перегруппировки
6.реакции, сопровождающиеся образованием двойной
![Факторы, определяющие активность ферментов [E], [S], [P], Km. Влияние pH, [P],](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1292266/slide-14.jpg)
Факторы, определяющие активность ферментов [E], [S], [P], Km.
Влияние pH, [P],
Влияние pH, [P],

Существенное влияние на активность ферментов оказывает реакция среды. Для проявления
Существенное влияние на активность ферментов оказывает реакция среды. Для проявления

В некоторых случаях сдвиг pH на единицу снижает активность фермента на
В некоторых случаях сдвиг pH на единицу снижает активность фермента на
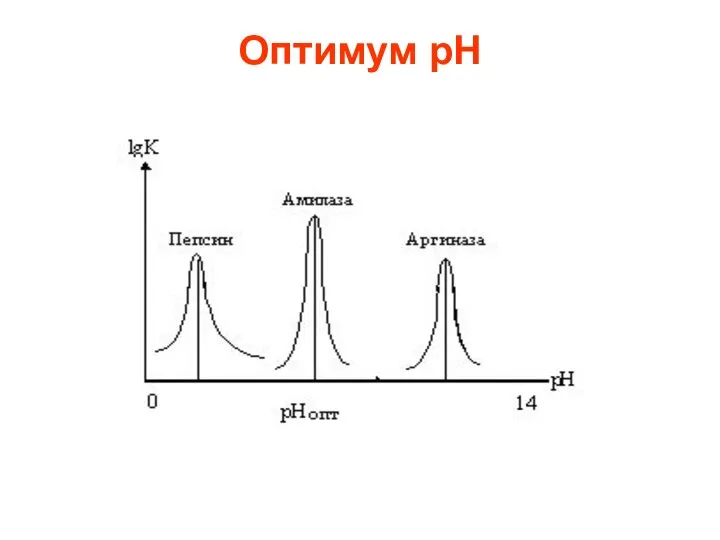
Оптимум рН
Оптимум рН
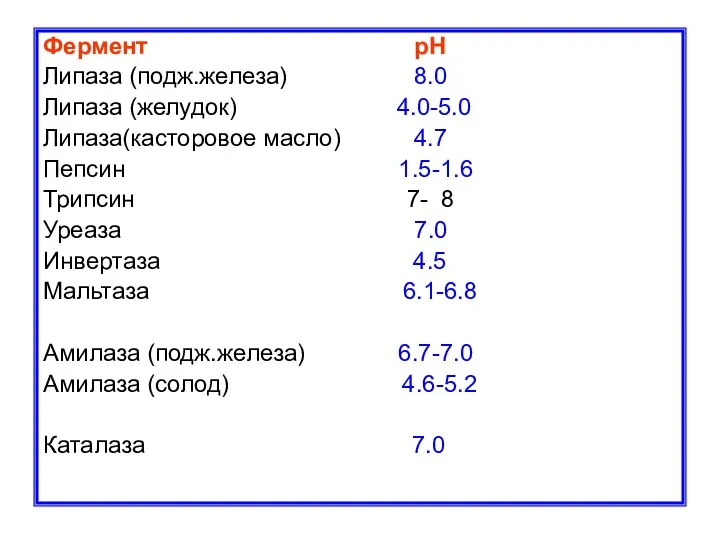
Фермент pH
Липаза (подж.железа) 8.0
Липаза (желудок) 4.0-5.0
Липаза(касторовое масло) 4.7
Пепсин 1.5-1.6
Трипсин 7-
Фермент pH
Липаза (подж.железа) 8.0
Липаза (желудок) 4.0-5.0
Липаза(касторовое масло) 4.7
Пепсин 1.5-1.6
Трипсин 7-
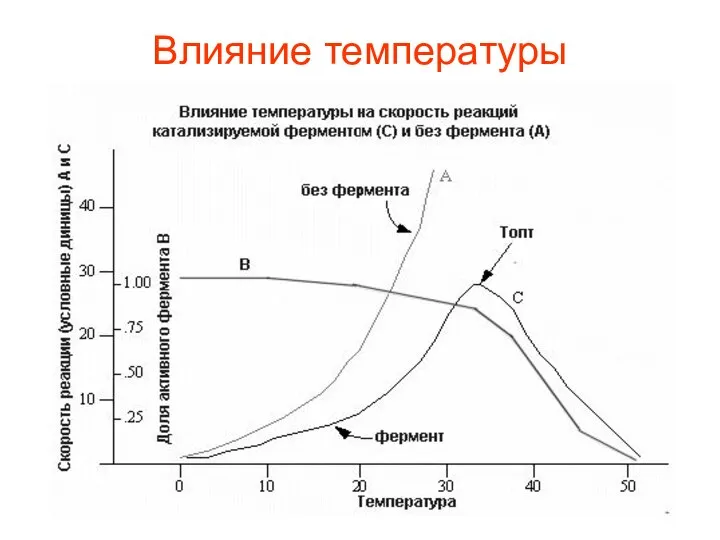
Влияние температуры
Влияние температуры

Так как все ферменты являются белками, а белки при температуре выше
Так как все ферменты являются белками, а белки при температуре выше

Активность фермента повышается при повышении температуры. Начиная с определенной температуры, совпадающей
Активность фермента повышается при повышении температуры. Начиная с определенной температуры, совпадающей

Специфичность ферментов
Специфичность у разных ферментов может проявляться по-разному. Ферменты как
Специфичность ферментов
Специфичность у разных ферментов может проявляться по-разному. Ферменты как

Правда, лишь небольшая часть ферментов обладает абсолютной специфичностью, т.е. катализирует превращение
Правда, лишь небольшая часть ферментов обладает абсолютной специфичностью, т.е. катализирует превращение

Регуляция активности ферментов
Регуляция активности ферментов бывает пассивная (с помощью изменения условий
Регуляция активности ферментов
Регуляция активности ферментов бывает пассивная (с помощью изменения условий

Активная регуляция:
изостерическая;( изос- равный) регуляция с помощью субстрата (S)и продукта (Р)
Активная регуляция:
изостерическая;( изос- равный) регуляция с помощью субстрата (S)и продукта (Р)

Регуляция путем изменения количества фермента.
У бактерий хорошо изучен феномен индуцированного синтеза
Регуляция путем изменения количества фермента.
У бактерий хорошо изучен феномен индуцированного синтеза

Замена глюкозы на лактозу приводит к индуцированному синтезу фермента галактозидазы, расщепляющей
Замена глюкозы на лактозу приводит к индуцированному синтезу фермента галактозидазы, расщепляющей

В животных тканях подобный быстрый синтез ферментов наблюдается реже, однако при
В животных тканях подобный быстрый синтез ферментов наблюдается реже, однако при

С другой стороны, иногда под действием этих гидроксилаз чужеродные вещества превращаются
С другой стороны, иногда под действием этих гидроксилаз чужеродные вещества превращаются

Регуляция активности по принципу обратной связи.
Допустим в клетке есть многоступенчатый
Регуляция активности по принципу обратной связи.
Допустим в клетке есть многоступенчатый

Аллостерическая регуляция.
Аллостерические ферменты - это ферменты, располагающиеся в начале
Аллостерическая регуляция.
Аллостерические ферменты - это ферменты, располагающиеся в начале

Свойства аллостерических ферментов:
1. Являются олигомерами состоящими из протомеров.
2. Имеют как минимум два
Свойства аллостерических ферментов:
1. Являются олигомерами состоящими из протомеров.
2. Имеют как минимум два

Существует 2 вида веществ (эффекторы), которые оказывают на фермент двоякое действие:
1)активаторы;
Существует 2 вида веществ (эффекторы), которые оказывают на фермент двоякое действие:
1)активаторы;

При взаимодействии аллостерического фермента с аллостерическим активатором резко возрастает степень сродства
При взаимодействии аллостерического фермента с аллостерическим активатором резко возрастает степень сродства

Кинетика ферментативных реакций
Кинетика ферментативных реакций

Имеется реакция:
S → P + Q
Представим эту реакцию в виде
Имеется реакция:
S → P + Q
Представим эту реакцию в виде

Основы термодинамики катализа
Д. Кошланд предположил, что с термодинамической точки зрения ферменты
Основы термодинамики катализа
Д. Кошланд предположил, что с термодинамической точки зрения ферменты

Энергия активации
Энергия активации - энергия, необходимая для перевода всех молекул моля
Энергия активации
Энергия активации - энергия, необходимая для перевода всех молекул моля
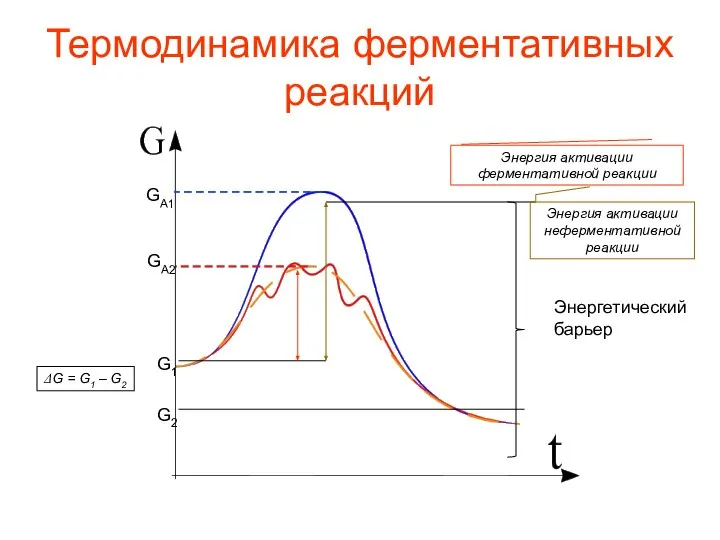
Термодинамика ферментативных реакций
Энергетический
барьер
G2
G1
ΔG = G1 – G2
GA1
GA2
Энергия активации неферментативной реакции
Энергия активации
Термодинамика ферментативных реакций
Энергетический
барьер
G2
G1
ΔG = G1 – G2
GA1
GA2
Энергия активации неферментативной реакции
Энергия активации
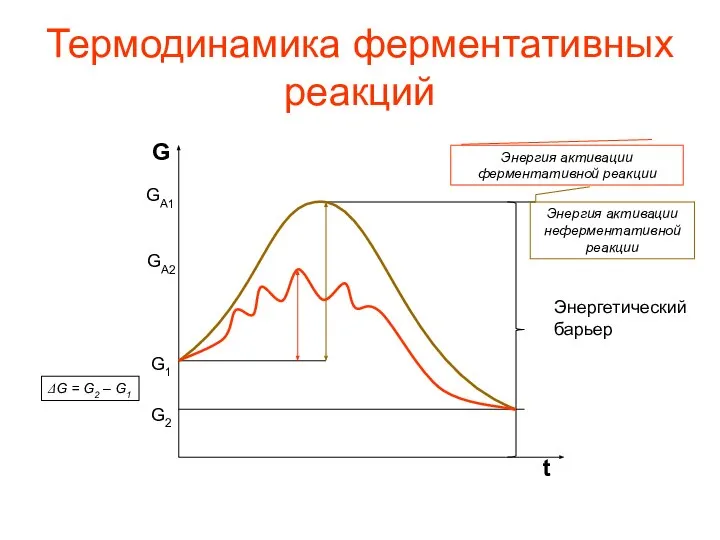
Термодинамика ферментативных реакций
Энергетический
барьер
G2
G1
ΔG = G2 – G1
GA1
GA2
Энергия активации неферментативной реакции
Энергия активации
Термодинамика ферментативных реакций
Энергетический
барьер
G2
G1
ΔG = G2 – G1
GA1
GA2
Энергия активации неферментативной реакции
Энергия активации
![Кинетика ферментативных реакций. Концентрация фермента. Чем выше концентрация E, тем выше скорость реакции. [E] V](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1292266/slide-41.jpg)
Кинетика ферментативных реакций. Концентрация фермента.
Чем выше концентрация E, тем выше скорость
Кинетика ферментативных реакций. Концентрация фермента.
Чем выше концентрация E, тем выше скорость
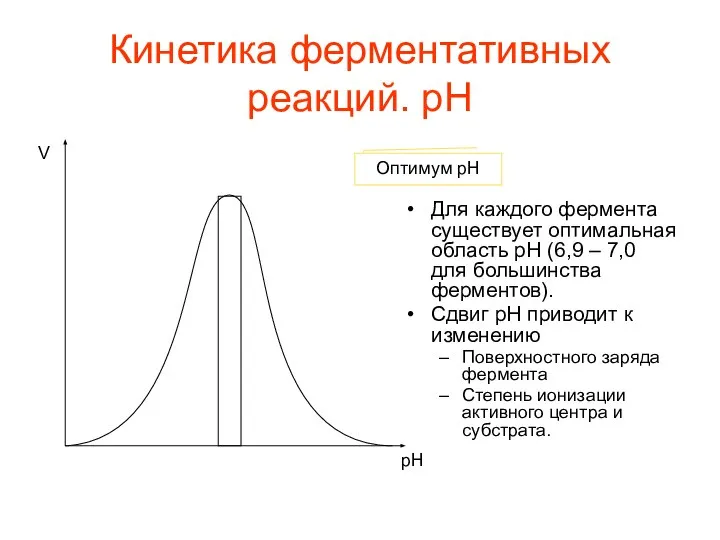
Кинетика ферментативных реакций. pH
Для каждого фермента существует оптимальная область pH (6,9
Кинетика ферментативных реакций. pH
Для каждого фермента существует оптимальная область pH (6,9
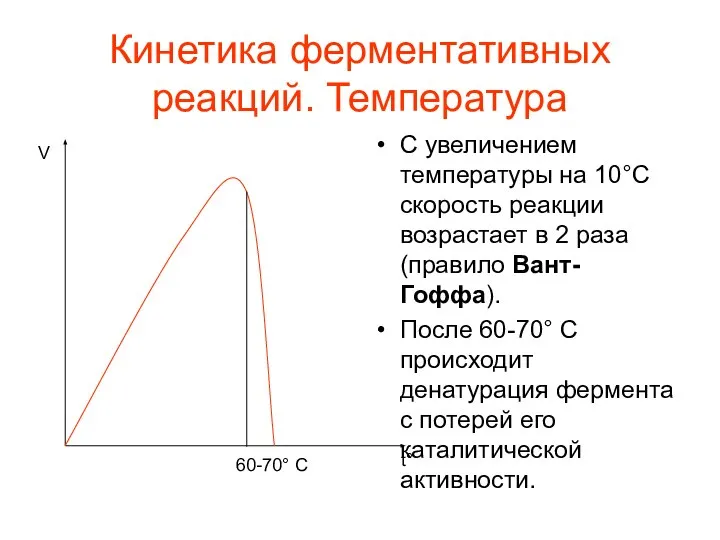
Кинетика ферментативных реакций. Температура
С увеличением температуры на 10°C скорость реакции возрастает
Кинетика ферментативных реакций. Температура
С увеличением температуры на 10°C скорость реакции возрастает

Активность фермента зависит от концентрации субстратов.
Исследование зависимости скорости ферментативных реакций
Активность фермента зависит от концентрации субстратов.
Исследование зависимости скорости ферментативных реакций
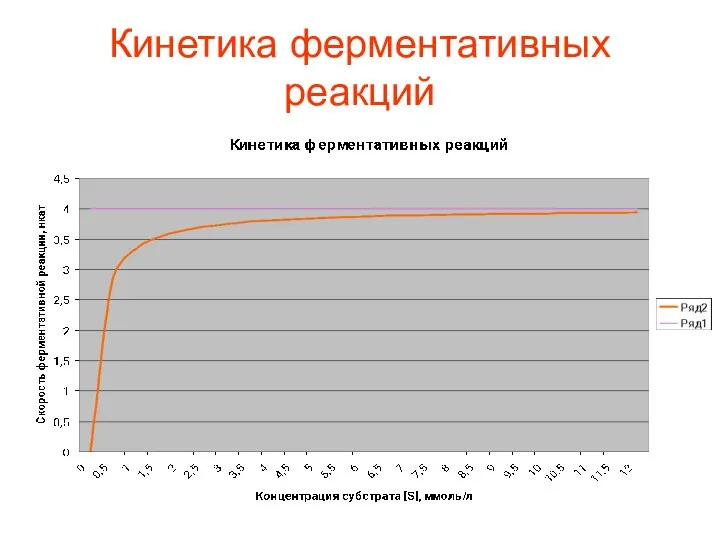
Кинетика ферментативных реакций
Кинетика ферментативных реакций
![Если концентрация субстрата [S] очень низкая, ограничивающим скорость реакции становится этап](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1292266/slide-46.jpg)
Если концентрация субстрата [S] очень низкая, ограничивающим скорость реакции становится этап
Если концентрация субстрата [S] очень низкая, ограничивающим скорость реакции становится этап
![Если концентрация субстрата [S] высокая, количество образующегося комплекса ES зависит от](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1292266/slide-47.jpg)
Если концентрация субстрата [S] высокая, количество образующегося комплекса ES зависит
Если концентрация субстрата [S] высокая, количество образующегося комплекса ES зависит

Ограничивающим скорость реакции становится этап образования продукта (катализ). В реакции, катализируемой
Ограничивающим скорость реакции становится этап образования продукта (катализ). В реакции, катализируемой
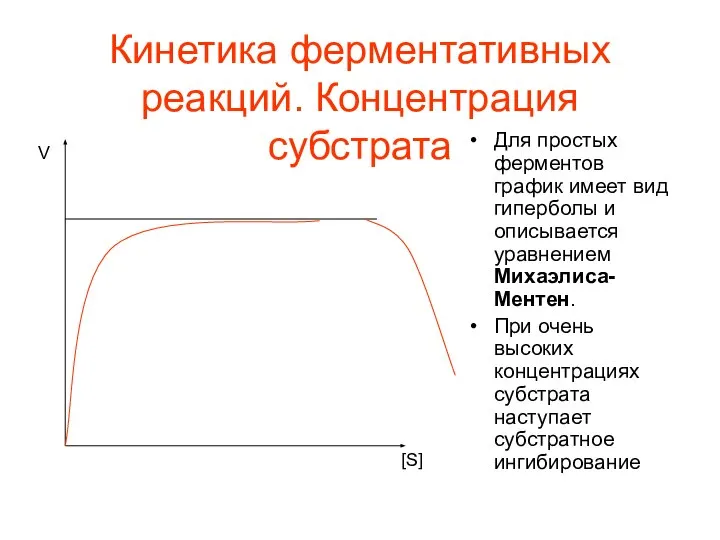
Кинетика ферментативных реакций. Концентрация субстрата
Для простых ферментов график имеет вид гиперболы
Кинетика ферментативных реакций. Концентрация субстрата
Для простых ферментов график имеет вид гиперболы
![[ES]max = [Et]общ Общее количество фермента в системе, равно сумме [E]](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1292266/slide-50.jpg)
[ES]max = [Et]общ
Общее количество фермента в системе, равно сумме [E]
[ES]max = [Et]общ
Общее количество фермента в системе, равно сумме [E]

Важным для правильной оценки результатов исследования зависимости скорости реакции от
Важным для правильной оценки результатов исследования зависимости скорости реакции от

При взаимодействии фермента и субстрата очень быстро наступает равновесие между скоростью
При взаимодействии фермента и субстрата очень быстро наступает равновесие между скоростью

В реакции, катализируемой ферментом можно выделить четыре реакции, каждая из которых
В реакции, катализируемой ферментом можно выделить четыре реакции, каждая из которых

Важной качественной характеристикой фермента является константа Михаэлиса
Воспользовавшись предположениями, высказанными Генри,
Важной качественной характеристикой фермента является константа Михаэлиса
Воспользовавшись предположениями, высказанными Генри,

Основная гипотеза: этапом, ограничивающим скорость ферментативной реакции является ( ES →
Основная гипотеза: этапом, ограничивающим скорость ферментативной реакции является ( ES →
![тогда, количество свободного фермента: [Et] - [ES] так как [S] >>](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1292266/slide-56.jpg)
тогда, количество свободного фермента: [Et] - [ES]
так как [S] >>
тогда, количество свободного фермента: [Et] - [ES]
так как [S] >>
![[этап 2] гипотеза: Образование фермент-субстратного комплекса самая быстрая реакция, результатом которой](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1292266/slide-57.jpg)
[этап 2] гипотеза: Образование фермент-субстратного комплекса самая быстрая реакция, результатом которой является
[этап 2] гипотеза: Образование фермент-субстратного комплекса самая быстрая реакция, результатом которой является
![[Этап 3] k1 [Et] [S] - k1 [ES] [S] = [ES]](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1292266/slide-58.jpg)
[Этап 3] k1 [Et] [S] - k1 [ES] [S] = [ES]
[Этап 3] k1 [Et] [S] - k1 [ES] [S] = [ES]
![отсюда v0 = k3 [ES] = k3 {[Et] [S] / {](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1292266/slide-59.jpg)
отсюда v0 = k3 [ES] = k3 {[Et] [S] / {
отсюда v0 = k3 [ES] = k3 {[Et] [S] / {
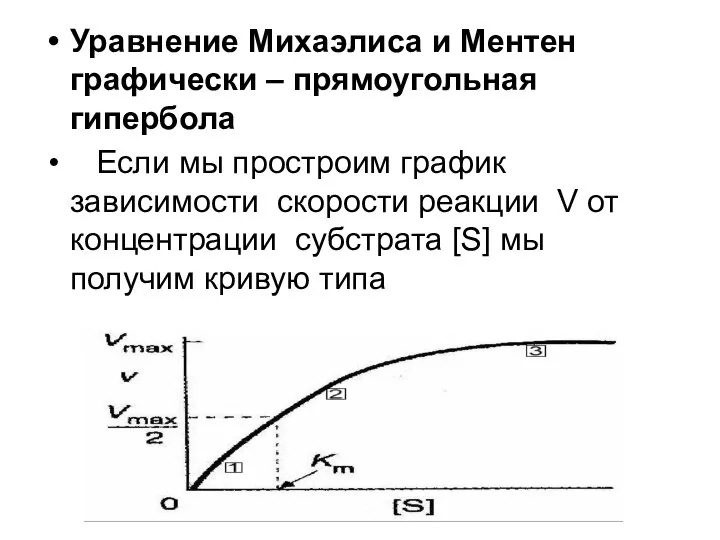
Уравнение Михаэлиса и Ментен графически – прямоугольная гипербола
Если мы простроим
Уравнение Михаэлиса и Ментен графически – прямоугольная гипербола
Если мы простроим
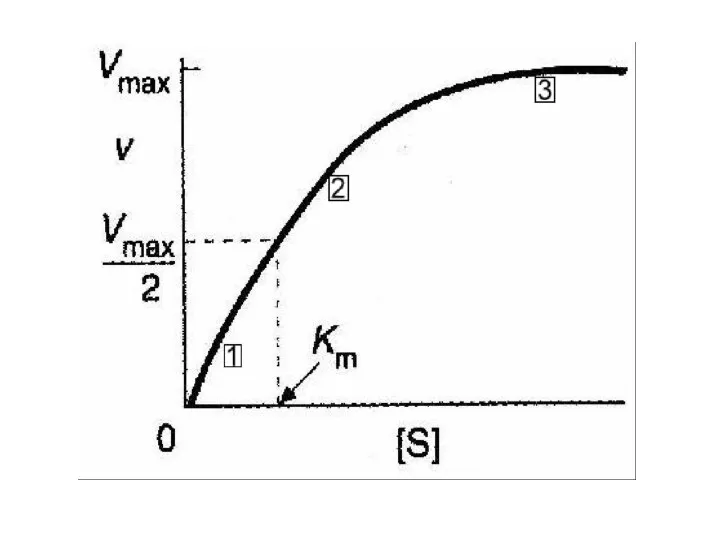

Каково физическое значение Km? Уравнение Михаэлиса-Ментен можно преобразовать к такому виду
Каково физическое значение Km? Уравнение Михаэлиса-Ментен можно преобразовать к такому виду
![Из этого уравнения легко показать, что при [S] =10 Kmv/Vmax =](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1292266/slide-63.jpg)
Из этого уравнения легко показать, что
при [S] =10 Kmv/Vmax =
Из этого уравнения легко показать, что
при [S] =10 Kmv/Vmax =

При условии, что k3 << k2, константа Михаэлиса становится хорошим показателем
При условии, что k3 << k2, константа Михаэлиса становится хорошим показателем

Значение Km дает также некоторые представления относительно эффективности катализа и регуляции.
Значение Km дает также некоторые представления относительно эффективности катализа и регуляции.
![Если [S]](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1292266/slide-66.jpg)
Если [S] << 0.1 Km, эффективность реакции низка, но имеется хорошее
Если [S] << 0.1 Km, эффективность реакции низка, но имеется хорошее

Наиболее удобное сочетание эффективности и контроля соблюдается при условии, если концентрация
Наиболее удобное сочетание эффективности и контроля соблюдается при условии, если концентрация

Знание Км позволит Вам оценивать концентрацию субстрата, необходимую для гарантии насыщения.
Знание Км позволит Вам оценивать концентрацию субстрата, необходимую для гарантии насыщения.

В физиологических условиях, для эффективной работы, концентрация субстрата должна быть на
В физиологических условиях, для эффективной работы, концентрация субстрата должна быть на

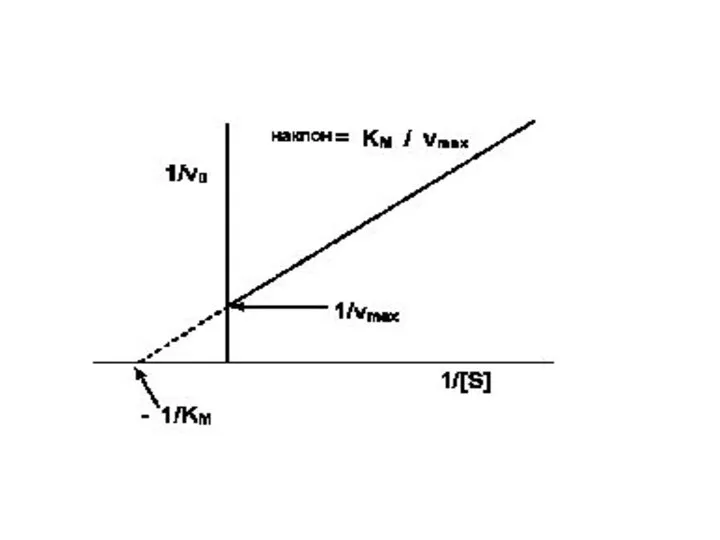
Практически рассчитать значения Км и Vmax, пользуясь кривой, описываемой уравнением
Практически рассчитать значения Км и Vmax, пользуясь кривой, описываемой уравнением

Такой способ выражения позволяет более точно рассчитать значения Км и
Такой способ выражения позволяет более точно рассчитать значения Км и

Примеры использования данных кинетических исследований ферментов в медицине
Некоторые люди обладают
Примеры использования данных кинетических исследований ферментов в медицине
Некоторые люди обладают

Альдегиддегидрогеназа обычно существует в двух формах, с высоким сродством (низкие значения
Альдегиддегидрогеназа обычно существует в двух формах, с высоким сродством (низкие значения

Семейная пара с генетической предрасположенностью к болезни Неймана-Пика ожидает ребенка. Им
Семейная пара с генетической предрасположенностью к болезни Неймана-Пика ожидает ребенка. Им

Если сфингомиэлин не распадается должным образом, нарушается нейронная передача. Физиологические последствия
Если сфингомиэлин не распадается должным образом, нарушается нейронная передача. Физиологические последствия

Семейная пара с генетической предрасположенностью к болезни Неймана-Пика ожидает ребенка. Им
Семейная пара с генетической предрасположенностью к болезни Неймана-Пика ожидает ребенка. Им

У беременной были получены клетки плода (путем амниоцентеза) и размножены методом
У беременной были получены клетки плода (путем амниоцентеза) и размножены методом

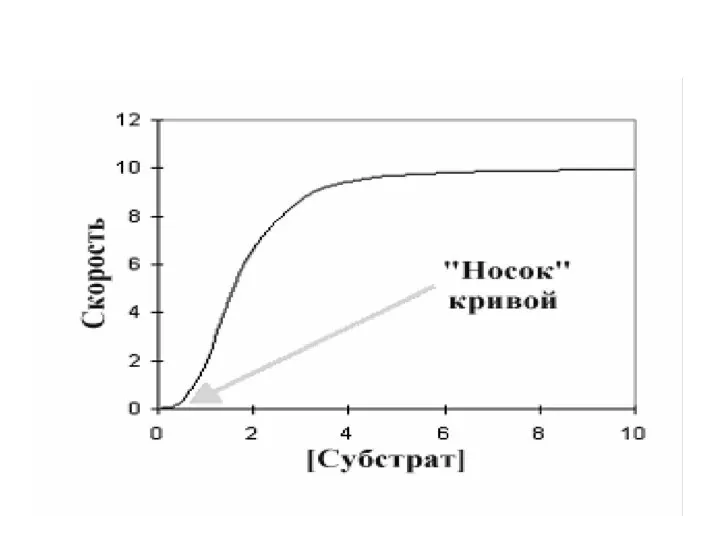
При знакомстве с материалом по кинетике ферментов могло сложиться впечатление, что кинетика
При знакомстве с материалом по кинетике ферментов могло сложиться впечатление, что кинетика

У ферментов, подчиняющимся принципам кинетики Михаэлиса и Ментен с позиций процессов,
У ферментов, подчиняющимся принципам кинетики Михаэлиса и Ментен с позиций процессов,

Таким образом, природа вынуждена обратиться к "кооперативным" системам, в которых маленькие
Таким образом, природа вынуждена обратиться к "кооперативным" системам, в которых маленькие

Аллостерический белок определяется как белок, содержащий два или больше топологически различающихся
Аллостерический белок определяется как белок, содержащий два или больше топологически различающихся

Кооперативность - модификация константы связывания лиганда белком предшествующим связыванием другого лиганда.
Кооперативность - модификация константы связывания лиганда белком предшествующим связыванием другого лиганда.

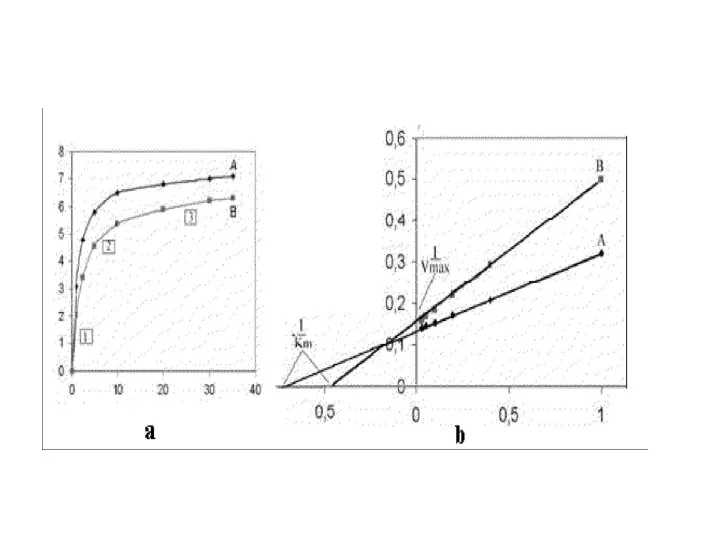
. Км обычно принимается как константа связывания субстрата, поскольку ее проще
. Км обычно принимается как константа связывания субстрата, поскольку ее проще

Значения Vmax и Km определяются при экстраполяции линий до пересечения с
Значения Vmax и Km определяются при экстраполяции линий до пересечения с

Два лиганда один из которых влияет на связывание другого, могут быть
Два лиганда один из которых влияет на связывание другого, могут быть

У аллостерических ферментов особые свойства
Полимерная структура
Сигмоидная ( в отличие от гиперболической
У аллостерических ферментов особые свойства
Полимерная структура
Сигмоидная ( в отличие от гиперболической

Аллостерический фермент содержит ряд активных центров, в самом простом случае по
Аллостерический фермент содержит ряд активных центров, в самом простом случае по

которое увеличит способность других активных центров связывать лиганды (положительная кооперативность. K2
которое увеличит способность других активных центров связывать лиганды (положительная кооперативность. K2
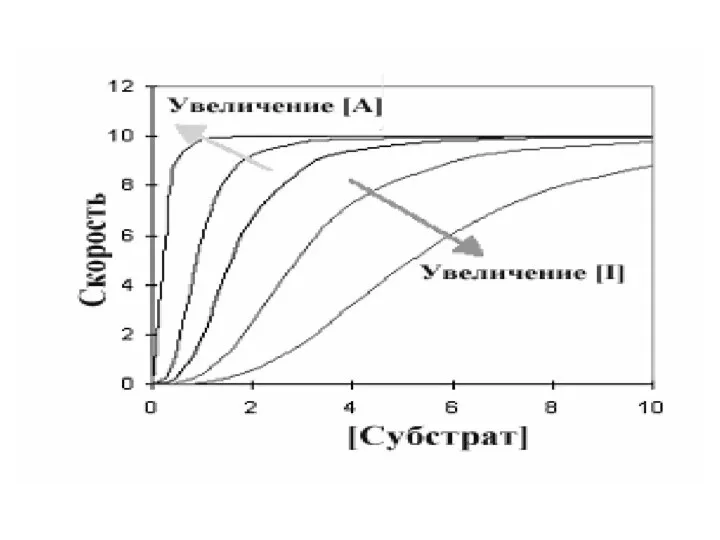

Центральная линия графика - типичная сигмовидная кривая в отсутствии любого эффектора.
Центральная линия графика - типичная сигмовидная кривая в отсутствии любого эффектора.

Денатурация – нарушение пространственной структуры фермента с последующей потерей активности фермента.
Денатурация – нарушение пространственной структуры фермента с последующей потерей активности фермента.

Ингибиторы бывают разные: обратимые и необратимые
Вещества со свойствами ингибиторов ферментов можно
Ингибиторы бывают разные: обратимые и необратимые
Вещества со свойствами ингибиторов ферментов можно

Необратимые ингибиторы известны также как инактиваторы фермента. Они связываются с ферментом,
Необратимые ингибиторы известны также как инактиваторы фермента. Они связываются с ферментом,

Среди примеров необратимо действующих ингибиторов можно назвать диизопропилфторфосфат (ДИПФФ). Это соединение
Среди примеров необратимо действующих ингибиторов можно назвать диизопропилфторфосфат (ДИПФФ). Это соединение
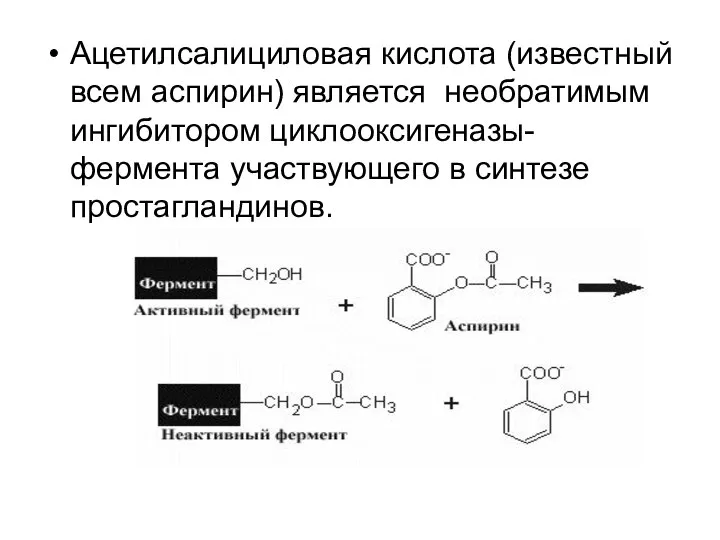
Ацетилсалициловая кислота (известный всем аспирин) является необратимым ингибитором циклооксигеназы- фермента участвующего
Ацетилсалициловая кислота (известный всем аспирин) является необратимым ингибитором циклооксигеназы- фермента участвующего

Ингибитором синтеза протеогликанов стенки бактерий является пенилиллин, структура которого напоминает D-аланин,
Ингибитором синтеза протеогликанов стенки бактерий является пенилиллин, структура которого напоминает D-аланин,

Предшествующее связывание субстрата к активному центру в свою очередь, вызывает изменения
Предшествующее связывание субстрата к активному центру в свою очередь, вызывает изменения

Различают два механизма конкурентного торможения.
1.Конкурентное торможение путем связывания активного центра.
2.
Различают два механизма конкурентного торможения.
1.Конкурентное торможение путем связывания активного центра.
2.

Конкурентные ингибиторы не влияют на Vmax, они понижают Км.
Оказывая эффект на:
Конкурентные ингибиторы не влияют на Vmax, они понижают Км.
Оказывая эффект на:

Малоновая кислота тормозит активность дегидрогеназы янтарной кислоты, занимая активный центр на
Малоновая кислота тормозит активность дегидрогеназы янтарной кислоты, занимая активный центр на

Конкурентное торможение путем изменения конформации фермента.
В отличие от классического варианта,
Конкурентное торможение путем изменения конформации фермента.
В отличие от классического варианта,

Регуляция активности ферментов путем химической модификации: Реакции ограниченного протеолиза, аденилирования, рибозилирования,
Регуляция активности ферментов путем химической модификации: Реакции ограниченного протеолиза, аденилирования, рибозилирования,

Активность фермента можно изменить путем ковалентной модификации его структуры.
Ковалентная модификация структуры
Активность фермента можно изменить путем ковалентной модификации его структуры.
Ковалентная модификация структуры

Регуляция активности ферментов
Гормональная регуляция осуществляется на генетическом уровне путем обратного фосфорилирования.
Регуляция активности ферментов
Гормональная регуляция осуществляется на генетическом уровне путем обратного фосфорилирования.

Гормон, циркулирующий в крови, попадает в межклеточную жидкость и контактирует с
Гормон, циркулирующий в крови, попадает в межклеточную жидкость и контактирует с

Уровень, образовавшийся под действием АЦ, цАМФ определяется не только активностью АЦ,
Уровень, образовавшийся под действием АЦ, цАМФ определяется не только активностью АЦ,

цГМФ оказывает эффекты противоположные цАМФ.
цАМФ находится в тесном контакте с ионами
цГМФ оказывает эффекты противоположные цАМФ.
цАМФ находится в тесном контакте с ионами

Изоферменты, их природа, биологическая роль, строение ЛДГ.
Изоферменты - это группа родственных
Изоферменты, их природа, биологическая роль, строение ЛДГ.
Изоферменты - это группа родственных

1) скорстью катализа;
2) направлением катализируемой реакции;
3) условиями протекания реакции;
4) чувствительностью к регуляторам, факторам среды. (Более
1) скорстью катализа;
2) направлением катализируемой реакции;
3) условиями протекания реакции;
4) чувствительностью к регуляторам, факторам среды. (Более

Изоферменты имеют адаптивное значение, т. е. придают специфику метаболизма.
Изоферменты обеспечивают межорганную
Изоферменты имеют адаптивное значение, т. е. придают специфику метаболизма.
Изоферменты обеспечивают межорганную

ЛДГ4,5
в печени: ПВК -----> лактат
ЛДГ1,2
в сердце: лактат ------> ПВК
ЛДГ - олигомерный
ЛДГ4,5
в печени: ПВК -----> лактат
ЛДГ1,2
в сердце: лактат ------> ПВК
ЛДГ - олигомерный

Существует 5 изоферментных форм:
HHHH HHHM HHMM HMMM MMMM
H4 H3M H2M2 HM3 M4
ЛДГ1,
Существует 5 изоферментных форм:
HHHH HHHM HHMM HMMM MMMM
H4 H3M H2M2 HM3 M4
ЛДГ1,

Остальные изоферменты занимают промежуточное положение.
Изоферменты ЛДГ локализованы в различных тканях:
ЛДГ1,2 ---->
Остальные изоферменты занимают промежуточное положение.
Изоферменты ЛДГ локализованы в различных тканях:
ЛДГ1,2 ---->

Изменение активности ферментов в онтогенезе.
Онтогенез человека развивается по определенной генетической
Изменение активности ферментов в онтогенезе.
Онтогенез человека развивается по определенной генетической

Глюкоза утилизируется анаэробным путем (анаэробный гликолиз), т. к. ткани плода
Глюкоза утилизируется анаэробным путем (анаэробный гликолиз), т. к. ткани плода

3. Грудной
Потребность в глюкозе резко возрастает, она начинает утилизироваться аэробно,
3. Грудной
Потребность в глюкозе резко возрастает, она начинает утилизироваться аэробно,
 Масленица в картинах русских художников
Масленица в картинах русских художников Этническая общность, русские
Этническая общность, русские Производственный контур систем управления предприятием
Производственный контур систем управления предприятием Презентация на тему "Научно-исследовательская деятельность как средство саморазвития обучающихся" - скачать презентации по
Презентация на тему "Научно-исследовательская деятельность как средство саморазвития обучающихся" - скачать презентации по  Введение в CSS
Введение в CSS Электрические цепи постоянного тока
Электрические цепи постоянного тока Урок -игра Путешествие в страну Информашка 5 класс
Урок -игра Путешествие в страну Информашка 5 класс  Мини-футбол
Мини-футбол Паттерны проектирования: Шаблонный метод
Паттерны проектирования: Шаблонный метод Сельские праздники
Сельские праздники ОСЛОЖНЕНИЯ ОСЛОЖНЕНИЯ ИНФАРКТА МИОКАРДА
ОСЛОЖНЕНИЯ ОСЛОЖНЕНИЯ ИНФАРКТА МИОКАРДА Горнолыжные трассы
Горнолыжные трассы Языки программирования Fortran, Lisp, Cobol
Языки программирования Fortran, Lisp, Cobol Искусство изразцов. 5 класс
Искусство изразцов. 5 класс Механизм рынка несовершенной конкуренции
Механизм рынка несовершенной конкуренции «В начале было Слово...» Библейские сказания
«В начале было Слово...» Библейские сказания Презентация на тему "Здоровые зубы - здоровое тело." - скачать презентации по Медицине
Презентация на тему "Здоровые зубы - здоровое тело." - скачать презентации по Медицине Урок 8. Тела, вещества, частицы.
Урок 8. Тела, вещества, частицы. Схема цепей приведения в движение тепловоза ЧМЭ3
Схема цепей приведения в движение тепловоза ЧМЭ3 Общество с ограниченной ответственностью «Горстройсеть»
Общество с ограниченной ответственностью «Горстройсеть» Режимы роботы и конструкция асинхронных машин
Режимы роботы и конструкция асинхронных машин История изобретения радио
История изобретения радио Программирование на языке Python
Программирование на языке Python ПРАВОСУДДЯ В УКРАЇНІ
ПРАВОСУДДЯ В УКРАЇНІ Презентация на тему "Я, ты, он, она, мы - здоровая семья!" - скачать презентации по Медицине
Презентация на тему "Я, ты, он, она, мы - здоровая семья!" - скачать презентации по Медицине Детали и механизмы машин. Паровая машина
Детали и механизмы машин. Паровая машина Меры по демократизации и общественной жизни во второй половине 1950-х –начале 1960-х
Меры по демократизации и общественной жизни во второй половине 1950-х –начале 1960-х  Причины и последствия мировых войн XX века
Причины и последствия мировых войн XX века