- Ферменты

Содержание
- 2. Содержание: 1.Механизм регуляции активности ферментов. Аллостерическая регуляция. 2.Характеристика аллостерических ферментов. Виды ингибирования( обратимое, необратимое, конкурентное, неконкурентное,
- 4. При знакомстве с материалом по кинетике ферментов могло сложиться впечатление, что кинетика всех ферментов основана на
- 5. У ферментов, подчиняющимся принципам кинетики Михаэлиса и Ментен с позиций процессов, протекающих в клетке, имеется ряд
- 6. Поэтому природа вынуждена обратиться к "кооперативным" системам, в которых маленькие изменения в одном параметре, например концентрации
- 7. Графически результат работы такой кооперативной системы (график зависимости скорости реакции от концентрации субстрата) выражается не гиперболой,
- 9. Аллостерический белок определяется как белок, содержащий два или больше топологически различающихся центра связывания лигандов (субстраты, ингибиторы
- 10. Кооперативность - это модификация константы связывания (Кs)лиганда, белком, предшествующим связыванию другого лиганда. Константы связывания - подобны
- 11. Км обычно принимается как константа связывания субстрата, поскольку ее проще измерять, чем Ks. Понятие кооперативность означает,
- 13. Значения Vmax и Km определяются при экстраполяции линий до пересечения с осью абсцисс и ординат. Как
- 14. Два лиганда один из которых влияет на связывание другого, могут быть химически идентичны, например, одна молекула
- 15. У аллостерических ферментов особые свойства Полимерная структура Сигмоидная ( в отличие от гиперболической для не аллостерических)
- 17. Центральная линия графика - типичная сигмовидная кривая в отсутствии любого эффектора. В присутствии активатора (А) скорость
- 18. Ингибитор увеличил сигмовидную форму, удлиняя «носок» кривой, в то время как активатор оказывал противоположный эффект. При
- 19. Денатурация – нарушение пространственной структуры фермента с последующей потерей активности фермента. Денатурация вызывается рядом факторов, включая
- 20. Инггибиторы бывают обратимые и необратимые Вещества со свойствами ингибиторов ферментов можно грубо разделить на обратимые и
- 21. Следствием такого связывания обратимого ингибитора является эффективное мгновенное действие, но после удаления ингибитора фермент сохраняет свою
- 22. Необратимые ингибиторы известны также как инактиваторы фермента. Они связываются с ферментом, формируя прочные, обычно ковалентные связи:
- 23. Среди примеров необратимо действующих ингибиторов можно назвать диизопропилфторфосфат (ДИПФФ). Это соединение вошло в историю энзимологии как
- 24. ДИПФФ ковалентно связывается с гидроксильной группой серина и если эта группа важна в катализе реакции, фермент
- 25. Ацетилсалициловая кислота (известный всем аспирин) является необратимым ингибитором циклооксигеназы- фермента участвующего в синтезе простагландинов.
- 26. Ингибитором синтеза протеогликанов стенки бактерий является пенилиллин, структура которого напоминает D-аланин, встраиваемый в структуру протеогликанов. Связываясь
- 27. Различают два механизма конкурентного торможения. 1.Конкурентное торможение путем связывания активного центра. 2. Конкурентное торможение путем изменения
- 28. Если связывание субстрата с АЦ фермента предшествует изменению конформации центра связывания ингибитора, то последний не связывает
- 29. И субстрат и ингибитор не могут одновременно связаться с ферментом. В этом виде конкурентного торможения ингибитор
- 30. Конкурентные ингибиторы не влияют на Vmax, они понижают Км. Оказывая эффект на: Km, тем самым оказывают
- 31. Малоновая кислота тормозит активность дегидрогеназы янтарной кислоты, занимая активный центр на ферменте. Учитывая обратимость реакции, избыток
- 32. Конкурентное торможение путем изменения конформации фермента. В отличие от классического варианта, ингибитор связывается не с активным
- 33. Регуляция активности ферментов путем химической модификации: Реакции ограниченного протеолиза, аденилирования, рибозилирования, ацетилирования, фосфорилирования (роль гормонов, АЦ-комплекса,
- 34. Активность фермента можно изменить путем ковалентной модификации его структуры. Ковалентная модификация структуры ферментов может быть обратимой
- 35. Гормональная регуляция активности ферментов Осуществляется при: -функционировании АЦ комплекса, - с помощью вторичных посредников- цАМФ, цГМФ,
- 36. Регуляция активности ферментов Гормональная регуляция осуществляется на генетическом уровне путем обратного фосфорилирования, и осуществляется при: -функционировании
- 37. Аденилатциклазный комплекс В ходе этого процесса образуется небелковое соединение - цАМФ, цАМФ - внутриклеточный гормон (вторичный
- 38. В ходе этого процесса образуется небелковое соединение - цАМФ, цАМФ - внутриклеточный гормон (вторичный посредник) яв-ся
- 39. См. мультимедийную презентацию-Сигнальная трансдукция.
- 40. Гормон, циркулирующий в крови, попадает в межклеточную жидкость и контактирует с поверхностью клетки, где расположены рецепторы
- 41. Уровень, образовавшийся под действием АЦ, цАМФ определяется не только активностью АЦ, но и активностью фосфодиэстераз, которые
- 42. Наибольшее значение имеет цГМФ. Она образуется под действием гуанилатциклазы, расположенной как в наружной мембране, так и
- 43. цГМФ оказывает эффекты противоположные цАМФ. цАМФ находится в тесном контакте с ионами Ca2+: высокая концентрация цАМФ
- 44. Аденилатциклазный механизм
- 45. Некоторые клеточные ответы на действие гормонов, опосредуемое циклическим АМФ
- 46. ИФ3 обеспечивает повышение уровня кальция
- 48. Сигнальные роли ДАГ
- 49. Если ИФ3 повышает концентрацию ионов Са2+ в цитозоле, другой продукт расщепления фосфатидилинозитол -4,5- дифосфата – ДАГ
- 50. Фермент, активируемый ДАГ, называется протеинкиназой С (ПКС) или С–киназой, так как активность его зависит от уровня
- 51. Связывание диацилглицерола изменяет конформацию протеинкиназы С, что сопровождается повышением ее сродства к ионам Са2+ и липидам.
- 52. С-киназа, активированная диацилглицеролом и Са2+, переносит концевую фосфатную группу с АТФ на специфические сериновые или треониновые
- 53. Концентрация С-киназы выше всего в нейронах головного мозга, где, помимо, прочего она фосфорилирует ионные каналы нейронов
- 54. Взаимосвязь цАМФ и ИФ3 зависимых путей
- 55. Изоферменты, их природа, биологическая роль, строение ЛДГ. Изоферменты - это группа родственных ферментов, катализирующих одну и
- 56. Изоферменты
- 57. 1) скоростью катализа; 2) направлением катализируемой реакции; 3) условиями протекания реакции; 4) чувствительностью к регуляторам, факторам
- 58. Тетрамерный фермент, состоящий из двух кодируемых генетически субъединиц А и В может быть представлен пятью изоферментами
- 59. Изоферменты имеют адаптивное значение, т. е. придают специфику метаболизма. Изоферменты обеспечивают межорганную связь, например, в процессе
- 60. ЛДГ4, и 5 в печени: ПВК -----> лактат ЛДГ1, и 2 в сердце: лактат ------> ПВК
- 61. Существует 5 изоферментных форм: HHHH HHHM HHMM HMMM MMMM H4 H3M H2M2 HM3 M4 ЛДГ1, ЛДГ2,
- 62. Остальные изоферменты занимают промежуточное положение. Изоферменты ЛДГ локализованы в различных тканях: ЛДГ1,2 ----> мозг, аэробные ткани
- 63. Изменение активности ферментов в онтогенезе. Онтогенез человека развивается по определенной генетической программе, которая записана на уровне
- 64. Глюкоза утилизируется анаэробным путем (анаэробный гликолиз), т. к. ткани плода не способны к ГНГ, и идет
- 65. 3. Грудной Потребность в глюкозе резко возрастает, она начинает утилизироваться аэробно, но примерно до двух лет
- 66. 6. Зрелый. Происходит стабилизация массы тела, репродуктивного гомеостаза. После 35-40 лет основным источником энергии являются опять
- 67. Активная регуляция ферментов возможна - на геномном уровне ( лактозный оперон); Регуляция конститутивными и индуцибельными ферментами.
- 68. Роль ПКС в регуляции транскрипции
- 69. Известны два внутриклеточных пути, через которые активированная С-киназа усиливает транскрипцию генов. В одном, С-киназа активирует протеинкиназный
- 70. Elk-1 связан с короткой последовательностью ДНК (обозначаемой serum response element – SRE) и ассоциирован с другим
- 71. В другом пути, активация С-киназы приводит к фосфорилированию ингибиторного белка Ik-B, что сопровождается высвобождением из комплекса
- 72. Существует группа соединений, среди которых наиболее хорошо изучены форболовые эфиры, которые являются мощными активаторами ПКС. Они
- 73. Метаболизм ферментов Превращение ферментов сходно с обменом других белков. Определенный уровень ферментов в тканях поддерживается их
- 75. Имея высокую молекулярную массу, большая часть ферментов не экскретируется с мочой. Ферменты экскретируемые с желчью -ЩФ,
- 76. Активность фермента Активность - это изменение количества субстрата под влиянием фермента в единицу времени. Под изменением
- 77. Понятие "активность фермента" по сути дела идентична понятию "скорость ферментативной" реакции. Ферментативная активность выражается в единицах
- 78. В системе СИ в качестве единицы ферментативной активности используют "катал" (kat). Катал определяется как 1 моль/сек.
- 79. Размерность её слишком велика, на практике пользуются меньшими кратными значениями, начиная с нанокатала (нкат). Это одна
- 80. В практике лабораторий широко пользуются понятием удельная активность. Для этого число cтандартных единиц пересчитывают на какую-либо
- 81. При исследовании кинетики реакций используется и такое понятие как молекулярная активность. Она показывает, сколько молекул субстрата
- 82. Пример для вычисления активности фермента Исходные данные:25.0 x 10-3 моль л--1 пептида-субстрата, объем реакционной смеси 2.5
- 83. Изменение активности ферментов у больного ИМ в разные сроки заболевания
- 84. Номенклатура и классификация ферментов Каждый фермент получает специфический кодовый номер-шифр фермента, отражающий его положение в классификации:
- 85. Шифр КФК и ее место в классификации ферментов
- 86. Каждый подподкласс представляет собой список ферментов. Порядковый номер фермента в этом списке – четвертая цифра кода.
- 87. Оксидоредуктазы Оксидоредуктазы (1.0.0.0.) 1.1.0.0. Действуют на СН-ОН группы доноров 1.1.1.0. НАД+ или НАДФ+ в качестве акцепторов
- 88. Трансферазы Трансферазы (2.0.0.0.) 2.1.0.0.Переносят одноуглеродные группы 2.1.1.0. Метилтрансферазы 2.1.1.1. Никотинамид метилтрансфераза 2.1.1.45. Тимидилат синтаза 2.3.0.0. Ацилтрансферазы
- 89. Гидролазы (3.0.0.0.) 3.1.0.0.Действуют на эфирные связи 3.1.1.0.Гидролазы эфиров карбоновых кислот 3.1.1.17. Ацетилхолинэстераза 3.2.1.0. Гликозидгидролазы 3.2.1.1. a-амилаза
- 90. Лиазы(4.0.0.0) 4.1.0.0.Углерод-углерод лиазы 4.1.1.0.Карбокси лиазы 4.1.1.1. Пируватдекарбоксилаза 4.2.0.0. Углерод-кислород-лиазы 4.2.1.0. Гидролиазы 4.2.1.11. Енолаза 4.2.1.12. Фосфоглюконатдегидраза
- 91. Лигазы (6.0.0.0) 6.1.0.0. Образуют С-О связи 6.1.1.0.Образуют молекулы аминоацил-тРНК и родственные им соединения. 6.1.1.1. Тирозил-тРНК синтаза
- 93. Скачать презентацию

Содержание:
1.Механизм регуляции активности ферментов. Аллостерическая регуляция.
2.Характеристика аллостерических ферментов. Виды ингибирования(
Содержание:
1.Механизм регуляции активности ферментов. Аллостерическая регуляция.
2.Характеристика аллостерических ферментов. Виды ингибирования(


При знакомстве с материалом по кинетике ферментов могло сложиться впечатление, что кинетика
При знакомстве с материалом по кинетике ферментов могло сложиться впечатление, что кинетика

У ферментов, подчиняющимся принципам кинетики Михаэлиса и Ментен с позиций процессов,
У ферментов, подчиняющимся принципам кинетики Михаэлиса и Ментен с позиций процессов,

Поэтому природа вынуждена обратиться к "кооперативным" системам, в которых маленькие изменения
Поэтому природа вынуждена обратиться к "кооперативным" системам, в которых маленькие изменения

Графически результат работы такой кооперативной системы (график зависимости скорости реакции от
Графически результат работы такой кооперативной системы (график зависимости скорости реакции от


Аллостерический белок определяется как белок, содержащий два или больше топологически различающихся
Аллостерический белок определяется как белок, содержащий два или больше топологически различающихся

Кооперативность - это модификация константы связывания (Кs)лиганда, белком, предшествующим связыванию другого
Кооперативность - это модификация константы связывания (Кs)лиганда, белком, предшествующим связыванию другого

Км обычно принимается как константа связывания субстрата, поскольку ее проще
Км обычно принимается как константа связывания субстрата, поскольку ее проще


Значения Vmax и Km определяются при экстраполяции линий до пересечения с
Значения Vmax и Km определяются при экстраполяции линий до пересечения с

Два лиганда один из которых влияет на связывание другого, могут быть
Два лиганда один из которых влияет на связывание другого, могут быть

У аллостерических ферментов особые свойства
Полимерная структура
Сигмоидная ( в отличие от гиперболической
У аллостерических ферментов особые свойства
Полимерная структура
Сигмоидная ( в отличие от гиперболической


Центральная линия графика - типичная сигмовидная кривая в отсутствии любого эффектора.
Центральная линия графика - типичная сигмовидная кривая в отсутствии любого эффектора.

Ингибитор увеличил сигмовидную форму, удлиняя «носок» кривой, в то время как
Ингибитор увеличил сигмовидную форму, удлиняя «носок» кривой, в то время как

Денатурация – нарушение пространственной структуры фермента с последующей потерей активности фермента.
Денатурация – нарушение пространственной структуры фермента с последующей потерей активности фермента.

Инггибиторы бывают обратимые и необратимые
Вещества со свойствами ингибиторов ферментов можно грубо
Инггибиторы бывают обратимые и необратимые
Вещества со свойствами ингибиторов ферментов можно грубо

Следствием такого связывания обратимого ингибитора является эффективное мгновенное действие, но после
Следствием такого связывания обратимого ингибитора является эффективное мгновенное действие, но после

Необратимые ингибиторы известны также как инактиваторы фермента. Они связываются с
Необратимые ингибиторы известны также как инактиваторы фермента. Они связываются с

Среди примеров необратимо действующих ингибиторов можно назвать диизопропилфторфосфат (ДИПФФ). Это соединение
Среди примеров необратимо действующих ингибиторов можно назвать диизопропилфторфосфат (ДИПФФ). Это соединение

ДИПФФ ковалентно связывается с гидроксильной группой серина и если эта
ДИПФФ ковалентно связывается с гидроксильной группой серина и если эта

Ацетилсалициловая кислота (известный всем аспирин) является необратимым ингибитором циклооксигеназы- фермента
Ацетилсалициловая кислота (известный всем аспирин) является необратимым ингибитором циклооксигеназы- фермента

Ингибитором синтеза протеогликанов стенки бактерий является пенилиллин, структура которого напоминает
Ингибитором синтеза протеогликанов стенки бактерий является пенилиллин, структура которого напоминает

Различают два механизма конкурентного торможения.
1.Конкурентное торможение путем связывания активного центра.
2.
Различают два механизма конкурентного торможения.
1.Конкурентное торможение путем связывания активного центра.
2.

Если связывание субстрата с АЦ фермента предшествует изменению конформации центра
Если связывание субстрата с АЦ фермента предшествует изменению конформации центра

И субстрат и ингибитор не могут одновременно связаться с ферментом.
И субстрат и ингибитор не могут одновременно связаться с ферментом.

Конкурентные ингибиторы не влияют на Vmax, они понижают Км.
Оказывая эффект
Конкурентные ингибиторы не влияют на Vmax, они понижают Км.
Оказывая эффект

Малоновая кислота тормозит активность дегидрогеназы янтарной кислоты, занимая активный центр
Малоновая кислота тормозит активность дегидрогеназы янтарной кислоты, занимая активный центр

Конкурентное торможение путем изменения конформации фермента.
В отличие от классического
Конкурентное торможение путем изменения конформации фермента.
В отличие от классического

Регуляция активности ферментов путем химической модификации: Реакции ограниченного протеолиза, аденилирования, рибозилирования,
Регуляция активности ферментов путем химической модификации: Реакции ограниченного протеолиза, аденилирования, рибозилирования,

Активность фермента можно изменить путем ковалентной модификации его структуры.
Ковалентная
Активность фермента можно изменить путем ковалентной модификации его структуры.
Ковалентная

Гормональная регуляция активности ферментов
Осуществляется при:
-функционировании АЦ комплекса,
- с помощью вторичных
Гормональная регуляция активности ферментов
Осуществляется при:
-функционировании АЦ комплекса,
- с помощью вторичных

Регуляция активности ферментов
Гормональная регуляция осуществляется на генетическом уровне путем обратного
Регуляция активности ферментов
Гормональная регуляция осуществляется на генетическом уровне путем обратного

Аденилатциклазный комплекс
В ходе этого процесса образуется небелковое соединение - цАМФ, цАМФ
Аденилатциклазный комплекс
В ходе этого процесса образуется небелковое соединение - цАМФ, цАМФ

В ходе этого процесса образуется небелковое соединение - цАМФ, цАМФ
В ходе этого процесса образуется небелковое соединение - цАМФ, цАМФ

См. мультимедийную презентацию-Сигнальная трансдукция.
См. мультимедийную презентацию-Сигнальная трансдукция.

Гормон, циркулирующий в крови, попадает в межклеточную жидкость и контактирует
Гормон, циркулирующий в крови, попадает в межклеточную жидкость и контактирует

Уровень, образовавшийся под действием АЦ, цАМФ определяется не только активностью
Уровень, образовавшийся под действием АЦ, цАМФ определяется не только активностью

Наибольшее значение имеет цГМФ. Она образуется под действием гуанилатциклазы, расположенной
Наибольшее значение имеет цГМФ. Она образуется под действием гуанилатциклазы, расположенной

цГМФ оказывает эффекты противоположные цАМФ.
цАМФ находится в тесном контакте
цГМФ оказывает эффекты противоположные цАМФ.
цАМФ находится в тесном контакте
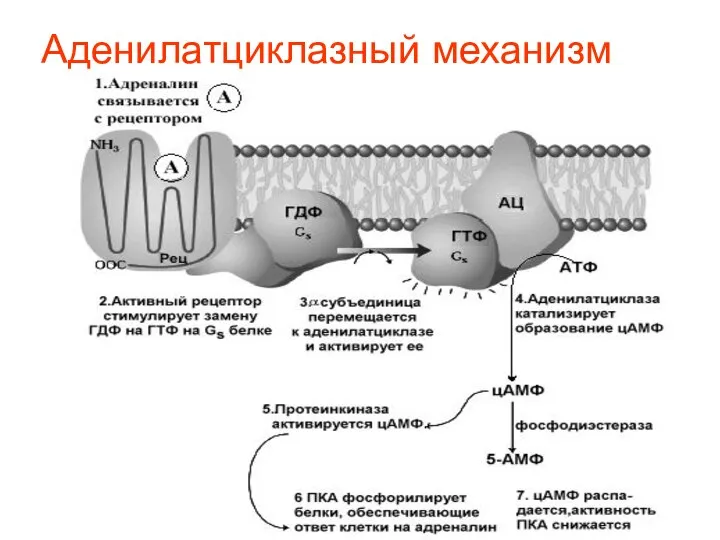
Аденилатциклазный механизм
Аденилатциклазный механизм

Некоторые клеточные ответы на действие гормонов,
опосредуемое циклическим АМФ
Некоторые клеточные ответы на действие гормонов,
опосредуемое циклическим АМФ
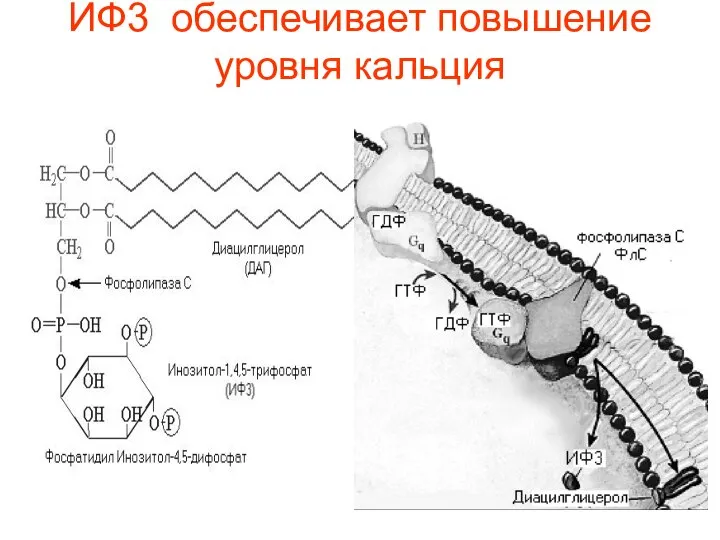
ИФ3 обеспечивает повышение уровня кальция
ИФ3 обеспечивает повышение уровня кальция


Сигнальные роли ДАГ
Сигнальные роли ДАГ

Если ИФ3 повышает концентрацию ионов Са2+ в цитозоле, другой продукт
Если ИФ3 повышает концентрацию ионов Са2+ в цитозоле, другой продукт

Фермент, активируемый ДАГ, называется протеинкиназой С (ПКС) или С–киназой, так как
Фермент, активируемый ДАГ, называется протеинкиназой С (ПКС) или С–киназой, так как

Связывание диацилглицерола изменяет конформацию протеинкиназы С, что сопровождается повышением ее сродства
Связывание диацилглицерола изменяет конформацию протеинкиназы С, что сопровождается повышением ее сродства

С-киназа, активированная диацилглицеролом и Са2+, переносит концевую фосфатную группу с АТФ
С-киназа, активированная диацилглицеролом и Са2+, переносит концевую фосфатную группу с АТФ

Концентрация С-киназы выше всего в нейронах головного мозга, где, помимо, прочего
Концентрация С-киназы выше всего в нейронах головного мозга, где, помимо, прочего
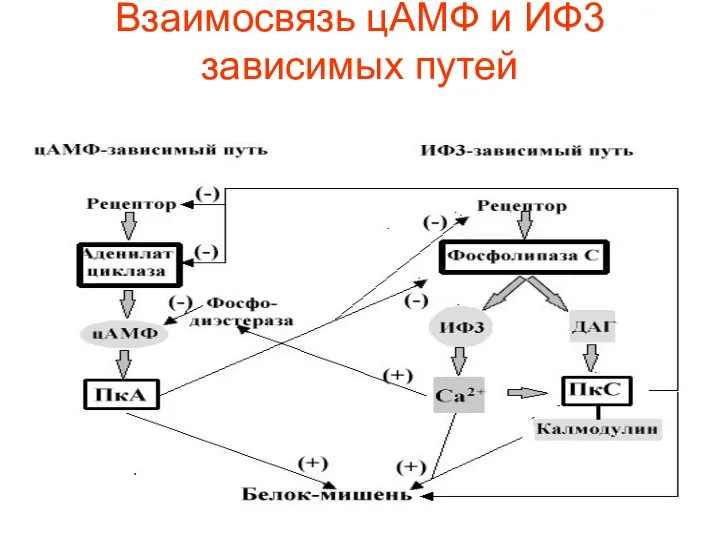
Взаимосвязь цАМФ и ИФ3 зависимых путей
Взаимосвязь цАМФ и ИФ3 зависимых путей

Изоферменты, их природа, биологическая роль, строение ЛДГ.
Изоферменты - это группа родственных
Изоферменты, их природа, биологическая роль, строение ЛДГ.
Изоферменты - это группа родственных
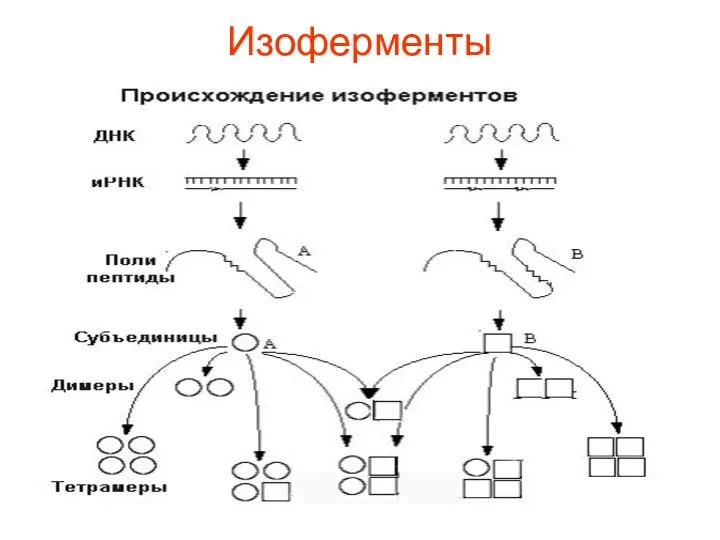
Изоферменты
Изоферменты

1) скоростью катализа;
2) направлением катализируемой реакции;
3) условиями протекания реакции;
4) чувствительностью к
1) скоростью катализа;
2) направлением катализируемой реакции;
3) условиями протекания реакции;
4) чувствительностью к

Тетрамерный фермент, состоящий из двух кодируемых генетически субъединиц А и В
Тетрамерный фермент, состоящий из двух кодируемых генетически субъединиц А и В

Изоферменты имеют адаптивное значение, т. е. придают специфику метаболизма.
Изоферменты
Изоферменты имеют адаптивное значение, т. е. придают специфику метаболизма.
Изоферменты

ЛДГ4, и 5
в печени: ПВК -----> лактат
ЛДГ1, и
ЛДГ4, и 5
в печени: ПВК -----> лактат
ЛДГ1, и

Существует 5 изоферментных форм:
HHHH HHHM HHMM HMMM MMMM
H4
Существует 5 изоферментных форм:
HHHH HHHM HHMM HMMM MMMM
H4

Остальные изоферменты занимают промежуточное положение.
Изоферменты ЛДГ локализованы в различных
Остальные изоферменты занимают промежуточное положение.
Изоферменты ЛДГ локализованы в различных

Изменение активности ферментов в онтогенезе.
Онтогенез человека развивается по определенной генетической
Изменение активности ферментов в онтогенезе.
Онтогенез человека развивается по определенной генетической

Глюкоза утилизируется анаэробным путем (анаэробный гликолиз), т. к. ткани плода
Глюкоза утилизируется анаэробным путем (анаэробный гликолиз), т. к. ткани плода

3. Грудной
Потребность в глюкозе резко возрастает, она начинает утилизироваться аэробно,
3. Грудной
Потребность в глюкозе резко возрастает, она начинает утилизироваться аэробно,

6. Зрелый.
Происходит стабилизация массы тела, репродуктивного гомеостаза. После 35-40 лет
6. Зрелый.
Происходит стабилизация массы тела, репродуктивного гомеостаза. После 35-40 лет

Активная регуляция ферментов возможна - на геномном уровне ( лактозный
Активная регуляция ферментов возможна - на геномном уровне ( лактозный
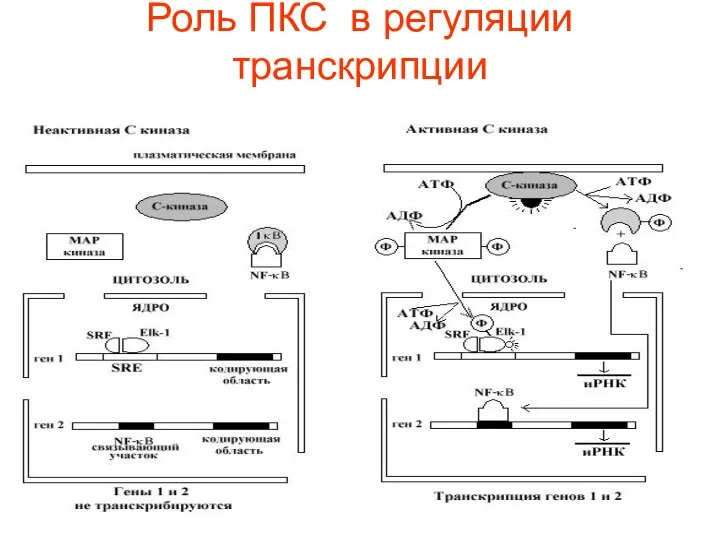
Роль ПКС в регуляции транскрипции
Роль ПКС в регуляции транскрипции

Известны два внутриклеточных пути, через которые активированная С-киназа усиливает транскрипцию
Известны два внутриклеточных пути, через которые активированная С-киназа усиливает транскрипцию

Elk-1 связан с короткой последовательностью ДНК (обозначаемой serum response element
Elk-1 связан с короткой последовательностью ДНК (обозначаемой serum response element

В другом пути, активация С-киназы приводит к фосфорилированию ингибиторного белка
В другом пути, активация С-киназы приводит к фосфорилированию ингибиторного белка

Существует группа соединений, среди которых наиболее хорошо изучены форболовые эфиры,
Существует группа соединений, среди которых наиболее хорошо изучены форболовые эфиры,

Метаболизм ферментов
Превращение ферментов сходно с обменом других белков. Определенный уровень ферментов
Метаболизм ферментов
Превращение ферментов сходно с обменом других белков. Определенный уровень ферментов


Имея высокую молекулярную массу, большая часть ферментов не экскретируется с
Имея высокую молекулярную массу, большая часть ферментов не экскретируется с

Активность фермента
Активность - это изменение количества субстрата под влиянием фермента
Активность фермента
Активность - это изменение количества субстрата под влиянием фермента

Понятие "активность фермента" по сути дела идентична понятию "скорость ферментативной"
Понятие "активность фермента" по сути дела идентична понятию "скорость ферментативной"

В системе СИ в качестве единицы ферментативной активности используют
В системе СИ в качестве единицы ферментативной активности используют

Размерность её слишком велика, на практике пользуются меньшими кратными значениями,
Размерность её слишком велика, на практике пользуются меньшими кратными значениями,

В практике лабораторий широко пользуются понятием удельная активность. Для этого
В практике лабораторий широко пользуются понятием удельная активность. Для этого

При исследовании кинетики реакций используется и такое понятие как молекулярная
При исследовании кинетики реакций используется и такое понятие как молекулярная

Пример для вычисления активности фермента
Исходные данные:25.0 x 10-3 моль л--1
Пример для вычисления активности фермента
Исходные данные:25.0 x 10-3 моль л--1

Изменение активности ферментов у больного ИМ в разные сроки заболевания
Изменение активности ферментов у больного ИМ в разные сроки заболевания

Номенклатура и классификация ферментов
Каждый фермент получает специфический кодовый номер-шифр фермента, отражающий
Номенклатура и классификация ферментов
Каждый фермент получает специфический кодовый номер-шифр фермента, отражающий
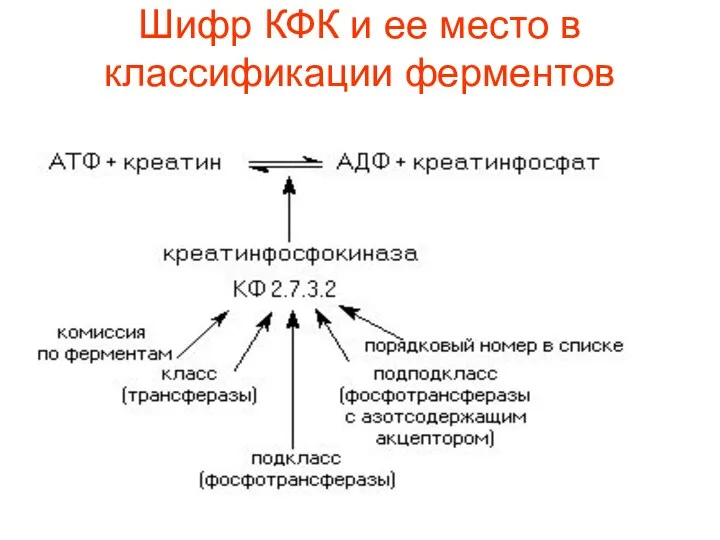
Шифр КФК и ее место в классификации ферментов
Шифр КФК и ее место в классификации ферментов

Каждый подподкласс представляет собой список ферментов. Порядковый номер фермента в
Каждый подподкласс представляет собой список ферментов. Порядковый номер фермента в

Оксидоредуктазы
Оксидоредуктазы (1.0.0.0.)
1.1.0.0. Действуют на СН-ОН группы доноров
1.1.1.0. НАД+
Оксидоредуктазы
Оксидоредуктазы (1.0.0.0.)
1.1.0.0. Действуют на СН-ОН группы доноров
1.1.1.0. НАД+

Трансферазы
Трансферазы (2.0.0.0.)
2.1.0.0.Переносят одноуглеродные
группы
2.1.1.0. Метилтрансферазы
2.1.1.1. Никотинамид метилтрансфераза
2.1.1.45. Тимидилат
Трансферазы
Трансферазы (2.0.0.0.)
2.1.0.0.Переносят одноуглеродные
группы
2.1.1.0. Метилтрансферазы
2.1.1.1. Никотинамид метилтрансфераза
2.1.1.45. Тимидилат

Гидролазы (3.0.0.0.)
3.1.0.0.Действуют на эфирные связи 3.1.1.0.Гидролазы эфиров карбоновых кислот
Гидролазы (3.0.0.0.)
3.1.0.0.Действуют на эфирные связи 3.1.1.0.Гидролазы эфиров карбоновых кислот
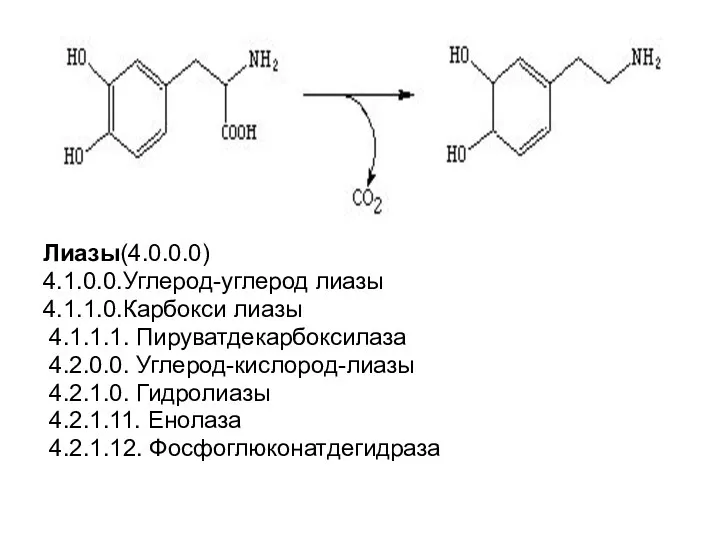
Лиазы(4.0.0.0)
4.1.0.0.Углерод-углерод лиазы
4.1.1.0.Карбокси лиазы
4.1.1.1. Пируватдекарбоксилаза
4.2.0.0. Углерод-кислород-лиазы
4.2.1.0. Гидролиазы
4.2.1.11. Енолаза
Лиазы(4.0.0.0)
4.1.0.0.Углерод-углерод лиазы
4.1.1.0.Карбокси лиазы
4.1.1.1. Пируватдекарбоксилаза
4.2.0.0. Углерод-кислород-лиазы
4.2.1.0. Гидролиазы
4.2.1.11. Енолаза

Лигазы (6.0.0.0)
6.1.0.0. Образуют С-О связи
6.1.1.0.Образуют молекулы аминоацил-тРНК
Лигазы (6.0.0.0)
6.1.0.0. Образуют С-О связи
6.1.1.0.Образуют молекулы аминоацил-тРНК
 Архитектура персональных компьютеров
Архитектура персональных компьютеров Қазақ тілі мен әдебиет
Қазақ тілі мен әдебиет  Знакомься_другие части слов
Знакомься_другие части слов Тракт генерирования и излучения зондирующего сигнала ПРВ. Особенности построения передающего устройства изделия 1РЛ130. (ПРВ-13)
Тракт генерирования и излучения зондирующего сигнала ПРВ. Особенности построения передающего устройства изделия 1РЛ130. (ПРВ-13) Судовой газотурбинный двигатель
Судовой газотурбинный двигатель Политическая элита и политическое лидерство
Политическая элита и политическое лидерство Распорядительные документы
Распорядительные документы Современное комплексное решение для образовательных учреждений от компании GST-Партнер
Современное комплексное решение для образовательных учреждений от компании GST-Партнер Статья 185.6. Неправомерное использование инсайдерской информации _
Статья 185.6. Неправомерное использование инсайдерской информации _ Физминутка Мурляндия Составила учитель начальных классов МАОУ Ильинской СОШ Домодедовского р-на Московской обл Белозёрова Т.В
Физминутка Мурляндия Составила учитель начальных классов МАОУ Ильинской СОШ Домодедовского р-на Московской обл Белозёрова Т.В Свободное движение тела
Свободное движение тела МЕЖДУНАРОДНАЯ ТОРГОВЛЯ
МЕЖДУНАРОДНАЯ ТОРГОВЛЯ  Презентация Сергей Викторович Лавров
Презентация Сергей Викторович Лавров  MS Project 2007 Краткое введение в инструментарий
MS Project 2007 Краткое введение в инструментарий Основы web-технологий. Технологии создания web-сайтов
Основы web-технологий. Технологии создания web-сайтов Скоростные способности и основы методики их развития
Скоростные способности и основы методики их развития Види і функції сучасних медіа
Види і функції сучасних медіа Русский пейзаж в поэзии и живописи Альбом 6 «Б» класса Презентация Субботина Георгия Школа №1173, Москва 2007 год
Русский пейзаж в поэзии и живописи Альбом 6 «Б» класса Презентация Субботина Георгия Школа №1173, Москва 2007 год Использование ЦОР Общие вопросы
Использование ЦОР Общие вопросы Антонио Гауди - гениальный архитектор, великий модернист
Антонио Гауди - гениальный архитектор, великий модернист Правовая защита государственной и частной компьютеризированной информации в РФ
Правовая защита государственной и частной компьютеризированной информации в РФ Предписания органов муниципального контроля
Предписания органов муниципального контроля Системы пожарной сигнализации
Системы пожарной сигнализации Классный час в УКПСИЗО Учитель Афанасьева Н.Н.
Классный час в УКПСИЗО Учитель Афанасьева Н.Н. Защита от электромагнитных излучений
Защита от электромагнитных излучений Globalization : good or bad? Victoria Yurlova
Globalization : good or bad? Victoria Yurlova Java. (Лекция 4)
Java. (Лекция 4) Religion. How does religion effect the modern life?
Religion. How does religion effect the modern life?