- Физиологические свойства нервных проводников

Содержание
- 2. План лекции 1. Физиологические свойства нервных проводников. Типы нервных волокон. 2. Механизмы и особенности проведения возбуждения
- 3. ФИЗИОЛОГИЯ НЕРВОВ ИЛИ НЕРВНЫХ ПРОВОДНИКОВ Нервное волокно (нервный проводник) представляет собой отросток нейрона(аксон), заключенный в глиальную
- 4. КЛАССИФИКАЦИЯ НЕРВНЫХ ВОЛОКОН ПО ЭЛАНГЕРУ-ГАССЕРУ
- 5. Законы проведения возбуждения в нервных волокнах • 1. Закон двустороннего проведения. • 2. Закон анатомической и
- 6. Закон двустороннего проведения • Возбуждение, возникающее в одном участке нерва, распространяется в обе стороны от места
- 7. Закон анатомической и физиологической целостности • Возбуждение может распространяться по нервному волокну только в случае его
- 8. Закон изолированного проведения • Возбуждение, распространяющееся по волокну, входящему в состав нерва, не передается на соседние
- 9. Закон бездекрементного проведения • Амплитуда потенциала действия не изменяется с увеличением расстояния от места его возникновения.
- 13. Механизм мышечного сокращения Электромеханическое сопряжение. • Распространение ПД по сарколемме в Т- трубочке приводит к активации
- 15. Механизм мышечного сокращения Скольжение нитей {сокращение саркомера). • АТФаза миозиновой головки вызывает гидролиз АТФ до АДФ
- 17. Механизм мышечного сокращения Расслабление миофибрилл. Для расслабления миофибрилл необходимы 2 главных условия: наличие достаточного уровня АТФ
- 21. Скачать презентацию

План лекции
1. Физиологические свойства нервных проводников. Типы нервных волокон.
2. Механизмы и
План лекции
1. Физиологические свойства нервных проводников. Типы нервных волокон.
2. Механизмы и
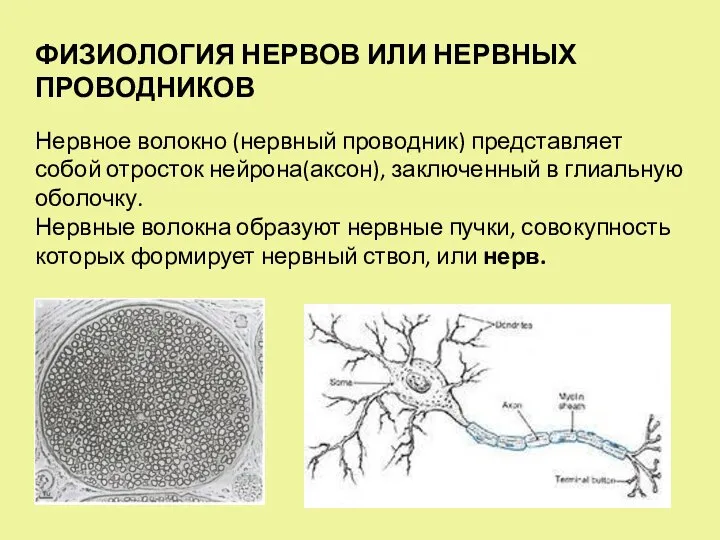
ФИЗИОЛОГИЯ НЕРВОВ ИЛИ НЕРВНЫХ ПРОВОДНИКОВ
Нервное волокно (нервный проводник) представляет собой отросток
ФИЗИОЛОГИЯ НЕРВОВ ИЛИ НЕРВНЫХ ПРОВОДНИКОВ
Нервное волокно (нервный проводник) представляет собой отросток
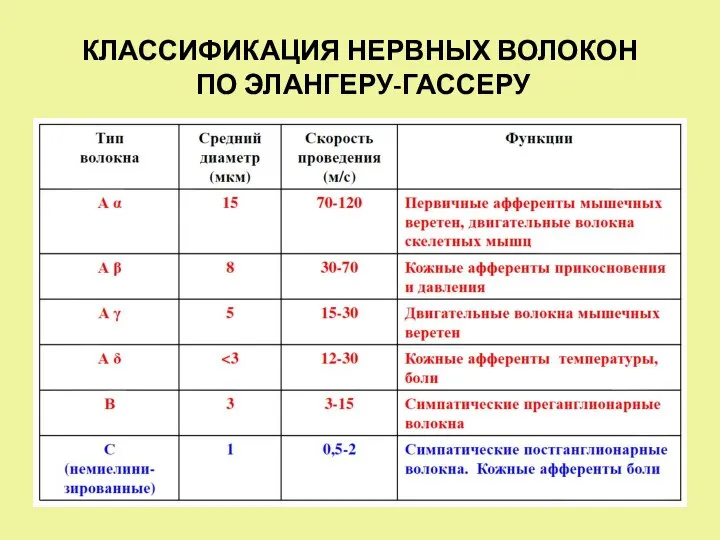
КЛАССИФИКАЦИЯ НЕРВНЫХ ВОЛОКОН
ПО ЭЛАНГЕРУ-ГАССЕРУ
КЛАССИФИКАЦИЯ НЕРВНЫХ ВОЛОКОН
ПО ЭЛАНГЕРУ-ГАССЕРУ

Законы проведения возбуждения в
нервных волокнах
• 1. Закон двустороннего проведения.
• 2. Закон
Законы проведения возбуждения в
нервных волокнах
• 1. Закон двустороннего проведения.
• 2. Закон

Закон двустороннего проведения
• Возбуждение, возникающее в одном участке нерва, распространяется в
Закон двустороннего проведения
• Возбуждение, возникающее в одном участке нерва, распространяется в

Закон анатомической и физиологической целостности
• Возбуждение может распространяться по нервному волокну
Закон анатомической и физиологической целостности
• Возбуждение может распространяться по нервному волокну

Закон изолированного проведения
• Возбуждение, распространяющееся по волокну,
входящему в состав нерва, не
Закон изолированного проведения
• Возбуждение, распространяющееся по волокну,
входящему в состав нерва, не

Закон бездекрементного проведения
• Амплитуда потенциала действия не
изменяется с увеличением расстояния
от места
Закон бездекрементного проведения
• Амплитуда потенциала действия не
изменяется с увеличением расстояния
от места
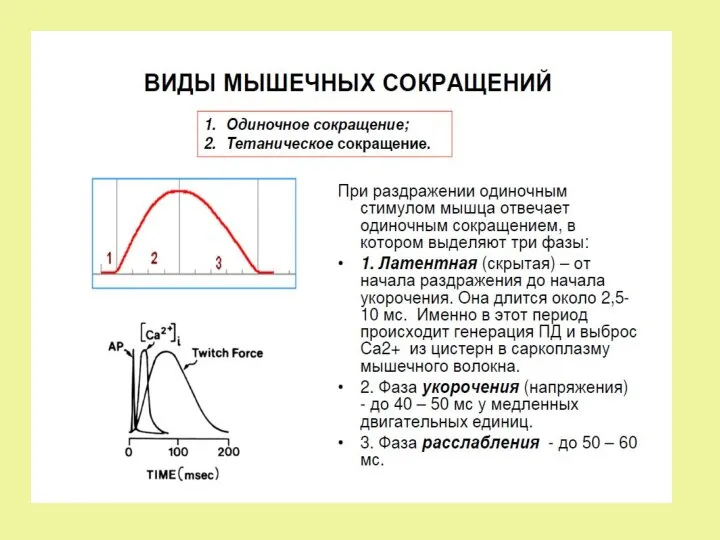

Механизм мышечного сокращения
Электромеханическое сопряжение.
• Распространение ПД по сарколемме в Т- трубочке
Механизм мышечного сокращения
Электромеханическое сопряжение.
• Распространение ПД по сарколемме в Т- трубочке
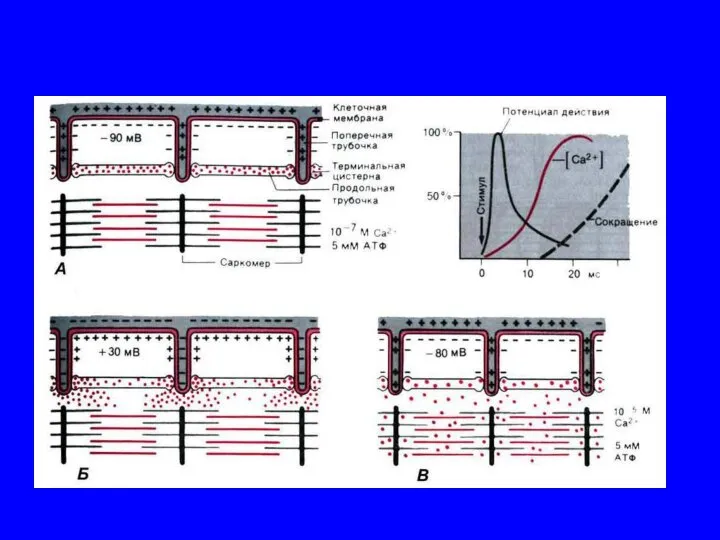

Механизм мышечного сокращения
Скольжение нитей {сокращение саркомера).
• АТФаза миозиновой головки вызывает гидролиз
Механизм мышечного сокращения
Скольжение нитей {сокращение саркомера).
• АТФаза миозиновой головки вызывает гидролиз
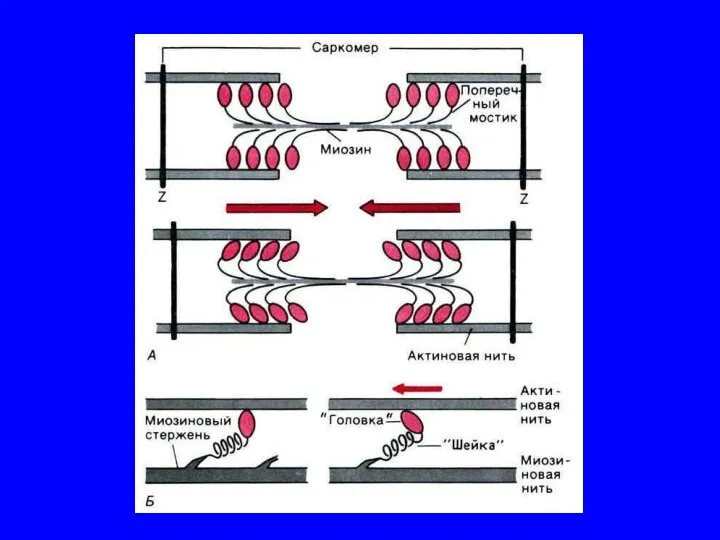

Механизм мышечного сокращения
Расслабление миофибрилл.
Для расслабления миофибрилл необходимы 2 главных условия:
Механизм мышечного сокращения
Расслабление миофибрилл.
Для расслабления миофибрилл необходимы 2 главных условия:

 Комплекс упражнений "Полет к звезде" - презентация для начальной школы_
Комплекс упражнений "Полет к звезде" - презентация для начальной школы_ Анализ рисков для устойчивого развития жилищного строительства и их количественная оценка
Анализ рисков для устойчивого развития жилищного строительства и их количественная оценка Освоение подземного пространства как одно из мероприятий экореконструкции
Освоение подземного пространства как одно из мероприятий экореконструкции Основы теории государства и права. Лекция 1
Основы теории государства и права. Лекция 1 Узлы и веревки
Узлы и веревки Интерференция света
Интерференция света Что такое погода - презентация для начальной школы_
Что такое погода - презентация для начальной школы_ Презентация "Как глобальные информационные системы, подобные Интернет способствуют развитию логистики" - скачать презентаци
Презентация "Как глобальные информационные системы, подобные Интернет способствуют развитию логистики" - скачать презентаци Формальная постановка комбинаторно-оптимизационных задач. (Тема 3)
Формальная постановка комбинаторно-оптимизационных задач. (Тема 3) Классы и объекты
Классы и объекты "Чипсы - Вредно или нет?" - презентация для начальной школы_
"Чипсы - Вредно или нет?" - презентация для начальной школы_ Прецеденты. Вопросы, возникающие при определении исполнителей и задач. (Лекция 3)
Прецеденты. Вопросы, возникающие при определении исполнителей и задач. (Лекция 3) 2. БАРЬЕР И ТРАМПЛИН ИССЛЕДОВАНИЯ. Определение проблемы как начало исследования Проблема – это белое пятно на карте науки, знание
2. БАРЬЕР И ТРАМПЛИН ИССЛЕДОВАНИЯ. Определение проблемы как начало исследования Проблема – это белое пятно на карте науки, знание  Технология проектной деятельности
Технология проектной деятельности История лампы накаливания
История лампы накаливания Задачи с использованием одномерных массивов
Задачи с использованием одномерных массивов Процесс моделирования
Процесс моделирования ПОНЯТИЕ «МЕТОДА», «МЕТОДИКИ» И «МЕТОДОЛОГИИ» НАУЧНОГО ПОЗНАНИЯ. ИХ ВЗАИМОСВЯЗЬ. Подготовила студентка 1 курса ФТД группы Т-1207 Пон
ПОНЯТИЕ «МЕТОДА», «МЕТОДИКИ» И «МЕТОДОЛОГИИ» НАУЧНОГО ПОЗНАНИЯ. ИХ ВЗАИМОСВЯЗЬ. Подготовила студентка 1 курса ФТД группы Т-1207 Пон Electronics
Electronics Индийский океан
Индийский океан «Наурызым-мерекем, Наурызым-берекем» (әдеби-музыкалық композиция)
«Наурызым-мерекем, Наурызым-берекем» (әдеби-музыкалық композиция) Организация ремонта радиоэлектронной техники (РЭТ)
Организация ремонта радиоэлектронной техники (РЭТ) Юри на льду
Юри на льду Suomen kielen viidestoista 15. oppitunti
Suomen kielen viidestoista 15. oppitunti Лентивирусы
Лентивирусы Транзисторы и их применение
Транзисторы и их применение Клавиатура. Типы клавиатур
Клавиатура. Типы клавиатур Характеристика группы 80-83 Выполнила Карпенко Татьяна Т-114
Характеристика группы 80-83 Выполнила Карпенко Татьяна Т-114