- ФИЗИОЛОГИЯ СЕНСОРНЫХ СИСТЕМ

Содержание
- 2. Анализатор (сенсорная система) – часть нервной системы, включающая 1. Рецепторный аппарат (периферический отдел), 2. Нервные клетки
- 3. Функции сенсорных систем Обнаружение сигналов Различение сигналов Кодирование информации Передача и преобразование сигналов Осознание образов
- 4. Структура отражения Явления внешнего мира Сенсорные стимулы Сенсорные рецепторы Афферентные нервные волокна Сенсорные центры Допсихическое отражение
- 11. ОСНОВНЫЕ ХАРАКТЕРИСТИКИ (РАЗМЕРНОСТИ) ОЩУЩЕНИЯ Пространство (местоположение, пространственная протяженность); Время (определение начала и продолжительности ощущения); Модальность (зрение,
- 14. ОБЩИЕ ПРИНЦИПЫ СТРОЕНИЯ АНАЛИЗАТОРОВ 1. Многослойность . 2. Многоканальность . специфический канал ассоциативный канал неспецифический канал.
- 15. Многослойность Наличие нескольких слоев нервных клеток, начиная с рецепторов и заканчивая корковыми нейронами. Между собой слои
- 16. специфический канал неспецифический канал ассоциативный канал 1 – чувствительные нейроны одной сенсорной системы, 2 – чувствительный
- 17. Специфический канал Осуществляет передачу сигналов определенной модальности и оценивает физические или химические параметры раздражителя. Импульсация распространяется
- 18. В строении специфических путей можно выделить следующие особенности: Каналы имеют малое количество переключений, т.е. специфический путь
- 19. Рецептивное поле – это совокупность рецепторов, импульсы от которых поступают к данному нейрону. Проекционное поле –
- 20. Специфический канал Соматотопическая организация корковой зоны SI человека Изображения на поперечном срезе мозга и их обозначения
- 21. Корковые поля по Бродману
- 22. Ассоциативный канал Нейроны этого канала имеют высокую степень конвергенции, Функция - обеспечивает интегрированную межсенсорную афферентацию в
- 23. Ассоциативные зоны обеспечивают процессы межсенсорной интеграции и играют большую роль в формировании полисенсорного образа и оценке
- 24. Неспецифический канал Формируется в результате дивергенции специфических нервных волокон на различных уровнях ЦНС (нейроны стволовой и
- 25. Значение неспецифической системы мозга: Поддерживает общий уровень возбудимости мозговых структур, участвует в их активации, т.е. быстрой
- 26. Сенсорная система не просто пассивный канал связей. Она включает и аппарат управления. Каждый уровень системы работает
- 27. 3. Многоуровневость рецепторный; спинальный; стволовой; таламический; кортикальный. В каждой СС выделяют следующие обязательные уровни переключений и
- 28. 4. Принцип парного строения Любая сенсорная система построена по принципу двухсторонней симметрии. Однако этот принцип достаточно
- 29. Основной механизм парной деятельности сенсорных систем – механизм функциональной асимметрии при действии различным образом локализованных в
- 30. Левое полушарие дискретное, обрабатывает информацию по принципу индукции – от частного к общему. Правое полушарие –
- 31. СЕНСОРНЫЕ РЕЦЕПТОРЫ
- 32. Клеточная и сенсорная рецепция Клеточная рецепция - процесс восприятия и преобразования химического сигнала в сложную последовательность
- 33. Классификации рецепторов Психофизиологическая классификация Рецепторы разделяются 1. по модальности ощущений, возникающих при их раздражении - слуховые,
- 34. Первичные и вторичные рецепторы 1 – тело чувствительного нейрона; 2 – периферический отросток чувствительного нейрона (дендрит);
- 35. Первичночувствующие рецепторы – это нервные окончания концевые участки дендрита чувствительного нейрона
- 36. Вторичночувствующие рецепторы Состоят из 2-х синаптически связанных элементов: клетки эпителиального либо нервного происхождения (волосковые клетки улитки)
- 37. СВОЙСТВА И ФУНКЦИИ РЕЦЕПТОРОВ
- 38. Свойства рецепторов 1. Модальная специфичность рецепторов - это способность рецепторов одного типа при взаимодействии с вышележащими
- 39. 2. Высокая чувствительность к адекватному раздражителю - это способность рецепторов возбуждаться при действии слабых по интенсивности
- 40. 3. Высокая специализация рецепторов одной модальности – Способность рецепторов реагировать не на весь диапазон адекватных стимулов,
- 41. 4. Способность к адаптации Адаптация сенсорных рецепторов – это процесс уменьшения или прекращения их активности по
- 42. Адаптация тонических ( А ), промежуточных ( Б ) и фазных рецепторов ( В ) к
- 43. Функции рецепторов Обнаружение; Кодирование; Различение.
- 44. ОБНАРУЖЕНИЕ СИГНАЛОВ - это трансформация (преобразование) физической и химической энергии раздражителя в импульсную активность (ПД). Этапы:
- 45. Закономерности преобразования энергии внешнего раздражителя в серию нервных импульсов: чем выше сила действующего раздражителя, тем больше
- 46. КОДИРОВАНИЕ ПАРАМЕТРОВ РАЗДРАЖИТЕЛЯ Кодирование – это процесс преобразования какой-либо информации в условный код – упорядоченный набор
- 47. Временное кодирование Временное кодирование – информация кодируется в виде определенной пачки ПД. Информационное значение имеет серия
- 48. Пространственное кодирование Пространственное кодирование – информация кодируется количеством и определенной топографией нервных волокон, приносящих возбуждение в
- 49. КОДИРОВАНИЕ ПАРАМЕТРОВ СЕНСОРНЫХ ОЩУЩЕНИЙ
- 50. Кодировании модальности и качества 1. Кодирование меченными линиями (пространственный принцип); 2. Кодирование определенным паттерном возбуждения
- 51. Кодирование интенсивности Более сильное раздражение вызывает более сильное ощущение. Осуществляется 2 способами: Частотой импульсов в отдельных
- 52. Кодирование места, формы и размеров Происходит по пространственному принципу. Данный способ кодирования хорошо выражен в зрительной
- 53. РАЗЛИЧЕНИЕ СИГНАЛОВ
- 54. Различение интенсивности Различение интенсивности стимулов происходит в соответствии с законом Вебера-Фехнера. Рецепторы могут различить по интенсивности
- 55. Вебер: Минимальное различимое изменение интенсивности стимуляции (∆φ) составляет постоянную долю (с) ее исходной интенсивности (φ): ∆φ=с∙φ
- 56. Фехнер предложил использовать дифференциальные пороги для измерения субъективных ощущений и построения шкалы интенсивности ощущений (ψ). Психофизический
- 57. Различение пространственных параметров Пространственное различение стимулов возможно в том случае, если между возбужденными при их действии
- 58. Различение временных параметров Для временного различения раздражителей важно, чтобы они не сливались во времени, т.е. второй
- 59. Функции сенсорных систем Обнаружение сигналов Различение сигналов Кодирование информации Передача и преобразование сигналов Осознание образов
- 60. 4. Передача и преобразование сигналов
- 61. Преобразования сигналов могут быть разделены на: Пространственные (усиление сигнала) – изменение соотношения разных частей сигнала за
- 62. Пространственные (усиление сигнала) преобразования сигналов
- 63. «Сенсорный гомункулюс» В зрительной и соматосенсорных системах на корковом уровне значительно искажаются геометрические пропорции представительства отдельных
- 65. 1981- теория колончатой организации коры (в 60-х годах прошлого века V.Mountcastle) Сенсорная кора построена из функциональных
- 66. Рецептивные поля интернейронов Рецептивным полем нейрона называют множество рецепторов, функционально связанных с этим нейроном. Рецептивное поле
- 67. Перекрытие рецептивных полей. Взаимодействие рецепторов в рецептивном поле. За счет конвергенции и дивергенции сенсорная информация передается
- 68. Подавление информации о менее существенных сигналах происходит за счет возвратного и латерального торможения. Например, на сетчатку
- 69. В ходе преобразования сигналов происходит: Ограничение избыточности информации и выделение существенных признаков сигнала. Подавление информации о
- 70. Детектирование сигналов Это избирательное выделение сенсорным нейроном признака раздражителя, имеющего поведенческое значение. Такой анализ осуществляют нейроны-детекторы,
- 72. Зрительные корковые нейроны с простым рецептивным полем активируются при воздействии на фоторецепторы световым стимулом в виде
- 73. Для активации нейронов со сложным рецептивным полем необходимы не только оформленный и пространственно-ориентированный стимул, но и
- 74. Иллюзия Мюллера-Лайера также объясняется организацией сложных рецептивных полей корковых нейронов, возбуждение которых определяется углом между двумя
- 75. 5. Опознание образов
- 76. Это конечная и наиболее сложная функция сенсорной системы. Она заключается в отнесении образа к определенному классу
- 77. ВЗАИМОДЕЙСТВИЕ СЕНСОРНЫХ СИСТЕМ Взаимодействие сенсорных систем осуществляется на спинальном, ретикулярном, таламическом и корковом уровнях. Особенно широка
- 79. Скачать презентацию

Анализатор (сенсорная система) – часть нервной системы, включающая
1. Рецепторный аппарат (периферический
Анализатор (сенсорная система) – часть нервной системы, включающая
1. Рецепторный аппарат (периферический

Функции сенсорных систем
Обнаружение сигналов
Различение сигналов
Кодирование информации
Передача и преобразование сигналов
Осознание образов
Функции сенсорных систем
Обнаружение сигналов
Различение сигналов
Кодирование информации
Передача и преобразование сигналов
Осознание образов
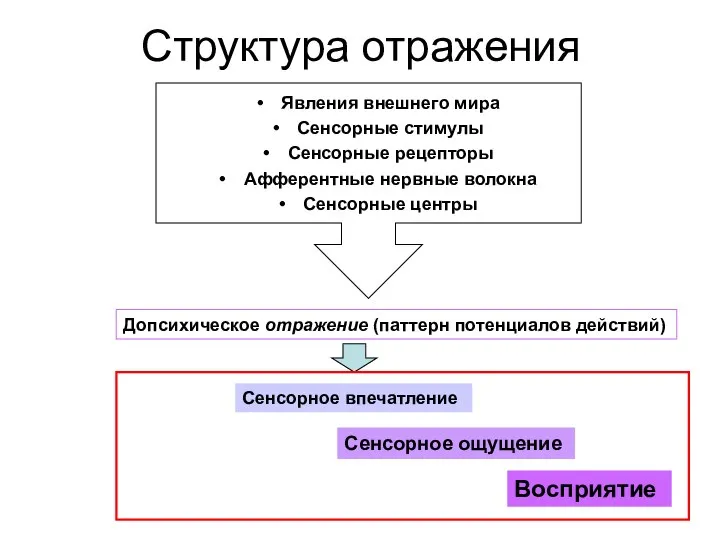
Структура отражения
Явления внешнего мира
Сенсорные стимулы
Сенсорные рецепторы
Афферентные нервные волокна
Сенсорные центры
Структура отражения
Явления внешнего мира
Сенсорные стимулы
Сенсорные рецепторы
Афферентные нервные волокна
Сенсорные центры






ОСНОВНЫЕ ХАРАКТЕРИСТИКИ (РАЗМЕРНОСТИ) ОЩУЩЕНИЯ
Пространство (местоположение, пространственная протяженность);
Время (определение начала и продолжительности
ОСНОВНЫЕ ХАРАКТЕРИСТИКИ (РАЗМЕРНОСТИ) ОЩУЩЕНИЯ
Пространство (местоположение, пространственная протяженность);
Время (определение начала и продолжительности

ОБЩИЕ ПРИНЦИПЫ СТРОЕНИЯ АНАЛИЗАТОРОВ
1. Многослойность .
2. Многоканальность .
специфический
ОБЩИЕ ПРИНЦИПЫ СТРОЕНИЯ АНАЛИЗАТОРОВ
1. Многослойность .
2. Многоканальность .
специфический

Многослойность
Наличие нескольких слоев нервных клеток, начиная с рецепторов и заканчивая корковыми
Многослойность
Наличие нескольких слоев нервных клеток, начиная с рецепторов и заканчивая корковыми
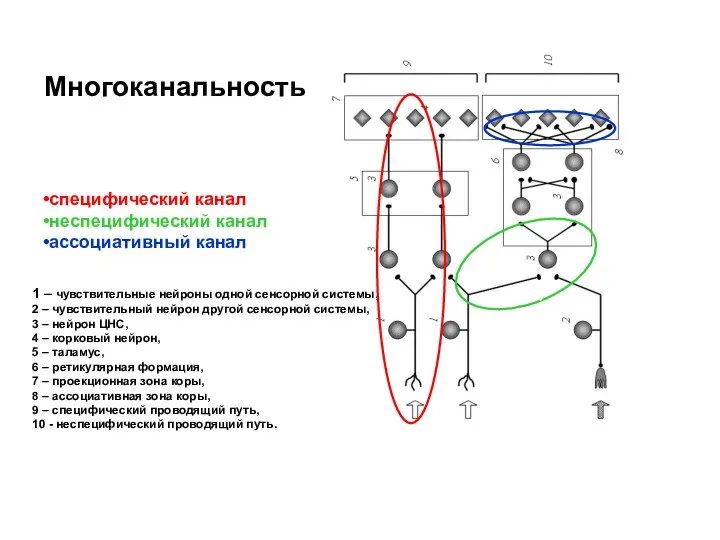
специфический канал
неспецифический канал
ассоциативный канал
1 – чувствительные нейроны одной сенсорной системы,
2 –
специфический канал
неспецифический канал
ассоциативный канал
1 – чувствительные нейроны одной сенсорной системы,
2 –

Специфический канал
Осуществляет передачу сигналов определенной модальности и оценивает физические или химические
Специфический канал
Осуществляет передачу сигналов определенной модальности и оценивает физические или химические

В строении специфических путей можно выделить следующие особенности:
Каналы имеют малое
В строении специфических путей можно выделить следующие особенности:
Каналы имеют малое

Рецептивное поле – это совокупность рецепторов, импульсы от которых поступают к
Рецептивное поле – это совокупность рецепторов, импульсы от которых поступают к
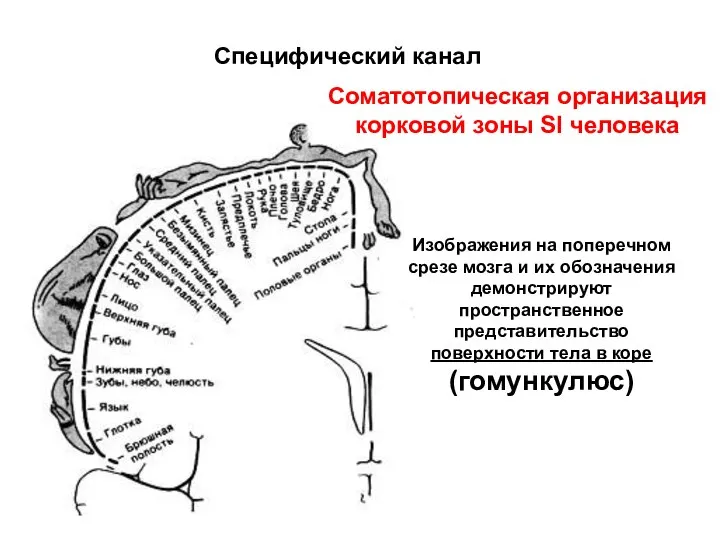
Специфический канал
Соматотопическая организация
корковой зоны SI человека
Изображения на поперечном срезе
Специфический канал
Соматотопическая организация
корковой зоны SI человека
Изображения на поперечном срезе
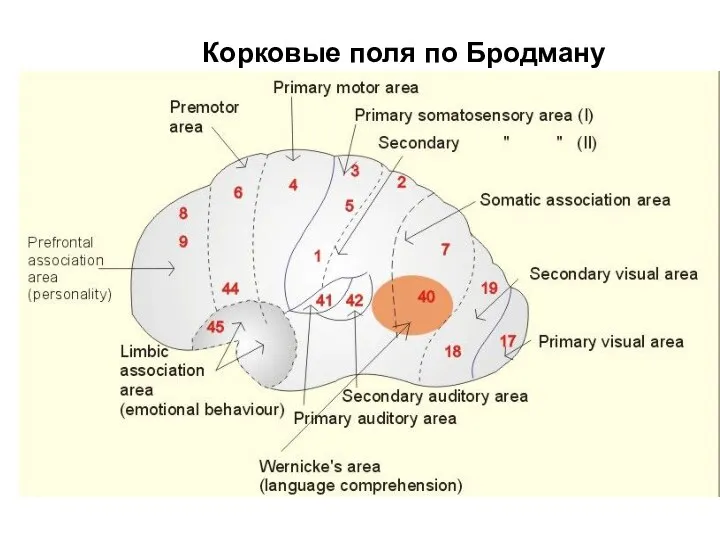
Корковые поля по Бродману
Корковые поля по Бродману

Ассоциативный канал
Нейроны этого канала имеют высокую степень конвергенции,
Функция - обеспечивает
Ассоциативный канал
Нейроны этого канала имеют высокую степень конвергенции,
Функция - обеспечивает

Ассоциативные зоны обеспечивают процессы межсенсорной интеграции и играют большую роль в
Ассоциативные зоны обеспечивают процессы межсенсорной интеграции и играют большую роль в

Неспецифический канал
Формируется в результате дивергенции специфических нервных волокон на различных уровнях
Неспецифический канал
Формируется в результате дивергенции специфических нервных волокон на различных уровнях

Значение неспецифической системы мозга:
Поддерживает общий уровень возбудимости мозговых структур, участвует в
Значение неспецифической системы мозга:
Поддерживает общий уровень возбудимости мозговых структур, участвует в

Сенсорная система не просто пассивный канал связей.
Она включает и аппарат
Сенсорная система не просто пассивный канал связей.
Она включает и аппарат
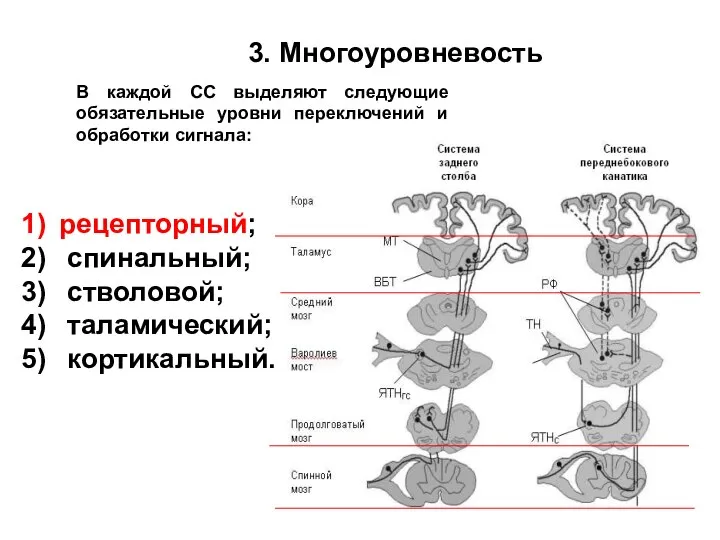
3. Многоуровневость
рецепторный;
спинальный;
стволовой;
таламический;
кортикальный.
В каждой
3. Многоуровневость
рецепторный;
спинальный;
стволовой;
таламический;
кортикальный.
В каждой

4. Принцип парного строения
Любая сенсорная система построена по принципу двухсторонней
4. Принцип парного строения
Любая сенсорная система построена по принципу двухсторонней

Основной механизм парной деятельности сенсорных систем – механизм функциональной асимметрии при
Основной механизм парной деятельности сенсорных систем – механизм функциональной асимметрии при

Левое полушарие дискретное, обрабатывает информацию по принципу индукции – от частного
Левое полушарие дискретное, обрабатывает информацию по принципу индукции – от частного

СЕНСОРНЫЕ
РЕЦЕПТОРЫ
СЕНСОРНЫЕ
РЕЦЕПТОРЫ

Клеточная и сенсорная рецепция
Клеточная рецепция - процесс восприятия и преобразования химического
Клеточная и сенсорная рецепция
Клеточная рецепция - процесс восприятия и преобразования химического

Классификации рецепторов
Психофизиологическая классификация
Рецепторы разделяются
1. по модальности ощущений, возникающих при их раздражении
Классификации рецепторов
Психофизиологическая классификация
Рецепторы разделяются
1. по модальности ощущений, возникающих при их раздражении
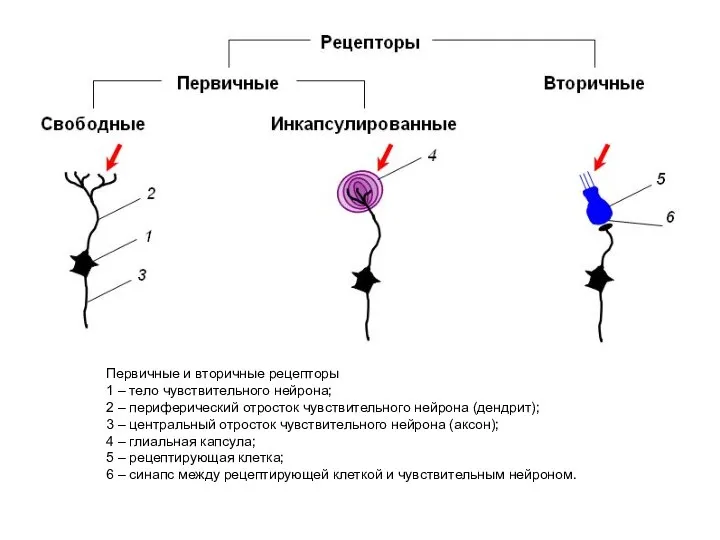
Первичные и вторичные рецепторы
1 – тело чувствительного нейрона;
2 – периферический
Первичные и вторичные рецепторы
1 – тело чувствительного нейрона;
2 – периферический
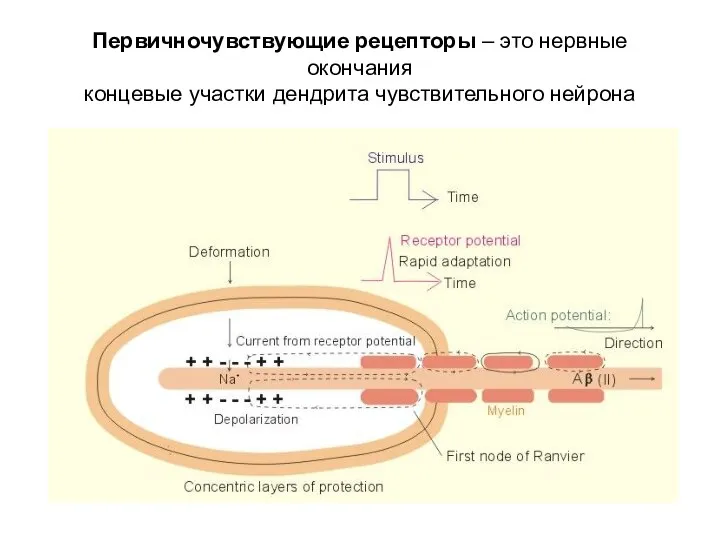
Первичночувствующие рецепторы – это нервные окончания
концевые участки дендрита чувствительного нейрона
Первичночувствующие рецепторы – это нервные окончания
концевые участки дендрита чувствительного нейрона

Вторичночувствующие рецепторы
Состоят из 2-х синаптически связанных элементов:
клетки эпителиального либо нервного происхождения
Вторичночувствующие рецепторы
Состоят из 2-х синаптически связанных элементов:
клетки эпителиального либо нервного происхождения

СВОЙСТВА И ФУНКЦИИ РЕЦЕПТОРОВ
СВОЙСТВА И ФУНКЦИИ РЕЦЕПТОРОВ

Свойства рецепторов
1. Модальная специфичность рецепторов - это способность рецепторов одного типа
Свойства рецепторов
1. Модальная специфичность рецепторов - это способность рецепторов одного типа

2. Высокая чувствительность к адекватному раздражителю - это способность рецепторов возбуждаться
2. Высокая чувствительность к адекватному раздражителю - это способность рецепторов возбуждаться
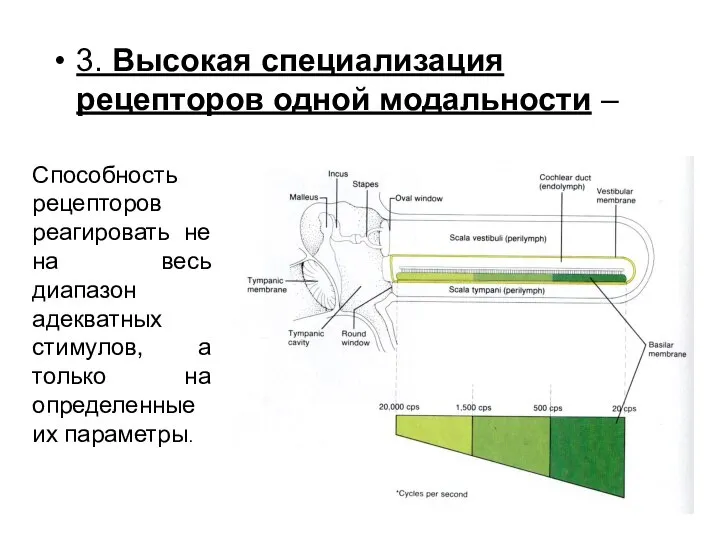
3. Высокая специализация рецепторов одной модальности –
Способность рецепторов реагировать не
3. Высокая специализация рецепторов одной модальности –
Способность рецепторов реагировать не

4. Способность к адаптации
Адаптация сенсорных рецепторов – это процесс уменьшения или
4. Способность к адаптации
Адаптация сенсорных рецепторов – это процесс уменьшения или
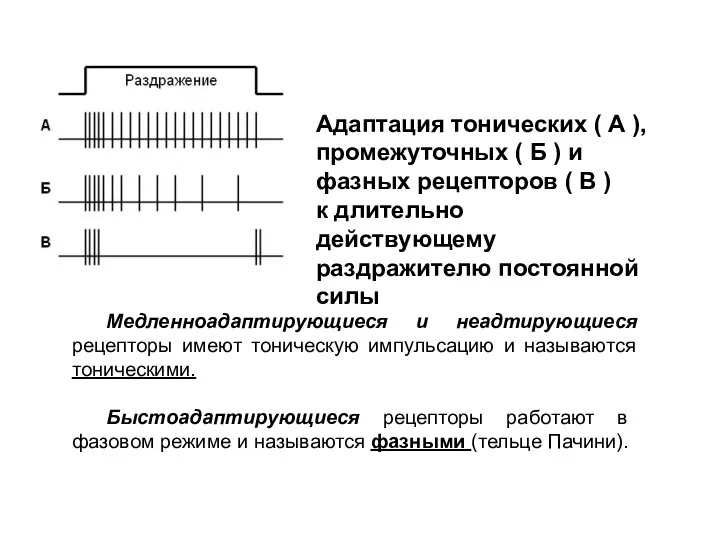
Адаптация тонических ( А ), промежуточных ( Б ) и
фазных
Адаптация тонических ( А ), промежуточных ( Б ) и
фазных

Функции рецепторов
Обнаружение;
Кодирование;
Различение.
Функции рецепторов
Обнаружение;
Кодирование;
Различение.

ОБНАРУЖЕНИЕ СИГНАЛОВ
- это трансформация (преобразование) физической и химической энергии раздражителя в
ОБНАРУЖЕНИЕ СИГНАЛОВ
- это трансформация (преобразование) физической и химической энергии раздражителя в
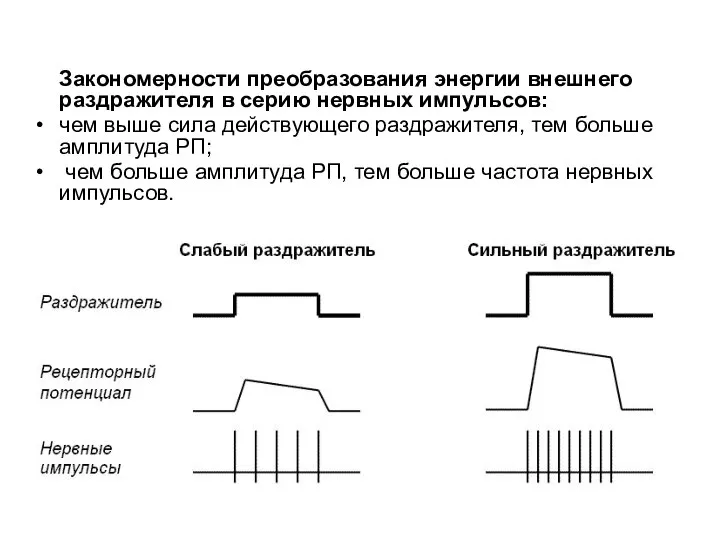
Закономерности преобразования энергии внешнего раздражителя в серию нервных импульсов:
чем выше сила
Закономерности преобразования энергии внешнего раздражителя в серию нервных импульсов:
чем выше сила

КОДИРОВАНИЕ ПАРАМЕТРОВ РАЗДРАЖИТЕЛЯ
Кодирование – это процесс преобразования какой-либо информации в условный
КОДИРОВАНИЕ ПАРАМЕТРОВ РАЗДРАЖИТЕЛЯ
Кодирование – это процесс преобразования какой-либо информации в условный

Временное кодирование
Временное кодирование – информация кодируется в виде определенной пачки ПД.
Временное кодирование
Временное кодирование – информация кодируется в виде определенной пачки ПД.

Пространственное кодирование
Пространственное кодирование – информация кодируется количеством и определенной топографией нервных
Пространственное кодирование
Пространственное кодирование – информация кодируется количеством и определенной топографией нервных

КОДИРОВАНИЕ ПАРАМЕТРОВ СЕНСОРНЫХ ОЩУЩЕНИЙ
КОДИРОВАНИЕ ПАРАМЕТРОВ СЕНСОРНЫХ ОЩУЩЕНИЙ
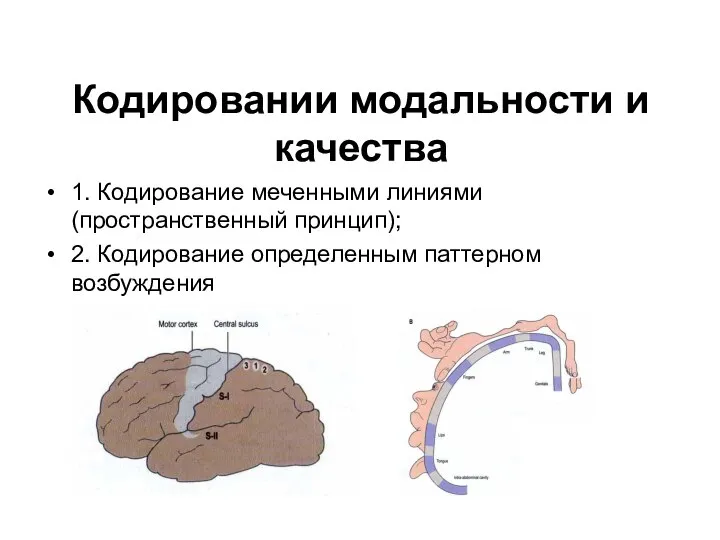
Кодировании модальности и качества
1. Кодирование меченными линиями (пространственный принцип);
2. Кодирование определенным
Кодировании модальности и качества
1. Кодирование меченными линиями (пространственный принцип);
2. Кодирование определенным

Кодирование интенсивности
Более сильное раздражение вызывает более сильное ощущение.
Осуществляется 2 способами:
Частотой импульсов
Кодирование интенсивности
Более сильное раздражение вызывает более сильное ощущение.
Осуществляется 2 способами:
Частотой импульсов

Кодирование места, формы и размеров
Происходит по пространственному принципу. Данный способ кодирования
Кодирование места, формы и размеров
Происходит по пространственному принципу. Данный способ кодирования

РАЗЛИЧЕНИЕ СИГНАЛОВ
РАЗЛИЧЕНИЕ СИГНАЛОВ

Различение интенсивности
Различение интенсивности стимулов происходит в соответствии с законом Вебера-Фехнера.
Рецепторы
Различение интенсивности
Различение интенсивности стимулов происходит в соответствии с законом Вебера-Фехнера.
Рецепторы
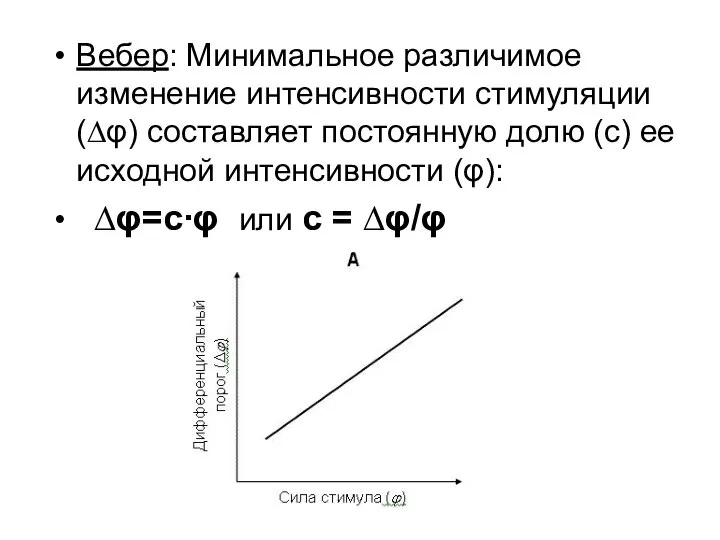
Вебер: Минимальное различимое изменение интенсивности стимуляции (∆φ) составляет постоянную долю (с)
Вебер: Минимальное различимое изменение интенсивности стимуляции (∆φ) составляет постоянную долю (с)

Фехнер предложил использовать дифференциальные пороги для измерения субъективных ощущений и построения
Фехнер предложил использовать дифференциальные пороги для измерения субъективных ощущений и построения

Различение пространственных параметров
Пространственное различение стимулов возможно в том случае, если между
Различение пространственных параметров
Пространственное различение стимулов возможно в том случае, если между

Различение временных параметров
Для временного различения раздражителей важно, чтобы они не сливались
Различение временных параметров
Для временного различения раздражителей важно, чтобы они не сливались

Функции сенсорных систем
Обнаружение сигналов
Различение сигналов
Кодирование информации
Передача и преобразование сигналов
Осознание образов
Функции сенсорных систем
Обнаружение сигналов
Различение сигналов
Кодирование информации
Передача и преобразование сигналов
Осознание образов

4. Передача и преобразование сигналов
4. Передача и преобразование сигналов

Преобразования сигналов могут быть разделены на:
Пространственные (усиление сигнала) – изменение соотношения
Преобразования сигналов могут быть разделены на:
Пространственные (усиление сигнала) – изменение соотношения

Пространственные (усиление сигнала) преобразования сигналов
Пространственные (усиление сигнала) преобразования сигналов

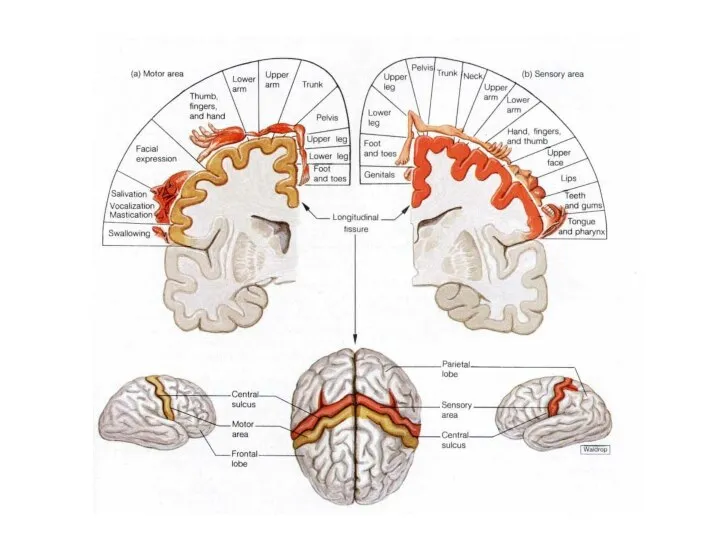
«Сенсорный гомункулюс»
В зрительной и соматосенсорных системах на корковом уровне значительно искажаются
«Сенсорный гомункулюс»
В зрительной и соматосенсорных системах на корковом уровне значительно искажаются

1981- теория колончатой организации коры
(в 60-х годах прошлого века V.Mountcastle)
1981- теория колончатой организации коры (в 60-х годах прошлого века V.Mountcastle)
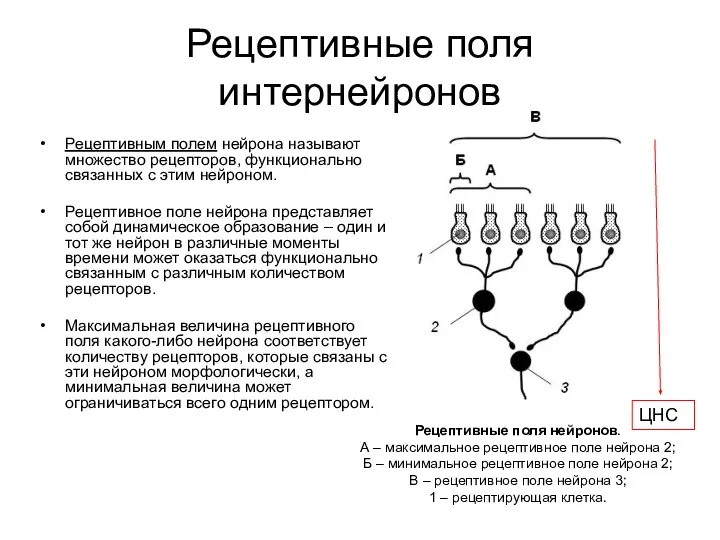
Рецептивные поля интернейронов
Рецептивным полем нейрона называют множество рецепторов, функционально связанных с
Рецептивные поля интернейронов
Рецептивным полем нейрона называют множество рецепторов, функционально связанных с

Перекрытие рецептивных полей.
Взаимодействие рецепторов в рецептивном поле.
За счет конвергенции
Перекрытие рецептивных полей.
Взаимодействие рецепторов в рецептивном поле.
За счет конвергенции

Подавление информации о менее существенных сигналах происходит за счет возвратного и
Подавление информации о менее существенных сигналах происходит за счет возвратного и

В ходе преобразования сигналов происходит:
Ограничение избыточности информации и выделение существенных признаков
В ходе преобразования сигналов происходит:
Ограничение избыточности информации и выделение существенных признаков

Детектирование сигналов
Это избирательное выделение сенсорным нейроном признака раздражителя, имеющего поведенческое
Детектирование сигналов
Это избирательное выделение сенсорным нейроном признака раздражителя, имеющего поведенческое


Зрительные корковые нейроны с простым рецептивным полем активируются при воздействии на
Зрительные корковые нейроны с простым рецептивным полем активируются при воздействии на

Для активации нейронов со сложным рецептивным полем необходимы не только оформленный
Для активации нейронов со сложным рецептивным полем необходимы не только оформленный

Иллюзия Мюллера-Лайера также объясняется организацией сложных рецептивных полей корковых нейронов, возбуждение
Иллюзия Мюллера-Лайера также объясняется организацией сложных рецептивных полей корковых нейронов, возбуждение

5. Опознание образов
5. Опознание образов

Это конечная и наиболее сложная функция сенсорной системы.
Она заключается в
Это конечная и наиболее сложная функция сенсорной системы.
Она заключается в

ВЗАИМОДЕЙСТВИЕ СЕНСОРНЫХ СИСТЕМ
Взаимодействие сенсорных систем осуществляется на спинальном, ретикулярном, таламическом и
ВЗАИМОДЕЙСТВИЕ СЕНСОРНЫХ СИСТЕМ
Взаимодействие сенсорных систем осуществляется на спинальном, ретикулярном, таламическом и
 ТОРГОВЛЯ «УНИВЕРСАМЫ,СУПЕРМАРКЕТЫ» 5 класс ГБС(К)ОУ школа –интернат VIIIвида станицы Медведовской Учитель социально-бытовой ор
ТОРГОВЛЯ «УНИВЕРСАМЫ,СУПЕРМАРКЕТЫ» 5 класс ГБС(К)ОУ школа –интернат VIIIвида станицы Медведовской Учитель социально-бытовой ор Нейронные сети
Нейронные сети Гармонический осциллятор
Гармонический осциллятор  Классификация образовательных электронных изданий: основные принципы и критерии Содержание. - презентация
Классификация образовательных электронных изданий: основные принципы и критерии Содержание. - презентация Менеджмент и управление: основные понятия
Менеджмент и управление: основные понятия Вольфганг Амадей Моцарт - презентация по МХК_
Вольфганг Амадей Моцарт - презентация по МХК_ Противодействие коррупции
Противодействие коррупции  Формы государственного управления
Формы государственного управления  Презентация на тему "Творческие задания как активизация мыслительной деятельности учащихся" - скачать презентации по Педаго
Презентация на тему "Творческие задания как активизация мыслительной деятельности учащихся" - скачать презентации по Педаго Основные средства оптимизация товарного ассортимента в международной деятельности фирмы Юрлова В. Нечаева А.
Основные средства оптимизация товарного ассортимента в международной деятельности фирмы Юрлова В. Нечаева А. Виды товарных знаков
Виды товарных знаков  Корейский язык
Корейский язык ЭЛАСТИЧНОСТЬ СПРОСА И ПРЕДЛОЖЕНИЯ Тема 5
ЭЛАСТИЧНОСТЬ СПРОСА И ПРЕДЛОЖЕНИЯ Тема 5  Архитектура предприятия
Архитектура предприятия Интеллектуальные остановки как средство формирования краеведческого кругозора горожан
Интеллектуальные остановки как средство формирования краеведческого кругозора горожан Поселок на 2500-3000 жителей
Поселок на 2500-3000 жителей «Улыбка Джоконды Леонардо да Винчи» Работу выполнили Опутина Анна, Тетеркина Кристина
«Улыбка Джоконды Леонардо да Винчи» Работу выполнили Опутина Анна, Тетеркина Кристина Презентация ВОИС (всемирная организация интеллектуальной собственности)
Презентация ВОИС (всемирная организация интеллектуальной собственности) Виды источника творчества в дизайне костюма. Этапы изучения творческого источника - предмета «Все, что вижу, слышу, знаю, превращу
Виды источника творчества в дизайне костюма. Этапы изучения творческого источника - предмета «Все, что вижу, слышу, знаю, превращу  Лекция 4.Автоматизация кассовых и банковских операций.
Лекция 4.Автоматизация кассовых и банковских операций. Хвойные ростения
Хвойные ростения Электротехника, электроника и схемотехника
Электротехника, электроника и схемотехника Введение в UML
Введение в UML Настройка и работа с антивирусными программами ПО ДОКТОР ВЭБ
Настройка и работа с антивирусными программами ПО ДОКТОР ВЭБ Физическая культура в общекультурной и профессиональной подготовке студентов
Физическая культура в общекультурной и профессиональной подготовке студентов Книга Иисуса Навина
Книга Иисуса Навина История развития оперативной памяти ЭВМ
История развития оперативной памяти ЭВМ Конструктивные разновидности и системы балок
Конструктивные разновидности и системы балок