- ФИЗИОЛОГИЯ ГЛАДКИХ МЫШЦ. ФИЗИОЛОГИЯ НЕРВОВ

Содержание
- 2. ПЛАН: Структурная и функциональная характеристика гладких мышц. Классификация нервов. Распространение возбуждения по безмиелиновым и миелиновым волокнам.
- 3. СТРУКТУРНАЯ И ФУНКЦИОНАЛЬНАЯ ХАРАКТЕРИСТИКА ГЛАДКИХ МЫШЦ 1. В гладкомышечных клетках миофибриллы расположены беспорядочно, нет исчерченности, нет
- 4. СТРУКТУРНАЯ И ФУНКЦИОНАЛЬНАЯ ХАРАКТЕРИСТИКА ГЛАДКИХ МЫШЦ 4. Соотношение актина к миозину 15:1 (в скелетных мышцах 2:1).
- 5. СТРУКТУРНАЯ И ФУНКЦИОНАЛЬНАЯ ХАРАКТЕРИСТИКА ГЛАДКИХ МЫШЦ 7. В состав тонких филаментов, кроме актина, входят: кальдесмон, кальпонин,
- 6. КЛАССИФИКАЦИЯ НЕРВОВ 1. По направлению распространения возбуждения – афферентные, эфферентные (двигательные – вызывающие фазную и тоническую
- 7. РАСПРОСТРАНЕНИЕ ВОЗБУЖДЕНИЯ ПО НЕРВАМ Значение – один из способов передачи информации или контролирующих сигналов. 206
- 8. МЕХАНИЗМ распространения – электротон, т.е. с помощью силовых линий постоянного тока. Теория малых токов создана Германом
- 9. Возбуждение передается по цитоплазматической мембране нерва по поверхности, покрытой межклеточной жидкостью, а не по аксоплазме, обладающей
- 10. В каждом участке мембраны полюса источника напряжения находятся внутри и снаружи волокна и ток является чисто
- 11. В клетках, неспособных генерировать ПД (глия, эпителий, тонические мышечные волокна), распространение электротона является важным механизмом клеточной
- 12. СПОСОБЫ РАСПРОСТРАНЕНИЯ ВОЗБУЖДЕНИЯ ПО НЕРВУ В безмякотном волокне – непрерывно: потенциал действия воспроизводится в каждом участке.
- 13. Миелиновое нервное волокно состоит из осевого цилиндра (аксона), вокруг которого шванновские клетки образуют миелиновую оболочку за
- 14. МИЕЛИНОВОЕ НЕРВНОЕ ВОЛОКНО (A.Guyton, J.Hall, 2000) Миелиновая оболочка Цитоплазма Швановской клетки Ядро Швановской клетки Перехват Ранвье
- 15. В мякотном волокне возбуждение распространяется сальтаторно, т.к. миелин – диэлектрик. Полностью окружая аксон в межузловых промежутках,
- 16. В БЕЗМЯКОТНОМ ВОЛОКНЕ ПОТЕНЦИАЛ ДЕЙСТВИЯ ВОСПРОИЗВОДИТСЯ В КАЖДОМ УЧАСТКЕ 215
- 17. Фактор надежности или гарантийный фактор – отношение амплитуды потенциала действия к величине порога деполяризации ФН=ПД:Vt Vt
- 18. Миелиновые нервные волокна Безмиелиновые нервные волокна диаметр волокна, мкм скорость проведения возбуждения СКОРОСТЬ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ (впервые
- 19. ЗАКОНЫ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ ПО НЕРВУ 218
- 20. 1 закон – анатомической и физиологической целостности: для проведения возбуждения необходима не только механическая целостность нерва,
- 21. 2 закон – двустороннего проведения возбуждения: от места раздражения возбуждение по нерву распространяется в обе стороны.
- 22. 3 закон – изолированного проведения возбуждения: возбуждение, распространяющееся по волокну, не передается на соседние (миелин –
- 23. НЕРВНЫЙ СТВОЛ – пучок множества нервных волокон, имеющих общие эпителиальные и соединительнотканные оболочки. Обычно включает в
- 24. Схемы дисперсии компонентов составного однофазного потенциала действия нерва (связанных с А (α, β, γ, Δ) В-
- 25. КЛАССИФИКАЦИЯ НЕРВНЫХ ВОЛОКОН ПО СКОРОСТИ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ ТИП ВОЛОКНА ФУНКЦИЯ ДИАМЕТР, мкм C 0,4 - 1,2
- 26. КЛАССИФИКАЦИЯ ЭРЛАНГЕРА И ГАССЕРА (ЛАТИНСКИЕ БУКВЫ) И АЛЬТЕРНАТИВНЫЕ ДОБАВЛЕНИЯ (РИМСКИЕ ЦИФРЫ) Типы нервных волокон и их
- 27. ТРОФИЧЕСКАЯ ФУНКЦИЯ НЕРВНОЙ СИСТЕМЫ (И.П. ПАВЛОВ) Впервые установил И.П. Павлов, изучая регуляцию сердечной деятельности – открыл
- 28. Сперанский доказал, что соматические нервы также обладают трофическим действием – при длительном раздражении тройничного нерва, иннервирующего
- 29. Основной центр, регулирующий трофические влияния, расположен в области гипоталамуса, где сосредоточены высшие обменные центры (углеводного, жирового
- 30. Доказательство – М.К. Петрова заметила, что у собак, у которых долго вырабатывали условные рефлексы, появляются трофические
- 31. АКСОННЫЙ ТРАНСПОРТ Na+ На рисунке представлен гипотетический транспортный механизм нервного волокна. Микротрубочки и нейрофиламенты имеют тонкие
- 32. Упрощённо аксонный транспорт можно представить как систему, состоящую из нескольких элементов. В неё входят: груз белки-моторы,
- 33. ВИДЫ АКСОННОГО ТРАНСПОРТА Быстрый – осуществляется со скоростью до 400 мм в сутки. Так переносятся нецитозольные
- 34. ВИДЫ АКСОННОГО ТРАНСПОРТА Ретроградный - транспорт в обратном направлении. Антероградный - транспорт материалов от тела нейрона
- 35. НЕЙРОТРОФИНЫ (ТРОФИЧЕСКАЯ ПОДДЕРЖКА НЕЙРОНА) Это белковые вещества необходимые для жизнедеятельности и роста нейронов. Продуцируются мышцами или
- 36. НЕЙРОТРОФИНЫ развитии росте выживаемости пластичности нейронов. Связываются с рецепторами, расположенными на нервных окончаниях, затем интернализируются и
- 37. НЕЙРОТРОФИНЫ 236
- 38. ДРУГИЕ ФАКТОРЫ, ИЗМЕНЯЮЩИЕ РОСТ НЕЙРОНОВ Цилиарный нейротрофный фактор Лейкемический ингибиторный фактор Инсулино-подобный фактор роста I Трансформирующий
- 40. Скачать презентацию

ПЛАН:
Структурная и функциональная характеристика гладких мышц.
Классификация нервов. Распространение возбуждения по безмиелиновым
ПЛАН: Структурная и функциональная характеристика гладких мышц. Классификация нервов. Распространение возбуждения по безмиелиновым

СТРУКТУРНАЯ И ФУНКЦИОНАЛЬНАЯ ХАРАКТЕРИСТИКА ГЛАДКИХ МЫШЦ
1. В гладкомышечных клетках миофибриллы
СТРУКТУРНАЯ И ФУНКЦИОНАЛЬНАЯ ХАРАКТЕРИСТИКА ГЛАДКИХ МЫШЦ 1. В гладкомышечных клетках миофибриллы

СТРУКТУРНАЯ И ФУНКЦИОНАЛЬНАЯ ХАРАКТЕРИСТИКА ГЛАДКИХ МЫШЦ
4. Соотношение актина к миозину
СТРУКТУРНАЯ И ФУНКЦИОНАЛЬНАЯ ХАРАКТЕРИСТИКА ГЛАДКИХ МЫШЦ 4. Соотношение актина к миозину

СТРУКТУРНАЯ И ФУНКЦИОНАЛЬНАЯ ХАРАКТЕРИСТИКА ГЛАДКИХ МЫШЦ 7. В состав тонких филаментов,
СТРУКТУРНАЯ И ФУНКЦИОНАЛЬНАЯ ХАРАКТЕРИСТИКА ГЛАДКИХ МЫШЦ 7. В состав тонких филаментов,

КЛАССИФИКАЦИЯ НЕРВОВ
1. По направлению распространения возбуждения – афферентные, эфферентные (двигательные –
КЛАССИФИКАЦИЯ НЕРВОВ
1. По направлению распространения возбуждения – афферентные, эфферентные (двигательные –

РАСПРОСТРАНЕНИЕ ВОЗБУЖДЕНИЯ ПО НЕРВАМ
Значение – один из способов передачи информации
РАСПРОСТРАНЕНИЕ ВОЗБУЖДЕНИЯ ПО НЕРВАМ Значение – один из способов передачи информации
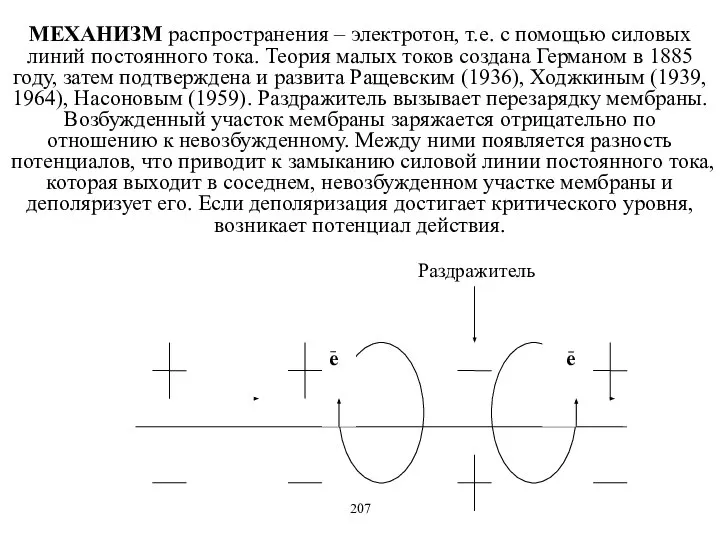
МЕХАНИЗМ распространения – электротон, т.е. с помощью силовых линий постоянного тока.
МЕХАНИЗМ распространения – электротон, т.е. с помощью силовых линий постоянного тока.

Возбуждение передается по цитоплазматической мембране нерва по поверхности, покрытой межклеточной жидкостью,
Возбуждение передается по цитоплазматической мембране нерва по поверхности, покрытой межклеточной жидкостью,
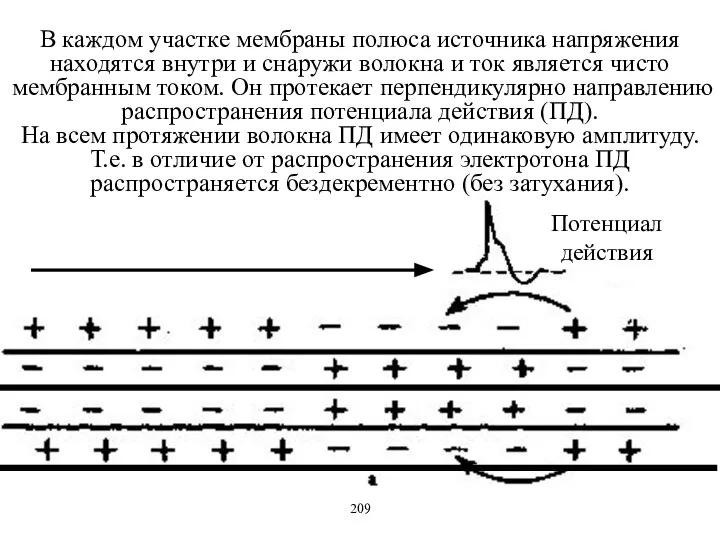
В каждом участке мембраны полюса источника напряжения находятся внутри и снаружи
В каждом участке мембраны полюса источника напряжения находятся внутри и снаружи

В клетках, неспособных генерировать ПД (глия, эпителий, тонические мышечные волокна), распространение
В клетках, неспособных генерировать ПД (глия, эпителий, тонические мышечные волокна), распространение
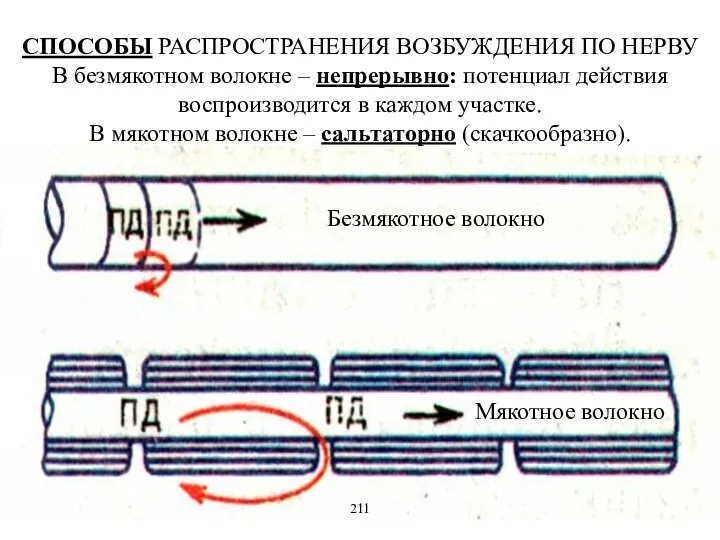
СПОСОБЫ РАСПРОСТРАНЕНИЯ ВОЗБУЖДЕНИЯ ПО НЕРВУ
В безмякотном волокне – непрерывно: потенциал действия
СПОСОБЫ РАСПРОСТРАНЕНИЯ ВОЗБУЖДЕНИЯ ПО НЕРВУ В безмякотном волокне – непрерывно: потенциал действия

Миелиновое нервное волокно состоит из осевого цилиндра (аксона), вокруг которого шванновские
Миелиновое нервное волокно состоит из осевого цилиндра (аксона), вокруг которого шванновские
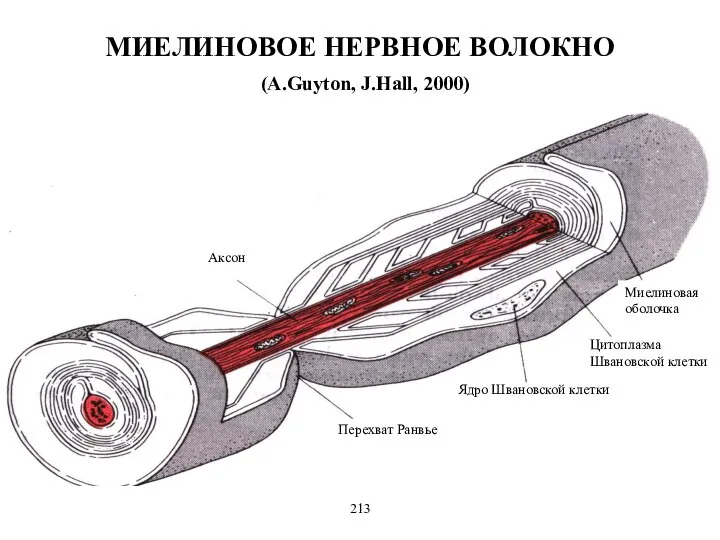
МИЕЛИНОВОЕ НЕРВНОЕ ВОЛОКНО
(A.Guyton, J.Hall, 2000)
Миелиновая оболочка
Цитоплазма Швановской клетки
Ядро Швановской клетки
Перехват
МИЕЛИНОВОЕ НЕРВНОЕ ВОЛОКНО
(A.Guyton, J.Hall, 2000)
Миелиновая оболочка
Цитоплазма Швановской клетки
Ядро Швановской клетки
Перехват

В мякотном волокне возбуждение распространяется сальтаторно, т.к. миелин – диэлектрик. Полностью
В мякотном волокне возбуждение распространяется сальтаторно, т.к. миелин – диэлектрик. Полностью

В БЕЗМЯКОТНОМ ВОЛОКНЕ ПОТЕНЦИАЛ ДЕЙСТВИЯ
ВОСПРОИЗВОДИТСЯ В КАЖДОМ УЧАСТКЕ
215
В БЕЗМЯКОТНОМ ВОЛОКНЕ ПОТЕНЦИАЛ ДЕЙСТВИЯ
ВОСПРОИЗВОДИТСЯ В КАЖДОМ УЧАСТКЕ
215
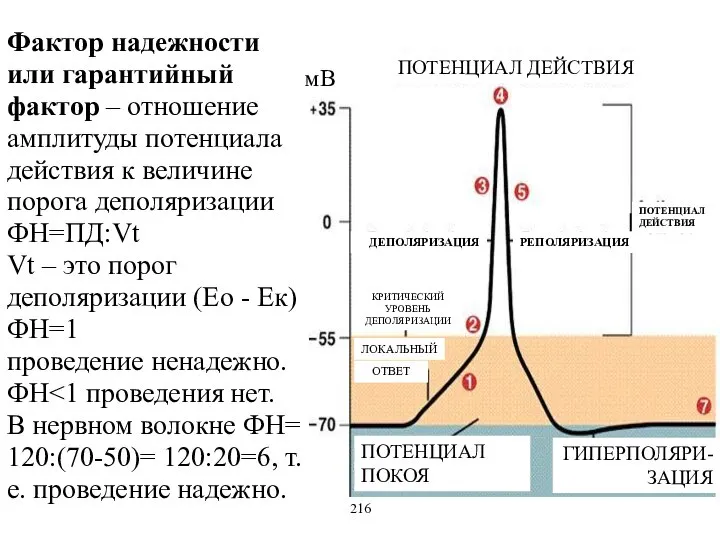
Фактор надежности или гарантийный фактор – отношение амплитуды потенциала действия к
Фактор надежности или гарантийный фактор – отношение амплитуды потенциала действия к
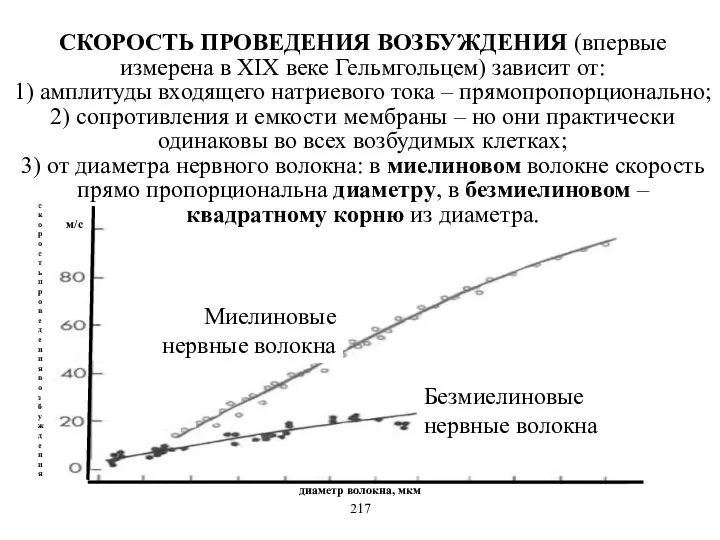
Миелиновые нервные волокна
Безмиелиновые нервные волокна
диаметр волокна, мкм
скорость проведения возбуждения
СКОРОСТЬ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ
Миелиновые нервные волокна
Безмиелиновые нервные волокна
диаметр волокна, мкм
скорость проведения возбуждения
СКОРОСТЬ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ

ЗАКОНЫ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ
ПО НЕРВУ
218
ЗАКОНЫ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ
ПО НЕРВУ
218

1 закон – анатомической и физиологической целостности:
для проведения возбуждения необходима не
1 закон – анатомической и физиологической целостности:
для проведения возбуждения необходима не

2 закон – двустороннего проведения возбуждения:
от места раздражения возбуждение по нерву
2 закон – двустороннего проведения возбуждения:
от места раздражения возбуждение по нерву

3 закон – изолированного проведения возбуждения:
возбуждение, распространяющееся по волокну, не передается
3 закон – изолированного проведения возбуждения:
возбуждение, распространяющееся по волокну, не передается
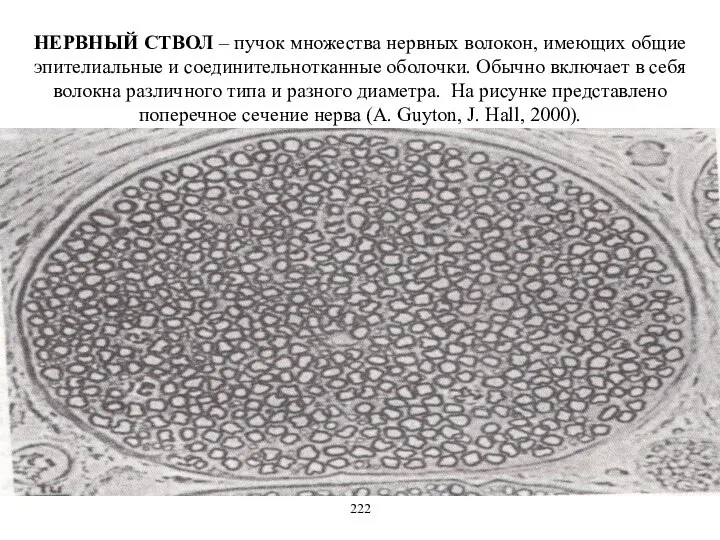
НЕРВНЫЙ СТВОЛ – пучок множества нервных волокон, имеющих общие эпителиальные и
НЕРВНЫЙ СТВОЛ – пучок множества нервных волокон, имеющих общие эпителиальные и
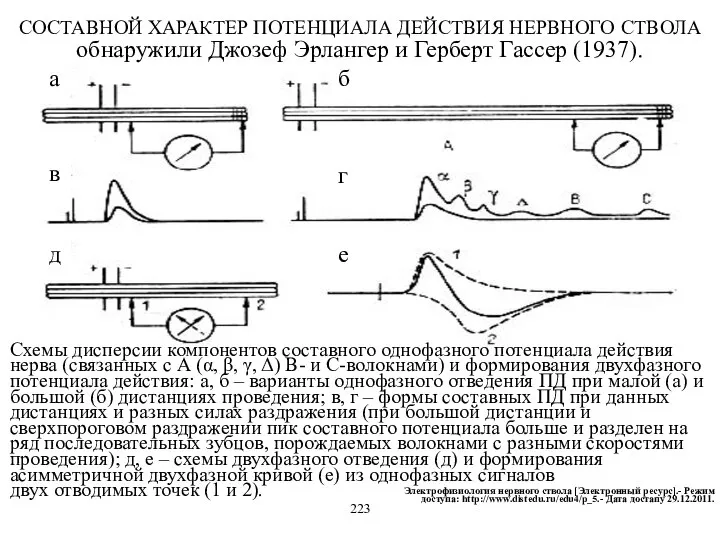
Схемы дисперсии компонентов составного однофазного потенциала действия нерва (связанных с А
Схемы дисперсии компонентов составного однофазного потенциала действия нерва (связанных с А
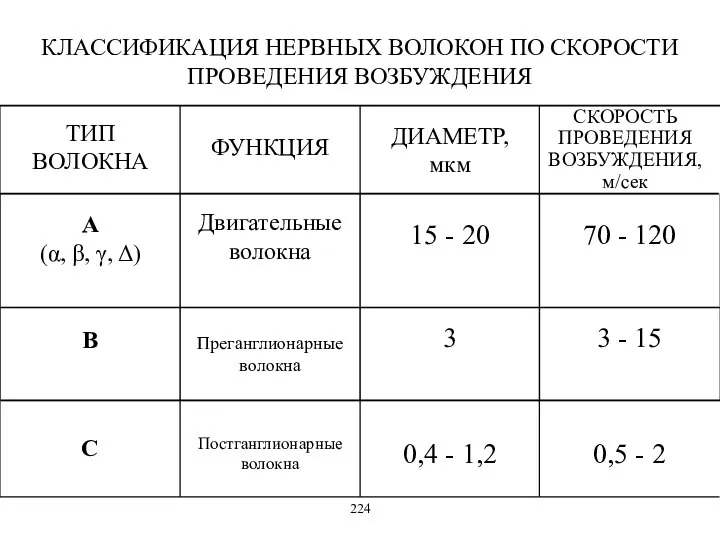
КЛАССИФИКАЦИЯ НЕРВНЫХ ВОЛОКОН ПО СКОРОСТИ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ
ТИП ВОЛОКНА
ФУНКЦИЯ
ДИАМЕТР,
мкм
C
0,4 - 1,2
0,5 -
КЛАССИФИКАЦИЯ НЕРВНЫХ ВОЛОКОН ПО СКОРОСТИ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ
ТИП ВОЛОКНА
ФУНКЦИЯ
ДИАМЕТР,
мкм
C
0,4 - 1,2
0,5 -

КЛАССИФИКАЦИЯ ЭРЛАНГЕРА И ГАССЕРА (ЛАТИНСКИЕ БУКВЫ) И АЛЬТЕРНАТИВНЫЕ ДОБАВЛЕНИЯ (РИМСКИЕ ЦИФРЫ)
Типы
КЛАССИФИКАЦИЯ ЭРЛАНГЕРА И ГАССЕРА (ЛАТИНСКИЕ БУКВЫ) И АЛЬТЕРНАТИВНЫЕ ДОБАВЛЕНИЯ (РИМСКИЕ ЦИФРЫ)
Типы

ТРОФИЧЕСКАЯ ФУНКЦИЯ НЕРВНОЙ СИСТЕМЫ (И.П. ПАВЛОВ)
Впервые установил И.П. Павлов, изучая регуляцию
ТРОФИЧЕСКАЯ ФУНКЦИЯ НЕРВНОЙ СИСТЕМЫ (И.П. ПАВЛОВ)
Впервые установил И.П. Павлов, изучая регуляцию

Сперанский доказал, что соматические нервы также обладают трофическим действием – при
Сперанский доказал, что соматические нервы также обладают трофическим действием – при
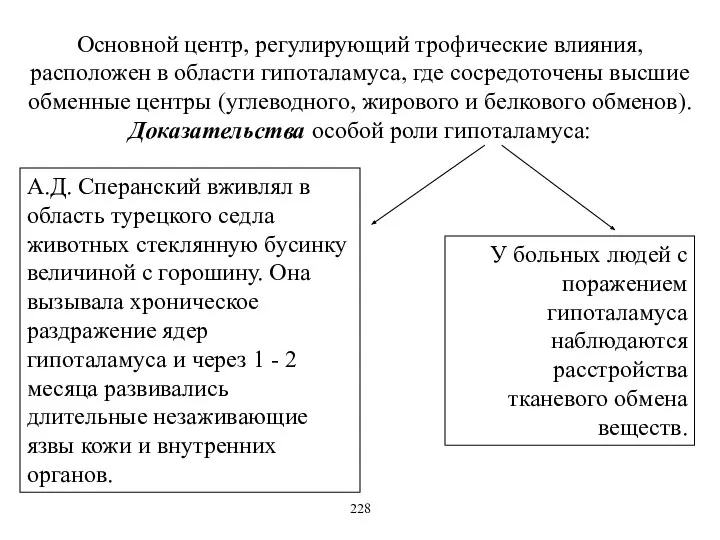
Основной центр, регулирующий трофические влияния, расположен в области гипоталамуса, где сосредоточены
Основной центр, регулирующий трофические влияния, расположен в области гипоталамуса, где сосредоточены

Доказательство – М.К. Петрова заметила, что у собак, у которых долго
Доказательство – М.К. Петрова заметила, что у собак, у которых долго
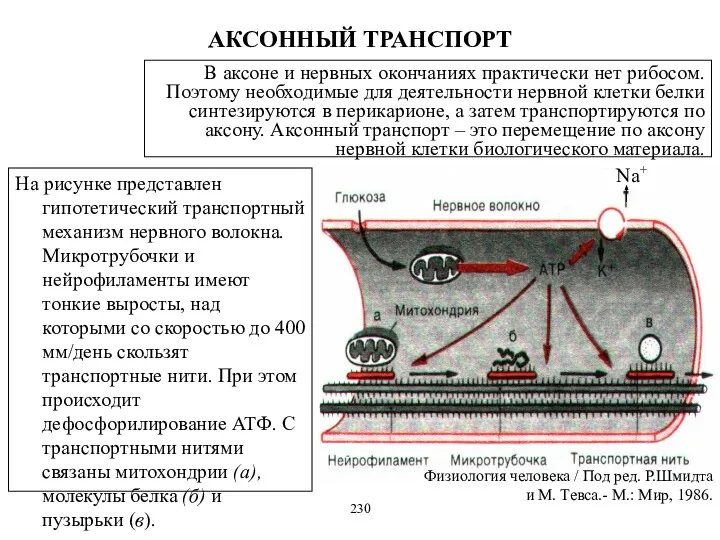
АКСОННЫЙ ТРАНСПОРТ
Na+
На рисунке представлен гипотетический транспортный механизм нервного волокна. Микротрубочки и
АКСОННЫЙ ТРАНСПОРТ
Na+
На рисунке представлен гипотетический транспортный механизм нервного волокна. Микротрубочки и
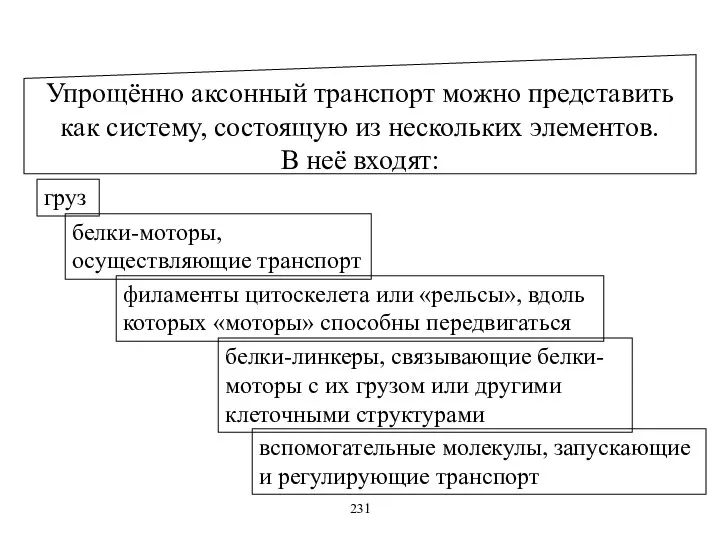
Упрощённо аксонный транспорт можно представить как систему, состоящую из нескольких элементов.
В
Упрощённо аксонный транспорт можно представить как систему, состоящую из нескольких элементов. В

ВИДЫ АКСОННОГО ТРАНСПОРТА
Быстрый – осуществляется со скоростью до 400 мм в
ВИДЫ АКСОННОГО ТРАНСПОРТА
Быстрый – осуществляется со скоростью до 400 мм в
ВИДЫ АКСОННОГО ТРАНСПОРТА
Ретроградный
- транспорт в обратном направлении.
Антероградный - транспорт материалов от
ВИДЫ АКСОННОГО ТРАНСПОРТА
Ретроградный
- транспорт в обратном направлении.
Антероградный - транспорт материалов от
НЕЙРОТРОФИНЫ
(ТРОФИЧЕСКАЯ ПОДДЕРЖКА НЕЙРОНА)
Это белковые вещества необходимые для жизнедеятельности и роста нейронов.
Продуцируются
НЕЙРОТРОФИНЫ
(ТРОФИЧЕСКАЯ ПОДДЕРЖКА НЕЙРОНА)
Это белковые вещества необходимые для жизнедеятельности и роста нейронов.
Продуцируются

НЕЙРОТРОФИНЫ
развитии
росте
выживаемости
пластичности нейронов.
Связываются с рецепторами, расположенными на нервных окончаниях, затем интернализируются и
НЕЙРОТРОФИНЫ
развитии
росте
выживаемости
пластичности нейронов.
Связываются с рецепторами, расположенными на нервных окончаниях, затем интернализируются и

НЕЙРОТРОФИНЫ
236
НЕЙРОТРОФИНЫ
236
ДРУГИЕ ФАКТОРЫ, ИЗМЕНЯЮЩИЕ РОСТ НЕЙРОНОВ
Цилиарный нейротрофный фактор
Лейкемический ингибиторный фактор
Инсулино-подобный фактор роста
ДРУГИЕ ФАКТОРЫ, ИЗМЕНЯЮЩИЕ РОСТ НЕЙРОНОВ
Цилиарный нейротрофный фактор
Лейкемический ингибиторный фактор
Инсулино-подобный фактор роста
 Авангардизм в натюрморте настроения
Авангардизм в натюрморте настроения Софийские соборы Киева и Новгорода
Софийские соборы Киева и Новгорода Состав слова - презентация для начальной школы_
Состав слова - презентация для начальной школы_ Виды, разрезы, сечения. ГОСТ 2.305-2008
Виды, разрезы, сечения. ГОСТ 2.305-2008 Обеспечение общественного порядка и противодействие преступности
Обеспечение общественного порядка и противодействие преступности Многогранники и их виды. Графические модели многогранников. (Лекция 5)
Многогранники и их виды. Графические модели многогранников. (Лекция 5) Восприятие русской аудиторией образа женщины-воина в современной японской массовой культуре
Восприятие русской аудиторией образа женщины-воина в современной японской массовой культуре 10 занятие_стом.ppt
10 занятие_стом.ppt Презентация на тему "ЛЕКАРСТВЕННЫЕ СРЕДСТВА ВЛИЯЮЩИЕ НА ФУНКЦИЮ ОРГАНОВ ПИЩЕВАРЕНИЯ" - скачать презентации по Медицине
Презентация на тему "ЛЕКАРСТВЕННЫЕ СРЕДСТВА ВЛИЯЮЩИЕ НА ФУНКЦИЮ ОРГАНОВ ПИЩЕВАРЕНИЯ" - скачать презентации по Медицине Презентация "Семь чудес света" - скачать презентации по МХК
Презентация "Семь чудес света" - скачать презентации по МХК Футбольная форма и инвентарь
Футбольная форма и инвентарь ЧРЕЗВЫЧАЙНЫЕ СИТУАЦИИ И МЕТОДЫ ЗАЩИТЫ В УСЛОВИЯХ ИХ РЕАЛИЗАЦИИ
ЧРЕЗВЫЧАЙНЫЕ СИТУАЦИИ И МЕТОДЫ ЗАЩИТЫ В УСЛОВИЯХ ИХ РЕАЛИЗАЦИИ Инфаркт миокарда
Инфаркт миокарда Нарышкинский детский сад «Солнышко».
Нарышкинский детский сад «Солнышко». Сommunication
Сommunication  Деятельность П.А.Сорокина Выполнили: Стародворов Вадим Ращупкин Артем
Деятельность П.А.Сорокина Выполнили: Стародворов Вадим Ращупкин Артем Циркуляция
Циркуляция День Святого Валентина
День Святого Валентина Бутик дизайнерской одежды “Salsa”. Бизнес-проект
Бутик дизайнерской одежды “Salsa”. Бизнес-проект Три медведя
Три медведя  ГОСУДАРСТВЕННЫЕ ЦЕННЫЕ БУМАГИ I. Общая характеристика II. Виды государственных ценных бумаг III. Ценные бумаги региональных ор
ГОСУДАРСТВЕННЫЕ ЦЕННЫЕ БУМАГИ I. Общая характеристика II. Виды государственных ценных бумаг III. Ценные бумаги региональных ор Расчет оборудования для транспортировки, питатели, дозаторы
Расчет оборудования для транспортировки, питатели, дозаторы Ми проти насильства та жорстокости
Ми проти насильства та жорстокости Турнир по теннису
Турнир по теннису Без названия
Без названия Обереги земли Тасу Ява в наше время
Обереги земли Тасу Ява в наше время Параметры зубошевинговальных станков
Параметры зубошевинговальных станков Инвестиционный проект строительства административного здания
Инвестиционный проект строительства административного здания