- гормоны

Содержание
- 2. Основные системы регуляции метаболизма и межклеточной коммуникации ЦНС и периферическая нервная система эндокринная система пара- и
- 3. Эрне́ст Ге́нри Ста́рлинг (1866 -1927) — английский физиолог Гормоны- биологически активные органические вещества, химические посредники, которые
- 5. Общая характеристика гормонов дистантность действия строгая специфичность и избирательность биологического действия высокая активность(10-7-10-12 М) осуществляют действие
- 6. ИЕРАРХИЯ ДЕЙСТВИЯ ГОРМОНОВ Внешние и внутренние сигналы, поступающие в ЦНС по нейронам поступают в гипоталамус, где
- 7. РЕГУЛЯЦИЯ СИНТЕЗА И СЕКРЕЦИИ ГОРМОНОВ ЭНДОКРИННЫХ ЖЕЛЕЗ
- 9. Пептидные (белковые) гормоны - гидрофильные: содержат одну или несколько полипептидных цепей, число аминокислот от 2-3 десятков
- 10. Гормоны из отдельных аминокислот могут быть гидрофильными и гидрофобными. Образованы из аминокислот тирозин и триптофан. Представители:
- 11. Стероидные гормоны гидрофобные гормоны, построены на основе кольца циклопентанпергидрофенантрена. Образуют группы: ● Кортикостероидные гормоны – образуются
- 13. Рецептор лютеинизирующего гормона СТРУКТУРА МЕМБРАННЫХ РЕЦЕПТОРОВ
- 14. СТРУКТУРА ЦИТОПЛАЗМАТИЧЕСКИХ РЕЦЕПТОРОВ
- 15. Для передачи сигнала с мембранного гормон-рецепторного комплекса внутрь клетки существует СИСТЕМА ВТОРИЧНЫХ ПОСРЕДНИКОВ : ● Циклические
- 16. цАМФ является продуктом превращения АТФ при участии фермента аденилатциклазы, который находится в мембране клетки:
- 17. МЕХАНИЗМЫ ПЕРЕДАЧИ ГОРМОНАЛЬНЫХ СИГНАЛОВ В КЛЕТКУ: МЕМБРАННЫЙ ТИП РЕЦЕПЦИИ ● Все представители белково-пептидных гормонов ● Гидрофильные
- 18. МЕМБРАННЫЙ ТИП РЕЦЕПЦИИ МЕХАНИЗМЫ ПЕРЕДАЧИ ГОРМОНАЛЬНЫХ СИГНАЛОВ
- 19. Рецептор G-белок; 3) Фосфотидилинозитол-4, 5-бисфосфат (ФИФ)– источник свободного инозитол-1,4,5-трифосфата (ИФ3) и диацилглицерола (ДАГ); 4)Фосфолипаза С; Все
- 20. ИФ3 и ДАГ
- 21. Схема Са2+-механизма передачи гормонального сигнала
- 22. Свободный Са2+ связывается со специфическим белком: кальциевым рецептором в клетке - кальмодулином. Образуется комплекс: Са-кальмодулин, который
- 23. Участие Са-КМ в передачи гормонального сигнала в клетку.
- 24. Диацилглицерид (ДАГ) так же участвует в механизме передачи гормонального сигнала в клетку, вступая во взаимодействие с
- 25. Гормоны с цитозольным механизмом действия: ● Стероидные гормоны ● Кальцитриол ● Гидрофобные гормоны из отдельных аминокислот
- 26. ЦИТОПЛАЗМАТИЧЕСКИЙ ТИП РЕЦЕПЦИИ МЕХАНИЗМЫ ПЕРЕДАЧИ ГОРМОНАЛЬНЫХ СИГНАЛОВ
- 27. ИНСУЛИН Полипептид, состоящий из 2-х цепей:А-21 амкт; В-30 амкт, соединенных дисульфидными мостиками.
- 28. Гормональные эффекты Быстрые эффекты (с-мин.) – регуляция метаболизма и функций (метаботропные, сопряженные с G-белками) Медленные или
- 29. ФОРМИРОВАНИЕ ЭНДОКРИННОЙ СИСТЕМЫ В ЭМБРИОГЕНЕЗЕ Секреция интестинальных гормонов (кл.кишечника) с 18-20 дня фетального периода. Синтез гормонов
- 30. Для периода новорожденности характерно влияние гормонов матери, поступивших трасплацентарно, и с грудным молоком (в первые 3
- 31. В первые дни жизни отмечается высокий уровень тропных гормонов СОМАТОТРОПНЫЙ ГОРМОН (ГОРМОН РОСТА, СТГ) Простой белок.
- 32. ОБМЕН КАЛЬЦИЯ И ФОСФАТОВ ПАРАТИРЕОИДНЫЙ ГОРМОН, паратгормон, паратирин Синтезируется паращитовидными железами. Простой белок. Органы мишени-кости и
- 33. Роберт Лефковиц Брайан Кобилка (Robert Lefkowitz) (Brian Kobilka) из Стэнфорда из университета Дьюка Нобелевская премия по
- 34. 1) Рецептор (Са2+-мобилизующий рецептор) 2) G-белок; 3) Фосфотидилинозитол – источник свободного инозитолфосфата (ФИФ, ФИФ2) и диацилглицерида
- 35. Схема Са2+-механизма передачи гормонального сигнала
- 36. Свободный Са2+ связывается со специфическим белком: кальциевым рецептором в клетке - кальмодулином. Образуется комплекс: Са-кальмодулин, который
- 37. Участие Са-КМ в передачи гормонального сигнала в клетку.
- 39. Скачать презентацию

Основные системы регуляции метаболизма и межклеточной коммуникации
ЦНС и периферическая нервная система
эндокринная
Основные системы регуляции метаболизма и межклеточной коммуникации
ЦНС и периферическая нервная система
эндокринная

Эрне́ст Ге́нри Ста́рлинг
(1866 -1927) — английский физиолог
Гормоны- биологически активные органические
Эрне́ст Ге́нри Ста́рлинг
(1866 -1927) — английский физиолог
Гормоны- биологически активные органические
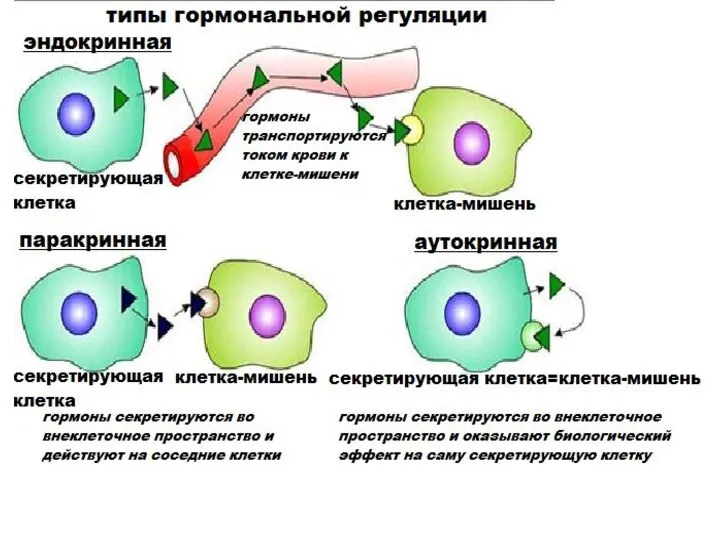

Общая характеристика гормонов
дистантность действия
строгая специфичность и избирательность биологического действия
высокая
Общая характеристика гормонов
дистантность действия
строгая специфичность и избирательность биологического действия
высокая
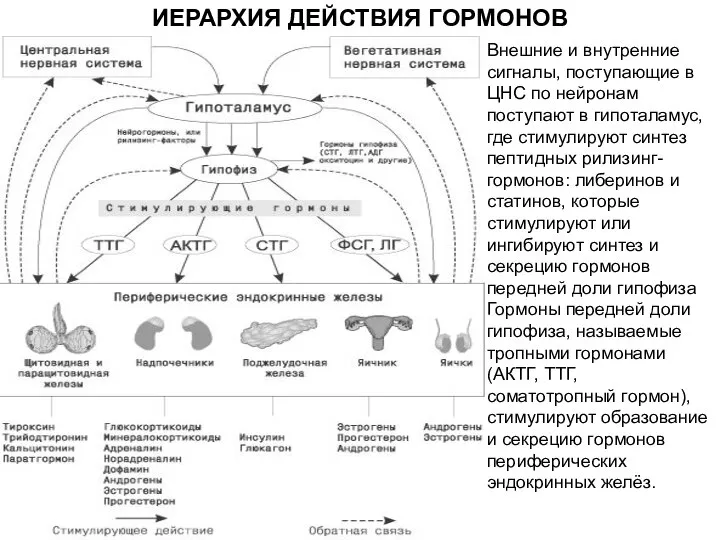
ИЕРАРХИЯ ДЕЙСТВИЯ ГОРМОНОВ
Внешние и внутренние сигналы, поступающие в ЦНС по нейронам
ИЕРАРХИЯ ДЕЙСТВИЯ ГОРМОНОВ
Внешние и внутренние сигналы, поступающие в ЦНС по нейронам
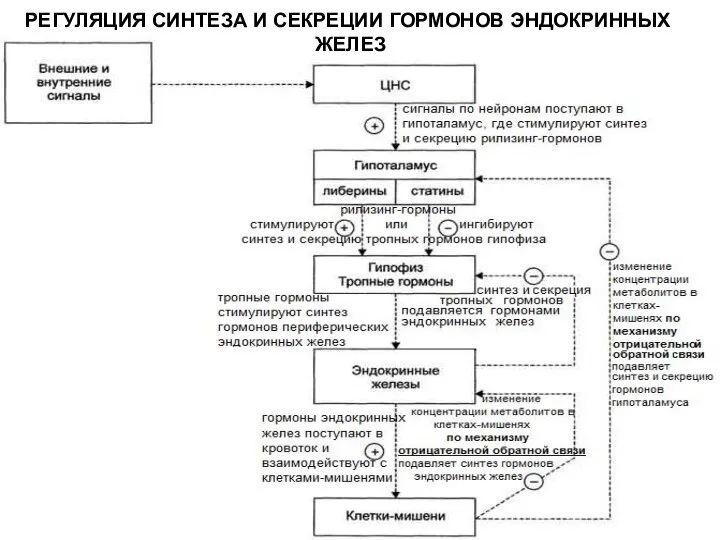
РЕГУЛЯЦИЯ СИНТЕЗА И СЕКРЕЦИИ ГОРМОНОВ ЭНДОКРИННЫХ
ЖЕЛЕЗ
РЕГУЛЯЦИЯ СИНТЕЗА И СЕКРЕЦИИ ГОРМОНОВ ЭНДОКРИННЫХ
ЖЕЛЕЗ
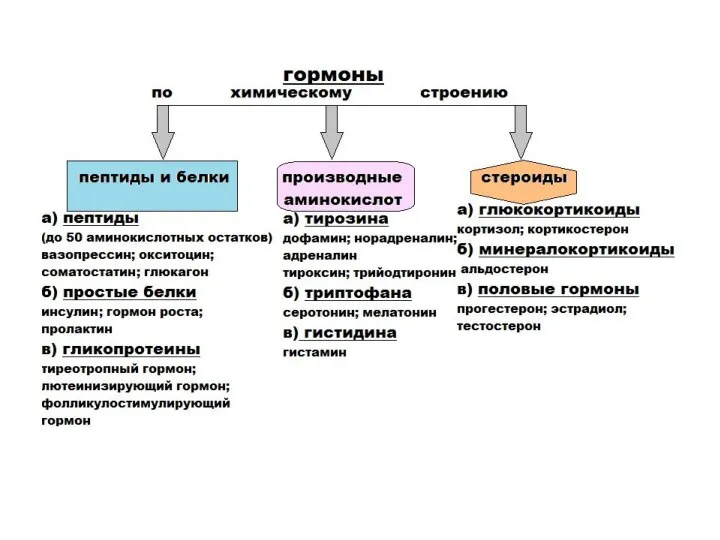

Пептидные (белковые) гормоны
- гидрофильные: содержат одну или несколько полипептидных цепей, число
Пептидные (белковые) гормоны
- гидрофильные: содержат одну или несколько полипептидных цепей, число

Гормоны из отдельных аминокислот
могут быть гидрофильными и гидрофобными. Образованы из
Гормоны из отдельных аминокислот
могут быть гидрофильными и гидрофобными. Образованы из

Стероидные гормоны
гидрофобные гормоны, построены на основе кольца циклопентанпергидрофенантрена.
Образуют группы:
●
Стероидные гормоны
гидрофобные гормоны, построены на основе кольца циклопентанпергидрофенантрена.
Образуют группы:
●
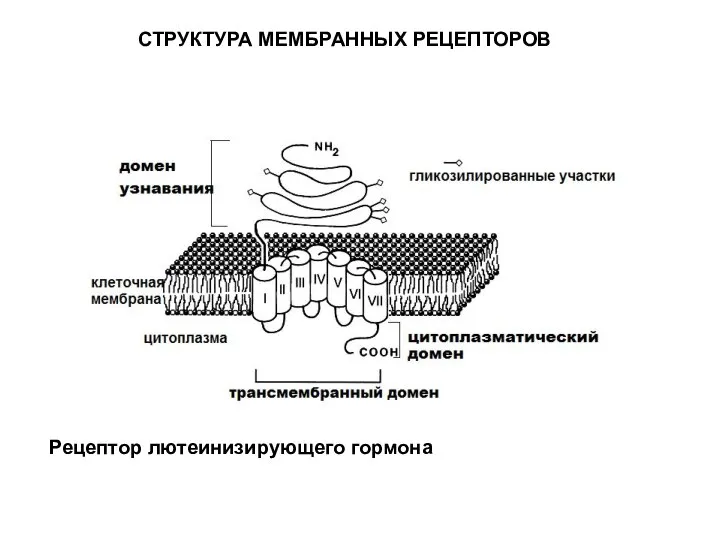
Рецептор лютеинизирующего гормона
СТРУКТУРА МЕМБРАННЫХ РЕЦЕПТОРОВ
Рецептор лютеинизирующего гормона
СТРУКТУРА МЕМБРАННЫХ РЕЦЕПТОРОВ
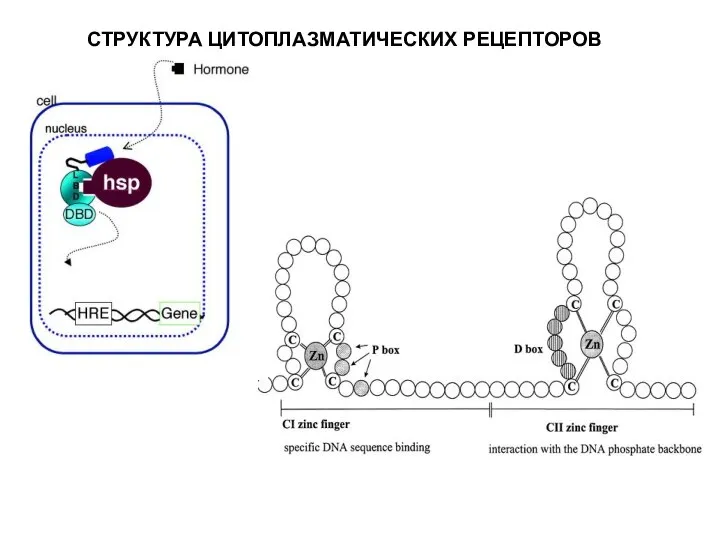
СТРУКТУРА ЦИТОПЛАЗМАТИЧЕСКИХ РЕЦЕПТОРОВ
СТРУКТУРА ЦИТОПЛАЗМАТИЧЕСКИХ РЕЦЕПТОРОВ

Для передачи сигнала с мембранного гормон-рецепторного комплекса внутрь клетки существует
СИСТЕМА
Для передачи сигнала с мембранного гормон-рецепторного комплекса внутрь клетки существует
СИСТЕМА
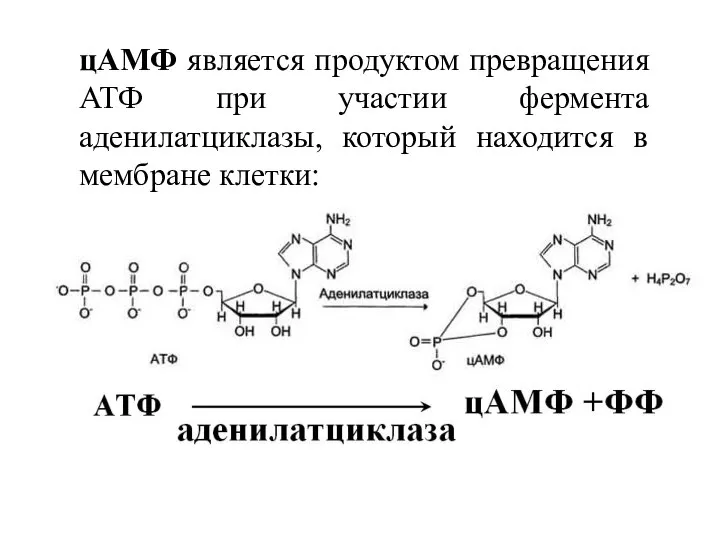
цАМФ является продуктом превращения АТФ при участии фермента аденилатциклазы, который находится
цАМФ является продуктом превращения АТФ при участии фермента аденилатциклазы, который находится
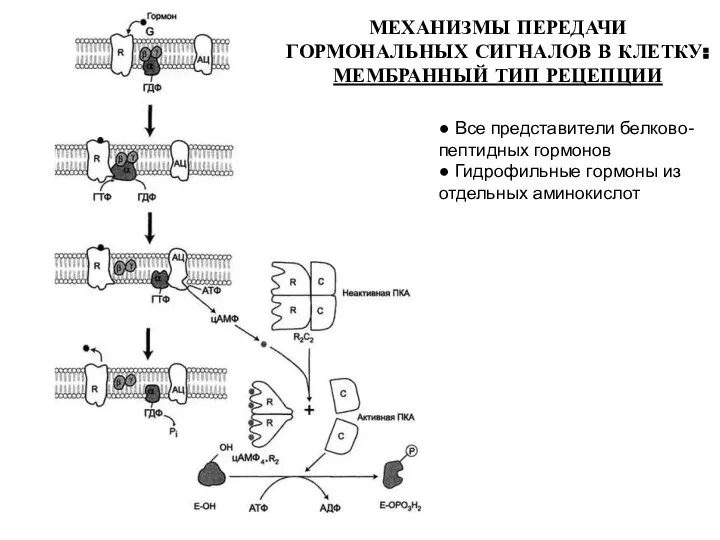
МЕХАНИЗМЫ ПЕРЕДАЧИ ГОРМОНАЛЬНЫХ СИГНАЛОВ В КЛЕТКУ: МЕМБРАННЫЙ ТИП РЕЦЕПЦИИ
● Все представители
МЕХАНИЗМЫ ПЕРЕДАЧИ ГОРМОНАЛЬНЫХ СИГНАЛОВ В КЛЕТКУ: МЕМБРАННЫЙ ТИП РЕЦЕПЦИИ
● Все представители
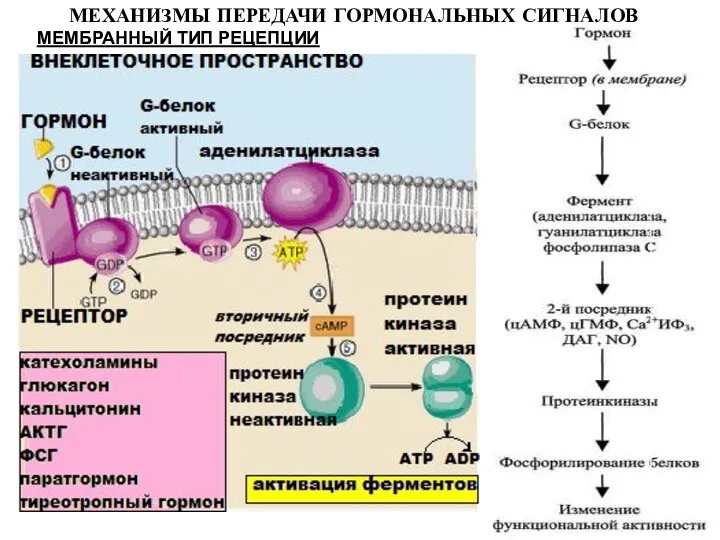
МЕМБРАННЫЙ ТИП РЕЦЕПЦИИ
МЕХАНИЗМЫ ПЕРЕДАЧИ ГОРМОНАЛЬНЫХ СИГНАЛОВ
МЕМБРАННЫЙ ТИП РЕЦЕПЦИИ
МЕХАНИЗМЫ ПЕРЕДАЧИ ГОРМОНАЛЬНЫХ СИГНАЛОВ

Рецептор
G-белок;
3) Фосфотидилинозитол-4, 5-бисфосфат (ФИФ)– источник свободного инозитол-1,4,5-трифосфата (ИФ3) и
Рецептор
G-белок;
3) Фосфотидилинозитол-4, 5-бисфосфат (ФИФ)– источник свободного инозитол-1,4,5-трифосфата (ИФ3) и
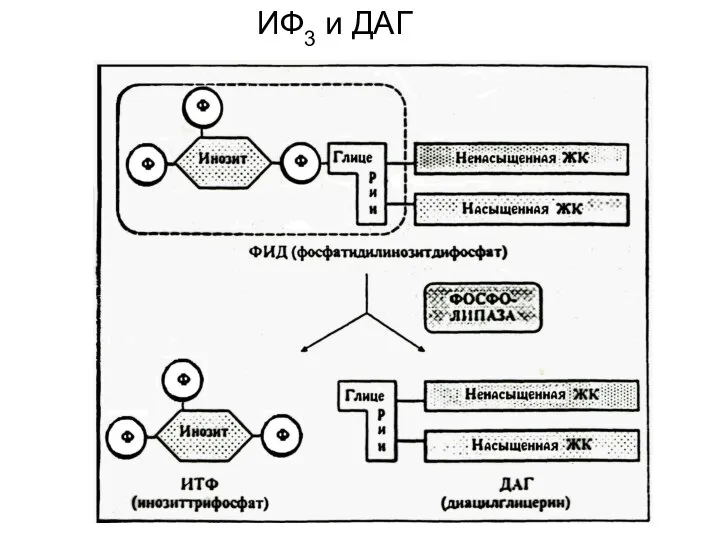
ИФ3 и ДАГ
ИФ3 и ДАГ
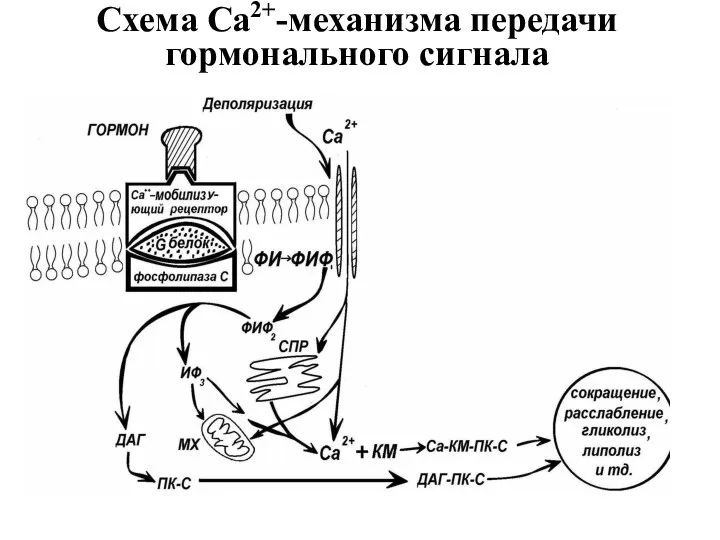
Схема Са2+-механизма передачи гормонального сигнала
Схема Са2+-механизма передачи гормонального сигнала

Свободный Са2+ связывается со специфическим белком: кальциевым рецептором в клетке -
Свободный Са2+ связывается со специфическим белком: кальциевым рецептором в клетке -
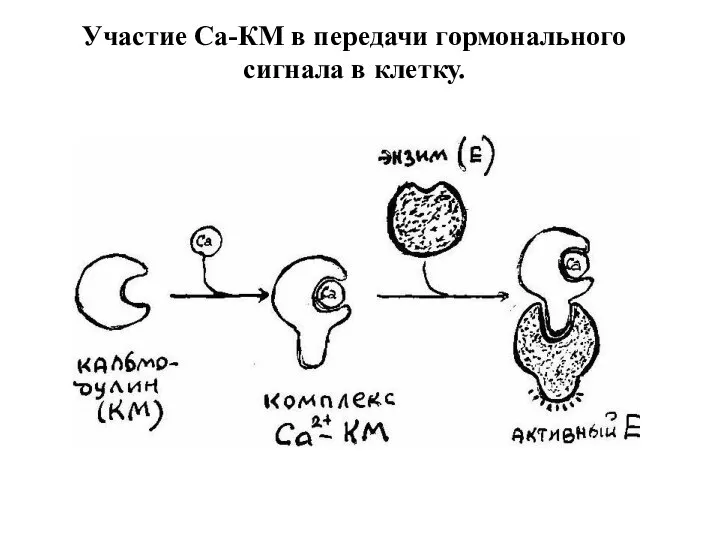
Участие Са-КМ в передачи гормонального сигнала в клетку.
Участие Са-КМ в передачи гормонального сигнала в клетку.

Диацилглицерид (ДАГ) так же участвует в механизме передачи гормонального сигнала в
Диацилглицерид (ДАГ) так же участвует в механизме передачи гормонального сигнала в

Гормоны с цитозольным механизмом действия:
● Стероидные гормоны
● Кальцитриол
● Гидрофобные гормоны
Гормоны с цитозольным механизмом действия:
● Стероидные гормоны
● Кальцитриол
● Гидрофобные гормоны
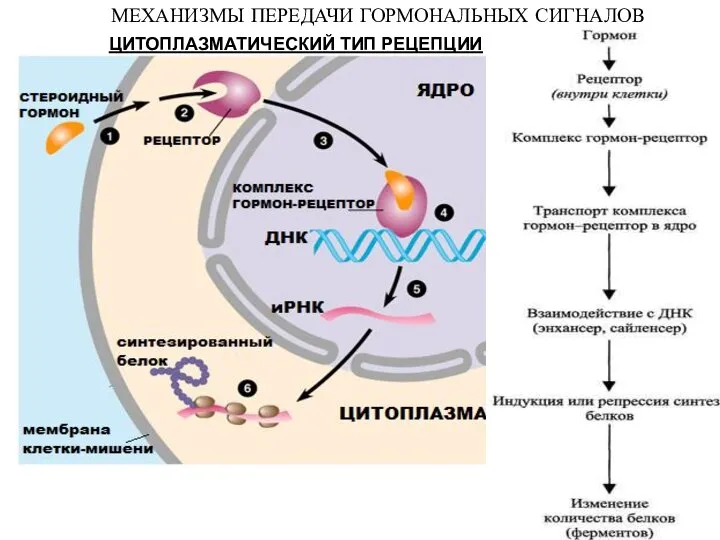
ЦИТОПЛАЗМАТИЧЕСКИЙ ТИП РЕЦЕПЦИИ
МЕХАНИЗМЫ ПЕРЕДАЧИ ГОРМОНАЛЬНЫХ СИГНАЛОВ
ЦИТОПЛАЗМАТИЧЕСКИЙ ТИП РЕЦЕПЦИИ
МЕХАНИЗМЫ ПЕРЕДАЧИ ГОРМОНАЛЬНЫХ СИГНАЛОВ
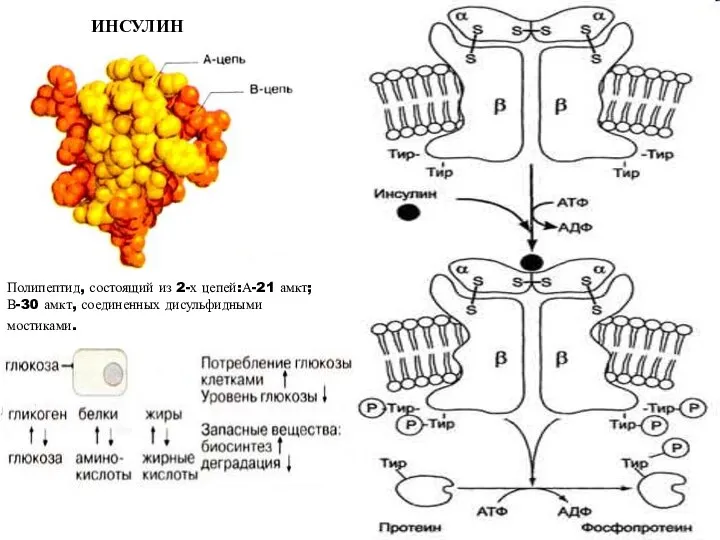
ИНСУЛИН
Полипептид, состоящий из 2-х цепей:А-21 амкт; В-30 амкт, соединенных дисульфидными мостиками.
ИНСУЛИН
Полипептид, состоящий из 2-х цепей:А-21 амкт; В-30 амкт, соединенных дисульфидными мостиками.

Гормональные эффекты
Быстрые эффекты (с-мин.) – регуляция метаболизма и функций (метаботропные, сопряженные
Гормональные эффекты
Быстрые эффекты (с-мин.) – регуляция метаболизма и функций (метаботропные, сопряженные
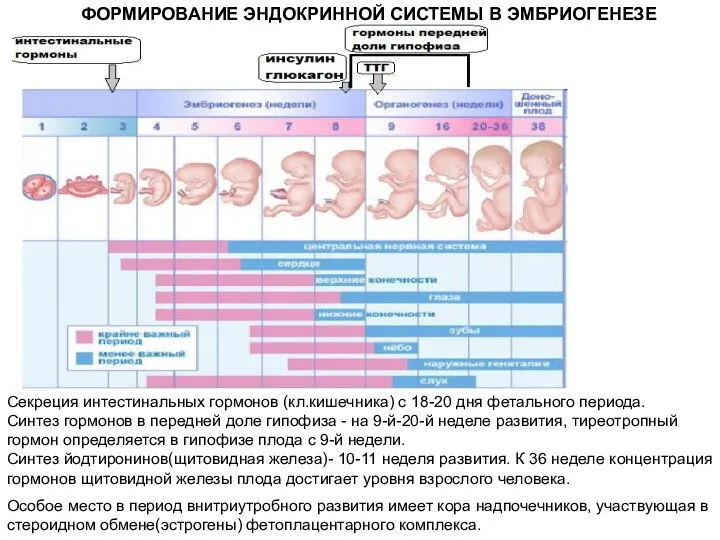
ФОРМИРОВАНИЕ ЭНДОКРИННОЙ СИСТЕМЫ В ЭМБРИОГЕНЕЗЕ
Секреция интестинальных гормонов (кл.кишечника) с 18-20 дня
ФОРМИРОВАНИЕ ЭНДОКРИННОЙ СИСТЕМЫ В ЭМБРИОГЕНЕЗЕ
Секреция интестинальных гормонов (кл.кишечника) с 18-20 дня
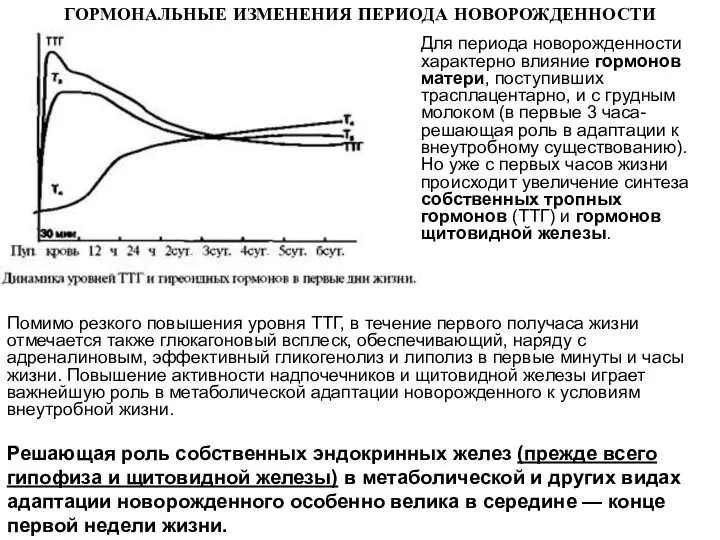
Для периода новорожденности характерно влияние гормонов матери, поступивших трасплацентарно, и с
Для периода новорожденности характерно влияние гормонов матери, поступивших трасплацентарно, и с
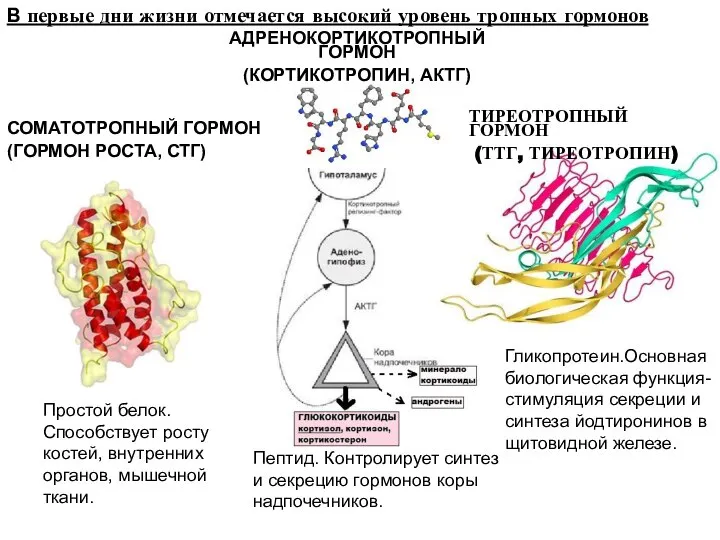
В первые дни жизни отмечается высокий уровень тропных гормонов
СОМАТОТРОПНЫЙ ГОРМОН
(ГОРМОН
В первые дни жизни отмечается высокий уровень тропных гормонов
СОМАТОТРОПНЫЙ ГОРМОН
(ГОРМОН
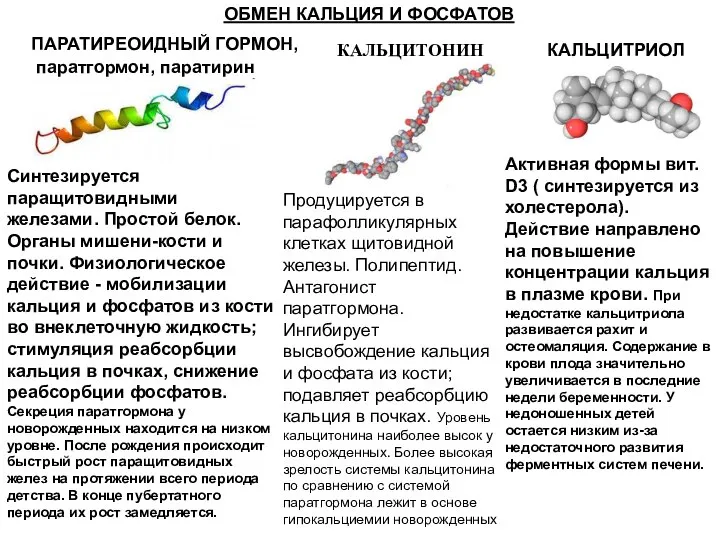
ОБМЕН КАЛЬЦИЯ И ФОСФАТОВ
ПАРАТИРЕОИДНЫЙ ГОРМОН,
паратгормон, паратирин
Синтезируется паращитовидными железами. Простой белок.
ОБМЕН КАЛЬЦИЯ И ФОСФАТОВ
ПАРАТИРЕОИДНЫЙ ГОРМОН,
паратгормон, паратирин
Синтезируется паращитовидными железами. Простой белок.

Роберт Лефковиц Брайан Кобилка
(Robert Lefkowitz) (Brian Kobilka)
из Стэнфорда
Роберт Лефковиц Брайан Кобилка
(Robert Lefkowitz) (Brian Kobilka)
из Стэнфорда

1) Рецептор (Са2+-мобилизующий рецептор)
2) G-белок;
3) Фосфотидилинозитол – источник свободного инозитолфосфата (ФИФ,
1) Рецептор (Са2+-мобилизующий рецептор)
2) G-белок;
3) Фосфотидилинозитол – источник свободного инозитолфосфата (ФИФ,
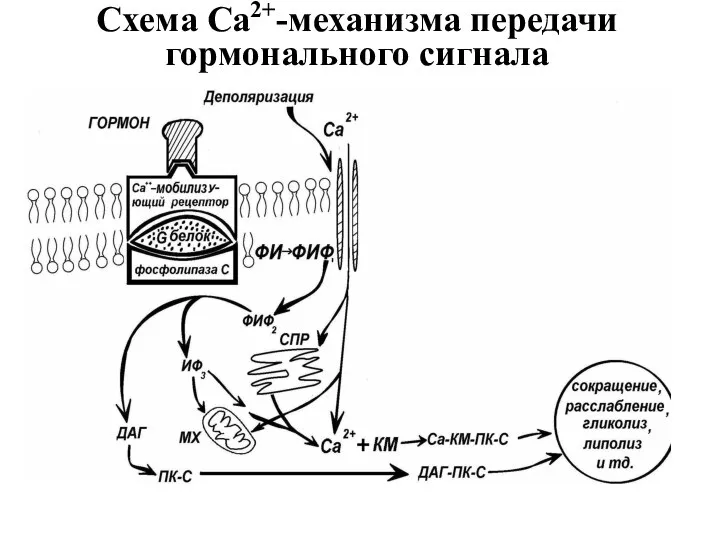
Схема Са2+-механизма передачи гормонального сигнала
Схема Са2+-механизма передачи гормонального сигнала

Свободный Са2+ связывается со специфическим белком: кальциевым рецептором в клетке -
Свободный Са2+ связывается со специфическим белком: кальциевым рецептором в клетке -
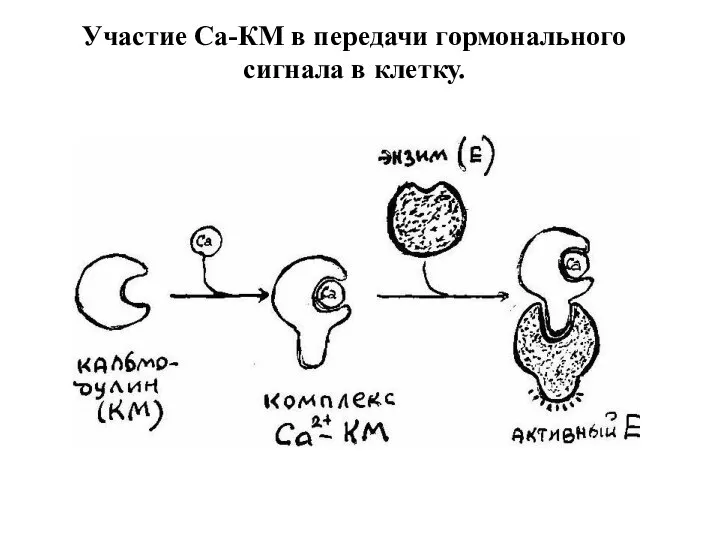
Участие Са-КМ в передачи гормонального сигнала в клетку.
Участие Са-КМ в передачи гормонального сигнала в клетку.
 Економічна теорія як наука
Економічна теорія як наука Презентация "Культура СССР начала ХХ века" - скачать презентации по МХК
Презентация "Культура СССР начала ХХ века" - скачать презентации по МХК Мочевыделительная система
Мочевыделительная система Программирование игр на Python
Программирование игр на Python Управление взаимоотношениями с клиентами
Управление взаимоотношениями с клиентами День Благодарения
День Благодарения Артриты и артрозы ВНЧС Выполнила: студентка 5 курса Группы 090716
Артриты и артрозы ВНЧС Выполнила: студентка 5 курса Группы 090716 Сердечная недостаточность
Сердечная недостаточность  ЭКЗИСТЕНЦИАЛЬНО-ГУМАНИСТИЧЕСКИЙ ПОДХОД В ПСИХОЛОГИИ И ПСИХОТЕРАПИИ
ЭКЗИСТЕНЦИАЛЬНО-ГУМАНИСТИЧЕСКИЙ ПОДХОД В ПСИХОЛОГИИ И ПСИХОТЕРАПИИ Жанерке слайд
Жанерке слайд Kinetika2
Kinetika2 Золотые травы Хохломы. «Чудо-ложка»
Золотые травы Хохломы. «Чудо-ложка» Оказание услуг в сфере декорирования джемперов
Оказание услуг в сфере декорирования джемперов Презентация по профессиональной стратификации
Презентация по профессиональной стратификации La arquitectura del Renacimiento (de Lan)
La arquitectura del Renacimiento (de Lan) Школа для развития своего таланта. Проект
Школа для развития своего таланта. Проект Даши Намдаков
Даши Намдаков Das schmeckt gut
Das schmeckt gut Адвокаттың кәсіби әдебі
Адвокаттың кәсіби әдебі О ходе модернизации системы подготовки спортивного резерва в Санкт-Петербурге
О ходе модернизации системы подготовки спортивного резерва в Санкт-Петербурге Поняття та види виборчих систем
Поняття та види виборчих систем УСТРОЙСТВО И ПРИНЦИП РАБОТЫ ЭВМ
УСТРОЙСТВО И ПРИНЦИП РАБОТЫ ЭВМ Лисичка-сестричка и волк
Лисичка-сестричка и волк  Обслуживание и ремонт двигателей постоянного тока
Обслуживание и ремонт двигателей постоянного тока Терминал. Планирование продаж
Терминал. Планирование продаж Современные технологии программирования. λ-выражения в Java 8. Функция как параметр
Современные технологии программирования. λ-выражения в Java 8. Функция как параметр МЕЖПРЕДМЕТНАЯ ИНТЕГРАЦИЯ В ШКОЛЬНОМ КУРСЕ ДИСЦИПЛИН ЕСТЕСТВЕННОГО ЦИКЛА НА ПРИМЕРЕ ПРЕПОДАВАНИЯ БИОЛОГИИ В ШКОЛЕ Аитова Фарид
МЕЖПРЕДМЕТНАЯ ИНТЕГРАЦИЯ В ШКОЛЬНОМ КУРСЕ ДИСЦИПЛИН ЕСТЕСТВЕННОГО ЦИКЛА НА ПРИМЕРЕ ПРЕПОДАВАНИЯ БИОЛОГИИ В ШКОЛЕ Аитова Фарид Филимоновская игрушка (средняя группа)
Филимоновская игрушка (средняя группа)