- дыхательная цепь

Содержание
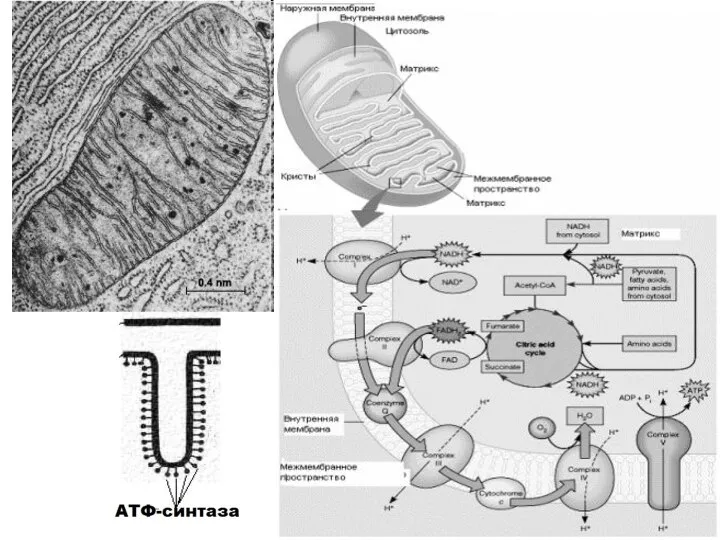
- 2. Дыхательная электронтранспортная цепь— комплекс структурно и функционально связанных ферментов, который расположен во внутренней мембране митохондрий и
- 4. Митохондриальная цепь переноса электронов
- 5. I комплекс дыхательной цепи – НАДН-дегидрогеназа – ФМН-содержащий фермент. В комплекс входит также группа железо-серных белков.
- 6. II комплекс дых. цепи – ФАД-содержащий фермент -сукцинатдегидрогеназа Субстрат: сукцинат (янтарная кислота)
- 7. Убихинон, принимая электроны и протоны, переходит в восстановленное состояние (КоQН2) УБИХИНОН (Коэнзим Q)
- 8. После КоQ в дыхательной цепи расположена группа цитохромов – гем-содержащих ферментов, обеспечивающих транспорт электронов. Различают несколько
- 9. III комплекс дыхательной цепи – цитохромС-редуктаза. В состав комплекса входит цитохромы В + С1, а так
- 10. Цитохром С
- 11. IV комплекс дыхательной цепи – цитохромоксидаза (ЦХО). Цитохромоксидаза представляет комплекс цитохрома А + А3, содержащий в
- 12. Изменение свободной энергии при переносе электронов по ЦПЭ. E-FMN - комплекс I; E-FAD - комплекс II;
- 13. 3Х7,3=21,9 ккал =3АТФ ΔG°’=52,6 ккал
- 16. Синтез АТФ – окислительное фосфорилирование - представляет процесс одновременного потребления О2, АДФ и Фн. В пределах
- 17. Функции дыхательной цепи: ●Транспорт электронов на О2 – тканевое дыхание. ●Преобразование энергии транспорта электронов в энергию
- 18. Синтез АТФ происходит при участии фермента- АТФ-синтетазы (протонная АТФаза) АТФ- СИНТЕТАЗА
- 19. Коэффициент фосфорилирования (Р/О)- количество молекул неорганического фосфата, которое включается в АТФ в расчете на 1 атом
- 20. Регуляция тканевого дыхания (транспорта электронов): ● Регуляция за счет концентрации О2 ●Дыхательный контроль (зависимость митохондриального окисления
- 21. Некоторые представители ингибиторов транспорта электронов
- 22. Регуляция синтеза АТФ: ● Регуляция протонного потока за счет разобщителей: - Протонофоры (динитрофенол, жирные кислоты, тироксин,
- 23. -субстраты цикла Кребса, -отношение [ATP]/[ADP] (обратимость АТФ-синтетазной реакции), -кислород (гипоксия), -физиологические разобщители (жирные кислоты, лизофосфолипиды, гормоны,
- 24. Особенности энергетического обмена у ребенка обусловлены его интенсивным ростом, биосинтетической деятельностью, функциональной незрелостью ряда регуляторных систем
- 25. Митохондриальный геном
- 26. Общая характеристика митохондриальных болезней Митохондриальные болезни – это мультисистемные заболевания, причиной или главным элементом патогенеза которых
- 27. Органы и системы, повреждаемые при митохондриальных болезнях Нервная система припадки, тремор, за- держка развития, глу- хота,
- 28. Первичные (врожденные) митохондриальные болезни могут быть обусловлены мутациями либо в ядерном, либо в митохондриальном геноме. Ядерные
- 31. Скачать презентацию

Дыхательная электронтранспортная цепь— комплекс структурно и функционально связанных ферментов, который расположен
Дыхательная электронтранспортная цепь— комплекс структурно и функционально связанных ферментов, который расположен
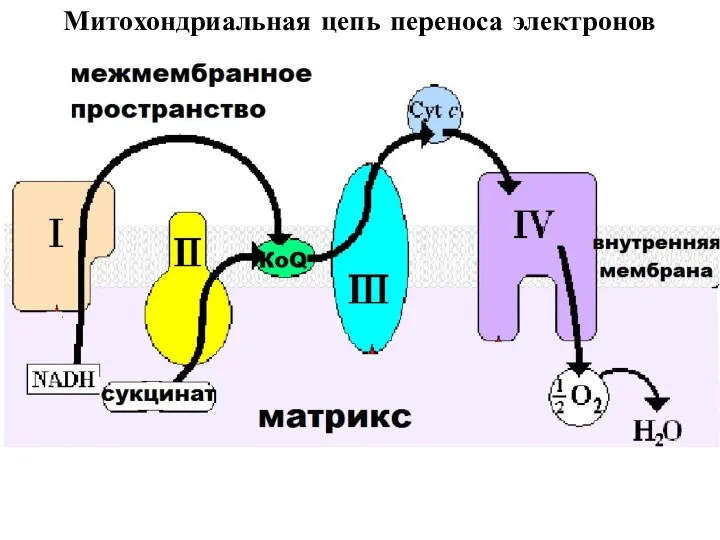
Митохондриальная цепь переноса электронов
Митохондриальная цепь переноса электронов
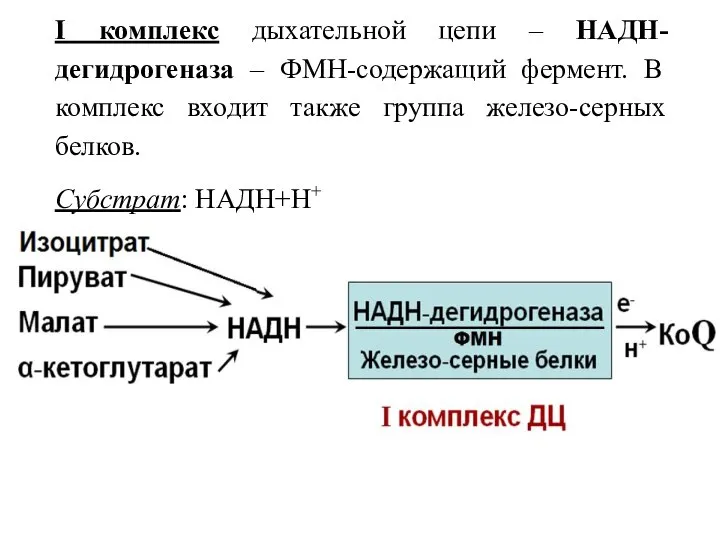
I комплекс дыхательной цепи – НАДН-дегидрогеназа – ФМН-содержащий фермент. В комплекс
I комплекс дыхательной цепи – НАДН-дегидрогеназа – ФМН-содержащий фермент. В комплекс
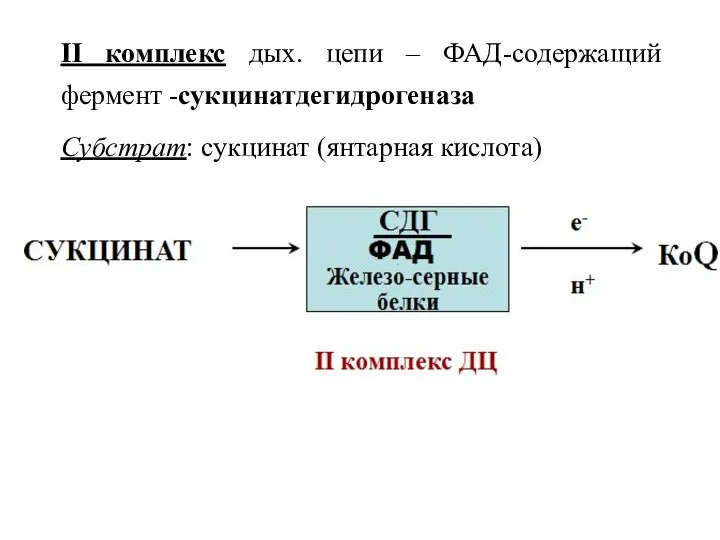
II комплекс дых. цепи – ФАД-содержащий фермент -сукцинатдегидрогеназа
Субстрат: сукцинат (янтарная кислота)
II комплекс дых. цепи – ФАД-содержащий фермент -сукцинатдегидрогеназа
Субстрат: сукцинат (янтарная кислота)
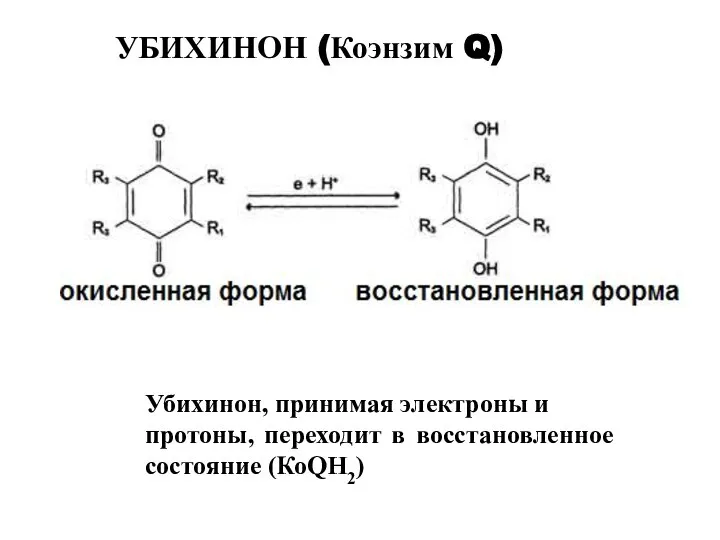
Убихинон, принимая электроны и
протоны, переходит в восстановленное состояние (КоQН2)
УБИХИНОН (Коэнзим Q)
Убихинон, принимая электроны и
протоны, переходит в восстановленное состояние (КоQН2)
УБИХИНОН (Коэнзим Q)

После КоQ в дыхательной цепи расположена группа цитохромов – гем-содержащих ферментов,
После КоQ в дыхательной цепи расположена группа цитохромов – гем-содержащих ферментов,
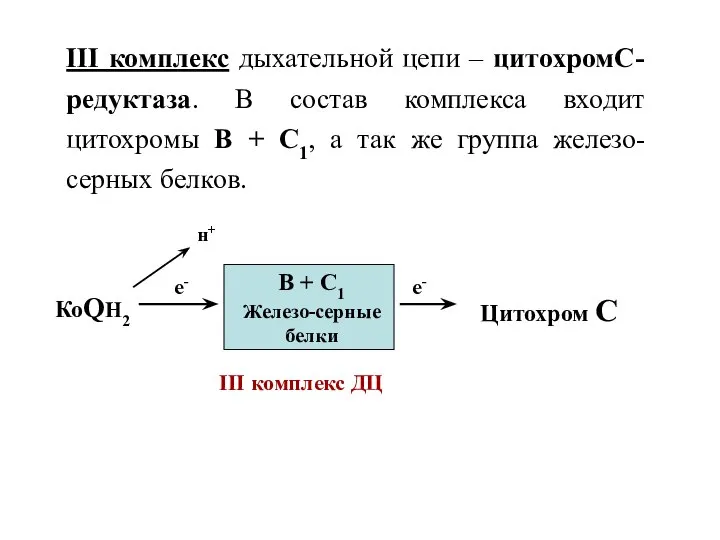
III комплекс дыхательной цепи – цитохромС-редуктаза. В состав комплекса входит цитохромы
III комплекс дыхательной цепи – цитохромС-редуктаза. В состав комплекса входит цитохромы
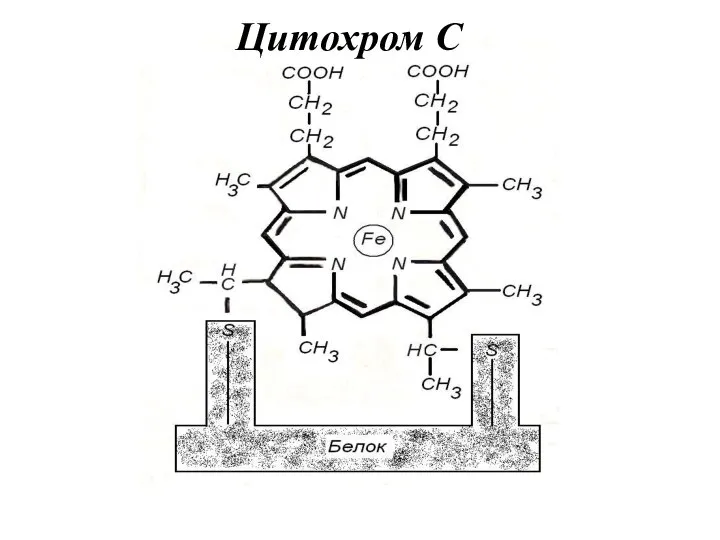
Цитохром С
Цитохром С
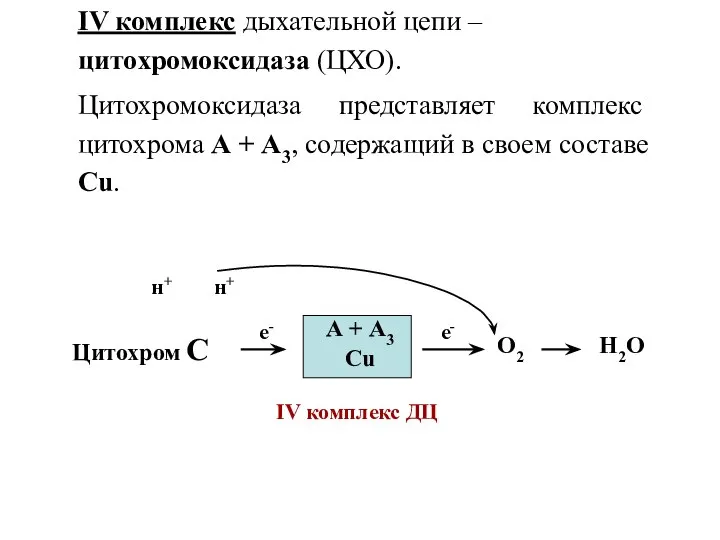
IV комплекс дыхательной цепи – цитохромоксидаза (ЦХО).
Цитохромоксидаза представляет комплекс цитохрома А
IV комплекс дыхательной цепи – цитохромоксидаза (ЦХО).
Цитохромоксидаза представляет комплекс цитохрома А
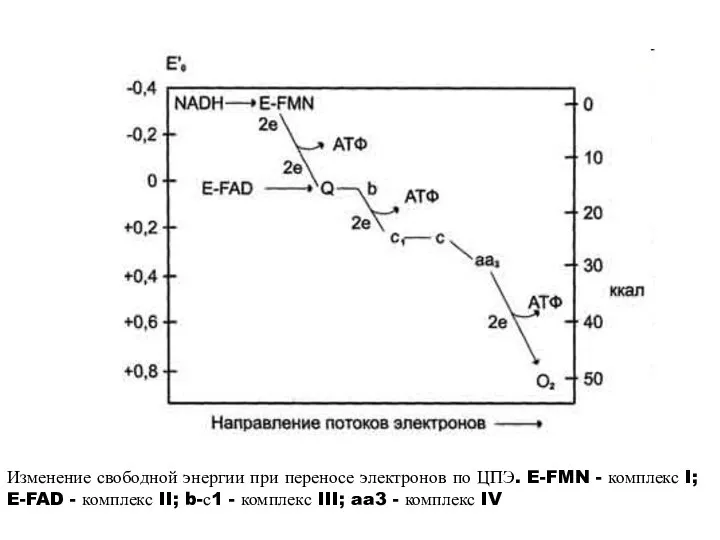
Изменение свободной энергии при переносе электронов по ЦПЭ. E-FMN - комплекс
Изменение свободной энергии при переносе электронов по ЦПЭ. E-FMN - комплекс
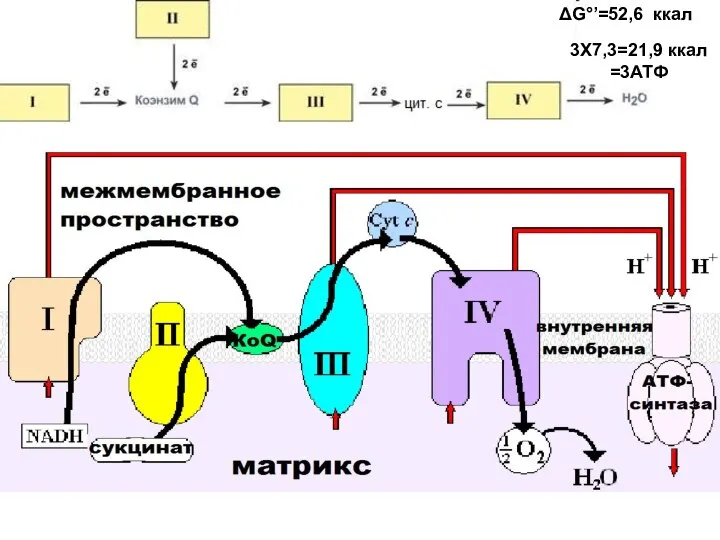
3Х7,3=21,9 ккал =3АТФ
ΔG°’=52,6 ккал
3Х7,3=21,9 ккал =3АТФ
ΔG°’=52,6 ккал
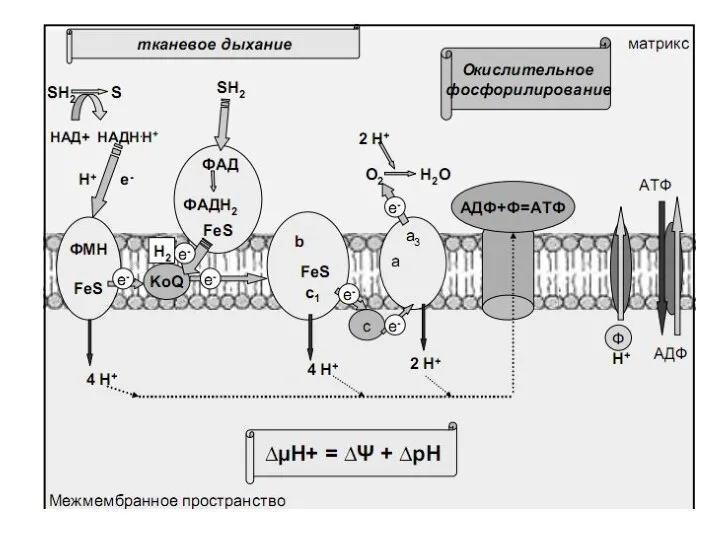


Синтез АТФ – окислительное фосфорилирование - представляет процесс одновременного потребления О2,
Синтез АТФ – окислительное фосфорилирование - представляет процесс одновременного потребления О2,

Функции дыхательной цепи:
●Транспорт электронов на О2 – тканевое дыхание.
●Преобразование энергии транспорта
Функции дыхательной цепи:
●Транспорт электронов на О2 – тканевое дыхание.
●Преобразование энергии транспорта
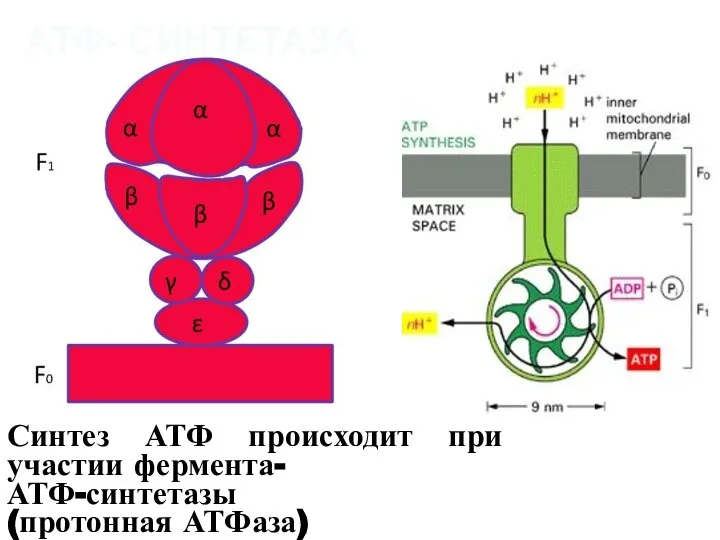
Синтез АТФ происходит при участии фермента-
АТФ-синтетазы
(протонная АТФаза)
АТФ- СИНТЕТАЗА
Синтез АТФ происходит при участии фермента-
АТФ-синтетазы
(протонная АТФаза)
АТФ- СИНТЕТАЗА

Коэффициент фосфорилирования (Р/О)- количество молекул неорганического фосфата, которое включается в АТФ
Коэффициент фосфорилирования (Р/О)- количество молекул неорганического фосфата, которое включается в АТФ

Регуляция тканевого дыхания (транспорта электронов):
● Регуляция за счет концентрации О2
●Дыхательный контроль
Регуляция тканевого дыхания (транспорта электронов):
● Регуляция за счет концентрации О2
●Дыхательный контроль
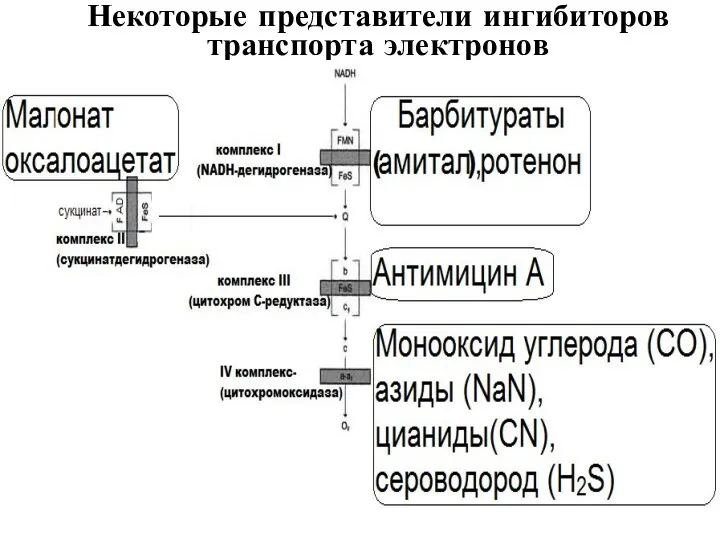
Некоторые представители ингибиторов транспорта электронов
Некоторые представители ингибиторов транспорта электронов

Регуляция синтеза АТФ:
● Регуляция протонного потока за счет разобщителей:
- Протонофоры (динитрофенол,
Регуляция синтеза АТФ:
● Регуляция протонного потока за счет разобщителей:
- Протонофоры (динитрофенол,
![-субстраты цикла Кребса, -отношение [ATP]/[ADP] (обратимость АТФ-синтетазной реакции), -кислород (гипоксия), -физиологические](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1298299/slide-22.jpg)
-субстраты цикла Кребса,
-отношение [ATP]/[ADP] (обратимость АТФ-синтетазной реакции),
-кислород (гипоксия),
-физиологические разобщители (жирные кислоты,
-субстраты цикла Кребса,
-отношение [ATP]/[ADP] (обратимость АТФ-синтетазной реакции),
-кислород (гипоксия),
-физиологические разобщители (жирные кислоты,

Особенности энергетического обмена у ребенка обусловлены его интенсивным ростом, биосинтетической деятельностью,
Особенности энергетического обмена у ребенка обусловлены его интенсивным ростом, биосинтетической деятельностью,
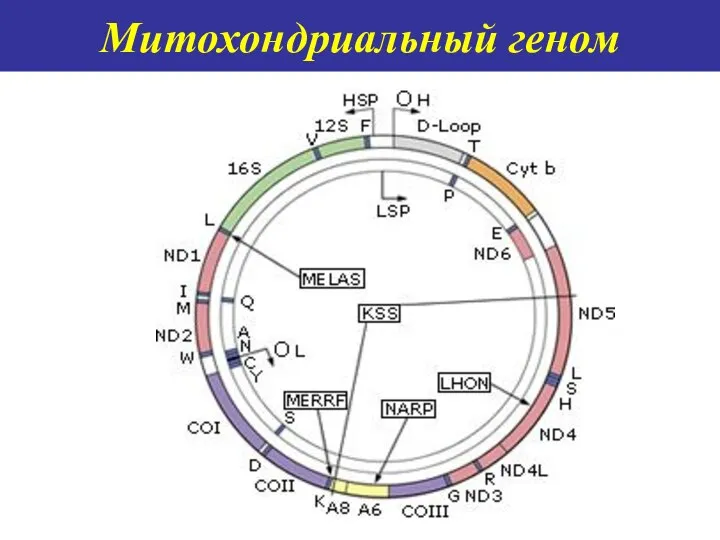
Митохондриальный геном
Митохондриальный геном

Общая характеристика митохондриальных болезней
Митохондриальные болезни – это мультисистемные заболевания, причиной
Общая характеристика митохондриальных болезней
Митохондриальные болезни – это мультисистемные заболевания, причиной
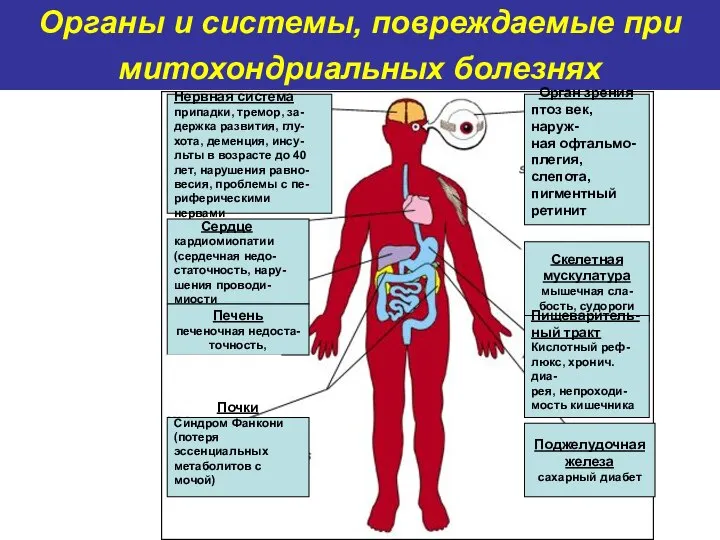
Органы и системы, повреждаемые при митохондриальных болезнях
Нервная система
припадки, тремор, за-
держка
Органы и системы, повреждаемые при митохондриальных болезнях
Нервная система
припадки, тремор, за-
держка

Первичные (врожденные) митохондриальные болезни могут быть обусловлены мутациями либо в
Первичные (врожденные) митохондриальные болезни могут быть обусловлены мутациями либо в

 Презентация "Скульптуры разных эпох и стилей" - скачать презентации по МХК
Презентация "Скульптуры разных эпох и стилей" - скачать презентации по МХК Гимнастическая терминология
Гимнастическая терминология Как я провёл Лето
Как я провёл Лето Проект фильма "Шалаш"
Проект фильма "Шалаш" Мастер-класс. Решение задач по разделу «Механика»
Мастер-класс. Решение задач по разделу «Механика» Спорт - Дело Тонкое. СпецВыпуск
Спорт - Дело Тонкое. СпецВыпуск Врубель Михаил Александрович ЖИЗНЬ И ТВОРЧЕСТВО ВЕЛИКОГО РУССКОГО ХУДОЖНИКА.
Врубель Михаил Александрович ЖИЗНЬ И ТВОРЧЕСТВО ВЕЛИКОГО РУССКОГО ХУДОЖНИКА.  Бог никогда не спит
Бог никогда не спит Интерьер-студия Felicita. Уют Вашего дома!
Интерьер-студия Felicita. Уют Вашего дома! Политология как наука и учебная дисциплина
Политология как наука и учебная дисциплина Наскільки важлива культура зовнішнього вигляду вчителя
Наскільки важлива культура зовнішнього вигляду вчителя МЕТОД ПРОЕКТОВ В ОБРАЗОВАНИИ "Метод проектов - это не алгоритм, состоящий из четких этапов, а модель творческого мышления и п
МЕТОД ПРОЕКТОВ В ОБРАЗОВАНИИ "Метод проектов - это не алгоритм, состоящий из четких этапов, а модель творческого мышления и п Средний восток в международных отношениях после окончания холодной войны
Средний восток в международных отношениях после окончания холодной войны Олимпийские медали
Олимпийские медали хип-хоп
хип-хоп Строительство и усиление военных дорог
Строительство и усиление военных дорог Сервисный тренинг EXD06 3,0l-V6-TDI-CR-двигатель в Phaeton
Сервисный тренинг EXD06 3,0l-V6-TDI-CR-двигатель в Phaeton The 19th amendment: the equal rights
The 19th amendment: the equal rights Устрій та бойове застосування КЗА 86Ж6. Методика розрахунку змінних величин. Запис змінних величин. (Тема 11.3)
Устрій та бойове застосування КЗА 86Ж6. Методика розрахунку змінних величин. Запис змінних величин. (Тема 11.3) Soudage, qu'est-ce que ce
Soudage, qu'est-ce que ce ВКР: Основные характеристики технологии PON
ВКР: Основные характеристики технологии PON Напряжения. Связь внутренних усилий и напряжений
Напряжения. Связь внутренних усилий и напряжений Презентация "Филимоновская игрушка" - скачать презентации по МХК
Презентация "Филимоновская игрушка" - скачать презентации по МХК Русская народная игрушка
Русская народная игрушка Аттестационная работа. Метод проектов на занятиях инженерной графикой
Аттестационная работа. Метод проектов на занятиях инженерной графикой Технология разработки программного обеспечения
Технология разработки программного обеспечения Принципы радиосвязи. (8 класс)
Принципы радиосвязи. (8 класс) Реализм в русской литературе 19 века
Реализм в русской литературе 19 века