- гормоны аггрессия

Содержание
- 2. Рис.1 Упрощенная схема синтеза стероидных гормонов
- 3. CYP зависимый метаболизм эстрадиола Е2.
- 4. Рис.2 Механизмы аггрессии под действием стероидных гормонов (А) гонадный тестостерон действует прямо на мозг (В) гонадный
- 5. Рис.3.Уровни DHEA, Т, Е2 (нг/мл) в плазме птиц в период не спаривания
- 6. Рис.4 Эффект введения DHEA на поведение самцов птиц в период не спаривания
- 7. Рис.5 (А) 3-бетта-гидростероид изомераза катализирует окисление и изомеризацию DHEA в андростенедион (АЕ); (В) эстроген попадает в
- 8. Рис.6 (А)Синтез гонадных стероидов и агрессивность (В) у самок и самцов морской свинки в результате «короткой
- 9. Рис.7 (А) Введение экзогенного мелатонина увеличивает количество атак самцов морской свинки. Эффект исчезает у самцов после
- 11. Скачать презентацию
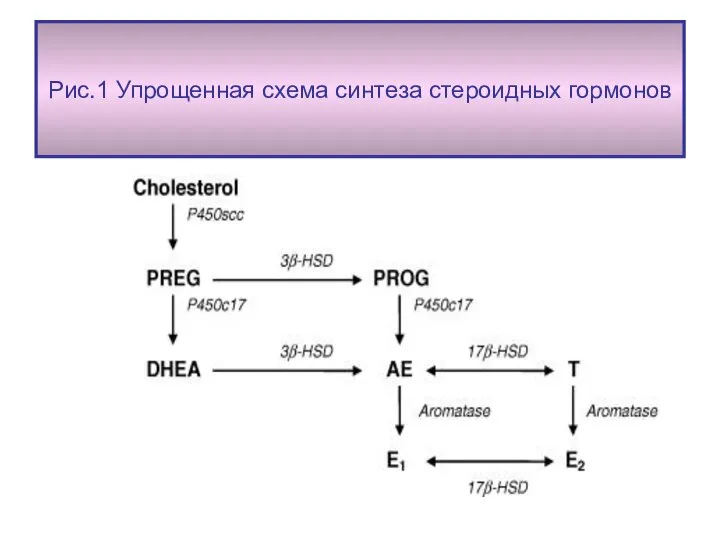
Рис.1 Упрощенная схема синтеза стероидных гормонов
Рис.1 Упрощенная схема синтеза стероидных гормонов
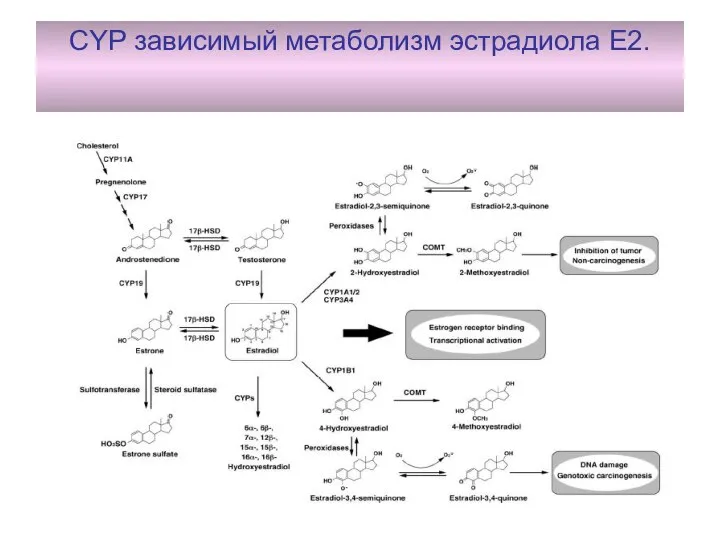
CYP зависимый метаболизм эстрадиола Е2.
CYP зависимый метаболизм эстрадиола Е2.
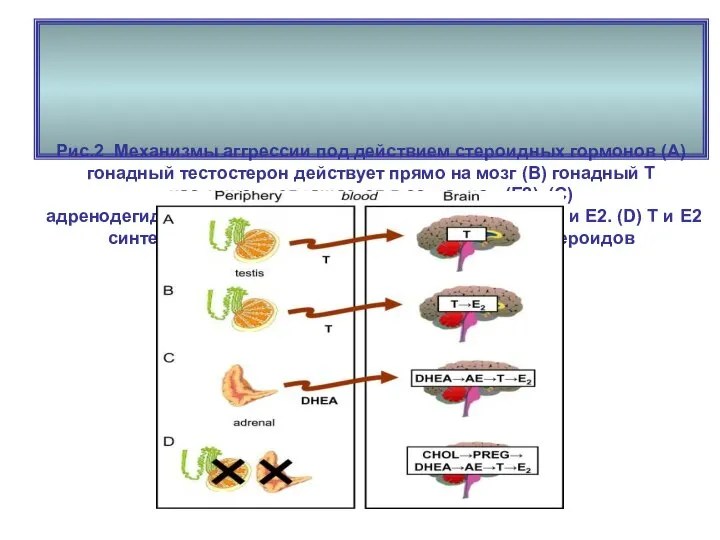
Рис.2 Механизмы аггрессии под действием стероидных гормонов (А) гонадный тестостерон действует
Рис.2 Механизмы аггрессии под действием стероидных гормонов (А) гонадный тестостерон действует
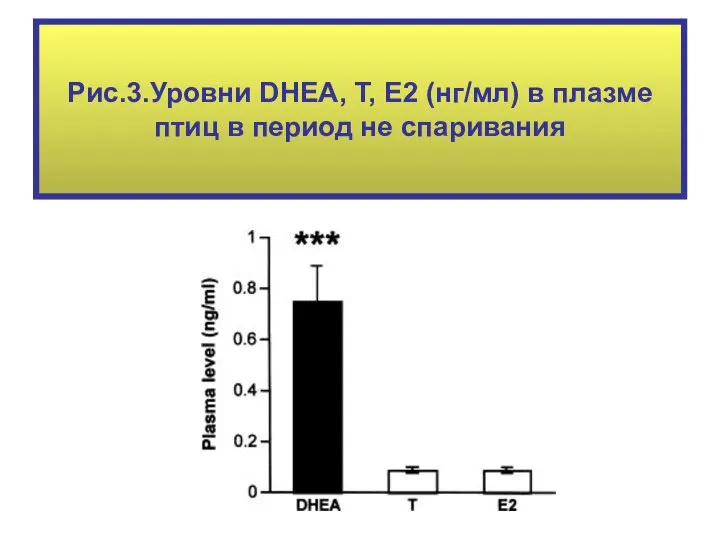
Рис.3.Уровни DHEA, Т, Е2 (нг/мл) в плазме птиц в период не
Рис.3.Уровни DHEA, Т, Е2 (нг/мл) в плазме птиц в период не
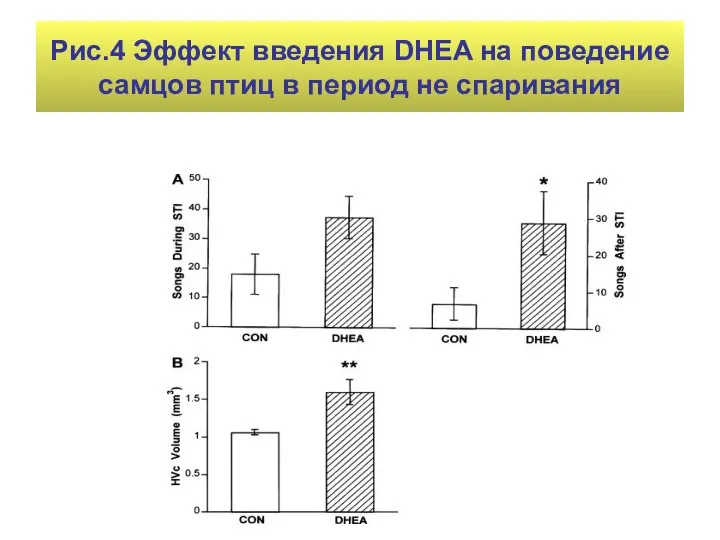
Рис.4 Эффект введения DHEA на поведение самцов птиц в период не
Рис.4 Эффект введения DHEA на поведение самцов птиц в период не
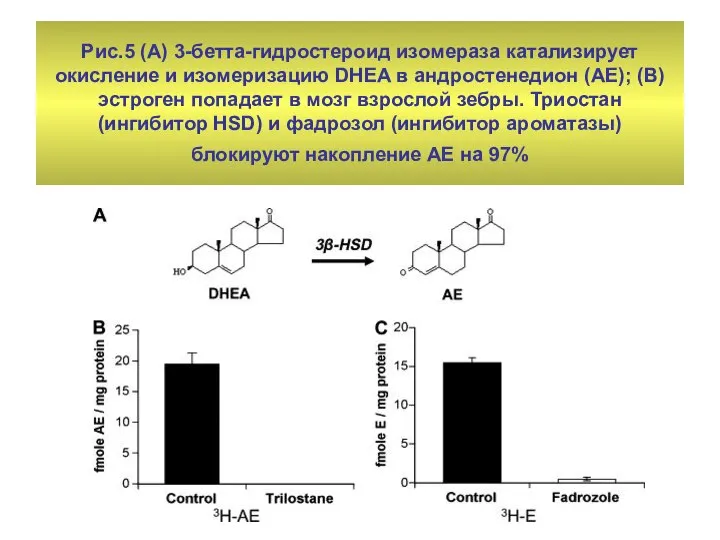
Рис.5 (А) 3-бетта-гидростероид изомераза катализирует окисление и изомеризацию DHEA в андростенедион
Рис.5 (А) 3-бетта-гидростероид изомераза катализирует окисление и изомеризацию DHEA в андростенедион
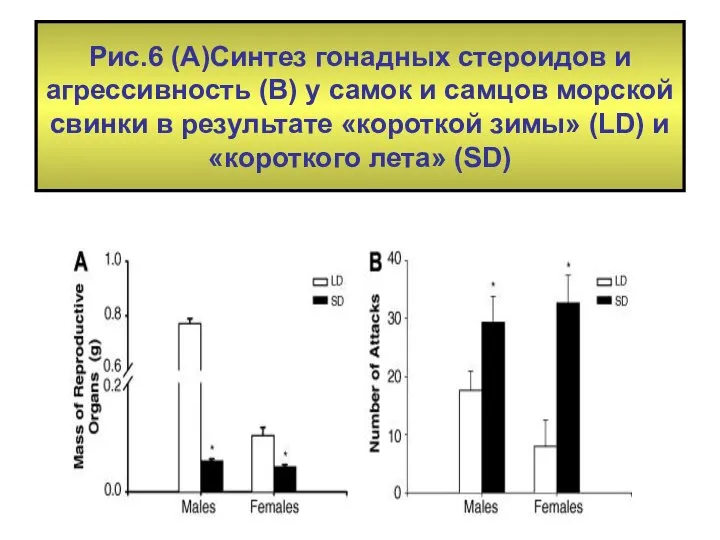
Рис.6 (А)Синтез гонадных стероидов и агрессивность (В) у самок и самцов
Рис.6 (А)Синтез гонадных стероидов и агрессивность (В) у самок и самцов
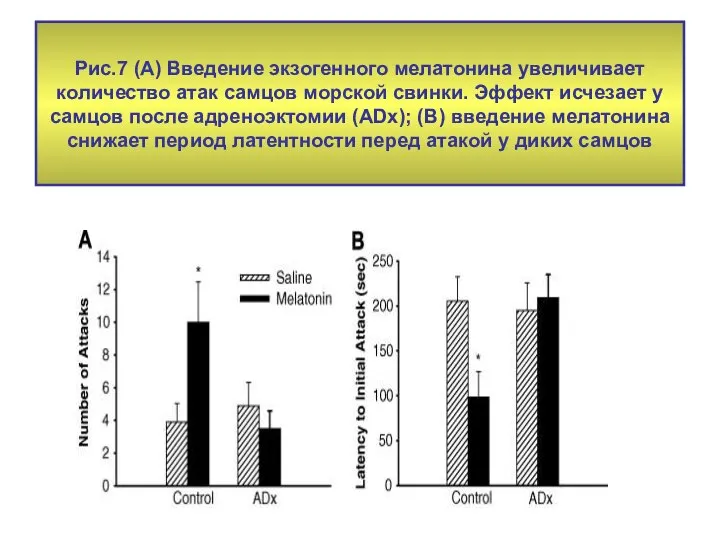
Рис.7 (А) Введение экзогенного мелатонина увеличивает количество атак самцов морской свинки.
Рис.7 (А) Введение экзогенного мелатонина увеличивает количество атак самцов морской свинки.
 Разрезы. Тест
Разрезы. Тест игра и игрушка как психолого-пед. инструмент
игра и игрушка как психолого-пед. инструмент  Илья Ефимович Репин (1844-1930 гг.)
Илья Ефимович Репин (1844-1930 гг.) Тхэквондо. Белый пояс
Тхэквондо. Белый пояс Презентация Коллективные переговоры и порядок их проведения
Презентация Коллективные переговоры и порядок их проведения Рассчитать технико-экономические показатели сборочно-сварочного участка по производству изделия
Рассчитать технико-экономические показатели сборочно-сварочного участка по производству изделия Азбука без букв (правила дорожного движения) Учитель 1-4 классов МБОУ «Кугесьский лицей» Чебоксарского района Чувашской Республи
Азбука без букв (правила дорожного движения) Учитель 1-4 классов МБОУ «Кугесьский лицей» Чебоксарского района Чувашской Республи Проект федерального закона «О внесении изменений и дополнений в ФЗ «О несостоятельности (банкротстве)» в части банкротства финанс
Проект федерального закона «О внесении изменений и дополнений в ФЗ «О несостоятельности (банкротстве)» в части банкротства финанс Государственный строй древнего периода
Государственный строй древнего периода Ця важка робота
Ця важка робота Философия Канта
Философия Канта Презентация на тему Сказка Колобок Мультфильм
Презентация на тему Сказка Колобок Мультфильм  Технология спирта ректификата
Технология спирта ректификата Грамматикалық формалар мен грамматикалық мағаналар
Грамматикалық формалар мен грамматикалық мағаналар Способы активизации познавательной деятельности учащихся на уроках черчения
Способы активизации познавательной деятельности учащихся на уроках черчения Авиаперевозки предложение о сотрудничестве
Авиаперевозки предложение о сотрудничестве Культура стран арабского халифата
Культура стран арабского халифата Самостоятельность как основа развития личности
Самостоятельность как основа развития личности Матч Украина-Латвия. Механизмы игры в футбол
Матч Украина-Латвия. Механизмы игры в футбол Наноэлектроника: устройства, цепи, архитектура
Наноэлектроника: устройства, цепи, архитектура Латинский язык. Занятие 3
Латинский язык. Занятие 3 Презентация "Баланс и счет бухгалтерского учета" - скачать презентации по Экономике
Презентация "Баланс и счет бухгалтерского учета" - скачать презентации по Экономике Оператор присваивания и процедуры ввода-вывода
Оператор присваивания и процедуры ввода-вывода Процедуры и функции, создаваемые пользователем. Лабораторная работа №11
Процедуры и функции, создаваемые пользователем. Лабораторная работа №11 МИРОВАЯ ХУДОЖЕСТВЕННАЯ КУЛЬТУРА Культура Двуречья (шумерская и ассиро-вавилонская)
МИРОВАЯ ХУДОЖЕСТВЕННАЯ КУЛЬТУРА Культура Двуречья (шумерская и ассиро-вавилонская) Математическая статистика в жизни класса
Математическая статистика в жизни класса Как заработать на арбитраже
Как заработать на арбитраже Презентация на тему "Компетенции заместителя директора школы" - скачать презентации по Педагогике
Презентация на тему "Компетенции заместителя директора школы" - скачать презентации по Педагогике