- Мышечная ткань

Содержание
- 2. Мышечными тканями называ-ются ткани, различные по про-исхождению, но сходные по способности к выраженным сокращениям. Они обеспечи-вают
- 3. Общая морфофункциональная характеристика мышечной ткани Удлинённая форма клеток Наличие миофибрилл и миофиламентов Расположение митохондрий рядом с
- 4. Классификация Морфофункциональная: - гладкая мышечная ткань - поперечно-полосатая: скелетная сердечная
- 5. Классификация Гистогенетическая: - мезенхимные (ГМТ) - эпидермальные (миоэпителиальные клетки) - нейральные (мышца суживающая и расширяющая зрачок)
- 6. Поперечнополосатая скелетная мышечная ткань Гистогенез – вначале образуются миобласты миотома. Клетки сливаются, образуя длинные симпласты. Одновременно
- 7. Строение ППМТ Структурная единица – мышечное волокно (мион), состоящее из миосимпласта и миосателлитоцитов, покрытое базальной мембраной.
- 8. Поперечнополосатая скелетная мышечная ткань
- 9. Строение миосимпласта Ядра в МС расположены под оболочкой волокна, их количество достигает 10000. Органеллы общего значения
- 10. Сократительный аппарат попе-речно-полосатой мышечной ткани Состоит из миофибрилл Каждая миофибрилла имеет поперечные тёмные и светлые диски
- 11. Структура скелетной мышцы
- 13. Границей саркомера является телофрагма (построена из a-акти-нина ), которая на препарате выглядит в виде Z линии
- 14. К мезофрагме прикреплены толстые нити сократительного белка миозина К телофрагме – тонкие нити белка актина Концы
- 17. Различают тёмный участок – диск А (анизотропный, соответствует длине миозиновой нити) И светлый диск – диск
- 19. Молекула миозина имеет длинный хвост и две головки. При повышении концентрации ионов Са молекула изменяет свою
- 20. Z- линии сближаются – саркомер укорачивается по длине (до 2-х раз) Альфа - актиновые сети Z-
- 21. Источником ионов Са служат цистерны АЭПС, они вытянуты вдоль каждого саркомера. На границе А- и I-
- 22. Саркоплазматическая сеть
- 23. Когда симпласт получает сигнал о начале сокращения, он перемещается по плаз-молемме в виде потенциала действия и
- 24. Гликоген – необходим для сокращения и поддержания теплового баланса Миоглобин – связывает кислород, когда мышца расслаблена.
- 25. Миосателлитоциты Малодифференцированные клетки, участвующие в регенерации мышечной ткани. Они находятся на поверхности симпласта и окружены общей
- 26. Типы мышечных волокон В связи с различными функциональ-ными условиями миофибриллы, мито-хондрии и миоглобин по-разному рас-пределяются в
- 27. Типы мышечных волокон
- 28. Регенерация скелетной мышечной ткани После повреждения мышечного волокна в участке травмы разрушенные фрагменты фагоцитируются макрофагами. В
- 29. Перерыв! Осторожно, яблоки в буфете могут быть с «червячком»!
- 30. Скелетная мышца как орган Со стороны сухожилий или надкостницы, к которым мышца прикреплена, прони-кают тонкие коллагеновые
- 31. Кровоснабжение Сосуды распространяются в прослойках соединительной ткани В перимизии находятся артериолы В эндомизии – капилляры Рядом
- 32. Кровоснабжение
- 33. Иннервация В мышцах присутствуют - эфферентные (миелиновые) - афферентные (безмиелиновые) - вегетативные нервные волокна (безмиелиновые) Отросток
- 34. Моторная бляшка
- 35. Группа мышечных волокон, иннервиру-емая одним мотонейроном, называется нервно-мышечная единица. Чувствительные нервные окончания свя-заны со специализированными мышечны-ми
- 36. Нервно-мышечное веретено
- 37. Интрафузальные специализиро-ванные мышечные волокна бывают двух типов: - волокна с ядерной сумкой - волокна с ядерной
- 38. При функционировании рабочих мышечных волокон изменяется натяжение соединительнотканной капсулы веретена. Возбуждаются чувствительные нервные окончания, возникают афферентные
- 39. Гладкая мышечная ткань Гистогенетические источники: - мезенхимные - эпидермальные - нейральные ------------------------------------------------------ ГМТ представлена в стенках
- 40. Гладкая мышечная ткань мезенхимного происхождения Структурная единица ГМТ – веретеновидная клетка (20-500 мкм) Ядро находится в
- 41. Сократительный аппарат предста-влен миофибриллами, филаменты актина которых образуют в цито-плазме трёхмерную сеть. В местах соединения с
- 42. Гладкая мышечная ткань
- 43. Сигнал для сокращения поступает по нервным волокнам. Медиаторы, выделяемые из терминалей, изме-няют состояние плазмолеммы. Она образует
- 44. Актиновые нити смещаются нав-стречу друг другу, усилия переда-ются на плазмолемму, вся клетка укорачивается. Когда поступление сигналов
- 45. Сердечная мышечная ткань Гистогенетический источник – миоэпикардиальные пластинки висцерального листка спланхнотома в шейной части зародыша. Структурной
- 46. Виды кардиомиоцитов: Рабочие (сократительные) Синусные (пейсмекерные = водители ритма сердца) Переходные Проводящие Секреторные.
- 47. Сократительные клетки имеют удли-нённую форму (до 150 мкм), их концы соединяются друг с другом, образуя функциональные
- 48. Сердечная мышечная ткань
- 49. Кардиомиоциты
- 50. Ядра КМЦ (1-2) овальной формы, распо-ложены в центре клетки. У полюсов ядра сосредоточены органел-лы общего значения.
- 51. Петли ЭПС вытянуты вдоль повер-хности миофибрилл и имеют лате-ральные утолщения (L -система), формирующие вместе с Т
- 52. Синусные (пейсмекерные) КМЦ способны генерировать импульс со скоростью около 70 в мин. Переходные КМЦ получают сигнал
- 53. Вставочные диски сердечной ткани образованы - интердигитациями - десмосомами К каждой десмосоме подходит мио-фибрилла, закрепляющаяся концом
- 54. Регенерация СМТ Стволовых клеток в сердечной мы-шечной ткани нет, поэтому погиба-ющие КМЦ не восстанавливаются. После инфаркта
- 55. Мышечная ткань эпидермального происхождения Миоэпителиальные клетки встреча-ются в потовых, молочных, слюнных и др. железах. Их называют
- 56. Мышечная ткань нейрального происхождения Миоциты этой ткани развиваются из клеток нейрального зачатка в составе внутренней стенки
- 57. Спасибо за внимание!
- 59. Скачать презентацию

Мышечными тканями называ-ются ткани, различные по про-исхождению, но сходные по способности
Мышечными тканями называ-ются ткани, различные по про-исхождению, но сходные по способности

Общая морфофункциональная характеристика мышечной ткани
Удлинённая форма клеток
Наличие миофибрилл и миофиламентов
Расположение митохондрий
Общая морфофункциональная характеристика мышечной ткани
Удлинённая форма клеток
Наличие миофибрилл и миофиламентов
Расположение митохондрий

Классификация
Морфофункциональная:
- гладкая мышечная ткань
- поперечно-полосатая:
скелетная
сердечная
Классификация
Морфофункциональная:
- гладкая мышечная ткань
- поперечно-полосатая:
скелетная
сердечная

Классификация
Гистогенетическая:
- мезенхимные (ГМТ)
- эпидермальные (миоэпителиальные клетки)
- нейральные (мышца
Классификация
Гистогенетическая:
- мезенхимные (ГМТ)
- эпидермальные (миоэпителиальные клетки)
- нейральные (мышца

Поперечнополосатая скелетная мышечная ткань
Гистогенез – вначале образуются миобласты миотома. Клетки сливаются,
Поперечнополосатая скелетная мышечная ткань
Гистогенез – вначале образуются миобласты миотома. Клетки сливаются,

Строение ППМТ
Структурная единица – мышечное волокно (мион), состоящее из миосимпласта и
Строение ППМТ
Структурная единица – мышечное волокно (мион), состоящее из миосимпласта и
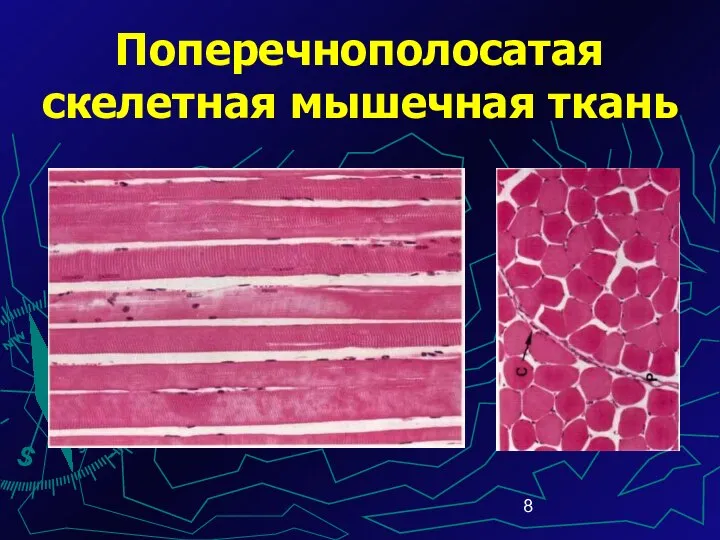
Поперечнополосатая скелетная мышечная ткань
Поперечнополосатая скелетная мышечная ткань

Строение миосимпласта
Ядра в МС расположены под оболочкой волокна, их количество достигает
Строение миосимпласта
Ядра в МС расположены под оболочкой волокна, их количество достигает

Сократительный аппарат попе-речно-полосатой мышечной ткани
Состоит из миофибрилл
Каждая миофибрилла имеет поперечные тёмные
Сократительный аппарат попе-речно-полосатой мышечной ткани
Состоит из миофибрилл
Каждая миофибрилла имеет поперечные тёмные
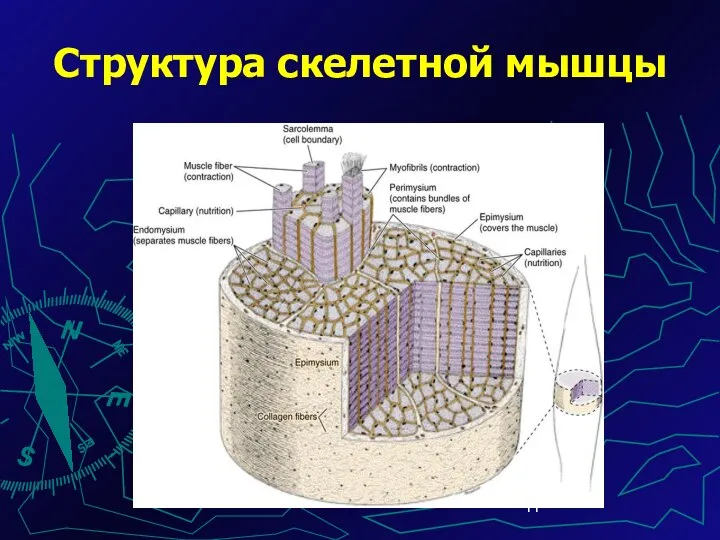
Структура скелетной мышцы
Структура скелетной мышцы


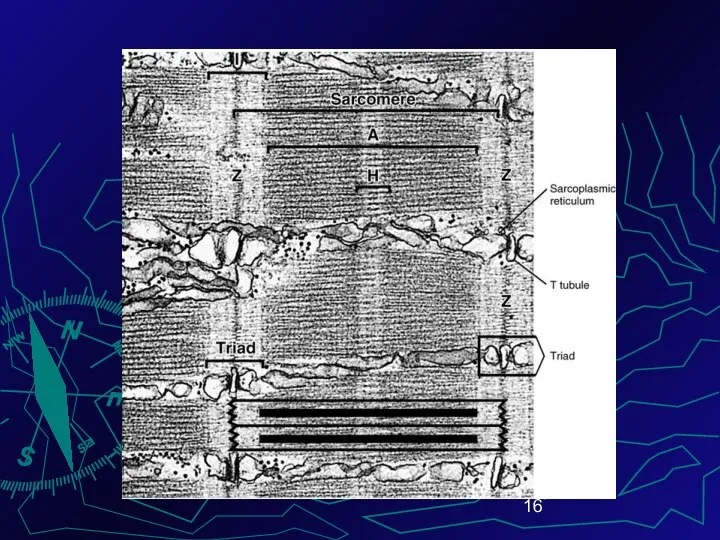
Границей саркомера является телофрагма (построена из a-акти-нина ), которая на препарате
Границей саркомера является телофрагма (построена из a-акти-нина ), которая на препарате

К мезофрагме прикреплены толстые нити сократительного белка миозина
К телофрагме – тонкие
К мезофрагме прикреплены толстые нити сократительного белка миозина
К телофрагме – тонкие


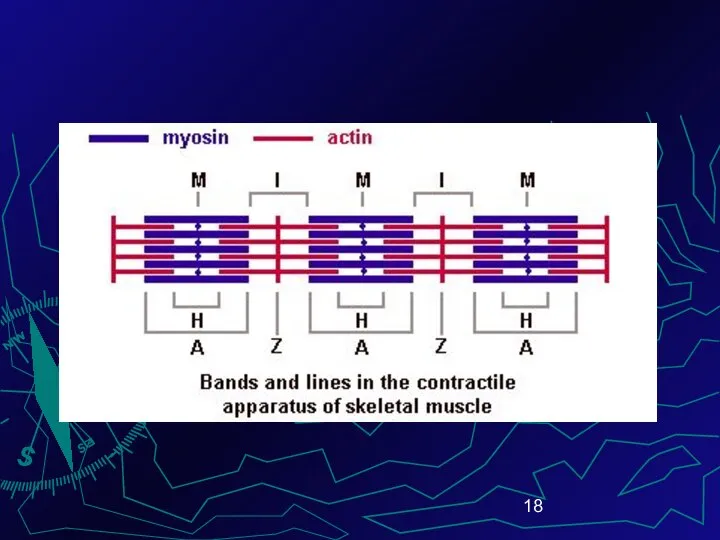
Различают тёмный участок – диск А (анизотропный, соответствует длине миозиновой нити)
И
Различают тёмный участок – диск А (анизотропный, соответствует длине миозиновой нити)
И

Молекула миозина имеет длинный хвост и две головки.
При повышении концентрации ионов
Молекула миозина имеет длинный хвост и две головки.
При повышении концентрации ионов

Z- линии сближаются – саркомер укорачивается по длине (до 2-х раз)
Z- линии сближаются – саркомер укорачивается по длине (до 2-х раз)

Источником ионов Са служат цистерны АЭПС, они вытянуты вдоль каждого саркомера.
Источником ионов Са служат цистерны АЭПС, они вытянуты вдоль каждого саркомера.
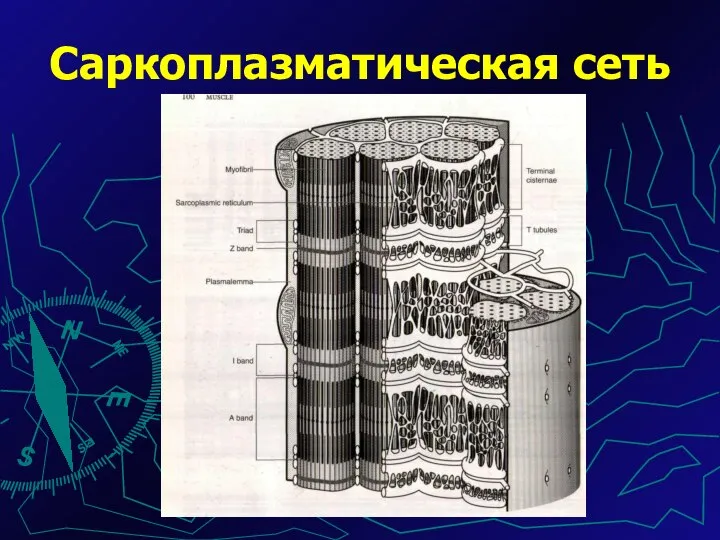
Саркоплазматическая сеть
Саркоплазматическая сеть

Когда симпласт получает сигнал о начале сокращения, он перемещается по плаз-молемме
Когда симпласт получает сигнал о начале сокращения, он перемещается по плаз-молемме

Гликоген – необходим для сокращения и поддержания теплового баланса
Миоглобин – связывает
Гликоген – необходим для сокращения и поддержания теплового баланса
Миоглобин – связывает

Миосателлитоциты
Малодифференцированные клетки, участвующие в регенерации мышечной ткани.
Они находятся на поверхности симпласта
Миосателлитоциты
Малодифференцированные клетки, участвующие в регенерации мышечной ткани.
Они находятся на поверхности симпласта

Типы мышечных волокон
В связи с различными функциональ-ными условиями миофибриллы, мито-хондрии и
Типы мышечных волокон
В связи с различными функциональ-ными условиями миофибриллы, мито-хондрии и
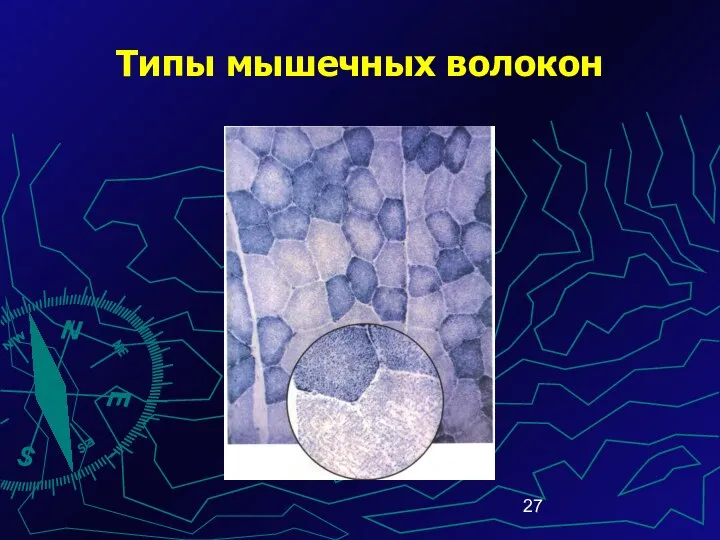
Типы мышечных волокон
Типы мышечных волокон

Регенерация скелетной мышечной ткани
После повреждения мышечного волокна в участке травмы разрушенные
Регенерация скелетной мышечной ткани
После повреждения мышечного волокна в участке травмы разрушенные

Перерыв!
Осторожно, яблоки в буфете могут быть с «червячком»!
Перерыв!
Осторожно, яблоки в буфете могут быть с «червячком»!

Скелетная мышца как орган
Со стороны сухожилий или надкостницы, к которым
Скелетная мышца как орган
Со стороны сухожилий или надкостницы, к которым

Кровоснабжение
Сосуды распространяются в прослойках соединительной ткани
В перимизии находятся артериолы
В эндомизии –
Кровоснабжение
Сосуды распространяются в прослойках соединительной ткани
В перимизии находятся артериолы
В эндомизии –
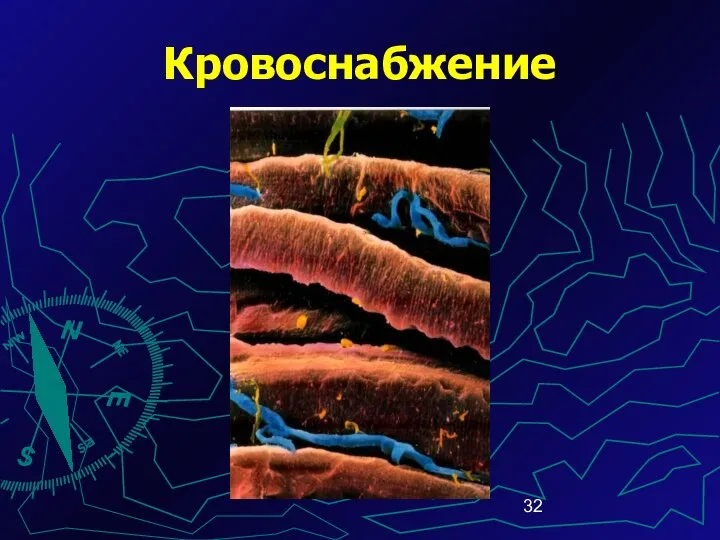
Кровоснабжение
Кровоснабжение

Иннервация
В мышцах присутствуют
- эфферентные (миелиновые)
- афферентные (безмиелиновые)
-
Иннервация
В мышцах присутствуют
- эфферентные (миелиновые)
- афферентные (безмиелиновые)
-
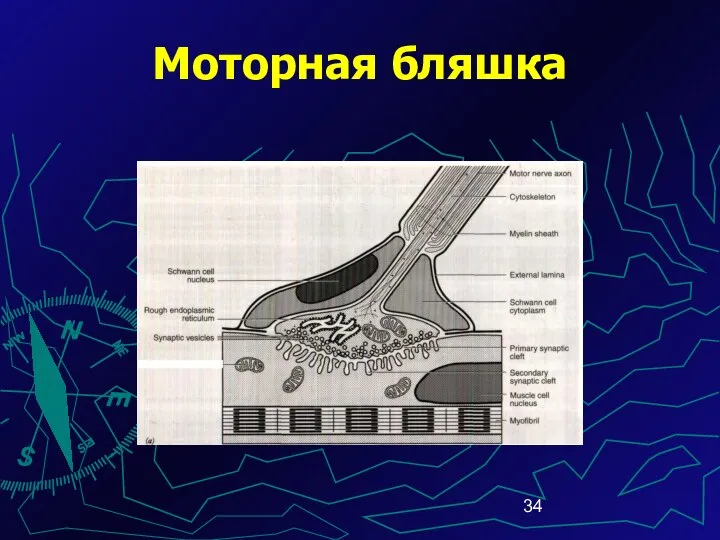
Моторная бляшка
Моторная бляшка

Группа мышечных волокон, иннервиру-емая одним мотонейроном, называется нервно-мышечная единица.
Чувствительные нервные окончания
Группа мышечных волокон, иннервиру-емая одним мотонейроном, называется нервно-мышечная единица.
Чувствительные нервные окончания
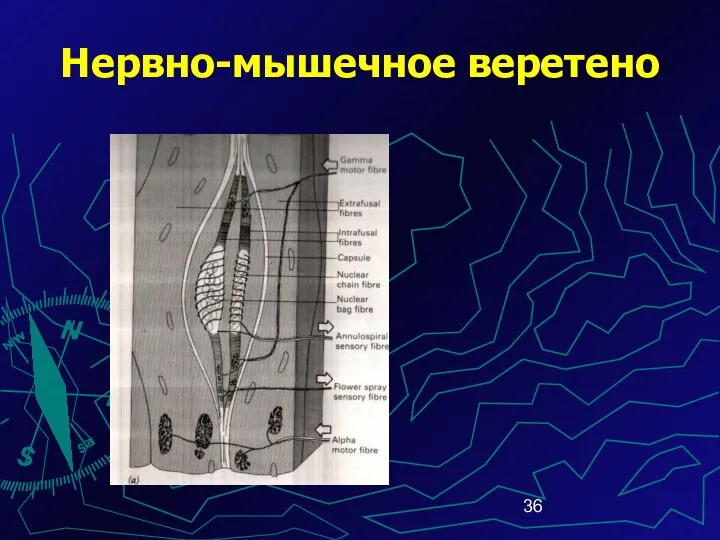
Нервно-мышечное веретено
Нервно-мышечное веретено

Интрафузальные специализиро-ванные мышечные волокна бывают двух типов:
- волокна с ядерной
Интрафузальные специализиро-ванные мышечные волокна бывают двух типов:
- волокна с ядерной

При функционировании рабочих мышечных волокон изменяется натяжение соединительнотканной капсулы веретена.
Возбуждаются чувствительные
При функционировании рабочих мышечных волокон изменяется натяжение соединительнотканной капсулы веретена.
Возбуждаются чувствительные

Гладкая мышечная ткань
Гистогенетические источники:
- мезенхимные
- эпидермальные
- нейральные
Гладкая мышечная ткань
Гистогенетические источники:
- мезенхимные
- эпидермальные
- нейральные
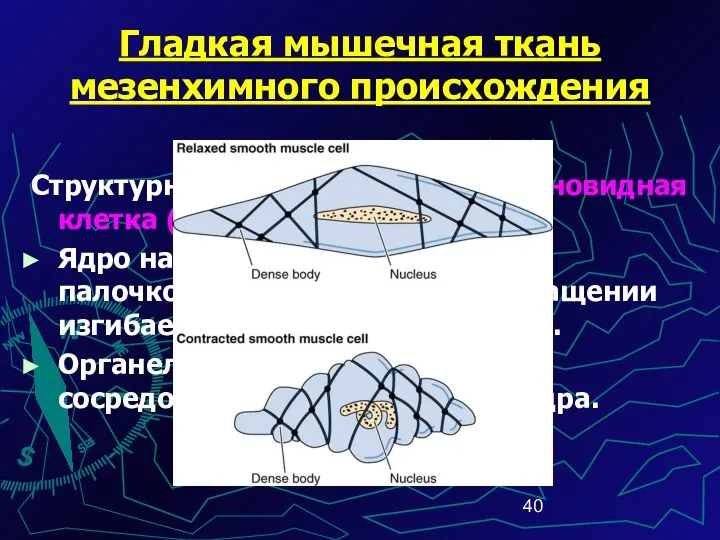
Гладкая мышечная ткань мезенхимного происхождения
Структурная единица ГМТ – веретеновидная клетка (20-500
Гладкая мышечная ткань мезенхимного происхождения
Структурная единица ГМТ – веретеновидная клетка (20-500

Сократительный аппарат предста-влен миофибриллами, филаменты актина которых образуют в цито-плазме трёхмерную
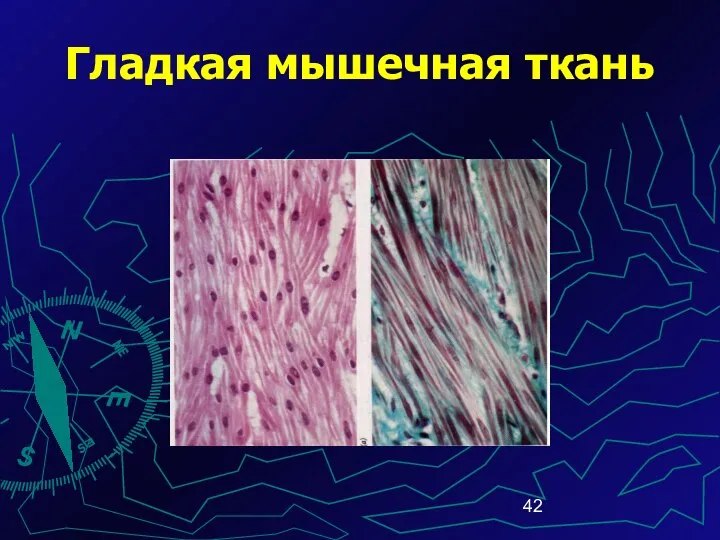
Гладкая мышечная ткань
Гладкая мышечная ткань

Сигнал для сокращения поступает по нервным волокнам. Медиаторы, выделяемые из терминалей,
Сигнал для сокращения поступает по нервным волокнам. Медиаторы, выделяемые из терминалей,

Актиновые нити смещаются нав-стречу друг другу, усилия переда-ются на плазмолемму, вся
Актиновые нити смещаются нав-стречу друг другу, усилия переда-ются на плазмолемму, вся

Сердечная мышечная ткань
Гистогенетический источник – миоэпикардиальные пластинки висцерального листка спланхнотома в
Сердечная мышечная ткань
Гистогенетический источник – миоэпикардиальные пластинки висцерального листка спланхнотома в

Виды кардиомиоцитов:
Рабочие (сократительные)
Синусные (пейсмекерные = водители ритма сердца)
Переходные
Проводящие
Секреторные.
Виды кардиомиоцитов:
Рабочие (сократительные)
Синусные (пейсмекерные = водители ритма сердца)
Переходные
Проводящие
Секреторные.

Сократительные клетки имеют удли-нённую форму (до 150 мкм), их концы соединяются
Сократительные клетки имеют удли-нённую форму (до 150 мкм), их концы соединяются
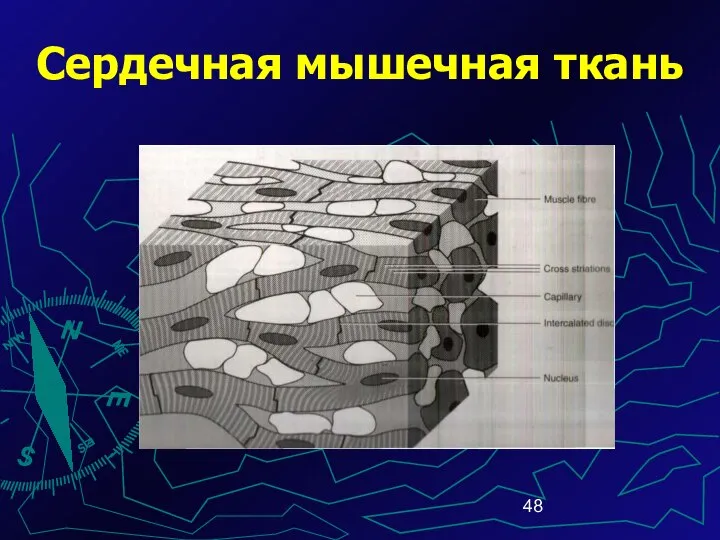
Сердечная мышечная ткань
Сердечная мышечная ткань
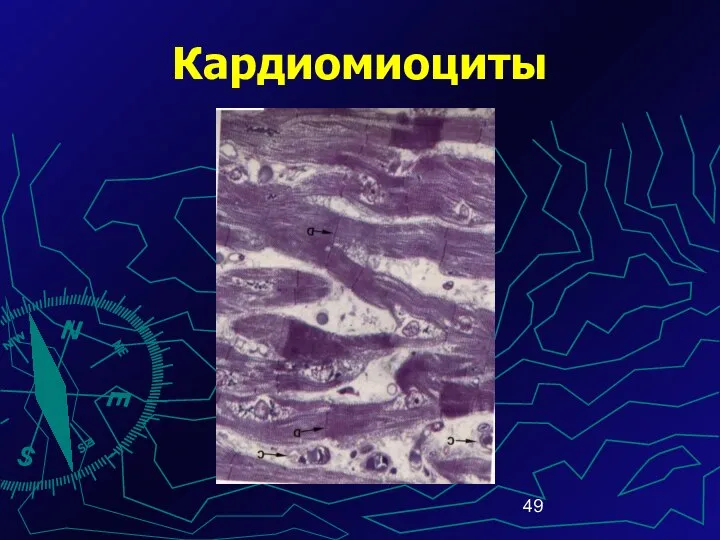
Кардиомиоциты
Кардиомиоциты

Ядра КМЦ (1-2) овальной формы, распо-ложены в центре клетки.
У полюсов ядра
Ядра КМЦ (1-2) овальной формы, распо-ложены в центре клетки.
У полюсов ядра

Петли ЭПС вытянуты вдоль повер-хности миофибрилл и имеют лате-ральные утолщения (L
Петли ЭПС вытянуты вдоль повер-хности миофибрилл и имеют лате-ральные утолщения (L

Синусные (пейсмекерные) КМЦ способны генерировать импульс со скоростью около 70 в
Синусные (пейсмекерные) КМЦ способны генерировать импульс со скоростью около 70 в

Вставочные диски сердечной ткани образованы
- интердигитациями
- десмосомами
К каждой
Вставочные диски сердечной ткани образованы
- интердигитациями
- десмосомами
К каждой

Регенерация СМТ
Стволовых клеток в сердечной мы-шечной ткани нет, поэтому погиба-ющие КМЦ
Регенерация СМТ
Стволовых клеток в сердечной мы-шечной ткани нет, поэтому погиба-ющие КМЦ

Мышечная ткань эпидермального происхождения
Миоэпителиальные клетки встреча-ются в потовых, молочных, слюнных и
Мышечная ткань эпидермального происхождения
Миоэпителиальные клетки встреча-ются в потовых, молочных, слюнных и

Мышечная ткань нейрального происхождения
Миоциты этой ткани развиваются из клеток нейрального зачатка
Мышечная ткань нейрального происхождения
Миоциты этой ткани развиваются из клеток нейрального зачатка

Спасибо за внимание!
Спасибо за внимание!
 Управління змістом робіт. Оцінювання тривалості та складності робіт
Управління змістом робіт. Оцінювання тривалості та складності робіт Юри на льду
Юри на льду Факты, тренды, технологии 05/04/2011 Санкт-Петербург Отель Сокос m.kisin@goodsmatrix.ru GoodsMatrix.ru Как платформа для продвижения потребительских то
Факты, тренды, технологии 05/04/2011 Санкт-Петербург Отель Сокос m.kisin@goodsmatrix.ru GoodsMatrix.ru Как платформа для продвижения потребительских то Контрольный тест. Электроника 6
Контрольный тест. Электроника 6 Судебный порядок защиты прав граждан в сфере государственного управления
Судебный порядок защиты прав граждан в сфере государственного управления Презентация Память и речь
Презентация Память и речь Петербургское Садовое кольцо. свой среди своих в курортном районе
Петербургское Садовое кольцо. свой среди своих в курортном районе Мотивация как функция менеджмента
Мотивация как функция менеджмента Технология «сухого» обогащения руд
Технология «сухого» обогащения руд Дом.ру. Продажа роутеров
Дом.ру. Продажа роутеров Роль религии в жизни общества
Роль религии в жизни общества Католицизм, или католичество
Католицизм, или католичество Optical access networks. Lecture 7
Optical access networks. Lecture 7 Олимпийские игры. Эстафета олимпийского огня
Олимпийские игры. Эстафета олимпийского огня сбытовая политика
сбытовая политика Общая электротехника. История развития электротехники
Общая электротехника. История развития электротехники Цель управления проектом и успешность проекта. SMART-критерии в целеполагании. (Тема 5)
Цель управления проектом и успешность проекта. SMART-критерии в целеполагании. (Тема 5) Общее учение о внутренностях
Общее учение о внутренностях  Особенности судебного разбирательства в суде с участием присяжных заседателей. (Тема 3.3)
Особенности судебного разбирательства в суде с участием присяжных заседателей. (Тема 3.3) ХУДОЖЕСТВЕННАЯ КУЛЬТУРА РОМАНТИЗМА: ЖИВОПИСЬ
ХУДОЖЕСТВЕННАЯ КУЛЬТУРА РОМАНТИЗМА: ЖИВОПИСЬ Как оформить выполнение работ, которые не входят в должностные обязанности
Как оформить выполнение работ, которые не входят в должностные обязанности ИЗДАНИЕ ДЛЯ АВТОМОБИЛИСТОВ auto.aviso.ua. РЕГУЛЯРНАЯ АУДИТОРИЯ в г. Киеве составляет 45,09 тыс. читателей, из которых по роду деятельности б
ИЗДАНИЕ ДЛЯ АВТОМОБИЛИСТОВ auto.aviso.ua. РЕГУЛЯРНАЯ АУДИТОРИЯ в г. Киеве составляет 45,09 тыс. читателей, из которых по роду деятельности б Информационные технологии в медицине
Информационные технологии в медицине Презентация "О чем поют колокола?" - скачать презентации по МХК
Презентация "О чем поют колокола?" - скачать презентации по МХК Делегирование и легитимность власти Подготовили: студентки 2 курса, ФТД, группы Т-114 Голофаст Н. и Зорина Д.
Делегирование и легитимность власти Подготовили: студентки 2 курса, ФТД, группы Т-114 Голофаст Н. и Зорина Д. Игрушка-дергунчик Дед Мороз
Игрушка-дергунчик Дед Мороз Основная образовательная программа высшего образования. Направление подготовки: менеджмент
Основная образовательная программа высшего образования. Направление подготовки: менеджмент ЖЕЛЧНОКАМЕННАЯ БОЛЕЗНЬ 7
ЖЕЛЧНОКАМЕННАЯ БОЛЕЗНЬ 7