- Нарушения структуры коллагена и их последствия

Содержание
- 2. Трансляция Это синтез белка на рибосомах. Матрицей является и РНК. Продукт трансляции – полипептидная цепь (первичная
- 3. Посттрансляционная модификация (ПТМ) Это и есть созревание белка после трансляции. В ходе ПТМ происходит внедрение в
- 4. Как правило, в ходе посттрансляционной модификации происходит частичное ОКИСЛЕНИЕ определенных аминокислот Часто к белку присоединяются углеводные
- 5. Условия успешной трансляции 1. Наличие в клетке малых и больших субъединиц рибосом 2. Поступление в цитоплазму
- 6. Закон «все или ничего» Если в клетке отсутствует хотя бы 1 незаменимая аминокислота, трансляция обрывается. Поэтому
- 7. Основные этапы трансляции 1. Инициация синтеза полипептидной цепи 2. Элонгация полипептидной цепи 3. Терминация синтеза полипептидной
- 8. Инициация трансляции Малая и большая субъединицы рибосомы в присутствии иона магния с затратой АТФ садятся на
- 9. Рибосома на этапе инициации
- 10. Элонгация Происходит удлинение полипептидной цепи на 1 аминокислоту, принесенную в рибосому новой молекулой тРНК За счет
- 11. Рибосома на этапе элонгации
- 12. Терминация В конце иРНК, перед полиаденилатным хвостом, всегда содержится «бессмысленный» кодон, не соответствующий ни одной тРНК.
- 13. Антибиотики и трансляция Многие «классические» антибиотики ингибируют один из этапов трансляции Поскольку рибосомы человека и бактерий
- 14. Реконструкция рибосомы, иРНК и полипептидной цепи по данным атомной силовой микроскопии
- 15. Примеры антибиотиков – ингибиторов трансляции
- 16. Коллаген и его роль Коллаген – структурный белок, один из наиболее распространенных белков организма. Имеет фибриллярную
- 17. Виды соединительной ткани (световая микроскопия)
- 18. Особенности аминокислотного состава коллагена (первичная структура) высокое содержание глицина отсутствие триптофана около 21 % аминокислот приходится
- 19. Вторичная структура β-складчатый лист (подобие «гармошки») 3 такие нити закручиваются в 3α-спираль Эта структура называется ТРОПОКОЛЛАГЕН
- 20. 3α-спираль тропоколлагена
- 21. Свойства тропоколлагена Это незрелый коллаген Легко вымывается водой Формирует фибриллу коллагена в присутствии ионов кальция
- 22. Основные этапы созревания коллагена (ПТМ) 1. Отщепление сигнальных пептидов от первичной последовательности 2. Гидроксилирование остатков пролина
- 24. После гликозилирования тропоколлаген становится высокоантигенным белком, уникальным для каждого человека Одна из причин отторжения органов после
- 25. Формирование коллагеновой фибриллы Коллагеновая фибрилла — это полукристаллическая структурная единица коллагена Состоит из молекул тропоколлагена, прикрепленных
- 26. Формирование коллагеновой фибриллы В фибриллах молекулы тропоколлагена смещены относительно друг друга примерно на 67нм (единица, которая
- 27. Формирование коллагеновой фибриллы Для правильной укладки молекул тропоколлагена нужны ионы кальция При дефиците кальция тропоколлаген укладывается
- 28. Коллагеновые фибриллы в электронном микроскопе
- 29. Взаимодействие молекул тропоколлагена при формировании фибриллы
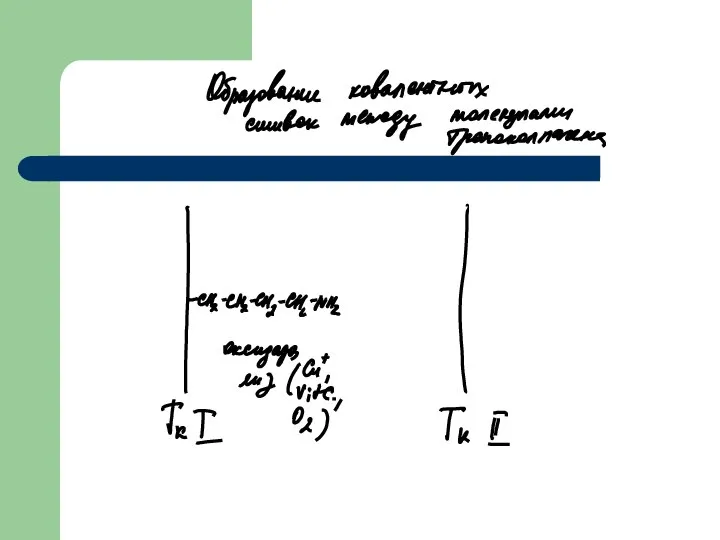
- 30. Придание коллагену прочностных свойств Внутри тропоколлагенов существует ковалентная связь между цепями Эти связи формируются между остатками
- 31. Нарушения синтеза коллагена Врожденные аномалии (дефицит гидроксилаз пролина и лизина) – синдром Эллерса-Данлоса Приобретенные аномалии –
- 32. Стоматологические проявления Ослабление связок зуба Расшатывание и самопроизвольное выпадение зубов Деструкция десневой ткани Разрушение тканей височно-нижнечелюстного
- 34. Основные типы коллагенов в организме человека (всего 28 видов) I тип Повсеместно в мягких и твердых
- 36. II тип Хрящи, стекловидное тело, межпозвоночные диски Ранний остеохондроз, ахондрогенез, высокая степень миопии (близорукости)
- 37. III тип Мягкие ткани и полые органы Синдром Элерса-Данлоса, фиброзно- мышечная дисплазия, аневризма аорты, варикозная болезнь
- 38. IV тип Базальные мембраны в легких, почках и др. органах Синдром Альпорта, синдром Гудпасчера (нарушение транспорта
- 40. Скачать презентацию

Трансляция
Это синтез белка на рибосомах. Матрицей является и РНК.
Продукт трансляции –
Трансляция
Это синтез белка на рибосомах. Матрицей является и РНК.
Продукт трансляции –

Посттрансляционная модификация (ПТМ)
Это и есть созревание белка после трансляции.
В ходе ПТМ
Посттрансляционная модификация (ПТМ)
Это и есть созревание белка после трансляции.
В ходе ПТМ

Как правило, в ходе посттрансляционной модификации происходит частичное ОКИСЛЕНИЕ определенных аминокислот
Часто
Как правило, в ходе посттрансляционной модификации происходит частичное ОКИСЛЕНИЕ определенных аминокислот
Часто

Условия успешной трансляции
1. Наличие в клетке малых и больших субъединиц рибосом
2.
Условия успешной трансляции
1. Наличие в клетке малых и больших субъединиц рибосом
2.

Закон «все или ничего»
Если в клетке отсутствует хотя бы 1 незаменимая
Закон «все или ничего»
Если в клетке отсутствует хотя бы 1 незаменимая

Основные этапы трансляции
1. Инициация синтеза полипептидной цепи
2. Элонгация полипептидной цепи
3. Терминация
Основные этапы трансляции
1. Инициация синтеза полипептидной цепи
2. Элонгация полипептидной цепи
3. Терминация

Инициация трансляции
Малая и большая субъединицы рибосомы в присутствии иона магния с
Инициация трансляции
Малая и большая субъединицы рибосомы в присутствии иона магния с

Рибосома на этапе инициации
Рибосома на этапе инициации

Элонгация
Происходит удлинение полипептидной цепи на 1 аминокислоту, принесенную в рибосому новой
Элонгация
Происходит удлинение полипептидной цепи на 1 аминокислоту, принесенную в рибосому новой

Рибосома на этапе элонгации
Рибосома на этапе элонгации

Терминация
В конце иРНК, перед полиаденилатным хвостом, всегда содержится «бессмысленный» кодон, не
Терминация
В конце иРНК, перед полиаденилатным хвостом, всегда содержится «бессмысленный» кодон, не

Антибиотики и трансляция
Многие «классические» антибиотики ингибируют один из этапов трансляции
Поскольку рибосомы
Антибиотики и трансляция
Многие «классические» антибиотики ингибируют один из этапов трансляции
Поскольку рибосомы
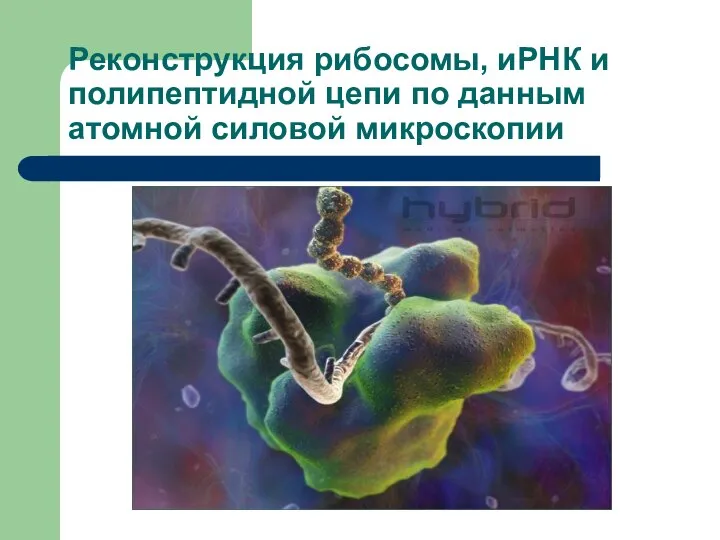
Реконструкция рибосомы, иРНК и полипептидной цепи по данным атомной силовой микроскопии
Реконструкция рибосомы, иРНК и полипептидной цепи по данным атомной силовой микроскопии

Примеры антибиотиков – ингибиторов трансляции
Примеры антибиотиков – ингибиторов трансляции

Коллаген и его роль
Коллаген – структурный белок, один из наиболее распространенных
Коллаген и его роль
Коллаген – структурный белок, один из наиболее распространенных

Виды соединительной ткани (световая микроскопия)
Виды соединительной ткани (световая микроскопия)

Особенности аминокислотного состава коллагена (первичная структура)
высокое содержание глицина
отсутствие триптофана
около 21 %
Особенности аминокислотного состава коллагена (первичная структура)
высокое содержание глицина
отсутствие триптофана
около 21 %

Вторичная структура
β-складчатый лист (подобие «гармошки»)
3 такие нити закручиваются в 3α-спираль
Эта
Вторичная структура
β-складчатый лист (подобие «гармошки»)
3 такие нити закручиваются в 3α-спираль
Эта
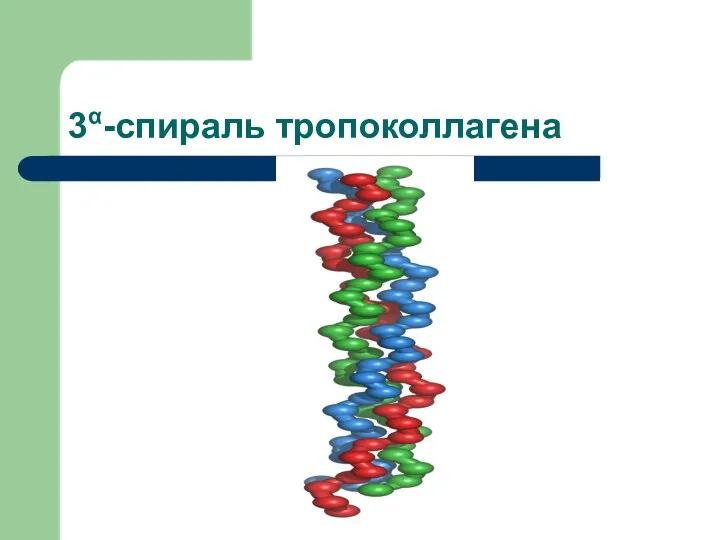
3α-спираль тропоколлагена
3α-спираль тропоколлагена

Свойства тропоколлагена
Это незрелый коллаген
Легко вымывается водой
Формирует фибриллу коллагена в присутствии ионов
Свойства тропоколлагена
Это незрелый коллаген
Легко вымывается водой
Формирует фибриллу коллагена в присутствии ионов

Основные этапы созревания коллагена (ПТМ)
1. Отщепление сигнальных пептидов от первичной последовательности
2.
Основные этапы созревания коллагена (ПТМ)
1. Отщепление сигнальных пептидов от первичной последовательности
2.

После гликозилирования тропоколлаген становится высокоантигенным белком, уникальным для каждого человека
Одна из
После гликозилирования тропоколлаген становится высокоантигенным белком, уникальным для каждого человека
Одна из

Формирование коллагеновой фибриллы
Коллагеновая фибрилла — это полукристаллическая структурная единица коллагена
Состоит
Формирование коллагеновой фибриллы
Коллагеновая фибрилла — это полукристаллическая структурная единица коллагена
Состоит

Формирование коллагеновой фибриллы
В фибриллах молекулы тропоколлагена смещены относительно друг друга примерно
Формирование коллагеновой фибриллы
В фибриллах молекулы тропоколлагена смещены относительно друг друга примерно

Формирование коллагеновой фибриллы
Для правильной укладки молекул тропоколлагена нужны ионы кальция
При дефиците
Формирование коллагеновой фибриллы
Для правильной укладки молекул тропоколлагена нужны ионы кальция
При дефиците

Коллагеновые фибриллы в электронном микроскопе
Коллагеновые фибриллы в электронном микроскопе

Взаимодействие молекул тропоколлагена при формировании фибриллы
Взаимодействие молекул тропоколлагена при формировании фибриллы

Придание коллагену прочностных свойств
Внутри тропоколлагенов существует ковалентная связь между цепями
Эти связи
Придание коллагену прочностных свойств
Внутри тропоколлагенов существует ковалентная связь между цепями
Эти связи

Нарушения синтеза коллагена
Врожденные аномалии (дефицит гидроксилаз пролина и лизина) –
Нарушения синтеза коллагена
Врожденные аномалии (дефицит гидроксилаз пролина и лизина) –

Стоматологические проявления
Ослабление связок зуба
Расшатывание и самопроизвольное выпадение зубов
Деструкция десневой ткани
Разрушение тканей
Стоматологические проявления
Ослабление связок зуба
Расшатывание и самопроизвольное выпадение зубов
Деструкция десневой ткани
Разрушение тканей


Основные типы коллагенов в организме человека (всего 28 видов)
I тип Повсеместно в
Основные типы коллагенов в организме человека (всего 28 видов)
I тип Повсеместно в


II тип Хрящи, стекловидное тело, межпозвоночные диски
Ранний остеохондроз, ахондрогенез, высокая степень
II тип Хрящи, стекловидное тело, межпозвоночные диски
Ранний остеохондроз, ахондрогенез, высокая степень

III тип Мягкие ткани и полые органы Синдром Элерса-Данлоса, фиброзно- мышечная дисплазия, аневризма аорты,
III тип Мягкие ткани и полые органы Синдром Элерса-Данлоса, фиброзно- мышечная дисплазия, аневризма аорты,

IV тип Базальные мембраны в легких, почках и др. органах
Синдром Альпорта, синдром
IV тип Базальные мембраны в легких, почках и др. органах
Синдром Альпорта, синдром
 Позитивизм
Позитивизм  Виклики, практики та стратегія реалізації реформ у регіонах
Виклики, практики та стратегія реалізації реформ у регіонах Маркетинг - promotion ( продвижение) 2
Маркетинг - promotion ( продвижение) 2 Понятия культуры и цивилизации. Виды и функции культуры
Понятия культуры и цивилизации. Виды и функции культуры России славные сыны…
России славные сыны… Цикл передачи системы ИКМ-30
Цикл передачи системы ИКМ-30 Узкополосный случайный процесс
Узкополосный случайный процесс Әмирхан Еники. Тормыш юлы һәм иҗаты
Әмирхан Еники. Тормыш юлы һәм иҗаты ВКР: Ручное изготовление изделия из группы сервировки стола с использованием традиционных материалов
ВКР: Ручное изготовление изделия из группы сервировки стола с использованием традиционных материалов Одежда говорит о человеке. 5 класс
Одежда говорит о человеке. 5 класс Дух предпринимательства преобразует экономику Д/З: § 4, читать, вопросы 1,2,5, пересказ
Дух предпринимательства преобразует экономику Д/З: § 4, читать, вопросы 1,2,5, пересказ Показатели радиоприемных устройств
Показатели радиоприемных устройств Grammaire Production orale Vidéo d’une expérience de voyage. Leçon 9
Grammaire Production orale Vidéo d’une expérience de voyage. Leçon 9 Презентация "Бюджет Российской Федерации" - скачать презентации по Экономике
Презентация "Бюджет Российской Федерации" - скачать презентации по Экономике 3. Essential Java Classes 5. Some Useful Classes
3. Essential Java Classes 5. Some Useful Classes Китай
Китай Бизнес-идея. Шаблон
Бизнес-идея. Шаблон Расчет тягово-динамический автобуса пригородного 5 класса с выполнением реальной части
Расчет тягово-динамический автобуса пригородного 5 класса с выполнением реальной части ФИЗОЛОГИЯ СТАРЕНИЯ Возрастная периодизация Общая характеристика процесса старения Изменения в органах и системах
ФИЗОЛОГИЯ СТАРЕНИЯ Возрастная периодизация Общая характеристика процесса старения Изменения в органах и системах  Java. Переменные и значения. (Лекция1)
Java. Переменные и значения. (Лекция1) Эффект изменения диаметра детали
Эффект изменения диаметра детали Тканевой обмен
Тканевой обмен Картина мира, созданная языком и культурой
Картина мира, созданная языком и культурой ТЕМА: Гнойная инфекция в хирургии Лекции для студентов III курса Кафедра общей хирургии составитель д.м.н., профессор Пушкаре
ТЕМА: Гнойная инфекция в хирургии Лекции для студентов III курса Кафедра общей хирургии составитель д.м.н., профессор Пушкаре Распорные сплошные ДК. Арки и рамы
Распорные сплошные ДК. Арки и рамы Энтропия
Энтропия Образование напряжений, деформаций и перемещений в сварных конструкциях. Лекция № 6
Образование напряжений, деформаций и перемещений в сварных конструкциях. Лекция № 6 Организация и содержание физминутки на уроке
Организация и содержание физминутки на уроке