- Тканевой обмен

Содержание
- 2. Содержание: 1.Метаболизм экзогенных и эндогенных липопротеидов (ЛП ). 2.Тканевой метаболизм липидов а).Механизм мобилизации жира( роль гормонов)
- 4. Основной массой пищевого жира яв-ся ТГ- нейтральный жир, поэтому создается 1-я форма транспорта прежде всего для
- 5. ХМ - частицы с диаметром от 90-1000 нм, и плотностью-ρ-0.93г/мл. Химический состав: - 88% ТГ, эф.ХС
- 6. Время жизни ХМ меньше 1 часа. Благодаря большим размерам ХМ не способны проникать из энтероцитов в
- 8. Отсюда проникают в кровяное русло. Уже через 1-2 часа после приема жирной пищи наблюдается алиментарная гиперлипемия-физиологическое
- 9. С током крови ХМ приносятся в жировую ткань, и подвергаются гидролизу на поверхности эндотелия капилляров жировой
- 10. Большая часть ЖК проходит внутрь адипоцитов, остальная часть связывается с альбуминами плазмы крови и уносятся с
- 12. В мышечной ткани также есть аналогичный ЛП-липазный фермент Обломки ХМ- ремнанты( первозданные ХМ- это насцентные), поступают
- 14. Главным липидным компонентом ЛПОНП являются триацилглицеролы. Однако, в отличие от хиломикронов, эти триацилглицеролы синтезируются в клетках
- 15. Основной функцией липопротеинов, содержащих апо- В, является транспорт ТАГ из печени к периферическим тканям, особенно в
- 16. Апо В-100 - это большой гидрофобный белок (4536 аминокислотных остатков), который синтезируется в печени. На его
- 17. Сборка липопротеинов, содержащих апо В-100, идет в эндоплазматическом ретикулуме; каждая частица ЛПОНП содержит один апо В-100.
- 18. Триацилглицеролы для ЛПОНП синтезируются путем эстерификации жирных кислот, поступающих в гепатоциты из плазмы крови (источником их
- 19. Уровень синтеза ЛПОНП регулируется также наличием холестерола, в особенности, образованием эфиров холестерола под действием ацил~КоА: холестеролацилтрансферазы
- 20. Сборка ЛПОНП регулируется на уровне посттрансляции за счет контроля наработки апо В-100. Значительное количество этого белка
- 21. Ассоциация апо В с ФХ сразу после трансляции обеспечивает возможность образования развернутой структуры белковой молекулы, необходимой
- 22. В регуляции сборки ЛПОНП чрезвычайно важную роль играют фосфатидилхолины. Об этом свидетельствует тот факт, что у
- 23. Примечательно, что блокируется секреция только ЛПОНП, в то время как секреция ЛПВП не изменяется. Внесение холина
- 24. Апопротеины ЛПОНП. Все белки, которые входят в состав липопротеинов, на пути своего образования проходят схожие этапы.
- 25. Новосинтезированная частица ЛПОНП содержит одну молекулу апо В-100. Апо С-II, апо С-III и апо Е поступают
- 26. В дополнению к обмену апопротеинами за счет ЛПОНП формируется поверхностный монослой ЛПВП. У ЛПОНП он становится
- 27. Перенос ЭХ осуществляется специальным белком, переносящим липиды (ЛПБ) также известен как белок, переносящий ЭХ (ЭХПБ) или
- 28. 1] Фермент секретируется в плазму крови из печени. МРНК ЛХАТ присутствует также в мозге. Однако белок,
- 29. ЛФХ является водорастворимым соединением, которое быстро удаляется из ЛПВП через водную фазу. В плазме он связывается
- 30. Сразу, вслед за ЛПОНП, печень посылает фермент ТГЛ- триглицеридлипазу-печеночную, которая выходит в кровоток и встречается ЛПОНП.
- 31. О регуляции печеночной липазы известно немного. Увеличение её активности происходит под влиянием тестостерона, других андрогенов и
- 32. Около 75% ЛППП попадает в печень после связывания апоЕ с рецепторами для ЛПНП или рецепторами для
- 33. В крови часть из ЛПОНП образуются ремнанты ЛППП ( ЛП промежуточной плотности). При электрофорезе они двигаются
- 35. Метаболизм ЛПНП Главным липидным компонентом ядра ЛПНП являются эфиры холестерола. Поэтому эти частицы являются основным средством
- 36. Ему принадлежит важная роль в прицельной доставке ЛПНП в клетку путем взаимодействия с рецепторами клеточной поверхности.
- 37. Количество таких рецепторов на поверхности клетки составляет от 15 000 до 70 000 . ЛПНП удаляются
- 38. Рецептор к ЛПНП
- 39. Такие рецепторы имеют низкую способность связывания с ЛПНП. Гораздо в большей степени у них выражена способность
- 40. В норме ЛПНП причаливают к печени в области рецептора и путем эндоцитоза проникают в клетку.Образуются эндосомы,
- 41. Схема поступления в клетки ЛПНП
- 42. Большинство тканей, в том числе и печень имеют рецепторы к ЛПНП. Эти рецепторы могут быть дефектными.,
- 43. Избыток эф.ХС подавляет процесс синтеза белков-рецепторов к ЛПОНП, который протекает в данной клетке, а также тормозит
- 44. ХС- это важнейший компонент биологических мембран -предшественник стероидных гормонов -источник желчных кислот -предшественник витамина D.
- 45. Извлечение избытка ХС из клетки осуществляется с помощью ЛПВП ( ЛП-высокой плотности)- антиатерогенного фракция ( синтезируется
- 46. ЛПВП подходит к клетке и с помощью фермента ЛХАТ( лецитин-холестерол-ацилтрансфераза), синтезированного в гепатоците, снимает ненасыщенную ЖК
- 50. Т.о. существуют 2 пути метаболизма ЛП-экзогенный и эндогенный. Экзогенный путь для ХС и ТГ, попадающих в
- 51. Аккумуляция холестерола в сосудистой стенке происходит вследствие дисбаланса между поступлением его в интиму сосудов и его
- 52. Во-первых, это изменения частиц ЛПНП (гликозилирование, перекисное окисление липидов, гидролиз фосфолипидов, окисление апо В). Поэтому они
- 53. Захват липопротеиновых частиц с помощью "мусорных" рецепторов протекает бесконтрольно. В отличие от апо В/Е - опосредованного
- 54. В результате макрофаги переполняются липидами, теряют функцию поглощения отходов и превращаются в пенистые клетки. Последние задерживаются
- 55. Во-вторых, это неэффективное высвобождение холестерола из эндотелия сосудистой стенки циркулирующими в крови ЛПВП[1]. [1] Антиатерогенные свойства
- 56. ЛПВП стимулируют образование простациклина и задерживают, следовательно, агрегацию тромбоцитов; они задерживают проникновение ЛПНП в интиму артерий;
- 57. Свойства клеточных рецепторов
- 59. Основные пути транспорта ХС в организме
- 60. Основные пути транспорта ХС в организме
- 61. Метаболизм липидов Л И П И Д Ы СЖК Ацил-КоА СН3-СО S-KoA ЦТК Синтез кетоновых тел
- 62. Главным эндогенным источником ЖК служит резерный жир, содержащийся в жировой ткани. Жировая ткань высокоспецифична. Ее функция
- 63. Распределени жира в организме зависит от нейрогуморальных факторов, половых и наследственных.
- 64. Мобилизация жира происходит при голодании, стрессе, физической нагрузке. В качестве источника энергии используются СНЖК, которые образуются
- 65. ТГ жировой ткани выполняют в обмене липидов такую же роль, как и гликоген печени в обмене
- 66. Свободные ЖК делятся на 3 группы: - насыщенные ЖК с четным числом атомов С. В животных
- 67. -Насыщенные ЖК с нечетным числом атомов С. Для них характерно ß- окисление до момента образования пропионил-S
- 68. В жировой ткани содержится много липаз, из которых наибольшее значение имеют ТГ-липаза (гормончувствительная) , ди и
- 69. ТГЛ, ДГЛ, МГЛ яв-ся клеточными липазами( их активность регулируется). Но при охлаждении они активируются. В плазме
- 72. При стрессе в результате мобилизации ТГ, ЖК в крови увеличиваются в 5 раз, благодаря чему глюкоза
- 73. При гидролизе ТГ, глицерин образуется в большем количестве, чем ЖК. Образованные ЖК нерастворимы в плазме и
- 74. ИТАК: источниками ЖК являются: липолиз под действием ТГЛ, ДГЛ, МГЛ распад ХМ под действием ЛПЛ распад
- 76. Основные параметры СЖК, циркулирующих в крови
- 77. человека
- 78. ß- окисление ЖК протекает в митохондриях и представляет собой последовательное ооооотщепление двухуглеродных фрагментов ( т.е. СН3-СО-S-КоА).
- 79. Реакция эта протекает, главным образом, в цитоплазме, в то время как процесс ß-окисления жирных кислот происходит
- 80. Образуется он из аминокислот лизина и метионина в печени и почках. На наружной стороне внутренней мембраны
- 83. Опосредованный карнитином перенос длинноцепочечного ацил-КоА в митохондриальный матрикс КПТн катализирует образование ацилкарнитинового комплекса из ацил-КоА и
- 84. Там совместное последовательное действие карнитин:ацилкарнитин транслоказы (Т) и КПТв обеспечивает поступление ацил-КоА в митохондриальный матрикс для
- 85. Ацилкарнитин обладает способностью проходить через внутреннюю мембрану митохондрий. На внутренней поверхности внутренней мембраны митохондрий ацилкарнитин взаимодействует
- 86. Далее митохондриальный ацил-КоА распадается в результате повторяющейся последовательности из четырех реакций окисления с участием флавинадениндинуклеотида (ФАД),
- 88. Регуляция ß- окисления Конкуренция глюкозы и жирных кислот за использование в качестве субстратов: цикл Рэндэла.
- 89. Увеличенное окисление жирных кислот ингибирует окисление глюкозы в клетках скелетных мышц и сердца за счет ингибирования
- 90. Однако у людей с высоким уровнем СЖК это является одной из причин устойчивости к действию инсулина
- 91. Это обусловлено тем, что регуляция поглощения жирных кислот митохондриями преимущественно осуществляется за счет контроля КПТI со
- 92. Гипергликемия частично подавляет липолиз. Энергетический выход окисления жирных кислот зависит от длины цепи. Можно подсчитать энергетический
- 93. При окислении каждого из этих НАДН через дыхательную цепь образуется три молекулы АТФ, тогда как при
- 94. Таким образом, энергетический выход 1 цикла b -окисления составляет 5 молекул АТФ + 12 молекул АТФ.
- 95. b-окисления (оно составляет n/2 - 1, где n - число углеродных атомов в составе жирной кислоты)
- 96. Реакции β-окисления тесно сопряжены друг с другом. Промежуточные продукты неизбежно переходят из одной реакции в другую;
- 97. Энергетический выход β-окисления на примере пальмитиновой кислоты. Образование АТФ (2 АТФ/ФАДН2; 3 АТФ/НАДН; 12 АТФ/ацетил~КоА; таким
- 98. Жирные кислоты с очень длинной цепью. Особенностью метаболизма жирных кислот в пероксисомах является расщепление тех из
- 99. Укорочение алкильной цепи в пероксисомах происходит до тех пор, пока не образуется ацил-КоА со средней длиной
- 100. Образующийся ацил-КоА с С-8 впоследствии подвергается дальнейшему окислению в митохондриях. Первоначальная стадия дегидрирования в ходе пероксисомального
- 101. Окисление дикарбоновых кислот. В пероксисомах происходит также окисление дикарбоновых кислот, образующихся в ходе ω-окисления. Само ω-окисление
- 102. Окисление ненасыщенных ЖК происходит также как и у насыщенных , но с предварительным переносом = связи
- 106. Начинается материал лекции Липиды-3
- 107. Малонил-КоА - это начальный промежуточный продукт в синтезе жирных кислот, образованный из ацетил-КоА в цитоплазме.
- 108. Избыток ацетил-КоА в митохондриях не может самостоятельно пройти в цитоплазму. Проход через митохондриальную мембрану становится возможным
- 109. На эту реакцию расходуется СО2 и АТФ. Таким образом, условия, которые способствуют липогенезу (наличие большого количества
- 110. Происхождение ненасыщенных жирных кислот в клетках организма. Метаболизм арахидоновой кислоты Незаменимые и заменимые - Среди ненасыщенных
- 111. К таким жирным кислотам относятся линолевая кислота (18:2, Δ9,12), линоленовая кислота (18:3, Δ9,12,15) и арахидоновая кислота
- 112. У человека при недостатке в пище незаменимых жирных кислот описаны дерматологические изменения. Обычный рацион взрослых людей
- 113. Случаи подобного дефицита наблюдаются и у пациентов, которые длительное время находятся на парентеральном питании, обедненном незаменимыми
- 114. Синтез ненасыщенных жирных кислот из насыщенных с параллельным удлинением цепи. Десатурация проходит под действием микросомального комплекса
- 115. Из этих компонентов образуется короткая цепь переноса электронов, с помощью которой на короткий период времени в
- 117. Образование и утилизация кетоновых тел Двумя основными видами ацетоновых тел являются ацетоацетат и β-гидроксибутират. β-гидроксибутират -
- 119. Первоначальная стадия этого процесса катализируется ферментом - β-кетотиолазой. Затем ацетоацетил-КоА конденсируется со следующей молекулой ацетил-КоА под
- 120. В дальнейшем ацетоуксусная кислота восстанавливается под влиянием фермента b-гидроксибутиратдегидрогеназы и в результате образуется b-оксимасляная кислота.
- 121. Затем фермент - ГОМГ-КоА лиаза катализирует расщепление ГОМГ-КоА на ацетоацетат и ацетил-КоА. В дальнейшем ацетоуксусная кислота
- 122. Количество ацетоацетата, которое восстанавливается в β-гидроксибутират, зависит от соотношения НАДН/НАД+. Восстановление это происходит под влиянием фермента
- 123. эти реакции происходят в митохондриях. В цитозоле имеются изоферменты - β-кетотиолазы и ГОМГ~КоА синтетазы, которые также
- 124. Образование кетоновых тел в печени контролируется состоянием питания. Такое контрольное действие усиливается инсулином и глюкагоном. Принятие
- 125. При голодании усиливается липолиз, растет уровень глюкагона и концентрация цАМФ в печени. Происходит фосфорилирование, тем самым
- 126. В норме кетоновые тела являются источником энергии для мышц; при продолжительном голодании они могут использоваться центральной
- 127. Такая избирательность обусловлена локализацией ферментов, катализирующих этот процесс. Сначала β-гидроксибутират дегидрогеназа катализирует окисление β-гидроксибутирата до ацетоацетата
- 128. Образуется ацетоацетил~КоА, который является промежуточным продуктом последнего витка β-окисления жирных кислот. Этот фермент в печени не
- 129. Зато спустя несколько суток после начала голодания в клетках мозга начинается экспрессия гена, кодирующего этот фермент.
- 130. Тиолаза довершает расщепление ацетоацетил-КоА, встраивая КоА по месту разрыва связи между α и β углеродными атомами.
- 131. Интенсивность окисления кетоновых тел во внепеченочных тканях пропорциональна их концентрации в крови. Общая концентрация кетоновых тел
- 132. В определенных метаболических условиях, когда происходит интенсивное окисление жирных кислот, в печени образуются значительные количества так
- 133. Состояние организма, при котором концентрация кетоновых тел в крови выше нормальной, называется кетонемией. Повышенное содержание кетоновых
- 134. Он обусловлен спонтанным декарбоксилированием ацетоацетата в ацетон. Эти три симптома - кетонемия, кетонурия и запах ацетона
- 135. Кетоз возникает в результате недостатка доступных углеводов. Например, при голодании их мало поступает (или не поступает)
- 136. Это приводит к дисбалансу между этерификацией и липолизом в жировой ткани в сторону интенсификации последнего. Он
- 138. Скачать презентацию

Содержание:
1.Метаболизм экзогенных и эндогенных липопротеидов (ЛП ).
2.Тканевой метаболизм липидов
а).Механизм
Содержание:
1.Метаболизм экзогенных и эндогенных липопротеидов (ЛП ).
2.Тканевой метаболизм липидов
а).Механизм


Основной массой пищевого жира яв-ся ТГ- нейтральный жир, поэтому
Основной массой пищевого жира яв-ся ТГ- нейтральный жир, поэтому

ХМ - частицы с диаметром от 90-1000 нм, и
ХМ - частицы с диаметром от 90-1000 нм, и

Время жизни ХМ меньше 1 часа. Благодаря большим размерам
Время жизни ХМ меньше 1 часа. Благодаря большим размерам


Отсюда проникают в кровяное русло. Уже через 1-2 часа
Отсюда проникают в кровяное русло. Уже через 1-2 часа

С током крови ХМ приносятся в жировую ткань, и подвергаются
С током крови ХМ приносятся в жировую ткань, и подвергаются

Большая часть ЖК проходит внутрь адипоцитов, остальная часть связывается
Большая часть ЖК проходит внутрь адипоцитов, остальная часть связывается


В мышечной ткани также есть аналогичный ЛП-липазный фермент
В мышечной ткани также есть аналогичный ЛП-липазный фермент


Главным липидным компонентом ЛПОНП являются триацилглицеролы. Однако, в отличие
Главным липидным компонентом ЛПОНП являются триацилглицеролы. Однако, в отличие

Основной функцией липопротеинов, содержащих апо- В, является транспорт ТАГ
Основной функцией липопротеинов, содержащих апо- В, является транспорт ТАГ

Апо В-100 - это большой гидрофобный белок (4536 аминокислотных
Апо В-100 - это большой гидрофобный белок (4536 аминокислотных

Сборка липопротеинов, содержащих апо В-100, идет в эндоплазматическом ретикулуме;
Сборка липопротеинов, содержащих апо В-100, идет в эндоплазматическом ретикулуме;

Триацилглицеролы для ЛПОНП синтезируются путем эстерификации жирных кислот, поступающих
Триацилглицеролы для ЛПОНП синтезируются путем эстерификации жирных кислот, поступающих

Уровень синтеза ЛПОНП регулируется также наличием холестерола, в особенности, образованием
Уровень синтеза ЛПОНП регулируется также наличием холестерола, в особенности, образованием

Сборка ЛПОНП регулируется на уровне посттрансляции за счет контроля наработки
Сборка ЛПОНП регулируется на уровне посттрансляции за счет контроля наработки

Ассоциация апо В с ФХ сразу после трансляции обеспечивает возможность
Ассоциация апо В с ФХ сразу после трансляции обеспечивает возможность

В регуляции сборки ЛПОНП чрезвычайно важную роль играют фосфатидилхолины. Об
В регуляции сборки ЛПОНП чрезвычайно важную роль играют фосфатидилхолины. Об

Примечательно, что блокируется секреция только ЛПОНП, в то время как
Примечательно, что блокируется секреция только ЛПОНП, в то время как

Апопротеины ЛПОНП. Все белки, которые входят в состав липопротеинов, на
Апопротеины ЛПОНП. Все белки, которые входят в состав липопротеинов, на

Новосинтезированная частица ЛПОНП содержит одну молекулу апо В-100. Апо
Новосинтезированная частица ЛПОНП содержит одну молекулу апо В-100. Апо

В дополнению к обмену апопротеинами за счет ЛПОНП формируется поверхностный монослой
В дополнению к обмену апопротеинами за счет ЛПОНП формируется поверхностный монослой

Перенос ЭХ осуществляется специальным белком, переносящим липиды (ЛПБ) также известен
Перенос ЭХ осуществляется специальным белком, переносящим липиды (ЛПБ) также известен
![1] Фермент секретируется в плазму крови из печени. МРНК ЛХАТ присутствует](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1292283/slide-27.jpg)
1] Фермент секретируется в плазму крови из печени. МРНК ЛХАТ присутствует
1] Фермент секретируется в плазму крови из печени. МРНК ЛХАТ присутствует

ЛФХ является водорастворимым соединением, которое быстро удаляется из ЛПВП через водную
ЛФХ является водорастворимым соединением, которое быстро удаляется из ЛПВП через водную

Сразу, вслед за ЛПОНП, печень посылает фермент ТГЛ- триглицеридлипазу-печеночную,
Сразу, вслед за ЛПОНП, печень посылает фермент ТГЛ- триглицеридлипазу-печеночную,

О регуляции печеночной липазы известно немного. Увеличение её активности происходит
О регуляции печеночной липазы известно немного. Увеличение её активности происходит

Около 75% ЛППП попадает в печень после связывания апоЕ с
Около 75% ЛППП попадает в печень после связывания апоЕ с

В крови часть из ЛПОНП образуются ремнанты ЛППП (
В крови часть из ЛПОНП образуются ремнанты ЛППП (


Метаболизм ЛПНП
Главным липидным компонентом ядра ЛПНП являются эфиры холестерола.
Метаболизм ЛПНП
Главным липидным компонентом ядра ЛПНП являются эфиры холестерола.

Ему принадлежит важная роль в прицельной доставке ЛПНП в клетку
Ему принадлежит важная роль в прицельной доставке ЛПНП в клетку

Количество таких рецепторов на поверхности клетки составляет от 15 000
Количество таких рецепторов на поверхности клетки составляет от 15 000
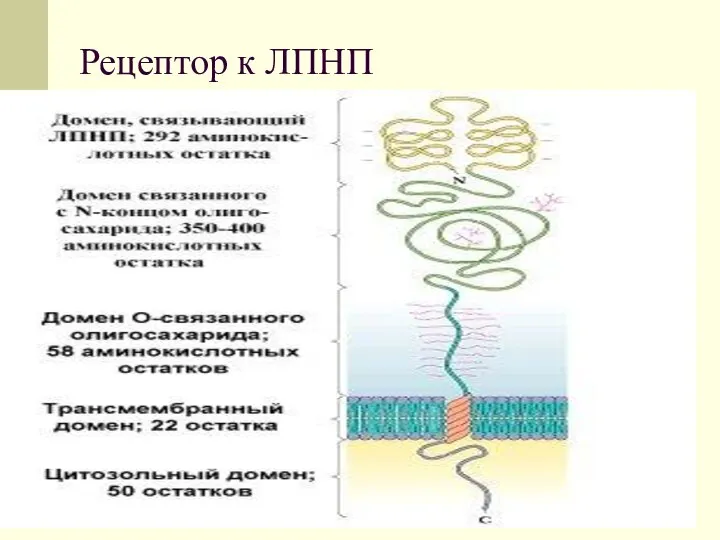
Рецептор к ЛПНП
Рецептор к ЛПНП

Такие рецепторы имеют низкую способность связывания с ЛПНП. Гораздо в большей
Такие рецепторы имеют низкую способность связывания с ЛПНП. Гораздо в большей

В норме ЛПНП причаливают к печени в области рецептора
В норме ЛПНП причаливают к печени в области рецептора
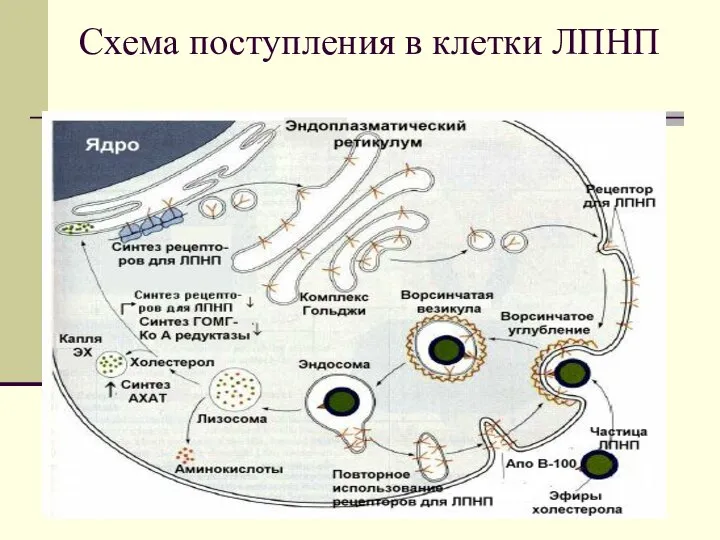
Схема поступления в клетки ЛПНП
Схема поступления в клетки ЛПНП

Большинство тканей, в том числе и печень имеют рецепторы к
Большинство тканей, в том числе и печень имеют рецепторы к

Избыток эф.ХС подавляет процесс синтеза белков-рецепторов к ЛПОНП, который протекает
Избыток эф.ХС подавляет процесс синтеза белков-рецепторов к ЛПОНП, который протекает

ХС- это важнейший компонент биологических мембран
-предшественник стероидных гормонов
-источник
ХС- это важнейший компонент биологических мембран
-предшественник стероидных гормонов
-источник

Извлечение избытка ХС из клетки осуществляется с помощью ЛПВП (
Извлечение избытка ХС из клетки осуществляется с помощью ЛПВП (

ЛПВП подходит к клетке и с помощью фермента ЛХАТ(
ЛПВП подходит к клетке и с помощью фермента ЛХАТ(




Т.о. существуют 2 пути метаболизма ЛП-экзогенный и эндогенный.
Экзогенный
Т.о. существуют 2 пути метаболизма ЛП-экзогенный и эндогенный.
Экзогенный

Аккумуляция холестерола в сосудистой стенке происходит вследствие дисбаланса между поступлением
Аккумуляция холестерола в сосудистой стенке происходит вследствие дисбаланса между поступлением

Во-первых, это изменения частиц ЛПНП (гликозилирование, перекисное окисление липидов, гидролиз
Во-первых, это изменения частиц ЛПНП (гликозилирование, перекисное окисление липидов, гидролиз

Захват липопротеиновых частиц с помощью "мусорных" рецепторов протекает бесконтрольно. В
Захват липопротеиновых частиц с помощью "мусорных" рецепторов протекает бесконтрольно. В

В результате макрофаги переполняются липидами, теряют функцию поглощения отходов и
В результате макрофаги переполняются липидами, теряют функцию поглощения отходов и

Во-вторых, это неэффективное высвобождение холестерола из эндотелия сосудистой стенки циркулирующими
Во-вторых, это неэффективное высвобождение холестерола из эндотелия сосудистой стенки циркулирующими

ЛПВП стимулируют образование простациклина и задерживают, следовательно, агрегацию тромбоцитов; они
ЛПВП стимулируют образование простациклина и задерживают, следовательно, агрегацию тромбоцитов; они

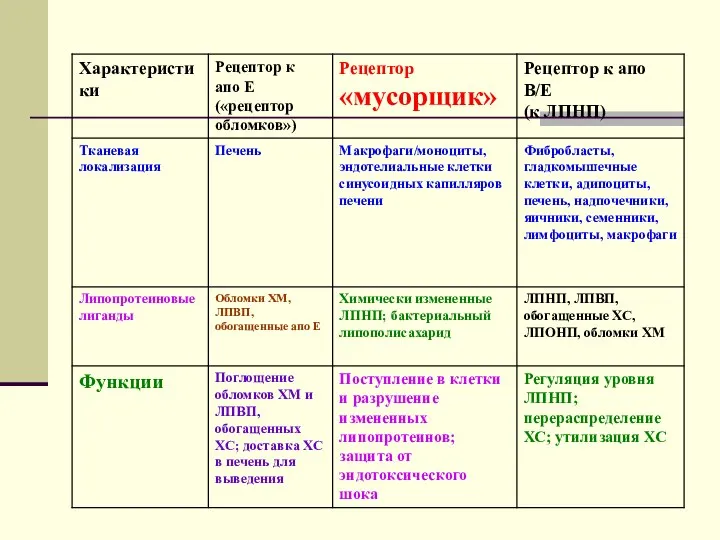
Свойства клеточных рецепторов

Основные пути
транспорта ХС в
организме
Основные пути
транспорта ХС в
организме
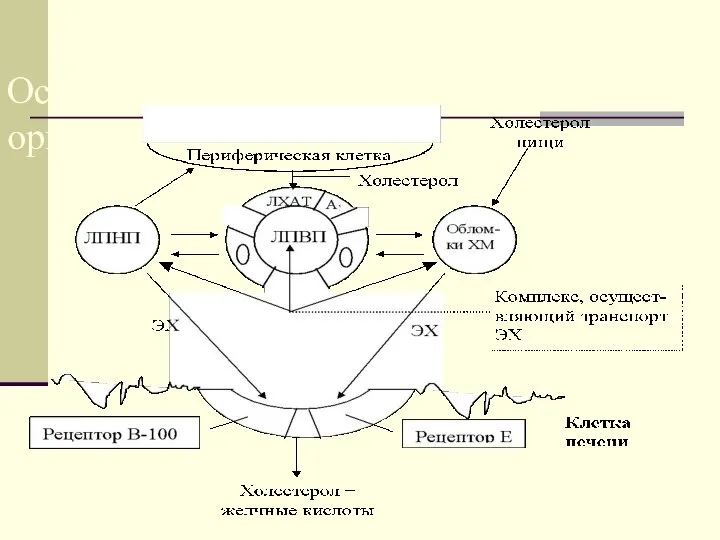
Основные пути транспорта ХС в организме
Основные пути транспорта ХС в организме
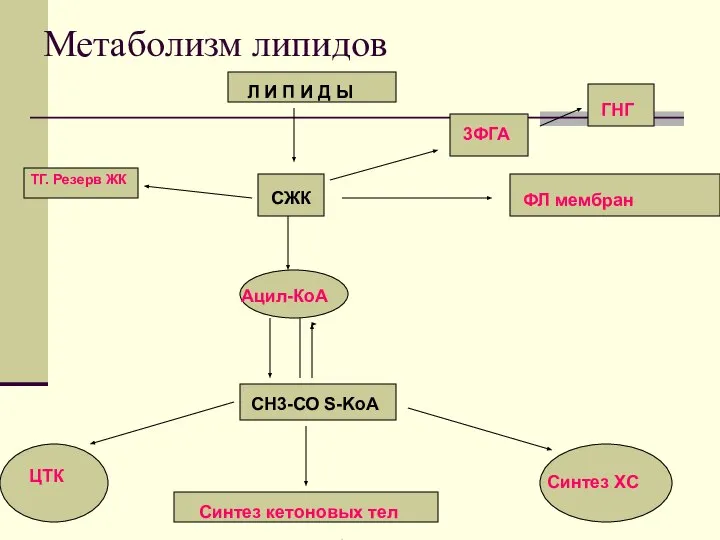
Метаболизм липидов
Л И П И Д Ы
СЖК
Ацил-КоА
СН3-СО
Метаболизм липидов
Л И П И Д Ы
СЖК
Ацил-КоА
СН3-СО

Главным эндогенным источником ЖК служит резерный жир, содержащийся в
Главным эндогенным источником ЖК служит резерный жир, содержащийся в

Распределени жира в организме зависит от нейрогуморальных факторов, половых
Распределени жира в организме зависит от нейрогуморальных факторов, половых

Мобилизация жира происходит при голодании, стрессе, физической нагрузке. В качестве
Мобилизация жира происходит при голодании, стрессе, физической нагрузке. В качестве

ТГ жировой ткани выполняют в обмене липидов такую же
ТГ жировой ткани выполняют в обмене липидов такую же

Свободные ЖК делятся на 3 группы:
- насыщенные ЖК с четным
Свободные ЖК делятся на 3 группы:
- насыщенные ЖК с четным

-Насыщенные ЖК с нечетным числом атомов С. Для них характерно
-Насыщенные ЖК с нечетным числом атомов С. Для них характерно

В жировой ткани содержится много липаз, из которых наибольшее значение
В жировой ткани содержится много липаз, из которых наибольшее значение

ТГЛ, ДГЛ, МГЛ яв-ся клеточными липазами( их активность регулируется). Но
ТГЛ, ДГЛ, МГЛ яв-ся клеточными липазами( их активность регулируется). Но



При стрессе в результате мобилизации ТГ, ЖК в крови увеличиваются
При стрессе в результате мобилизации ТГ, ЖК в крови увеличиваются

При гидролизе ТГ, глицерин образуется в большем количестве, чем ЖК.
При гидролизе ТГ, глицерин образуется в большем количестве, чем ЖК.

ИТАК: источниками ЖК являются:
липолиз под действием ТГЛ, ДГЛ, МГЛ
распад ХМ
ИТАК: источниками ЖК являются:
липолиз под действием ТГЛ, ДГЛ, МГЛ
распад ХМ


Основные параметры СЖК, циркулирующих в
крови
Основные параметры СЖК, циркулирующих в
крови

человека
человека

ß- окисление ЖК протекает в митохондриях и представляет собой последовательное
ß- окисление ЖК протекает в митохондриях и представляет собой последовательное

Реакция эта протекает, главным образом, в цитоплазме, в то
Реакция эта протекает, главным образом, в цитоплазме, в то

Образуется он из аминокислот лизина и метионина в печени
Образуется он из аминокислот лизина и метионина в печени
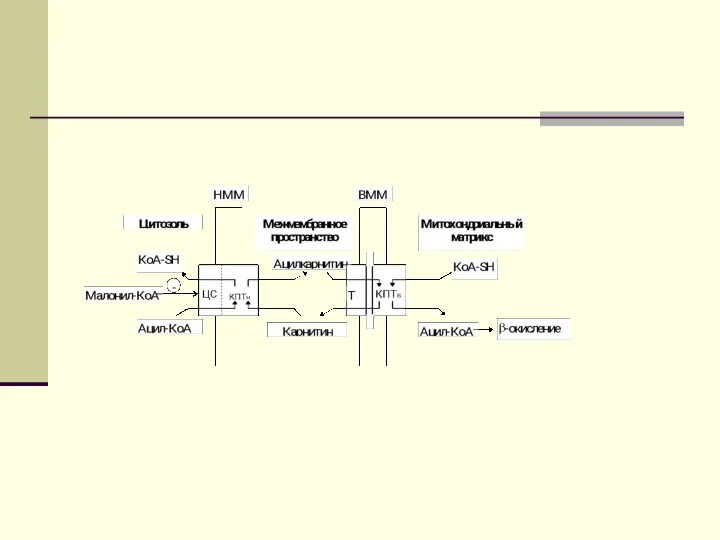
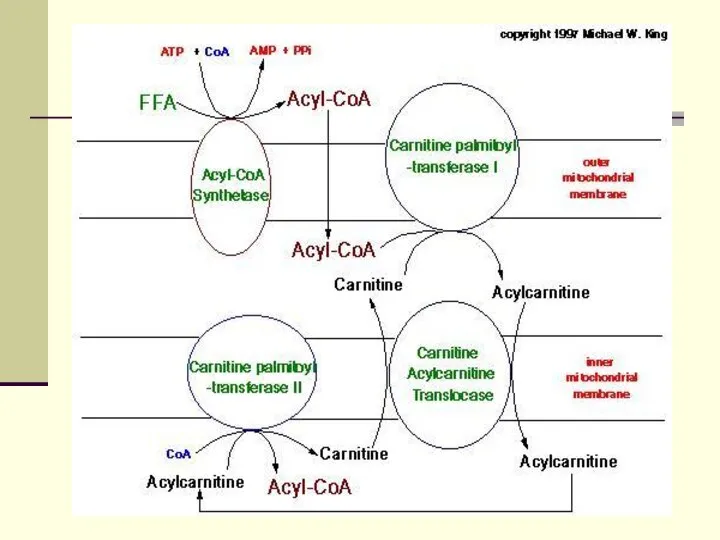

Опосредованный карнитином перенос длинноцепочечного ацил-КоА в митохондриальный матрикс
КПТн катализирует
Опосредованный карнитином перенос длинноцепочечного ацил-КоА в митохондриальный матрикс
КПТн катализирует

Там совместное последовательное действие карнитин:ацилкарнитин транслоказы (Т) и КПТв
Там совместное последовательное действие карнитин:ацилкарнитин транслоказы (Т) и КПТв

Ацилкарнитин обладает способностью проходить через внутреннюю мембрану митохондрий. На
Ацилкарнитин обладает способностью проходить через внутреннюю мембрану митохондрий. На

Далее митохондриальный ацил-КоА распадается в результате повторяющейся последовательности из четырех
Далее митохондриальный ацил-КоА распадается в результате повторяющейся последовательности из четырех


Регуляция ß- окисления
Конкуренция глюкозы и жирных кислот за использование
Регуляция ß- окисления
Конкуренция глюкозы и жирных кислот за использование

Увеличенное окисление жирных кислот ингибирует окисление глюкозы в клетках скелетных
Увеличенное окисление жирных кислот ингибирует окисление глюкозы в клетках скелетных

Однако у людей с высоким уровнем СЖК это является одной
Однако у людей с высоким уровнем СЖК это является одной

Это обусловлено тем, что регуляция поглощения жирных кислот митохондриями
Это обусловлено тем, что регуляция поглощения жирных кислот митохондриями

Гипергликемия частично подавляет липолиз. Энергетический выход окисления жирных кислот зависит
Гипергликемия частично подавляет липолиз. Энергетический выход окисления жирных кислот зависит

При окислении каждого из этих НАДН через дыхательную цепь образуется
При окислении каждого из этих НАДН через дыхательную цепь образуется

Таким образом, энергетический выход 1 цикла b -окисления составляет
Таким образом, энергетический выход 1 цикла b -окисления составляет

b-окисления (оно составляет n/2 - 1, где n - число
b-окисления (оно составляет n/2 - 1, где n - число

Реакции β-окисления тесно сопряжены друг с другом. Промежуточные продукты неизбежно переходят

Энергетический выход β-окисления на примере пальмитиновой кислоты. Образование АТФ (2 АТФ/ФАДН2;
Энергетический выход β-окисления на примере пальмитиновой кислоты. Образование АТФ (2 АТФ/ФАДН2;

Жирные кислоты с очень длинной цепью. Особенностью метаболизма жирных кислот в

Укорочение алкильной цепи в пероксисомах происходит до тех пор, пока
Укорочение алкильной цепи в пероксисомах происходит до тех пор, пока

Образующийся ацил-КоА с С-8 впоследствии подвергается дальнейшему окислению в митохондриях.
Первоначальная стадия
Образующийся ацил-КоА с С-8 впоследствии подвергается дальнейшему окислению в митохондриях.
Первоначальная стадия

Окисление дикарбоновых кислот. В пероксисомах происходит также окисление дикарбоновых кислот,
Окисление дикарбоновых кислот. В пероксисомах происходит также окисление дикарбоновых кислот,

Окисление ненасыщенных ЖК происходит также как и у насыщенных
Окисление ненасыщенных ЖК происходит также как и у насыщенных




Начинается материал лекции
Липиды-3
Липиды-3

Малонил-КоА - это начальный промежуточный продукт в синтезе жирных кислот,
Малонил-КоА - это начальный промежуточный продукт в синтезе жирных кислот,

Избыток ацетил-КоА в митохондриях не может самостоятельно пройти в цитоплазму.
Избыток ацетил-КоА в митохондриях не может самостоятельно пройти в цитоплазму.

На эту реакцию расходуется СО2 и АТФ. Таким образом,
На эту реакцию расходуется СО2 и АТФ. Таким образом,

Происхождение ненасыщенных жирных кислот в клетках организма. Метаболизм арахидоновой кислоты
Незаменимые и
Происхождение ненасыщенных жирных кислот в клетках организма. Метаболизм арахидоновой кислоты
Незаменимые и

К таким жирным кислотам относятся линолевая кислота (18:2, Δ9,12), линоленовая
К таким жирным кислотам относятся линолевая кислота (18:2, Δ9,12), линоленовая

У человека при недостатке в пище незаменимых жирных кислот описаны
У человека при недостатке в пище незаменимых жирных кислот описаны

Случаи подобного дефицита наблюдаются и у пациентов, которые длительное время
Случаи подобного дефицита наблюдаются и у пациентов, которые длительное время

Синтез ненасыщенных жирных кислот из насыщенных с параллельным удлинением цепи.
Синтез ненасыщенных жирных кислот из насыщенных с параллельным удлинением цепи.

Из этих компонентов образуется короткая цепь переноса электронов, с помощью
Из этих компонентов образуется короткая цепь переноса электронов, с помощью


Образование и утилизация кетоновых тел
Двумя основными видами ацетоновых тел являются ацетоацетат
Образование и утилизация кетоновых тел
Двумя основными видами ацетоновых тел являются ацетоацетат


Первоначальная стадия этого процесса катализируется ферментом - β-кетотиолазой. Затем
Первоначальная стадия этого процесса катализируется ферментом - β-кетотиолазой. Затем

В дальнейшем ацетоуксусная кислота восстанавливается под влиянием фермента b-гидроксибутиратдегидрогеназы
В дальнейшем ацетоуксусная кислота восстанавливается под влиянием фермента b-гидроксибутиратдегидрогеназы

Затем фермент - ГОМГ-КоА лиаза катализирует расщепление ГОМГ-КоА на
Затем фермент - ГОМГ-КоА лиаза катализирует расщепление ГОМГ-КоА на

Количество ацетоацетата, которое восстанавливается в β-гидроксибутират, зависит от соотношения
Количество ацетоацетата, которое восстанавливается в β-гидроксибутират, зависит от соотношения

эти реакции происходят в митохондриях. В цитозоле имеются изоферменты - β-кетотиолазы

Образование кетоновых тел в печени контролируется состоянием питания. Такое
Образование кетоновых тел в печени контролируется состоянием питания. Такое

При голодании усиливается липолиз, растет уровень глюкагона
и концентрация
При голодании усиливается липолиз, растет уровень глюкагона
и концентрация

В норме кетоновые тела являются источником энергии для мышц; при продолжительном

Такая избирательность обусловлена локализацией ферментов, катализирующих этот процесс.
Сначала
Такая избирательность обусловлена локализацией ферментов, катализирующих этот процесс.
Сначала

Образуется ацетоацетил~КоА, который является промежуточным продуктом последнего витка β-окисления жирных
Образуется ацетоацетил~КоА, который является промежуточным продуктом последнего витка β-окисления жирных

Зато спустя несколько суток после начала голодания в клетках
Зато спустя несколько суток после начала голодания в клетках

Тиолаза довершает расщепление ацетоацетил-КоА, встраивая КоА по месту разрыва
Тиолаза довершает расщепление ацетоацетил-КоА, встраивая КоА по месту разрыва

Интенсивность окисления кетоновых тел во внепеченочных тканях пропорциональна их
Интенсивность окисления кетоновых тел во внепеченочных тканях пропорциональна их

В определенных метаболических условиях, когда происходит интенсивное окисление жирных кислот,
В определенных метаболических условиях, когда происходит интенсивное окисление жирных кислот,

Состояние организма, при котором концентрация кетоновых тел в крови выше
Состояние организма, при котором концентрация кетоновых тел в крови выше

Он обусловлен спонтанным декарбоксилированием ацетоацетата в ацетон. Эти три
Он обусловлен спонтанным декарбоксилированием ацетоацетата в ацетон. Эти три

Кетоз возникает в результате недостатка доступных углеводов. Например, при
Кетоз возникает в результате недостатка доступных углеводов. Например, при

Это приводит к дисбалансу между этерификацией и липолизом в
Это приводит к дисбалансу между этерификацией и липолизом в
 Уж так задумано в природе Не знаю, в шутку иль всерьёз. На тёмно-синем небосводе Есть миллионы ярких звёзд
Уж так задумано в природе Не знаю, в шутку иль всерьёз. На тёмно-синем небосводе Есть миллионы ярких звёзд  Костная ткань
Костная ткань  синтез спиросоединений из антраниламида
синтез спиросоединений из антраниламида  Рынок биотехнологии РФ Abercade Consulting 2004
Рынок биотехнологии РФ Abercade Consulting 2004 Статические массивы
Статические массивы ЛОГИКА 2 класс урок 1 Тема: «Снова в школу»
ЛОГИКА 2 класс урок 1 Тема: «Снова в школу» Типы данных, адресация. Введение. Тема 1
Типы данных, адресация. Введение. Тема 1 Педагогический совет «Мотивация учебной деятельности обучающихся и обеспечение условий для ее развития» 13 декабря 2005 года
Педагогический совет «Мотивация учебной деятельности обучающихся и обеспечение условий для ее развития» 13 декабря 2005 года Региональные таможенные управления. _
Региональные таможенные управления. _ Тема №4: Методы принятия решений в сфере сервиса и туризма
Тема №4: Методы принятия решений в сфере сервиса и туризма  Достоинства и недостатки Window 7
Достоинства и недостатки Window 7 Cascading Style Sheets. Веб. CSS. п.9. Лекція 2
Cascading Style Sheets. Веб. CSS. п.9. Лекція 2 Как правильно?
Как правильно? Качественное сказуемое. Китайский язык
Качественное сказуемое. Китайский язык Организационная культура учреждения
Организационная культура учреждения Негосударственное дошкольное образовательное учреждение «Детский сад №5 ОАО «РЖД»
Негосударственное дошкольное образовательное учреждение «Детский сад №5 ОАО «РЖД» ВКР: Исследование судовых нагревательных, отопительных и холодильных установок
ВКР: Исследование судовых нагревательных, отопительных и холодильных установок Учитель ГОУ школы № 341 г. Санкт-Петербурга Паничева Марина Владимировна
Учитель ГОУ школы № 341 г. Санкт-Петербурга Паничева Марина Владимировна Международное финансирование развития
Международное финансирование развития  Git. Working directory
Git. Working directory Музей изобразительных искусств в Будапеште
Музей изобразительных искусств в Будапеште Особенности русского народного костюма
Особенности русского народного костюма Презентация "Коммерческие банки" - скачать презентации по Экономике
Презентация "Коммерческие банки" - скачать презентации по Экономике Числовые функции и способы их задания - презентация по Алгебре
Числовые функции и способы их задания - презентация по Алгебре Система частиц. Простейшие эффекты
Система частиц. Простейшие эффекты Военная служба как особый вид государственной службы
Военная служба как особый вид государственной службы I.От знания учебного предмета II.От знакомства с процедурой проведения экзамена III.От стратегии подготовки к экзамену IV.От психологи
I.От знания учебного предмета II.От знакомства с процедурой проведения экзамена III.От стратегии подготовки к экзамену IV.От психологи Петергоф. Дворцы
Петергоф. Дворцы