- Патофизиология клетки

Содержание
- 2. Рудольф Вирхов — основатель целлюлярной (клеточной) патологии 1821-1902 "omnis ceilula е cellula" (клетка происходит только от
- 3. 1858 год «Целлюлярная патология как учение, основанное на физиологической и патологической гистологии»
- 4. Основной постулат труда Р.Вирхова - Всякое болезненное изменение организма связано с патологическим процессом в клетках
- 5. Причины повреждения клеток Физические факторы: -Механические -Температура -Ионизирующая радиация -Ультрафиолетовое излучение -Электромагнитные
- 6. Химические факторы органические и неорганические кислоты, щелочи, соли тяжелых металлов, продукты нарушенного метаболизма, лекарственные препараты и
- 7. Биологические факторы Наибольшее значение среди них имеют вирусы, риккетсии, бактерии, паразиты, грибы
- 8. И другие факторы повреждения Гипоксия Гипероксия Иммунологические Нарушение КОС Нарушение водно-электролитного обмена Нарушение углеводного обмена Генетические
- 9. Реакция клеток на повреждение нарушение функции клеток нарушение клеточных защитных реакций – (активация или ингибиция) изменение
- 10. Изменение метаболизма - нарушение функции транспортных систем клеточной мембраны - повышение проницаемости внутриклеточных мембран и плазмалеммы
- 11. -нарушение дифференцировки и роста клеток (опухолевый рост) -патология репродукции клеток -патология движения клеток -патология ядра и
- 12. Вывод: повреждение клетки - это нарушение клеточного обмена веществ, внутриклеточного гомеостазиса.
- 13. Повреждения летальные сублетальные Специфические и неспецифические
- 14. Основные механизмы острого летального повреждения - чрезмерное подавление синтеза АТФ - разрушение митохондрий - нарушение работы
- 15. Специфическое повреждение -определяется этиологическим фактором Иммунный гемолиз, радиационное повреждение свободными радикалами, повреждение дыхательных ферментов цианистыми соединениями,
- 16. Неспецифическое повреждение Утрата принципа неравновесности К в клетке в 20-30 раз выше, чем вне клетки (в
- 17. - Внутриклеточный ацидоз - Появление свободных радикалов - Дефицит макроэргов Выделение энергии (тепловой, электрической – потенциал
- 18. Строение мембраны
- 19. Основные функции мембран Контролируют состав внутриклеточной среды Обеспечивают и облегчают межклеточную и и внутриклеточную передачу информации
- 20. Перенос веществ через мембраны Трансмембранный транспорт мелких молекул -фильтрация фагоцитоз и пиноцитоз наличие аквапоринов, или "водных
- 21. Аквапорины избирательно пропускают молекулы воды, позволяя ей поступать в клетку и покидать ее, в то же
- 22. Питер Эгр аквапорин
- 23. Функции аквапоринов Аквапорин - 1 в эритроцитах способствует регидратации эритроцитов, обезвоженных в гипертонической среде капилляров мозговой
- 24. Патология аквапоринов Болезнь Девика (БД, оптический нейромиелит) — одновременное или последовательное развитие неврита зрительных нервов и
- 25. Патология аквапоринов С возрастом, а также под воздействием ультрафиолета количество аквапоринов в клетках кожи сокращается. Объем
- 26. мембрана Внутреннее пространство Внешнее пространство Простая диффузия (СО2, мочевина) Белок переносчик Облегченная диффузия(аминокислоты, моносахара) АТФ Активный
- 27. Облегченная диффузия Канал всегда открыт, вещество проходит по градиенту концентрации или электрохимическому градиенту
- 28. Связывание вещества белком-переносчиком специфично и может быть конкурентно ингибировано этот процесс может быть лимитирован насыщением белка
- 29. Переносчики глюкозы — интегральные гликопротеины. Известно 6 кодируемых генами GLUT - трансмембранных переносчиков глюкозы из внеклеточной
- 31. Перенос веществ через мембраны Ионные каналы- это интегральные белки, найденные во всех клетках, которые регулируют прохождение
- 32. Выделяют 4 типа ионных каналов. 1. управляемые лигандом, который соединяется с рецептором (каналы для кальция) 2.
- 33. Потенциалзависимый ионный канал.
- 34. Селективный ионный канал Механочувствительный ионный канал
- 35. Активность разных каналов может изменяться под воздействием различных факторов: метаболических реакций, фосфолирирования, эффектов токсинов и лекарственных
- 36. В других случаях лекарственные препараты могут регулировать внутриклеточные процессы, например, сердечные глюкозиды влияют на характер сокращения
- 37. Обеспечивают межклеточную информацию, межклеточное взаимодействие феномен контактного торможения у нормальных клеток отсутствие этого феномена у опухолевых
- 38. Группа ученых университета Рочестера под руководством молекулярного биолога Веры Горбуновой выяснили механизм необычной устойчивости к развитию
- 39. Их возраст может превышать 30 лет – однако даже у самых «взрослых» особей не происходит спонтанного
- 40. Коннекси́ны — «двойные» поры, получающиеся за счёт совмещения друг с другом пор, принадлежащих контактирующим мембранам двух
- 41. Коннексины —интегральные мембранные белки образуют в мембранахобразуют в мембранах контактирующих клетокобразуют в мембранах контактирующих клеток сквозные
- 42. Патология коннексинов Соматические мутации в гене, кодирующем gap-junction protein connexin 40 (GJA5), миокардиальный белок, участвующий в
- 43. Патология коннексинов КОННЕКСиН-26. Ген коннексина-26 находит Ген коннексина-26 находится на 13-й хромосоме. Делеционная мутация мутация гена
- 44. Обеспечивают передачу информации Через поверхностные клеточные рецепторы Через интернализацию сигнала (рецептор-лиганд, инсулин) Перенос к внутриклеточным рецепторам
- 46. Клеточные мембранные антигены используются иммунной системой для разделения клеток на «свой» и «чужой» Антигены гистосовместимости
- 47. Антигены групп крови Модель мембраны эритроцита со встроенными молекулами групп крови разных систем. Таких систем на
- 48. Три важных принципа строения мембраны Мембраны не однородны Многие компоненты мембран находятся в непрерывном движении Компоненты
- 49. Липиды и белки формируют подвижную мозаичную мембрану. - молекулы липидов могут перемещаться параллельно поверхности мембраны, определяя
- 50. Текучие свойства мембраны определяются составом мембраны, так ненасыщенные жирные кислоты повышают текучесть холестерол ограничивает подвижность соседних
- 51. От текучести мембраны зависит проникновение воды, кислорода, углекислого газа, наркотических средств и т.д.
- 52. Компоненты мембран асимметричны фосфолипиды распределены асимметрично между внутренним и наружным монослоями цитоплазматической мембраны: в наружном монослое
- 53. В мембранах содержатся липиды трех классов: фосфолипиды, холестерин и гликолипиды. Наиболее важная группа, фосфолипиды, включает фосфатидилхолин
- 54. Нарушение метаболизма сфинголипидов (пример) Болезнь Тея — Сакса —наследственное заболевание нервной системы. Названо в честь британского
- 55. Новорожденные с данным заболеванием развиваются нормально в первые месяцы жизни. В возрасте около полугода возникает регресс
- 56. Для Болезни Тея—Сакса характерно наличие красного пятна, расположенного на сетчатке напротив зрачка. Это пятно можно увидеть
- 57. Некоторые токсины (холера, столбняк) способны вступать во взаимодействие с ганглиозидами мембран, пространственно или химически блокируя функции
- 58. Повреждение мембраны Хлорированные углеводороды могут приводить к фрагментации мембран адсорбция холестерола может привести к изменению текучих
- 59. Нарушения мембраны могут быть обусловлены свободно-радикальным (перекисным) окислением ненасыщенных жирных кислот в мембранных фосфолипидах. - действием
- 60. Следствием активации СРО: окисление сульфигидрильных групп мембранных белков, что может привести к помутнению хрусталика глаза; инактивация
- 61. Увеличение проницаемости мембраны клетки и выключение насосов приводит к утечке Са из митохондрий, возрастанию его в
- 62. Исходы повреждения Некроз- смерть поврежденной клетки, как завершающий этап клеточной дистрофии или как следствие действия повреждающего
- 63. Когда в делящихся клетках регенерирующих тканей накапливаются необратимые повреждения клетки прибегают к двум основным механизмам -
- 64. Зачем клетка погибает Конечное дифференцирование Клетка исчерпала "лимит Хейфлика" (состарилась) Апоптоз, аутофагия Клетка стала ненужной организму
- 65. Навсегда остановить клеточный цикл и войти в состояние покоя - сенесценцию Запустить механизмы запрограммированной смерти Следствия
- 66. Стимулы способствующие переходу клетки в состояние покоя – укорочение теломер, повреждение ДНК, оксидативный стресс и другие.
- 67. Теломеры (от др.-греч. τέλος — конец и μέρος — часть) концевые участки хромосом. В каждом цикле
- 68. Аутофагия – самопоедание Аутофагия - механизм ликвидации поврежденных органелл, долгоживущих и аномальных белков, излишних объёмов цитоплазмы.
- 70. Апоптоз- генетически активная программа клеточного суицида альтернативная клеточному делению. Морфологические признаки апоптоза – сморщивание и конденсация
- 71. Апоптоз - форма гибели клетки, проявляющаяся в уменьшении ее размера, конденсации (уплотнении) и фрагментации хроматина -
- 72. Апаптоз завершается фагоцитозом клеточных фрагментов. Главное значение апоптоза в норме удаление поврежденных, старых клеток. Уничтожение лишних
- 74. Усиление апоптоза Дефекты развития. .Дегенерация коронарной и легочной артерий, аномальная инволюция правого желудочка через усиление апоптоза
- 75. Ослабление апоптоза
- 76. Программа апоптоза включается через эндогенные сигналы - при невозможности репарации ДНК повреждении митохондрий выходу из митохондрий
- 77. Сенсором повреждения является Р53 ген кодирущий этот белок, находится в коротком плече 17 хромосомы. Его называют
- 78. Мутации гена р53 приводят к заболеваниям, например, мутации гена Р53 обнаруживаются в 50% раковых опухолей, что
- 79. Программу апоптоза запускает экзогенный сигнал, например ФНО или Т- киллеры. дефицит веществ необходимых для клетки. При
- 80. Информационный сигнал способствует активации протеолитических ферментов – каспаз Каспазы расщепляют белки в местах расположения аспарагиновых оснований.
- 82. Скачать презентацию

Рудольф Вирхов — основатель целлюлярной (клеточной) патологии
1821-1902
"omnis ceilula е cellula" (клетка происходит
Рудольф Вирхов — основатель целлюлярной (клеточной) патологии
1821-1902
"omnis ceilula е cellula" (клетка происходит

1858 год
«Целлюлярная патология как учение, основанное на физиологической и патологической
1858 год
«Целлюлярная патология как учение, основанное на физиологической и патологической

Основной постулат труда Р.Вирхова -
Всякое болезненное изменение организма связано с
Основной постулат труда Р.Вирхова -
Всякое болезненное изменение организма связано с

Причины повреждения клеток
Физические факторы:
-Механические
-Температура
-Ионизирующая радиация
-Ультрафиолетовое излучение
-Электромагнитные
Причины повреждения клеток
Физические факторы:
-Механические
-Температура
-Ионизирующая радиация
-Ультрафиолетовое излучение
-Электромагнитные

Химические факторы
органические и неорганические кислоты, щелочи, соли тяжелых металлов, продукты нарушенного
Химические факторы
органические и неорганические кислоты, щелочи, соли тяжелых металлов, продукты нарушенного

Биологические факторы
Наибольшее значение среди них имеют вирусы, риккетсии, бактерии, паразиты, грибы
Биологические факторы
Наибольшее значение среди них имеют вирусы, риккетсии, бактерии, паразиты, грибы

И другие факторы повреждения
Гипоксия
Гипероксия
Иммунологические
Нарушение КОС
Нарушение водно-электролитного обмена
Нарушение углеводного обмена
Генетические дефекты
Дисбаланс питания
Старение
И другие факторы повреждения
Гипоксия
Гипероксия
Иммунологические
Нарушение КОС
Нарушение водно-электролитного обмена
Нарушение углеводного обмена
Генетические дефекты
Дисбаланс питания
Старение

Реакция клеток на повреждение
нарушение функции клеток
нарушение клеточных защитных реакций – (активация
Реакция клеток на повреждение
нарушение функции клеток
нарушение клеточных защитных реакций – (активация

Изменение метаболизма
- нарушение функции транспортных систем клеточной мембраны
- повышение проницаемости внутриклеточных
Изменение метаболизма
- нарушение функции транспортных систем клеточной мембраны
- повышение проницаемости внутриклеточных

-нарушение дифференцировки и роста клеток (опухолевый рост)
-патология репродукции клеток
-патология движения клеток
-патология
-нарушение дифференцировки и роста клеток (опухолевый рост)
-патология репродукции клеток
-патология движения клеток
-патология

Вывод: повреждение клетки - это нарушение клеточного обмена веществ, внутриклеточного гомеостазиса.
Вывод: повреждение клетки - это нарушение клеточного обмена веществ, внутриклеточного гомеостазиса.
Повреждения
летальные
сублетальные
Специфические и неспецифические
Повреждения
летальные
сублетальные
Специфические и неспецифические

Основные механизмы острого летального повреждения
- чрезмерное подавление синтеза АТФ
-
Основные механизмы острого летального повреждения
- чрезмерное подавление синтеза АТФ
-

Специфическое повреждение -определяется этиологическим фактором
Иммунный гемолиз, радиационное повреждение свободными радикалами, повреждение
Специфическое повреждение -определяется этиологическим фактором
Иммунный гемолиз, радиационное повреждение свободными радикалами, повреждение

Неспецифическое повреждение
Утрата принципа неравновесности
К в клетке в 20-30 раз выше, чем
Неспецифическое повреждение
Утрата принципа неравновесности
К в клетке в 20-30 раз выше, чем

- Внутриклеточный ацидоз
- Появление свободных радикалов
- Дефицит макроэргов
Выделение энергии (тепловой, электрической
- Внутриклеточный ацидоз
- Появление свободных радикалов
- Дефицит макроэргов
Выделение энергии (тепловой, электрической
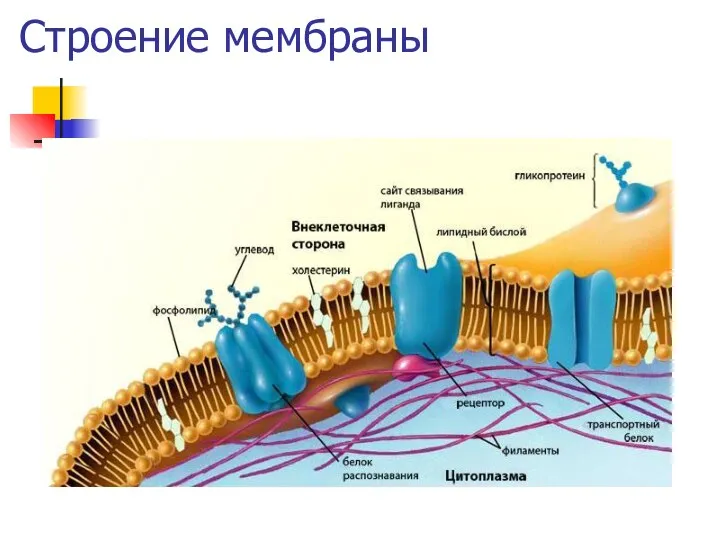
Строение мембраны
Строение мембраны

Основные функции мембран
Контролируют состав внутриклеточной среды
Обеспечивают и облегчают межклеточную и и
Основные функции мембран
Контролируют состав внутриклеточной среды
Обеспечивают и облегчают межклеточную и и

Перенос веществ через мембраны
Трансмембранный транспорт мелких молекул
-фильтрация
фагоцитоз и пиноцитоз
наличие аквапоринов, или
Перенос веществ через мембраны
Трансмембранный транспорт мелких молекул
-фильтрация
фагоцитоз и пиноцитоз
наличие аквапоринов, или

Аквапорины
избирательно пропускают молекулы воды, позволяя ей поступать в клетку и покидать
Аквапорины
избирательно пропускают молекулы воды, позволяя ей поступать в клетку и покидать
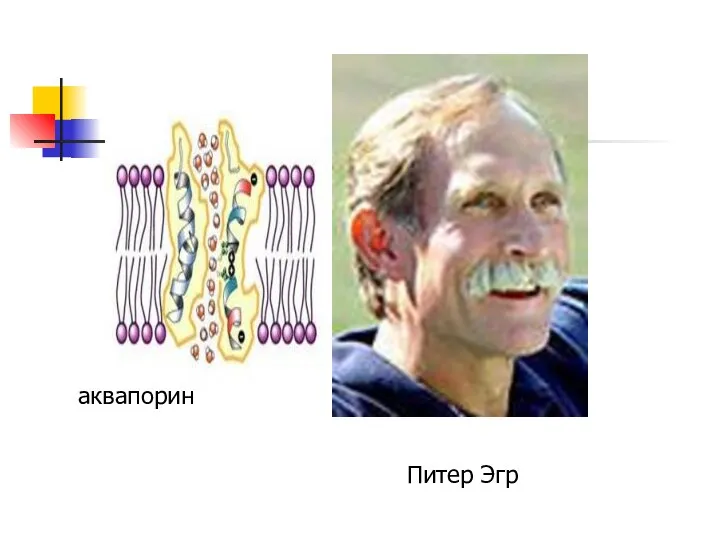
Питер Эгр
аквапорин
Питер Эгр
аквапорин

Функции аквапоринов
Аквапорин - 1 в эритроцитах способствует регидратации эритроцитов, обезвоженных в
Функции аквапоринов
Аквапорин - 1 в эритроцитах способствует регидратации эритроцитов, обезвоженных в

Патология аквапоринов
Болезнь Девика (БД, оптический нейромиелит) — одновременное или последовательное развитие
Патология аквапоринов
Болезнь Девика (БД, оптический нейромиелит) — одновременное или последовательное развитие

Патология аквапоринов
С возрастом, а также под воздействием ультрафиолета количество аквапоринов в
Патология аквапоринов
С возрастом, а также под воздействием ультрафиолета количество аквапоринов в

мембрана
Внутреннее пространство
Внешнее
пространство
Простая диффузия (СО2, мочевина)
Белок переносчик
Облегченная диффузия(аминокислоты, моносахара)
АТФ
Активный транспорт –
мембрана
Внутреннее пространство
Внешнее
пространство
Простая диффузия (СО2, мочевина)
Белок переносчик
Облегченная диффузия(аминокислоты, моносахара)
АТФ
Активный транспорт –
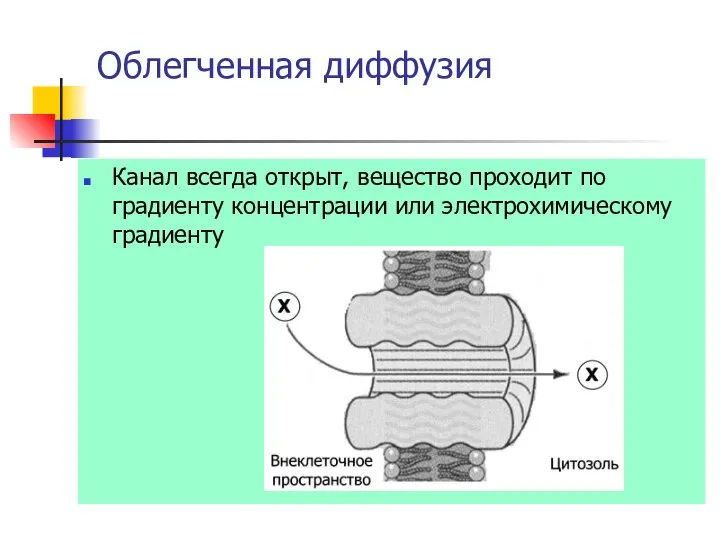
Облегченная диффузия
Канал всегда открыт, вещество проходит по градиенту концентрации или электрохимическому
Облегченная диффузия
Канал всегда открыт, вещество проходит по градиенту концентрации или электрохимическому

Связывание вещества белком-переносчиком специфично
и может быть конкурентно ингибировано
этот процесс может
Связывание вещества белком-переносчиком специфично
и может быть конкурентно ингибировано
этот процесс может

Переносчики глюкозы — интегральные гликопротеины.
Известно 6 кодируемых генами GLUT - трансмембранных
Переносчики глюкозы — интегральные гликопротеины.
Известно 6 кодируемых генами GLUT - трансмембранных


Перенос веществ через мембраны
Ионные каналы- это интегральные белки, найденные во всех
Перенос веществ через мембраны
Ионные каналы- это интегральные белки, найденные во всех

Выделяют 4 типа ионных каналов.
1. управляемые лигандом, который соединяется с рецептором
Выделяют 4 типа ионных каналов.
1. управляемые лигандом, который соединяется с рецептором

Потенциалзависимый ионный канал.
Потенциалзависимый ионный канал.
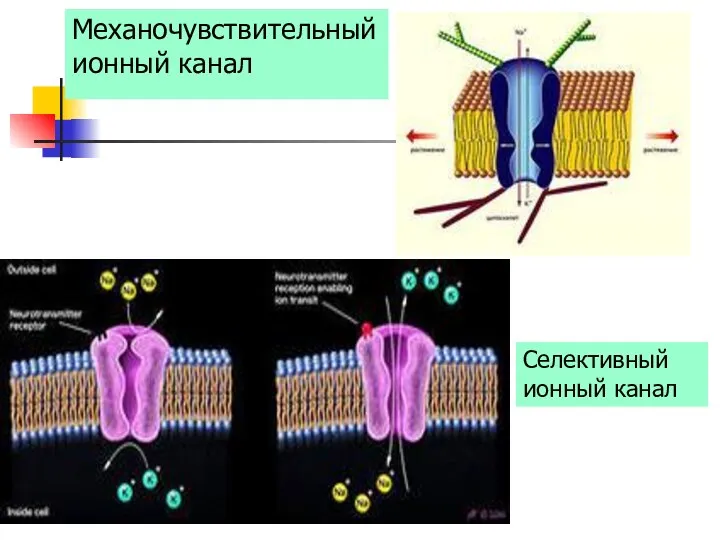
Селективный ионный канал
Механочувствительный ионный канал
Селективный ионный канал
Механочувствительный ионный канал

Активность разных каналов может изменяться
под воздействием различных факторов: метаболических реакций, фосфолирирования,
Активность разных каналов может изменяться
под воздействием различных факторов: метаболических реакций, фосфолирирования,

В других случаях
лекарственные препараты могут регулировать внутриклеточные процессы, например, сердечные глюкозиды
В других случаях
лекарственные препараты могут регулировать внутриклеточные процессы, например, сердечные глюкозиды

Обеспечивают межклеточную информацию, межклеточное взаимодействие
феномен контактного торможения у нормальных клеток
отсутствие
Обеспечивают межклеточную информацию, межклеточное взаимодействие
феномен контактного торможения у нормальных клеток
отсутствие

Группа ученых университета Рочестера
под руководством молекулярного биолога Веры Горбуновой выяснили механизм
Группа ученых университета Рочестера
под руководством молекулярного биолога Веры Горбуновой выяснили механизм
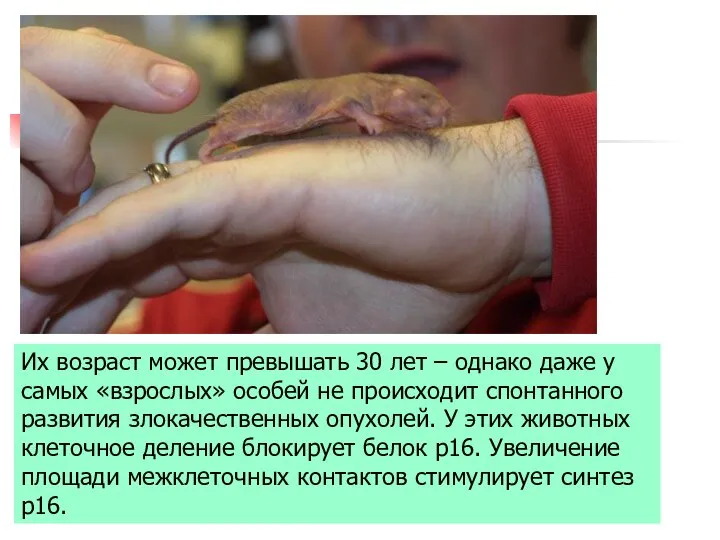
Их возраст может превышать 30 лет – однако даже у самых
Их возраст может превышать 30 лет – однако даже у самых
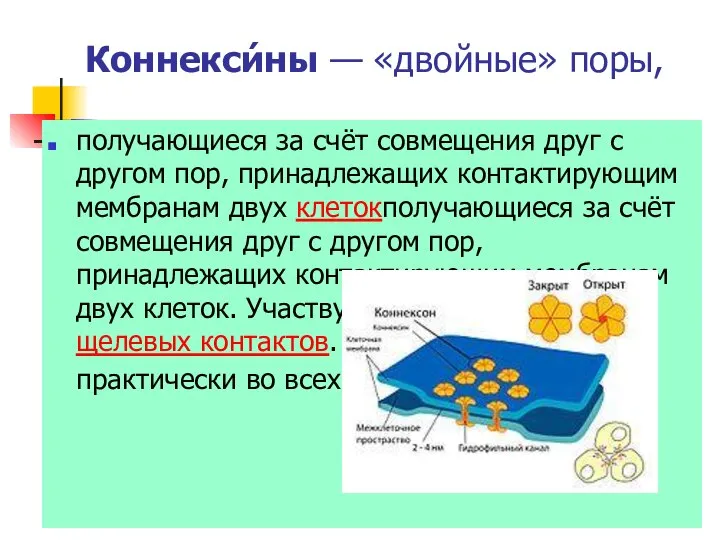
Коннекси́ны — «двойные» поры,
получающиеся за счёт совмещения друг с другом пор, принадлежащих
Коннекси́ны — «двойные» поры,
получающиеся за счёт совмещения друг с другом пор, принадлежащих

Коннексины —интегральные мембранные белки
образуют в мембранахобразуют в мембранах контактирующих клетокобразуют в мембранах
Коннексины —интегральные мембранные белки
образуют в мембранахобразуют в мембранах контактирующих клетокобразуют в мембранах

Патология коннексинов
Соматические мутации в гене, кодирующем gap-junction protein connexin 40 (GJA5),
Патология коннексинов
Соматические мутации в гене, кодирующем gap-junction protein connexin 40 (GJA5),

Патология коннексинов
КОННЕКСиН-26.
Ген коннексина-26 находит Ген коннексина-26 находится на 13-й хромосоме.
Патология коннексинов
КОННЕКСиН-26.
Ген коннексина-26 находит Ген коннексина-26 находится на 13-й хромосоме.

Обеспечивают передачу информации
Через поверхностные клеточные рецепторы
Через интернализацию сигнала (рецептор-лиганд, инсулин)
Обеспечивают передачу информации
Через поверхностные клеточные рецепторы
Через интернализацию сигнала (рецептор-лиганд, инсулин)

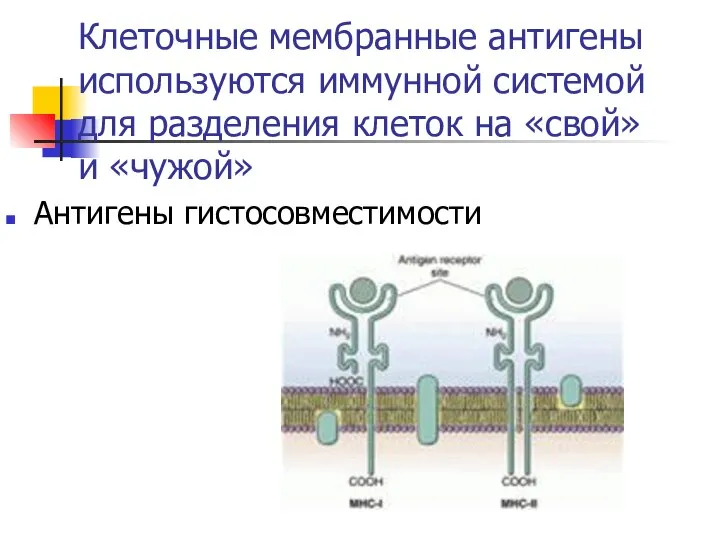
Клеточные мембранные антигены используются иммунной системой для разделения клеток на «свой»
Клеточные мембранные антигены используются иммунной системой для разделения клеток на «свой»
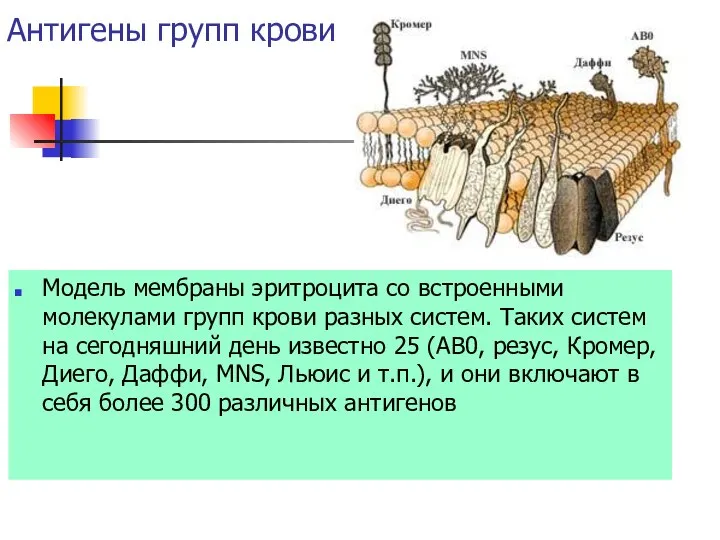
Антигены групп крови
Модель мембраны эритроцита со встроенными молекулами групп крови разных
Антигены групп крови
Модель мембраны эритроцита со встроенными молекулами групп крови разных

Три важных принципа строения мембраны
Мембраны не однородны
Многие компоненты мембран находятся в
Три важных принципа строения мембраны
Мембраны не однородны
Многие компоненты мембран находятся в

Липиды и белки формируют подвижную мозаичную мембрану.
- молекулы липидов могут перемещаться
параллельно
Липиды и белки формируют подвижную мозаичную мембрану.
- молекулы липидов могут перемещаться
параллельно

Текучие свойства мембраны определяются
составом мембраны, так ненасыщенные жирные кислоты повышают текучесть
Текучие свойства мембраны определяются
составом мембраны, так ненасыщенные жирные кислоты повышают текучесть

От текучести мембраны
зависит проникновение воды, кислорода, углекислого газа, наркотических средств и
От текучести мембраны
зависит проникновение воды, кислорода, углекислого газа, наркотических средств и

Компоненты мембран асимметричны
фосфолипиды распределены асимметрично между внутренним и наружным монослоями цитоплазматической
Компоненты мембран асимметричны
фосфолипиды распределены асимметрично между внутренним и наружным монослоями цитоплазматической

В мембранах содержатся липиды трех классов: фосфолипиды, холестерин и гликолипиды.
Наиболее
В мембранах содержатся липиды трех классов: фосфолипиды, холестерин и гликолипиды.
Наиболее

Нарушение метаболизма сфинголипидов (пример)
Болезнь Тея — Сакса —наследственное заболевание нервной системы. Названо
Нарушение метаболизма сфинголипидов (пример)
Болезнь Тея — Сакса —наследственное заболевание нервной системы. Названо

Новорожденные с данным заболеванием развиваются нормально в первые месяцы жизни.
В возрасте
Новорожденные с данным заболеванием развиваются нормально в первые месяцы жизни.
В возрасте

Для Болезни Тея—Сакса характерно
наличие красного пятна, расположенного на сетчатке напротив зрачка.
Для Болезни Тея—Сакса характерно
наличие красного пятна, расположенного на сетчатке напротив зрачка.

Некоторые токсины (холера, столбняк)
способны вступать во взаимодействие с ганглиозидами мембран, пространственно
Некоторые токсины (холера, столбняк)
способны вступать во взаимодействие с ганглиозидами мембран, пространственно

Повреждение мембраны
Хлорированные углеводороды могут приводить к фрагментации мембран
адсорбция холестерола может привести
Повреждение мембраны
Хлорированные углеводороды могут приводить к фрагментации мембран
адсорбция холестерола может привести

Нарушения мембраны могут быть обусловлены
свободно-радикальным (перекисным) окислением ненасыщенных жирных кислот
Нарушения мембраны могут быть обусловлены
свободно-радикальным (перекисным) окислением ненасыщенных жирных кислот

Следствием активации СРО:
окисление сульфигидрильных групп мембранных белков, что может привести к
Следствием активации СРО:
окисление сульфигидрильных групп мембранных белков, что может привести к

Увеличение проницаемости мембраны клетки и выключение насосов
приводит к утечке Са из
Увеличение проницаемости мембраны клетки и выключение насосов
приводит к утечке Са из
Исходы повреждения
Некроз- смерть поврежденной клетки, как завершающий этап клеточной дистрофии
Исходы повреждения
Некроз- смерть поврежденной клетки, как завершающий этап клеточной дистрофии

Когда в делящихся клетках регенерирующих тканей накапливаются необратимые повреждения
клетки прибегают к
Когда в делящихся клетках регенерирующих тканей накапливаются необратимые повреждения
клетки прибегают к
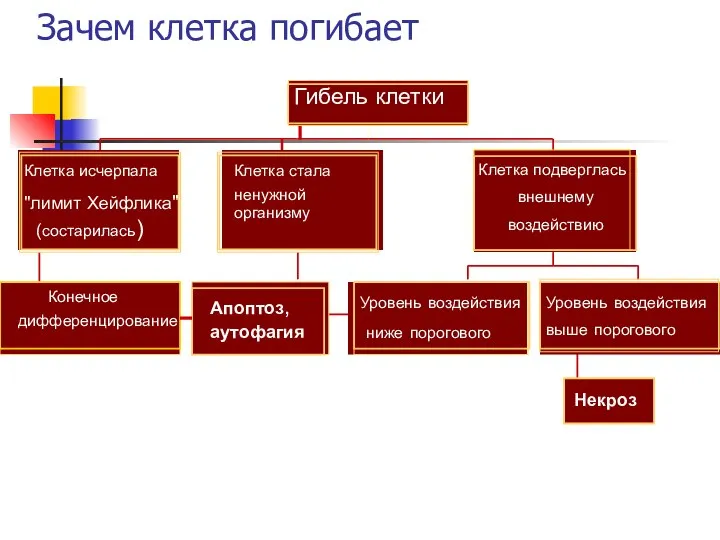
Зачем клетка погибает
Конечное
дифференцирование
Клетка исчерпала
"лимит Хейфлика"
(состарилась)
Апоптоз,
аутофагия
Клетка стала
ненужной
организму
Уровень воздействия
ниже порогового
Некроз
Уровень воздействия
выше порогового
Зачем клетка погибает
Конечное
дифференцирование
Клетка исчерпала
"лимит Хейфлика"
(состарилась)
Апоптоз,
аутофагия
Клетка стала
ненужной
организму
Уровень воздействия
ниже порогового
Некроз
Уровень воздействия
выше порогового

Навсегда остановить клеточный цикл и войти в состояние покоя - сенесценцию
Запустить
Навсегда остановить клеточный цикл и войти в состояние покоя - сенесценцию
Запустить

Стимулы способствующие переходу клетки в состояние покоя –
укорочение теломер, повреждение ДНК,
Стимулы способствующие переходу клетки в состояние покоя –
укорочение теломер, повреждение ДНК,

Теломеры (от др.-греч. τέλος — конец и μέρος — часть) концевые участки хромосом.
Теломеры (от др.-греч. τέλος — конец и μέρος — часть) концевые участки хромосом.

Аутофагия – самопоедание
Аутофагия - механизм ликвидации поврежденных органелл, долгоживущих и аномальных
Аутофагия – самопоедание
Аутофагия - механизм ликвидации поврежденных органелл, долгоживущих и аномальных


Апоптоз-
генетически активная программа клеточного суицида альтернативная клеточному делению.
Морфологические признаки апоптоза –
Апоптоз-
генетически активная программа клеточного суицида альтернативная клеточному делению.
Морфологические признаки апоптоза –

Апоптоз
- форма гибели клетки, проявляющаяся в уменьшении ее размера, конденсации (уплотнении)
Апоптоз
- форма гибели клетки, проявляющаяся в уменьшении ее размера, конденсации (уплотнении)
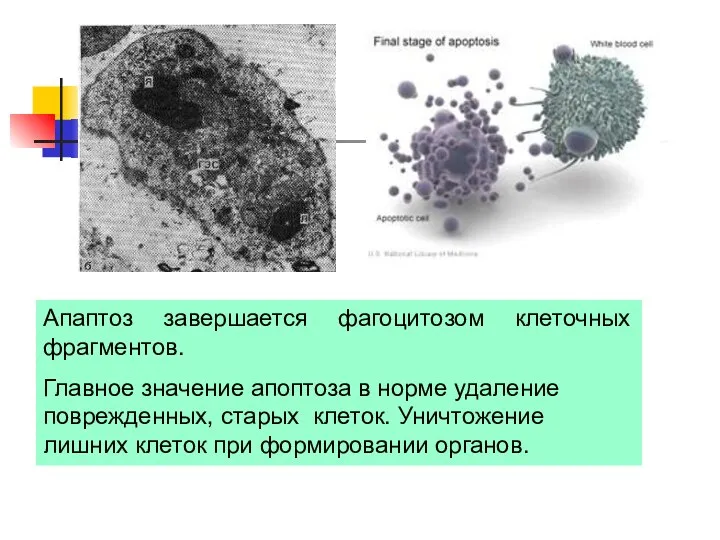
Апаптоз завершается фагоцитозом клеточных фрагментов.
Главное значение апоптоза в норме удаление поврежденных,
Апаптоз завершается фагоцитозом клеточных фрагментов.
Главное значение апоптоза в норме удаление поврежденных,


Усиление апоптоза
Дефекты развития.
.Дегенерация коронарной и легочной артерий, аномальная инволюция правого желудочка
Усиление апоптоза
Дефекты развития.
.Дегенерация коронарной и легочной артерий, аномальная инволюция правого желудочка

Ослабление апоптоза
Ослабление апоптоза

Программа апоптоза включается
через эндогенные сигналы - при невозможности репарации ДНК
повреждении митохондрий
Программа апоптоза включается
через эндогенные сигналы - при невозможности репарации ДНК
повреждении митохондрий

Сенсором повреждения является Р53
ген кодирущий этот белок, находится в коротком плече
Сенсором повреждения является Р53
ген кодирущий этот белок, находится в коротком плече

Мутации гена р53 приводят к заболеваниям, например,
мутации гена Р53 обнаруживаются в
Мутации гена р53 приводят к заболеваниям, например,
мутации гена Р53 обнаруживаются в

Программу апоптоза запускает
экзогенный сигнал, например ФНО или Т- киллеры.
Программу апоптоза запускает
экзогенный сигнал, например ФНО или Т- киллеры.

Информационный сигнал
способствует активации протеолитических ферментов – каспаз
Каспазы расщепляют белки в местах
Информационный сигнал
способствует активации протеолитических ферментов – каспаз
Каспазы расщепляют белки в местах
 Lektsia__3_NG_18
Lektsia__3_NG_18 Прийменники з подвійним керуванням
Прийменники з подвійним керуванням Всё о неравенствах Работу выполнил Попов Игорь ученик 9-класса
Всё о неравенствах Работу выполнил Попов Игорь ученик 9-класса  proect
proect Аттестационная работа. Методика по выполнению проектно – исследовательской работы с учащимися 8 класса «Нации села Каразей»
Аттестационная работа. Методика по выполнению проектно – исследовательской работы с учащимися 8 класса «Нации села Каразей» Буддизм
Буддизм Дипломатия второй половины ХХ века
Дипломатия второй половины ХХ века Аппаратное обеспечение ПК
Аппаратное обеспечение ПК Разработка технологии управления конфликтами на предприятии в условиях кризиса ПАО «Уралсиб»
Разработка технологии управления конфликтами на предприятии в условиях кризиса ПАО «Уралсиб» Переломы челюстей
Переломы челюстей  Схема разрывной машины с рычажно-маятниковым силоизмерителем
Схема разрывной машины с рычажно-маятниковым силоизмерителем Мода XVIII века. Дворянство
Мода XVIII века. Дворянство Презентация "Пушно-меховые и овчинно-шубные изделия" - скачать презентации по Экономике
Презентация "Пушно-меховые и овчинно-шубные изделия" - скачать презентации по Экономике Презентация РЕЧЬ ОРАТОРА В СУДЕ!!!
Презентация РЕЧЬ ОРАТОРА В СУДЕ!!! Реакция нейтрализации
Реакция нейтрализации Педагогические технологии личностно- ориентированного и развивающего обучения
Педагогические технологии личностно- ориентированного и развивающего обучения International Captain’s summit (HMBIA)
International Captain’s summit (HMBIA) Международно-правовые основы борьбы с терроризмом. Сравнение УК РФ и США
Международно-правовые основы борьбы с терроризмом. Сравнение УК РФ и США Строительные конструкции. Расчет прочности изгибаемых элементов
Строительные конструкции. Расчет прочности изгибаемых элементов Разработка опытного образца экзоскелета
Разработка опытного образца экзоскелета Элладская (Ахейская) Греция
Элладская (Ахейская) Греция Презентация на тему "Жанры живописи"
Презентация на тему "Жанры живописи" Казимир Северинович Малевич 23 февраля 1878 - Киев 15мая 1935 Ленинград
Казимир Северинович Малевич 23 февраля 1878 - Киев 15мая 1935 Ленинград Материаловедение. Свойства материалов, способы изготовления из них изделий
Материаловедение. Свойства материалов, способы изготовления из них изделий Культура Германии
Культура Германии рисуем пасхальное яйцо - презентация для начальной школы
рисуем пасхальное яйцо - презентация для начальной школы Великий и могучий русский язык
Великий и могучий русский язык Möchten + infinitiv
Möchten + infinitiv