- ПОДВИЖНОСТЬ ЛИПИДОВ В МЕМБРАНЕ

Содержание
- 2. ПОДВИЖНОСТЬ ЛИПИДОВ В МЕМБРАНЕ
- 3. ИЗМЕНЕНИЕ ОРИЕНТАЦИИ ПОЛЯРНЫХ ГОЛОВ ЛАТЕРАЛЬНОЕ ДВИЖЕНИЕ КОЛЕБАНИЯ АЦИЛЬНЫХ ЦЕПЕЙ ОБРАЗОВАНИЕ КИНКОВ И ПЕРЕМЕЩЕНИЕ ИХ ВДОЛЬ АЦИЛЬНЫХ
- 4. I – изменение ориентации полярных голов II – латеральная диффузия III – колебания жирнокислотных цепей IV
- 5. ДИНАМИКА ЛИПИДОВ В МЕМБРАНЕ а б в а) ротационная подвижность(10-9 с) б) латеральная подвижность(10-7 с) в)
- 6. УПАКОВКА ЛИПИДОВ В МЕМБРАНЕ, ПОДВИЖНОСТЬ ЖИРНОКИСЛОТНЫХ ХВОСТОВ, ОБРАЗОВАНИЕ КИНКОВ
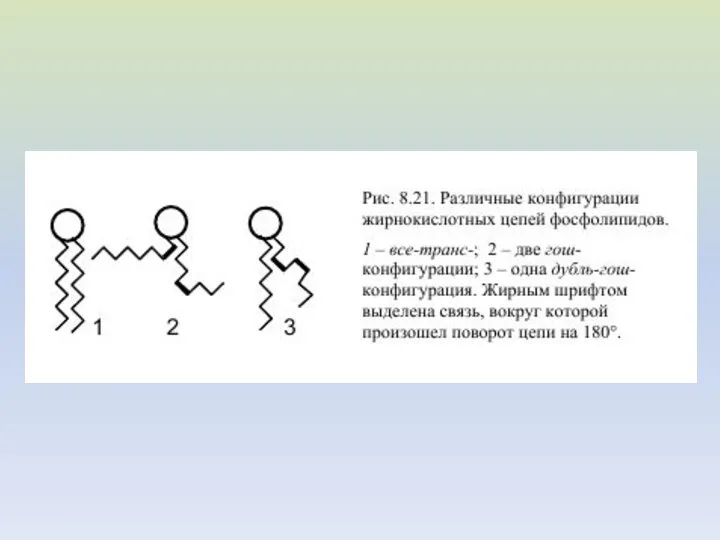
- 7. ПРОСТРАНСТВЕННАЯ КОНФОРМАЦИЯ ЖИРНЫХ КИСЛОТ 1 – насыщенная углеводородная цепь 2 – ненасыщенная цепь в цис-конформации 3
- 8. ИЗОМЕРИЯ ЖИРНЫХ КИСЛОТ ДВОЙНАЯ СВЯЗЬ
- 9. ОБРАЗОВАНИЕ ДЕФЕКТОВ В МЕМБРАННОМ БИСЛОЕ
- 10. ГЕОМЕТРИЯ БИСЛОЯ РАЗМЕРЫ МОЛЕКУЛЫ ЛИПИДА: ХВОСТ 2 НМ, ГОЛОВКА 0,5 – 0,7 НМ, РАССЧИТАННАЯ ТОЛЩИНА БИСЛОЯ
- 11. вращение вокруг С-С связей ничем не ограничено (I) образование скошенной (гош) конформации (II) или кинка (III)
- 12. I – кинки отсутствуют II – один кинк на жирнокислотную цепь III – два кинка на
- 13. ТРАНС-ГОШ-ИЗОМЕРИЗАЦИЯ – ИЗМЕНЕНИЕ КОНФОРМАЦИИ МОЛЕКУЛ ЗА СЧЕТ ПОВОРОТОВ ВОКРУГ ЕДИНИЧНЫХ С-С СВЯЗЕЙ ПРИ ПЕРЕХОДЕ ИЗ ТРАНС-
- 15. МЕХАНИЗМ ПЕРЕНОСА ИОНОВ ЧЕРЕЗ МЕМБРАНУ
- 16. ПЕРЕНОС ЧАСТИЦЫ ЧЕРЕЗ МЕМБРАНУ БЛАГОДАРЯ КИНКАМ На рисунке показаны не разные молекулы фосфолипидов в бислое, а
- 17. ФАЗОВЫЕ ПЕРЕХОДЫ В ЛИПИДНОМ БИСЛОЕ
- 18. Структуры, образуемые в водных суспензиях липидами, склонными к созданию ламеллярных образований (А) и небислойных гексагональных образований
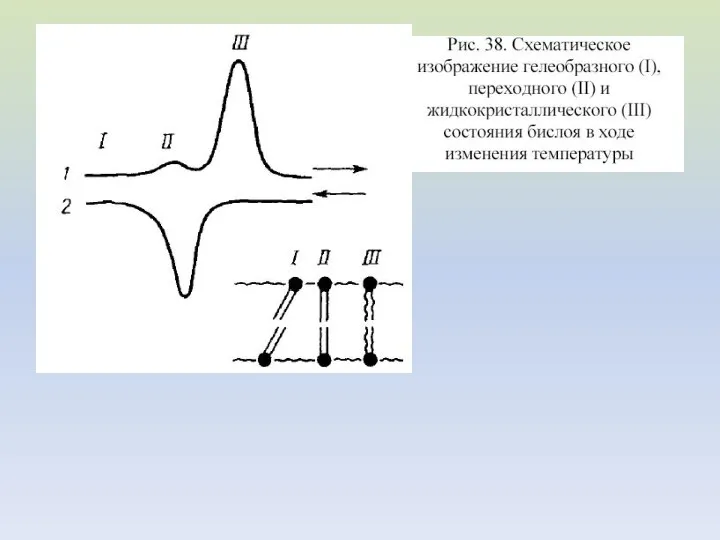
- 21. Схематическое изображение четырех фазовых состояний ламеллярного бислоя, образованного чистым димиристоилфосфатидилхолином Сплошная стрелка обозначает быстрые (порядка минут
- 22. РАЗДЕЛЕНИЕ ФАЗ В ГЕТЕРОГЕННОМ БИСЛОЕ (А) С ОДНОВРЕМЕННЫМ ФАЗОВЫМ ПЕРЕХОДОМ ЧАСТИ БИСЛОЯ (Б) ПОД ВЛИЯНИЕМ ТЕМПЕРАТУРЫ
- 24. МЕТОД ДИФФЕРЕНЦИАЛЬНОЙ СКАНИРУЮЩЕЙ МИКРОКАЛОРИМЕТРИИ Метод, основанный на измерении разницы тепловых потоков, идущих от испытуемого образца и
- 25. Фазовые переходы в суспензии фосфолипидных липосом
- 27. Анализ кривых ДСК Пусть удельная теплота плавления липида равна Qm, количество липидов в образце - m
- 28. В интервале температур от T1 до текущей температуры T расплавится количество молей липида ml, и при
- 31. Перед приготовлением липосом к фосфолипидам было добавлено разное количество холестерина; его содержание в молярных процентах указано
- 32. ИЗМЕНЕНИЕ УПАКОВКИ БИСЛОЯ ПРИ ТЕРМОИНДУЦИРОВАННОМ ФАЗОВОМ ПЕРЕХОДЕ (от А к Б) и ПРИ ВСТРАИВАНИИ В БИСЛОЙ
- 33. Гигантские везикулы, состоящих из насыщенного (ДПФХ) и ненасыщенного (ДОФХ) фосфолипидов, а также холестерола. Образующиеся в везикулах
- 34. ФЛУОРЕСЦЕНТНЫЕ МЕТОДЫ ИССЛЕДОВАНИЯ ФЛУОРЕСЦЕНТНЫЙ АНАЛИЗ ДАЕТ ВОЗМОЖНОСТЬ ИССЛЕДОВАТЬ ПОДВИЖНОСТЬ ФОСФОЛИПИДОВ В МЕМБРАНЕ, ОЦЕНИТЬ МИКРОВЯЗКОСТЬ МЕМБРАН ПРИМЕРЫ
- 35. Измерение флуоресценции зондов
- 38. Влияние препаратов на спектры флуоресценции пирена в мембранах эритроцитов
- 39. СПИНОВЫЕ ЗОНДЫ НИТРОКСИЛЬНЫЕ РАДИКАЛЫ устойчивы в широком интервале температур (до 100-200 СС) способны вступать в хим.
- 40. СПИНОВЫЙ ЗОНД ТЕМПО
- 41. Химическая "прививка" метки к макромолекулам с реакционно способными группами
- 42. Реакции макромолекул с бирадикалами и спиновыми ловушками. Спиновая ловушка - соединение, образующее стабильные радикалы при взаимодействии
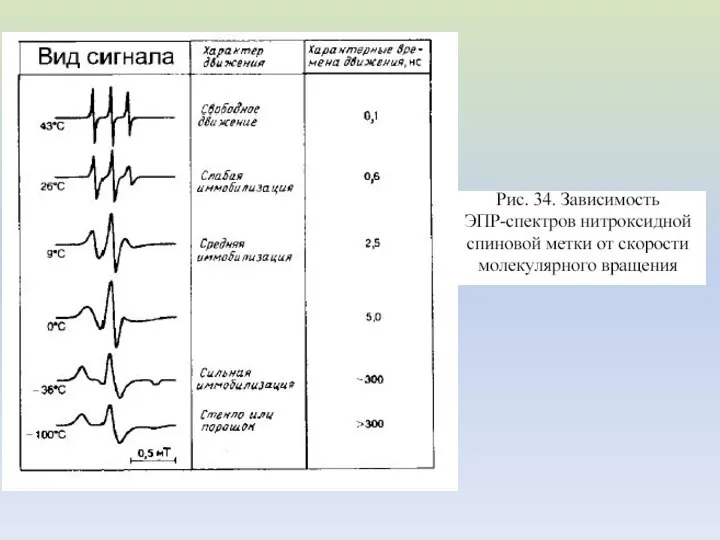
- 43. Спектры ЭПР Спектры ЭПР нитроксильных радикалов в вязких средах при временах корреляции вращения 5·10-10 с (a)
- 45. ИЗМЕНЕНИЕ СПЕКТРА ЭПР ПРИ УМЕНЬШЕНИИ МИКРОВЯЗКОСТИ η Н Н I
- 46. ДВА СПОСОБА ПРИКРЕПЛЕНИЯ СПИНОВОЙ МЕТКИ К МОЛЕКУЛЕ ФОСФОЛИПИДА И РАЗЛИЧИЕ СПЕКТРОВ ЭПР
- 47. Спектры 2Н-ЯМР димиристоилфофатидилхолина, дейтерированного по разным положениям ацильной цепи ЯМР
- 49. Скачать презентацию

ПОДВИЖНОСТЬ ЛИПИДОВ В МЕМБРАНЕ
ПОДВИЖНОСТЬ ЛИПИДОВ В МЕМБРАНЕ

ИЗМЕНЕНИЕ ОРИЕНТАЦИИ ПОЛЯРНЫХ ГОЛОВ
ЛАТЕРАЛЬНОЕ ДВИЖЕНИЕ
КОЛЕБАНИЯ АЦИЛЬНЫХ ЦЕПЕЙ
ОБРАЗОВАНИЕ КИНКОВ И ПЕРЕМЕЩЕНИЕ ИХ
ЛАТЕРАЛЬНОЕ ДВИЖЕНИЕ
КОЛЕБАНИЯ АЦИЛЬНЫХ ЦЕПЕЙ
ОБРАЗОВАНИЕ КИНКОВ И ПЕРЕМЕЩЕНИЕ ИХ
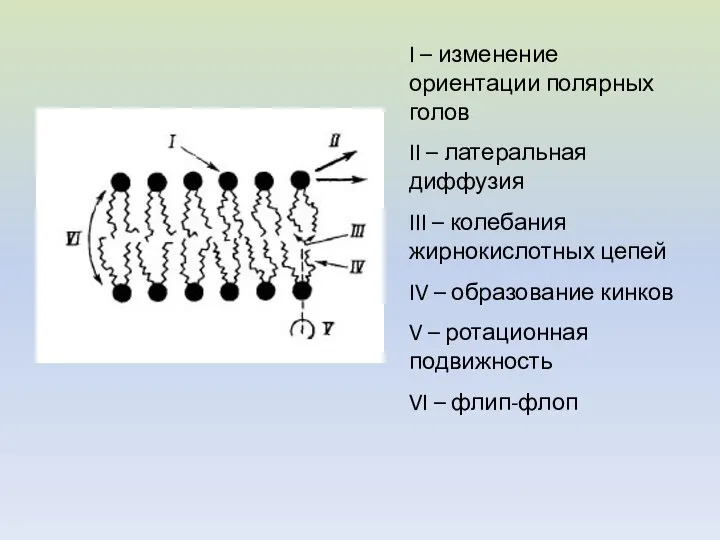
I – изменение ориентации полярных голов
II – латеральная диффузия
III – колебания
I – изменение ориентации полярных голов
II – латеральная диффузия
III – колебания
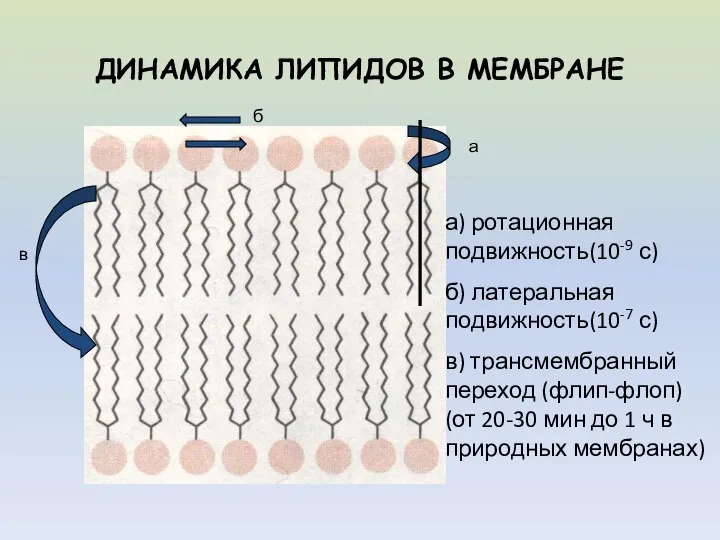
ДИНАМИКА ЛИПИДОВ В МЕМБРАНЕ
а
б
в
а) ротационная подвижность(10-9 с)
б) латеральная подвижность(10-7 с)
в) трансмембранный
ДИНАМИКА ЛИПИДОВ В МЕМБРАНЕ
а
б
в
а) ротационная подвижность(10-9 с)
б) латеральная подвижность(10-7 с)
в) трансмембранный

УПАКОВКА ЛИПИДОВ В МЕМБРАНЕ, ПОДВИЖНОСТЬ ЖИРНОКИСЛОТНЫХ ХВОСТОВ, ОБРАЗОВАНИЕ КИНКОВ
УПАКОВКА ЛИПИДОВ В МЕМБРАНЕ, ПОДВИЖНОСТЬ ЖИРНОКИСЛОТНЫХ ХВОСТОВ, ОБРАЗОВАНИЕ КИНКОВ
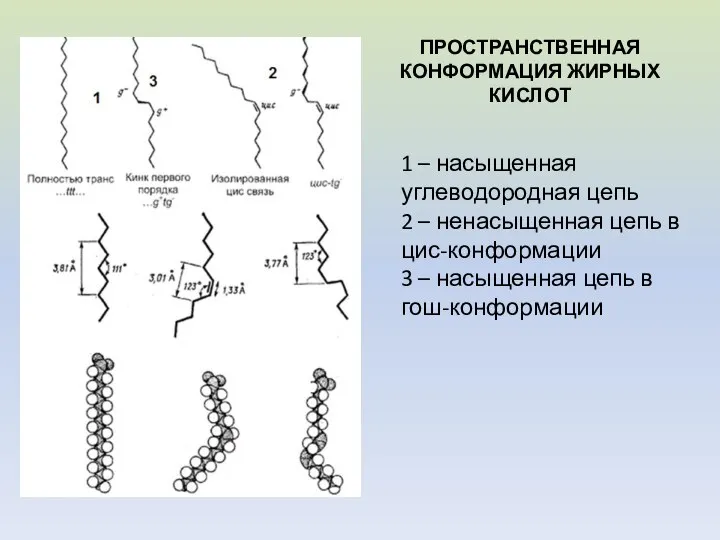
ПРОСТРАНСТВЕННАЯ КОНФОРМАЦИЯ ЖИРНЫХ КИСЛОТ
1 – насыщенная углеводородная цепь
2 – ненасыщенная цепь
ПРОСТРАНСТВЕННАЯ КОНФОРМАЦИЯ ЖИРНЫХ КИСЛОТ
1 – насыщенная углеводородная цепь
2 – ненасыщенная цепь
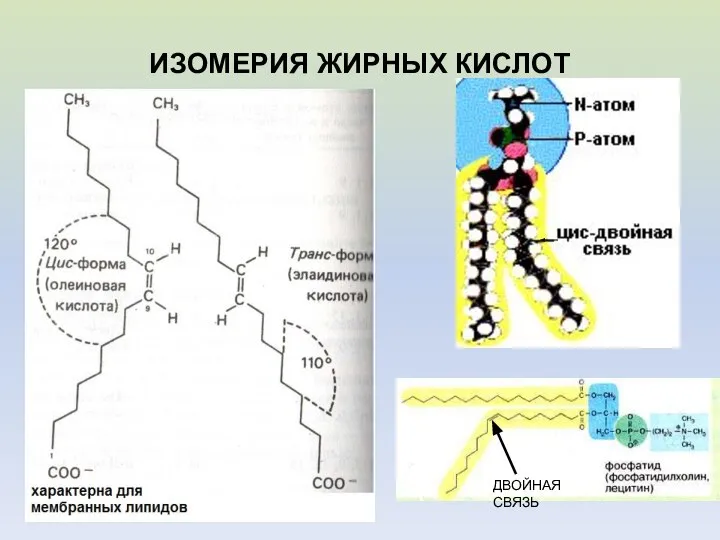
ИЗОМЕРИЯ ЖИРНЫХ КИСЛОТ
ДВОЙНАЯ СВЯЗЬ
ИЗОМЕРИЯ ЖИРНЫХ КИСЛОТ
ДВОЙНАЯ СВЯЗЬ

ОБРАЗОВАНИЕ ДЕФЕКТОВ В МЕМБРАННОМ БИСЛОЕ
ОБРАЗОВАНИЕ ДЕФЕКТОВ В МЕМБРАННОМ БИСЛОЕ

ГЕОМЕТРИЯ БИСЛОЯ
РАЗМЕРЫ МОЛЕКУЛЫ ЛИПИДА: ХВОСТ 2 НМ, ГОЛОВКА 0,5 – 0,7
ГЕОМЕТРИЯ БИСЛОЯ
РАЗМЕРЫ МОЛЕКУЛЫ ЛИПИДА: ХВОСТ 2 НМ, ГОЛОВКА 0,5 – 0,7
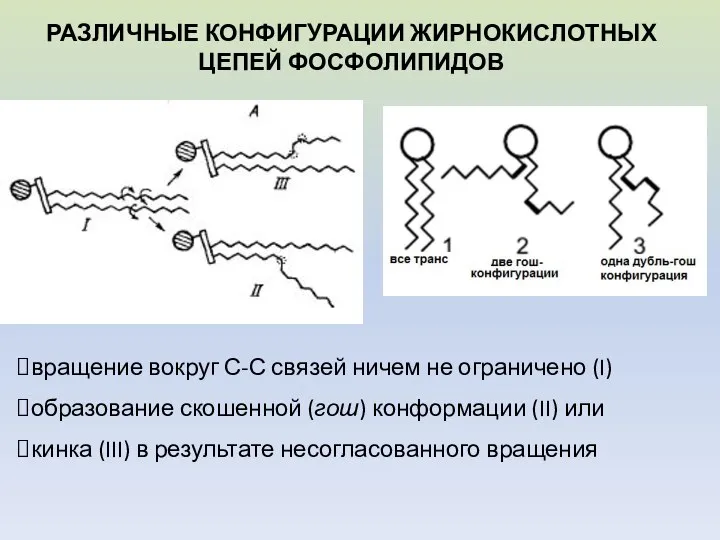
вращение вокруг С-С связей ничем не ограничено (I)
образование скошенной (гош) конформации
вращение вокруг С-С связей ничем не ограничено (I)
образование скошенной (гош) конформации
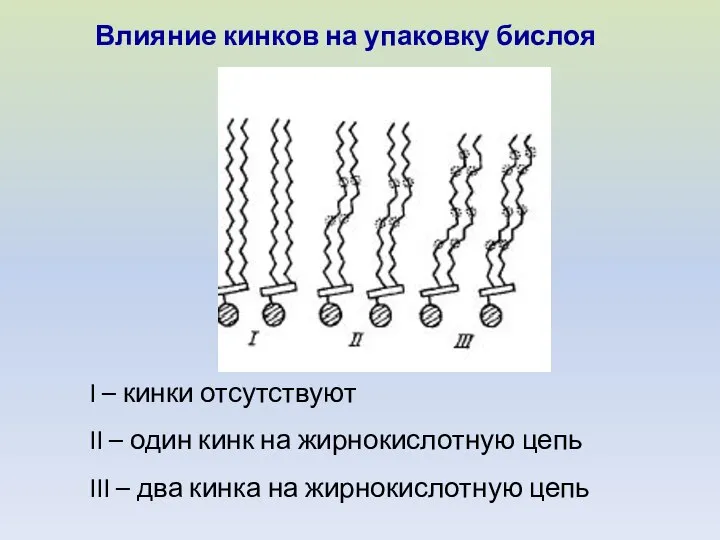
I – кинки отсутствуют
II – один кинк на жирнокислотную цепь
III –
I – кинки отсутствуют
II – один кинк на жирнокислотную цепь
III –
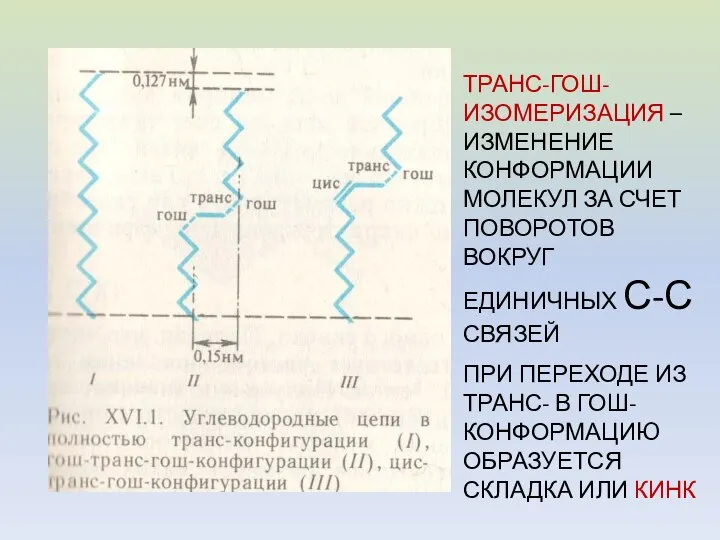
ТРАНС-ГОШ-ИЗОМЕРИЗАЦИЯ – ИЗМЕНЕНИЕ КОНФОРМАЦИИ МОЛЕКУЛ ЗА СЧЕТ ПОВОРОТОВ ВОКРУГ ЕДИНИЧНЫХ С-С
ТРАНС-ГОШ-ИЗОМЕРИЗАЦИЯ – ИЗМЕНЕНИЕ КОНФОРМАЦИИ МОЛЕКУЛ ЗА СЧЕТ ПОВОРОТОВ ВОКРУГ ЕДИНИЧНЫХ С-С
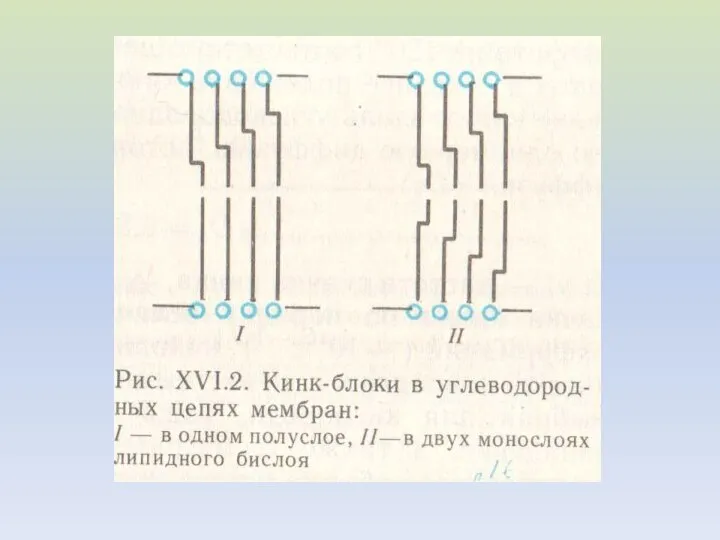
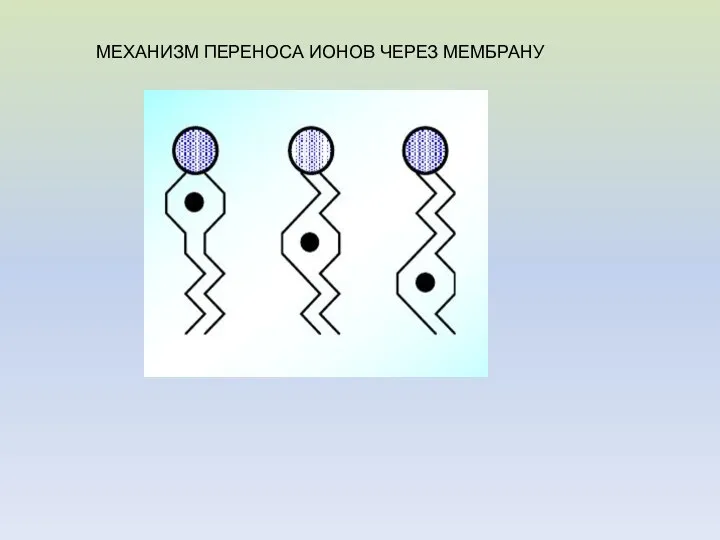
МЕХАНИЗМ ПЕРЕНОСА ИОНОВ ЧЕРЕЗ МЕМБРАНУ
МЕХАНИЗМ ПЕРЕНОСА ИОНОВ ЧЕРЕЗ МЕМБРАНУ
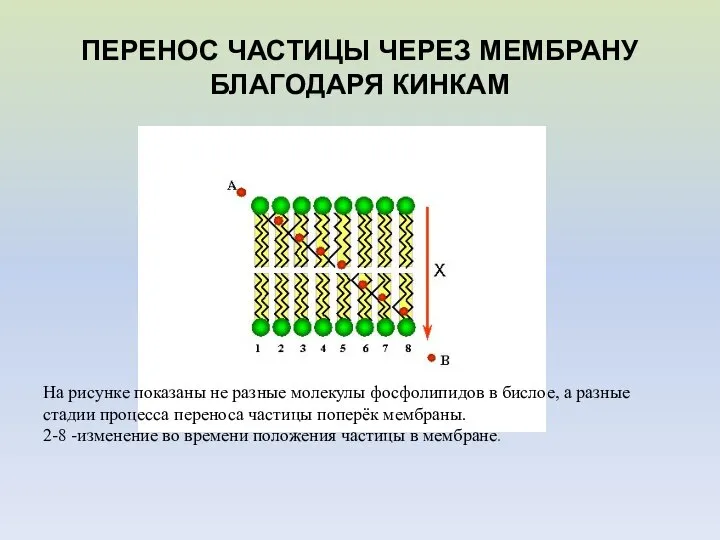
ПЕРЕНОС ЧАСТИЦЫ ЧЕРЕЗ МЕМБРАНУ БЛАГОДАРЯ КИНКАМ
На рисунке показаны не разные молекулы
ПЕРЕНОС ЧАСТИЦЫ ЧЕРЕЗ МЕМБРАНУ БЛАГОДАРЯ КИНКАМ
На рисунке показаны не разные молекулы

ФАЗОВЫЕ ПЕРЕХОДЫ В ЛИПИДНОМ БИСЛОЕ
ФАЗОВЫЕ ПЕРЕХОДЫ В ЛИПИДНОМ БИСЛОЕ
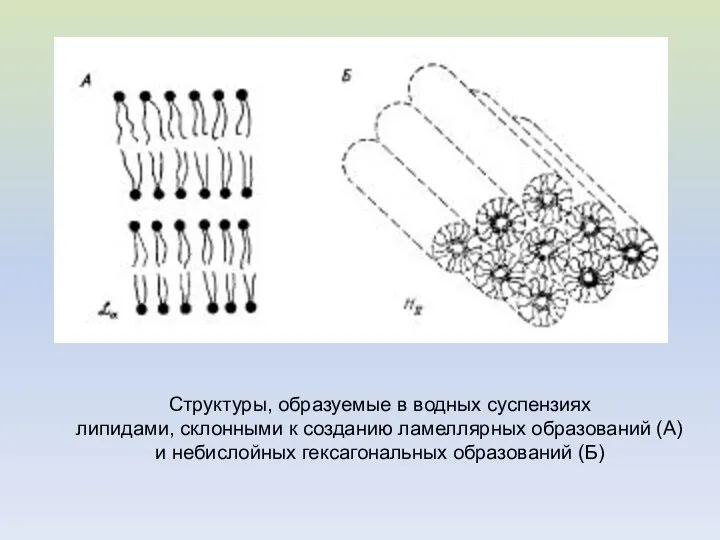
Структуры, образуемые в водных суспензиях
липидами, склонными к созданию ламеллярных образований
Структуры, образуемые в водных суспензиях
липидами, склонными к созданию ламеллярных образований
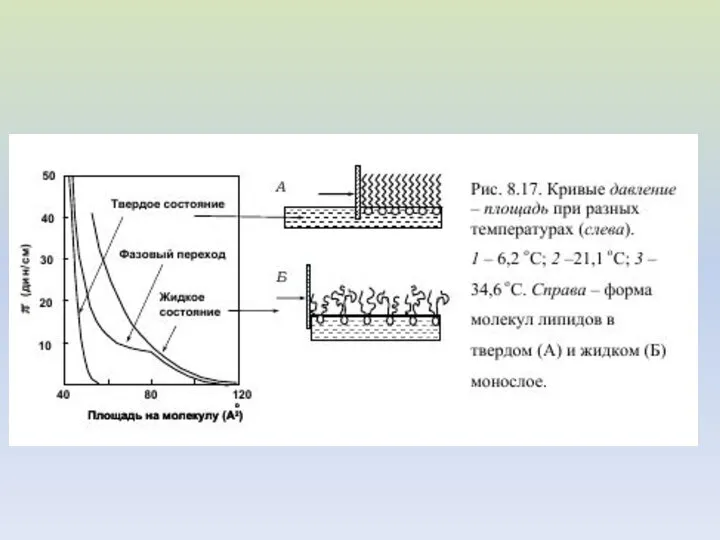
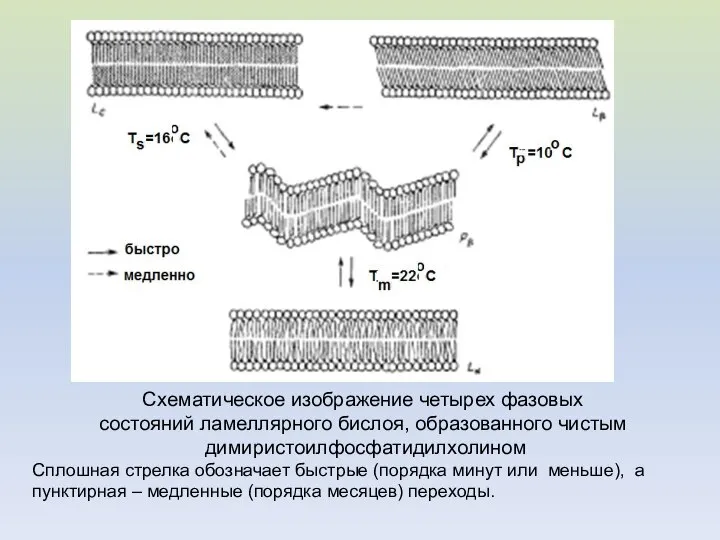
Схематическое изображение четырех фазовых
состояний ламеллярного бислоя, образованного чистым
димиристоилфосфатидилхолином
Схематическое изображение четырех фазовых
состояний ламеллярного бислоя, образованного чистым
димиристоилфосфатидилхолином
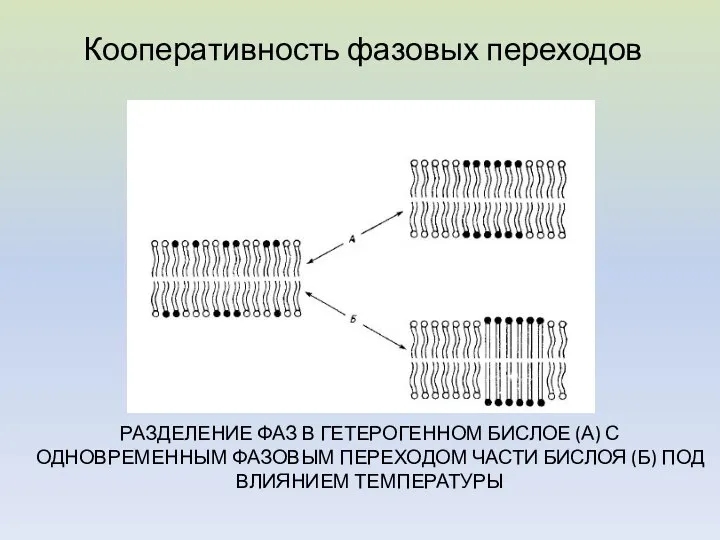
РАЗДЕЛЕНИЕ ФАЗ В ГЕТЕРОГЕННОМ БИСЛОЕ (А) С ОДНОВРЕМЕННЫМ ФАЗОВЫМ ПЕРЕХОДОМ ЧАСТИ
РАЗДЕЛЕНИЕ ФАЗ В ГЕТЕРОГЕННОМ БИСЛОЕ (А) С ОДНОВРЕМЕННЫМ ФАЗОВЫМ ПЕРЕХОДОМ ЧАСТИ

МЕТОД ДИФФЕРЕНЦИАЛЬНОЙ СКАНИРУЮЩЕЙ МИКРОКАЛОРИМЕТРИИ
Метод, основанный на измерении разницы тепловых потоков, идущих
МЕТОД ДИФФЕРЕНЦИАЛЬНОЙ СКАНИРУЮЩЕЙ МИКРОКАЛОРИМЕТРИИ
Метод, основанный на измерении разницы тепловых потоков, идущих
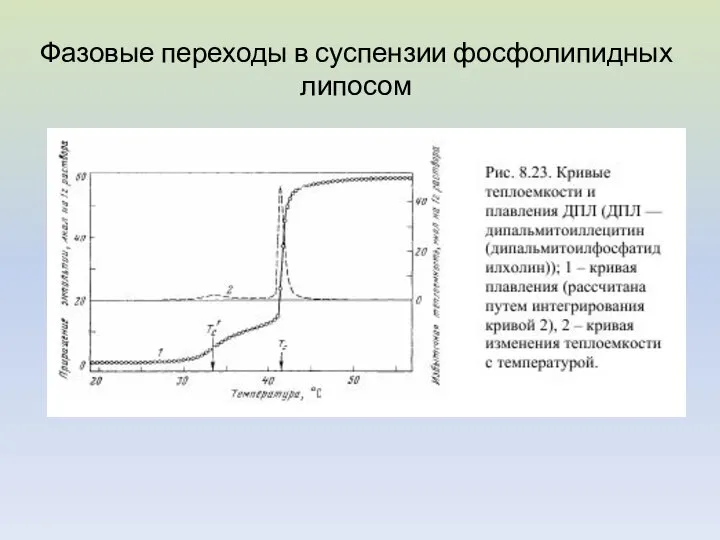
Фазовые переходы в суспензии фосфолипидных липосом
Фазовые переходы в суспензии фосфолипидных липосом
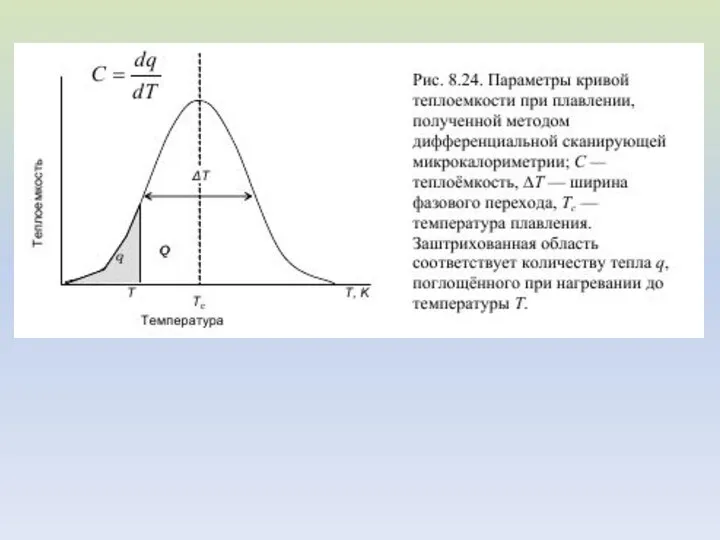

Анализ кривых ДСК
Пусть удельная теплота плавления липида равна Qm, количество
Анализ кривых ДСК
Пусть удельная теплота плавления липида равна Qm, количество

В интервале температур от T1 до текущей температуры T расплавится количество
В интервале температур от T1 до текущей температуры T расплавится количество
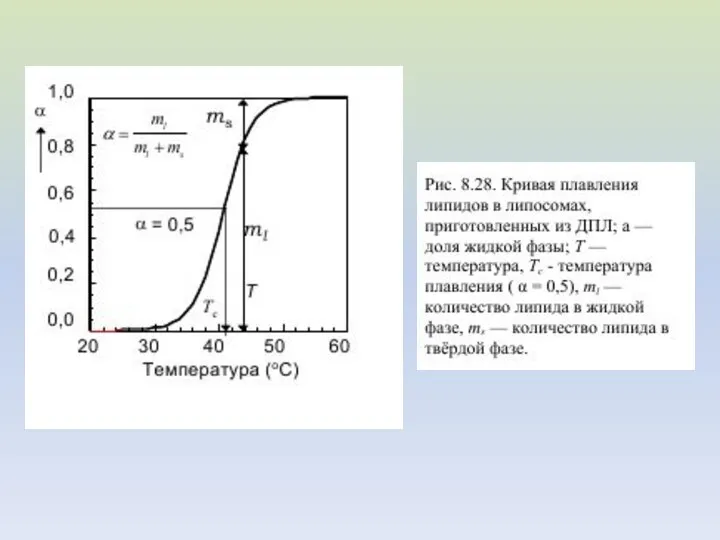
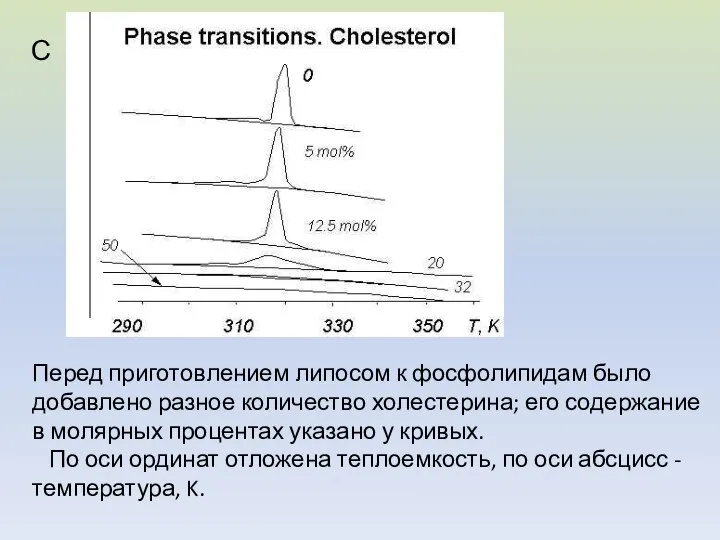
Перед приготовлением липосом к фосфолипидам было добавлено разное количество холестерина; его
Перед приготовлением липосом к фосфолипидам было добавлено разное количество холестерина; его
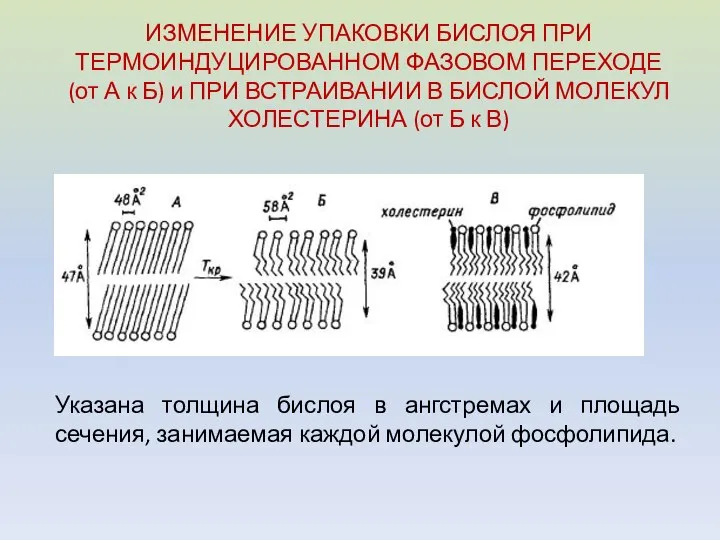
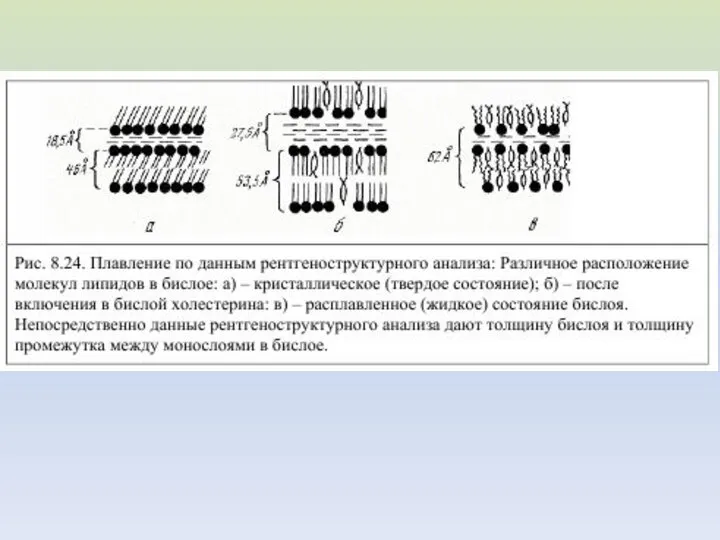
ИЗМЕНЕНИЕ УПАКОВКИ БИСЛОЯ ПРИ ТЕРМОИНДУЦИРОВАННОМ ФАЗОВОМ ПЕРЕХОДЕ (от А к Б)
ИЗМЕНЕНИЕ УПАКОВКИ БИСЛОЯ ПРИ ТЕРМОИНДУЦИРОВАННОМ ФАЗОВОМ ПЕРЕХОДЕ (от А к Б)
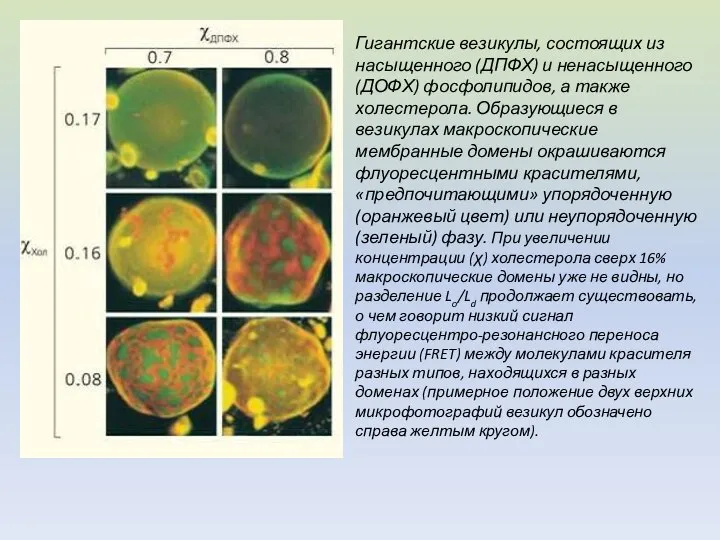
Гигантские везикулы, состоящих из насыщенного (ДПФХ) и ненасыщенного (ДОФХ) фосфолипидов, а
Гигантские везикулы, состоящих из насыщенного (ДПФХ) и ненасыщенного (ДОФХ) фосфолипидов, а
ФЛУОРЕСЦЕНТНЫЕ МЕТОДЫ ИССЛЕДОВАНИЯ
ФЛУОРЕСЦЕНТНЫЙ АНАЛИЗ ДАЕТ ВОЗМОЖНОСТЬ ИССЛЕДОВАТЬ ПОДВИЖНОСТЬ ФОСФОЛИПИДОВ В МЕМБРАНЕ,
ФЛУОРЕСЦЕНТНЫЕ МЕТОДЫ ИССЛЕДОВАНИЯ
ФЛУОРЕСЦЕНТНЫЙ АНАЛИЗ ДАЕТ ВОЗМОЖНОСТЬ ИССЛЕДОВАТЬ ПОДВИЖНОСТЬ ФОСФОЛИПИДОВ В МЕМБРАНЕ,
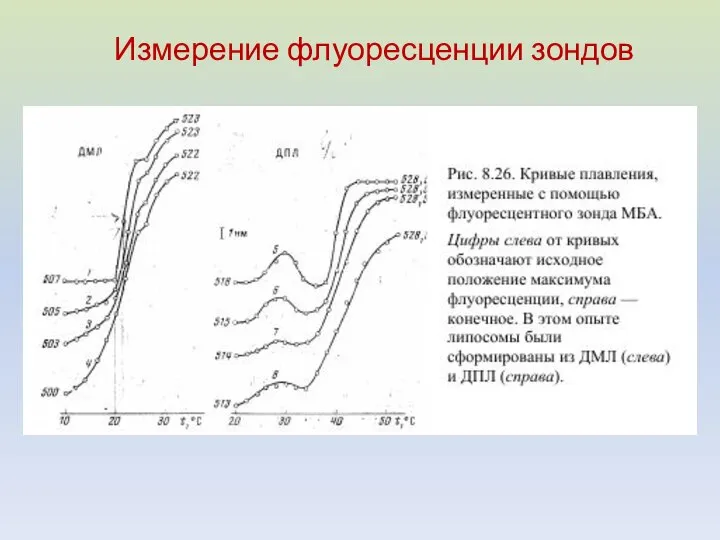
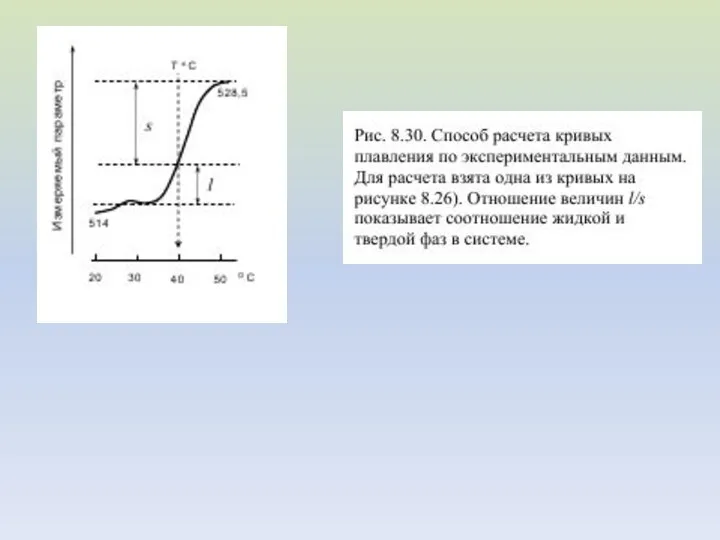
Измерение флуоресценции зондов
Измерение флуоресценции зондов
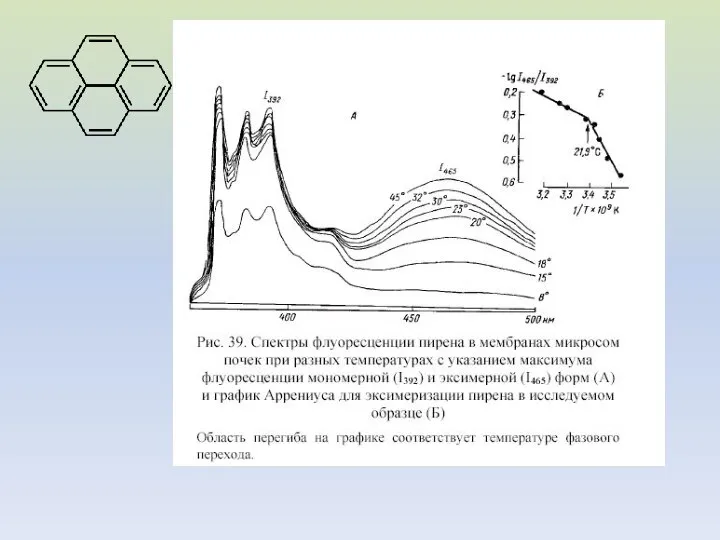
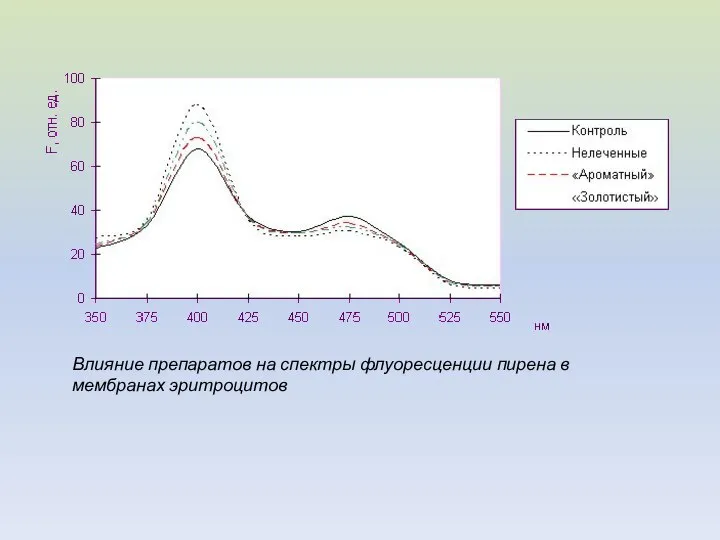
Влияние препаратов на спектры флуоресценции пирена в мембранах эритроцитов
Влияние препаратов на спектры флуоресценции пирена в мембранах эритроцитов
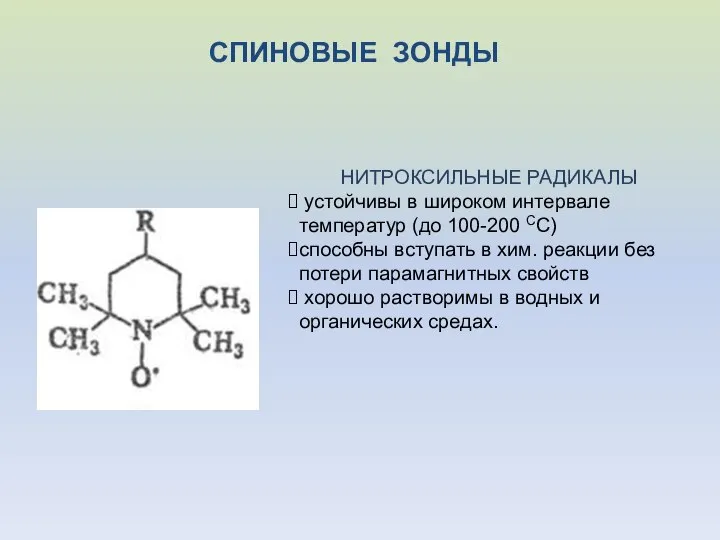
СПИНОВЫЕ ЗОНДЫ
НИТРОКСИЛЬНЫЕ РАДИКАЛЫ
устойчивы в широком интервале температур (до 100-200 СС)
СПИНОВЫЕ ЗОНДЫ
НИТРОКСИЛЬНЫЕ РАДИКАЛЫ
устойчивы в широком интервале температур (до 100-200 СС)
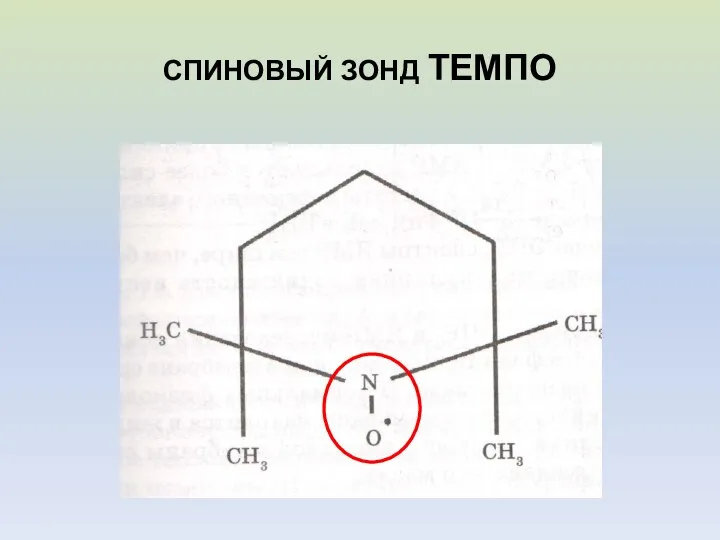
СПИНОВЫЙ ЗОНД ТЕМПО
СПИНОВЫЙ ЗОНД ТЕМПО
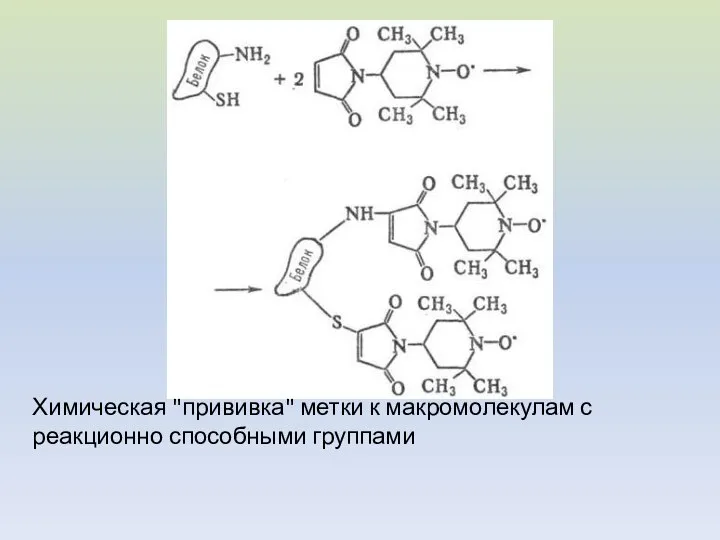
Химическая "прививка" метки к макромолекулам с реакционно способными группами
Химическая "прививка" метки к макромолекулам с реакционно способными группами

Реакции макромолекул с бирадикалами и спиновыми ловушками.
Спиновая ловушка - соединение, образующее
Реакции макромолекул с бирадикалами и спиновыми ловушками.
Спиновая ловушка - соединение, образующее
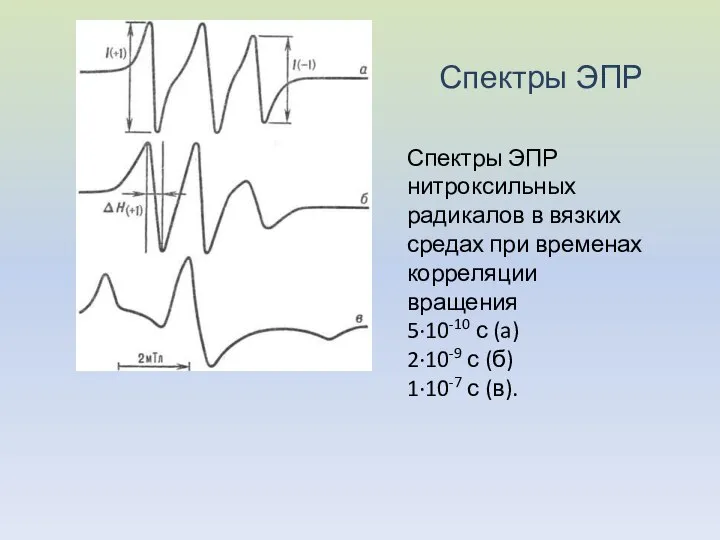
Спектры ЭПР
Спектры ЭПР нитроксильных радикалов в вязких средах при временах корреляции
Спектры ЭПР
Спектры ЭПР нитроксильных радикалов в вязких средах при временах корреляции
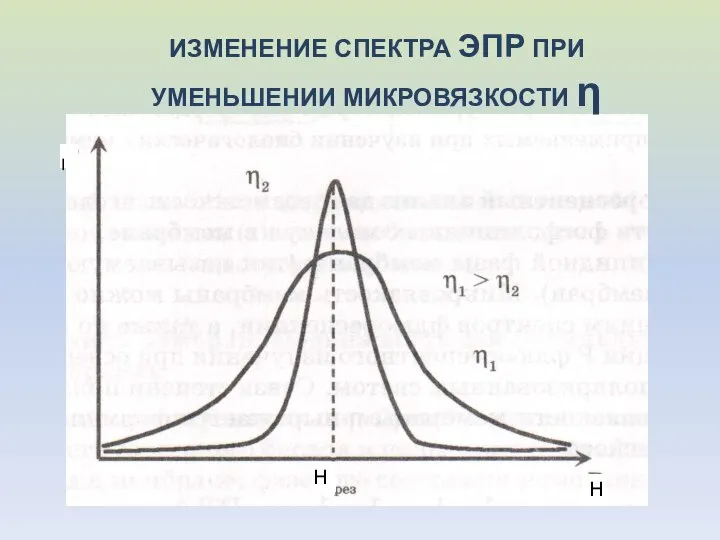
ИЗМЕНЕНИЕ СПЕКТРА ЭПР ПРИ УМЕНЬШЕНИИ МИКРОВЯЗКОСТИ η
Н
Н
I
ИЗМЕНЕНИЕ СПЕКТРА ЭПР ПРИ УМЕНЬШЕНИИ МИКРОВЯЗКОСТИ η
Н
Н
I
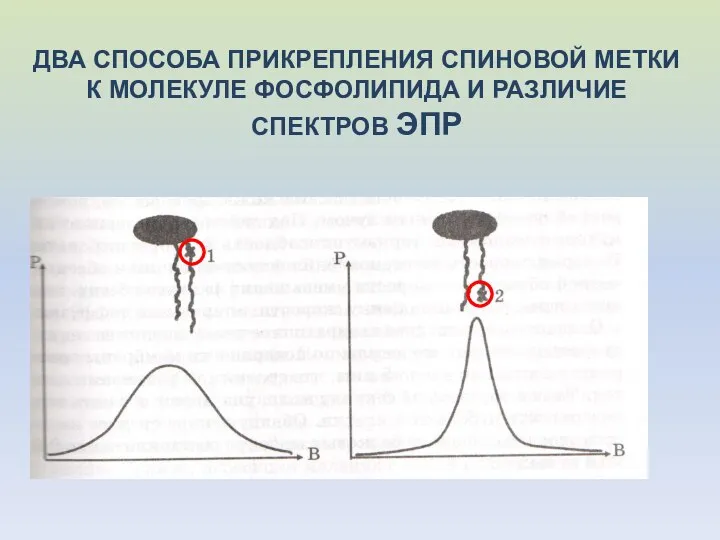
ДВА СПОСОБА ПРИКРЕПЛЕНИЯ СПИНОВОЙ МЕТКИ К МОЛЕКУЛЕ ФОСФОЛИПИДА И РАЗЛИЧИЕ СПЕКТРОВ
ДВА СПОСОБА ПРИКРЕПЛЕНИЯ СПИНОВОЙ МЕТКИ К МОЛЕКУЛЕ ФОСФОЛИПИДА И РАЗЛИЧИЕ СПЕКТРОВ
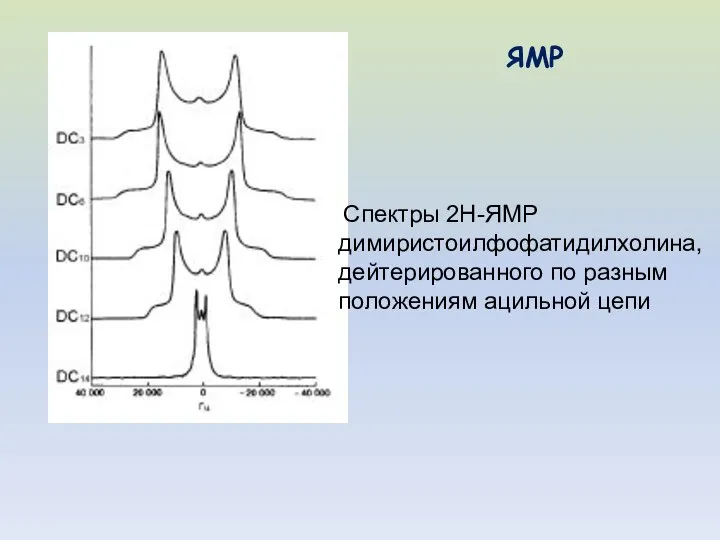
Спектры 2Н-ЯМР
димиристоилфофатидилхолина,
дейтерированного по разным
положениям ацильной цепи
ЯМР
Спектры 2Н-ЯМР
димиристоилфофатидилхолина,
дейтерированного по разным
положениям ацильной цепи
ЯМР
 Корпоративный менеджмент. Определения
Корпоративный менеджмент. Определения Резка ножницами с параллельными ножами с гидравлическим приводом
Резка ножницами с параллельными ножами с гидравлическим приводом Управление карьерой
Управление карьерой Аттестационая работа. Методическая разработка по проведению в начальной школе (3 класс) модуля «Путешествие по родному краю»
Аттестационая работа. Методическая разработка по проведению в начальной школе (3 класс) модуля «Путешествие по родному краю» Душа и природа в поэзии Ф.И. Тютчева
Душа и природа в поэзии Ф.И. Тютчева Права и свободы человека и гражданина
Права и свободы человека и гражданина Даже если поражение - нужно продолжать сражение. Детский футбольный клуб Пересвет
Даже если поражение - нужно продолжать сражение. Детский футбольный клуб Пересвет Қазіргі заман мәдениеті
Қазіргі заман мәдениеті Художественная культура Древнего и Средневекового Китая
Художественная культура Древнего и Средневекового Китая Продвижение бизнеса в социальных сетях
Продвижение бизнеса в социальных сетях Электроэнергетический комплекс РФ.
Электроэнергетический комплекс РФ.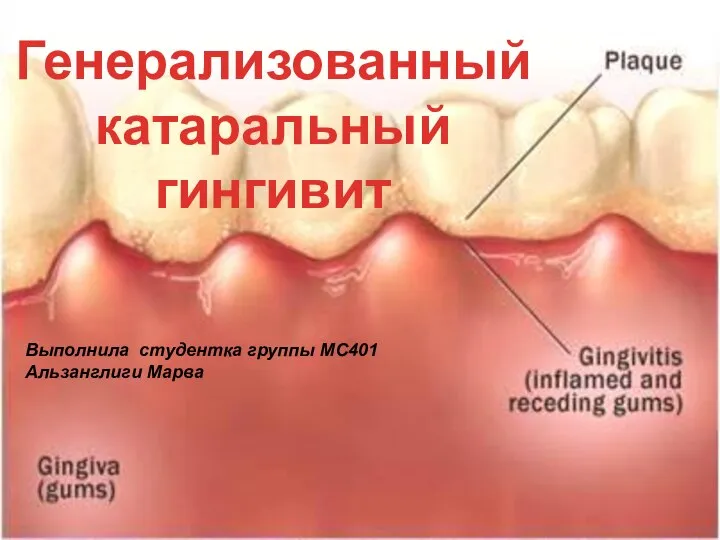 Презентация на тему "Генерализованный катаральный гингивит" - скачать презентации по Медицине
Презентация на тему "Генерализованный катаральный гингивит" - скачать презентации по Медицине РЕГУЛЯЦИЯ АКТИВНОСТИ ФЕРМЕНТОВ
РЕГУЛЯЦИЯ АКТИВНОСТИ ФЕРМЕНТОВ Нахождение характеристик рассеянной волны
Нахождение характеристик рассеянной волны Большекияикский ДЦ
Большекияикский ДЦ Диэлектрическая проницаемость электронного газа
Диэлектрическая проницаемость электронного газа Порядок осмотра, выявления неисправностей
Порядок осмотра, выявления неисправностей Женский западноевропейский костюм в раннем Средневековье
Женский западноевропейский костюм в раннем Средневековье Histoire de Disneyland Paris
Histoire de Disneyland Paris Пылающий огонь - презентация для начальной школы
Пылающий огонь - презентация для начальной школы Кадровое планирование в организации
Кадровое планирование в организации Презентация "представители фовизма" - скачать презентации по МХК
Презентация "представители фовизма" - скачать презентации по МХК Автомобильные дороги и городские улицы 5
Автомобильные дороги и городские улицы 5 Курс обучения тестированию
Курс обучения тестированию Авторитет судебных органов
Авторитет судебных органов Мезенская роспись
Мезенская роспись Биография и творчество А.Г.Алексина
Биография и творчество А.Г.Алексина Организация Объединённых Наций
Организация Объединённых Наций