- Углеводы и обмен углеводов

Содержание
- 2. Углеводы и обмен углеводов. Гликоген. Строение. Значение. Синтез гликогена. Ферменты. Мобилизация гликогена. Фосфоролиз. Ферменты. Связь гликогенолиза
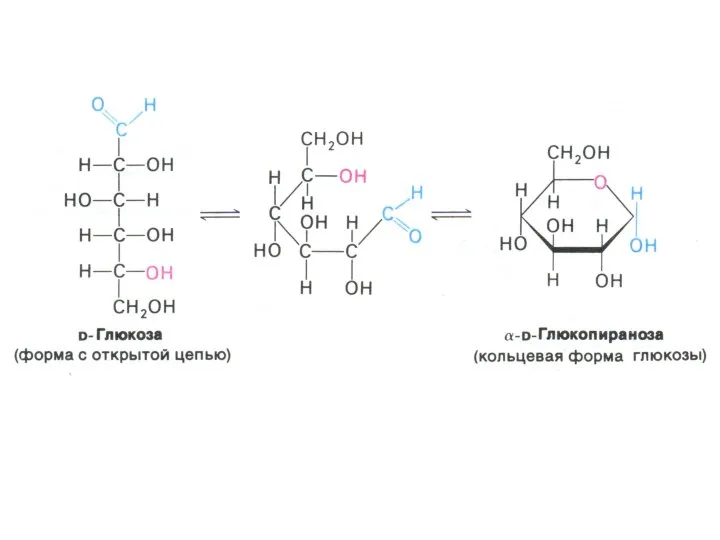
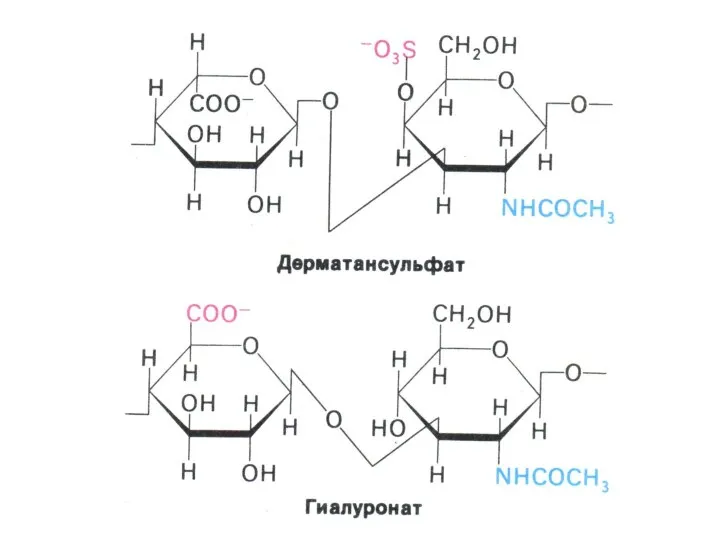
- 3. Углеводы и обмен углеводов Классификация углеводов (моно-, дисахариды, олигосахариды, полисахариды - нейтральные и кислые); Ацетилированные, аминированные,
- 9. Протеогликановый агрегат из эпифизарного хряща
- 10. Функции углеводов 1.Энергетическая (1г углеводов – 4,1 ккал) – глюкоза. Преимущество окисления углеводов в анаэробных условиях.
- 11. Функции углеводов Модификация белков – гликозилирование (норма и патология); Углеводы – как лекарственные препараты (глюкоза, гепарин,
- 12. Внешний обмен углеводов Эндоамилаза слюны и панкреатического сока: крахмал и гликоген ? декстрины, мальтотриозы, мальтоза. Гидролизуют
- 13. Промежуточный обмен углеводов Катаболизм: Гликолиз и пентозофосфатный шунт Окисление пирувата и ЦТК Гликогенолиз (расщепление гликогена) Анаболизм:
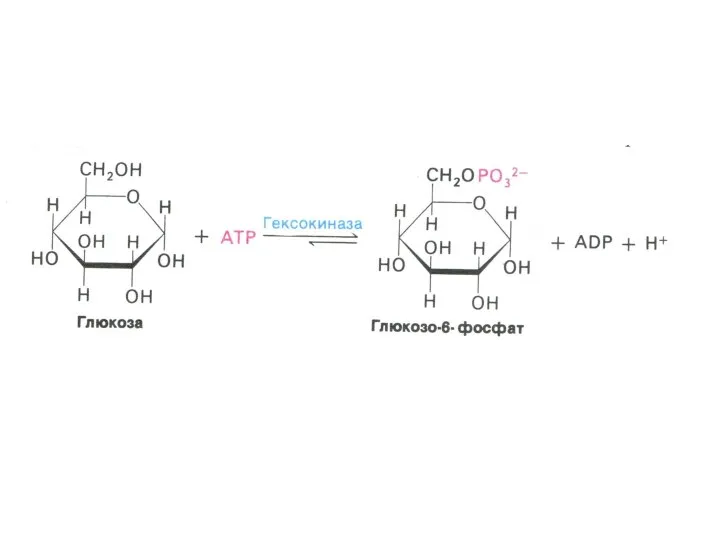
- 14. Особенность промежуточного обмена углеводов Сахара метаболизируют внутри клетки только в фосфорилированном виде. Фосфорилирование осуществляется гексокиназами, дефосфорилирование
- 15. Взаимопревращения сахаров При потреблении различных сахаров, в крови содержание глюкозы в сотни раз превышает таковое для
- 16. Расходование глюкозы клеткой Гликолиз и тканевое дыхание (65%) Запасание гликогена (5%) Синтез липидов при наличии избытка
- 17. ГЛИКОЛИЗ Центральный путь метаболизма глюкозы Самостоятельный способ получения энергии клеткой (в анаэробных условиях) и первый этап
- 30. Гликолиз, конечные продукты Гликолиз – окисление глюкозы, не требующее присутствия кислорода (брожение). В различных клетках и
- 32. Особенности гликолиза в разных тканях Наиболее распространенный шунт окисления глюкозы – пентозофосфатные превращения в печени, жировой
- 33. ПЕНТОЗОФОСФАТНЫЙ ПУТЬ окисления глюкозы Три ферментные системы, локализованные в цитоплазме: Окислительно-декарбоксилирующая (образование пентозы – рибулозо-5-фосфата и
- 41. Шунт Раппопорта в эритроцитах Особенностью гликолиза в эритроцитах является образование 2,3-глицерофосфата.(особенно усиливающегося в условиях гипоксии) 2,3-
- 43. Аэробный метаболизм глюкозы Окисление глюкозы продолжается в матриксе митохондрий, куда пируват поступает с помощь переносчика (пируват-
- 50. Окисление пирувата Необратимая реакция. Продукт окисления и декарбоксилирования – ацетил-КоА. Протон фиксируется в НАДН. 2-х углеродный
- 51. Структура и функции КоА Низкомолекулярный термостабильный (не связан с белком) фактор – активатор и переносчик ацетильных
- 52. Ацетил-КоА Ацетил - КоА – универсальное клеточное топливо, окисляется до СО2 – в цикле лимонной кислоты
- 53. Цикл трикарбоновых кислот (лимонной кислоты), цикл Кребса. 8 реакций, замкнутых в цикл («катализатором» является оксалоацетат), в
- 58. α –кетоглутаратдегидрогеназный комплекс Мультиферментный комплекс, осуществляющий декарбоксилирование и окисление кетоглутарата, аналогично пируватдегидрогеназному комплексу. Содержит 3 фермента
- 59. Субстратное фосфорилирование Сукцинил-КоА содержит энергию тиоэфирной связи, достаточную для обеспечения реакции фосфорилирования АДФ (ГДФ) до АТФ
- 62. Функции цикла Кребса Водороддонорная (генерация 4-х пар протонов, поступающих в цепь переноса электронов в составе 3
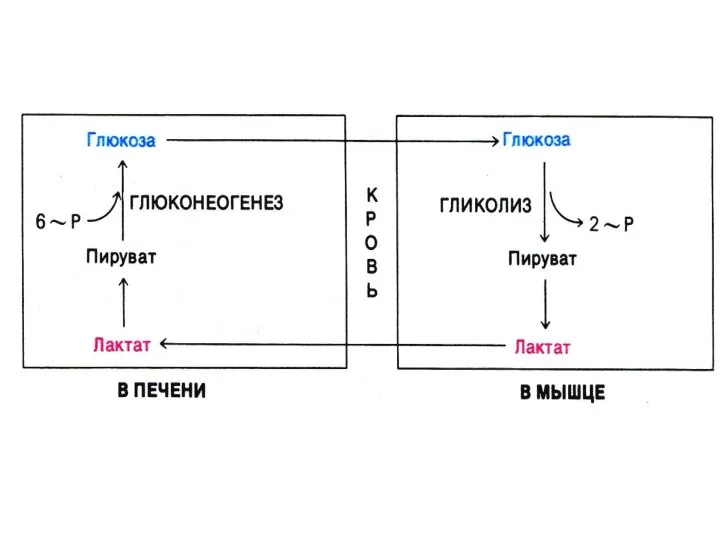
- 63. Глюконеогенез Образование глюкозы из неуглеводных предшественников (лактата, глицерина, аминокислот, метаболитов ЦТК). 4 адаптивных фермента, «обращающих» лимитирующие
- 68. Перемещение оксалоацетата из митохондрий в цитозоль Пируват, проникнув в митохондрии, карбоксилируется до оксалоацетата (с участием биотина).
- 69. Ферменты глюконеогенеза Активны в печени, мозге, почках. Нет в скелетных мышцах. ФЭП- карбоксикиназа; фруктозо-1,6- дифосфатаза; глюкозо-6
- 72. Метаболизм гликогена Запасы гликогена в клетках разных тканей. Значение. Структура гликогена. Синтез гликогена. Условия. Локализация. Затравка.
- 76. Фосфоролиз гликогена В отличие от внешнего обмена (ЖКТ) , в клетке гликоген распадается путем фосфоролиза с
- 80. Синтез гликогена Субстратом для синтеза гликогена является активированная форма глюкозы УДФ -1 – глюкоза. Гл ?
- 84. Синтез гликогена Синтез гликогена осуществляется ферментами: ГЛИКОГЕНСИНТАЗОЙ и амило -1,4 - 1,6 –гликозилтрансферазой Гликогенсинтаза образует 1,4
- 85. Регуляция метаболизма гликогена Синтез и распад гликогена регулируются по одним и тем же механизмам (фосфорилирование-дефосфорилирование ключевых
- 86. Регуляция метаболизма гликогена Гликогенсинтаза ингибируется фосфорилированием при участии фосфопротеинкиназы и активируется дефосфорилированием при участии протеинфосфатазы. В
- 87. Регуляция метаболизма гликогена В мышцах: В состоянии покоя гликогенфосфорилаза аллостерически активируется АМФ; При слабых и умеренных
- 91. Скачать презентацию

Углеводы и обмен углеводов.
Гликоген. Строение. Значение.
Синтез гликогена. Ферменты.
Мобилизация гликогена.
Углеводы и обмен углеводов.
Гликоген. Строение. Значение.
Синтез гликогена. Ферменты.
Мобилизация гликогена.

Углеводы и обмен углеводов
Классификация углеводов (моно-, дисахариды, олигосахариды, полисахариды - нейтральные
Углеводы и обмен углеводов
Классификация углеводов (моно-, дисахариды, олигосахариды, полисахариды - нейтральные



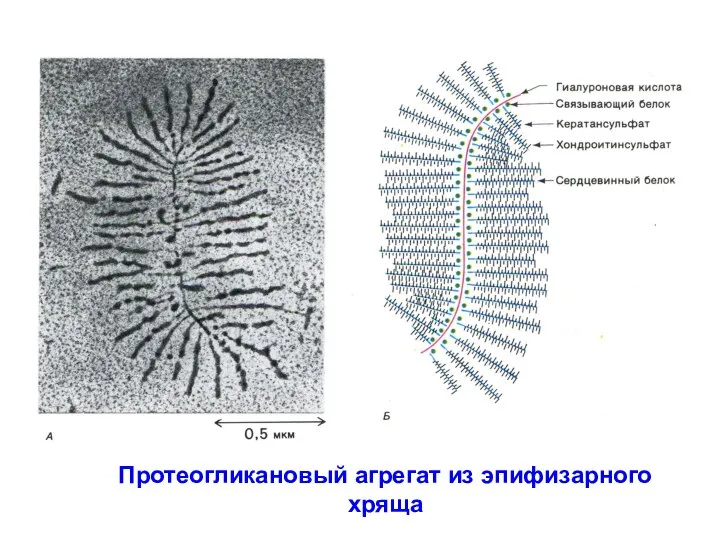
Протеогликановый агрегат из эпифизарного хряща
Протеогликановый агрегат из эпифизарного хряща

Функции углеводов
1.Энергетическая (1г углеводов – 4,1 ккал) – глюкоза.
Преимущество
Функции углеводов
1.Энергетическая (1г углеводов – 4,1 ккал) – глюкоза.
Преимущество

Функции углеводов
Модификация белков – гликозилирование (норма и патология);
Углеводы – как лекарственные
Функции углеводов
Модификация белков – гликозилирование (норма и патология);
Углеводы – как лекарственные

Внешний обмен углеводов
Эндоамилаза слюны и панкреатического сока: крахмал и гликоген ?
Внешний обмен углеводов
Эндоамилаза слюны и панкреатического сока: крахмал и гликоген ?

Промежуточный обмен углеводов
Катаболизм:
Гликолиз и пентозофосфатный шунт
Окисление пирувата и ЦТК
Гликогенолиз (расщепление
Промежуточный обмен углеводов
Катаболизм:
Гликолиз и пентозофосфатный шунт
Окисление пирувата и ЦТК
Гликогенолиз (расщепление

Особенность промежуточного обмена углеводов
Сахара метаболизируют внутри клетки только в фосфорилированном виде.
Фосфорилирование
Особенность промежуточного обмена углеводов
Сахара метаболизируют внутри клетки только в фосфорилированном виде.
Фосфорилирование

Взаимопревращения сахаров
При потреблении различных сахаров, в крови содержание глюкозы в сотни
Взаимопревращения сахаров
При потреблении различных сахаров, в крови содержание глюкозы в сотни

Расходование глюкозы клеткой
Гликолиз и тканевое дыхание (65%)
Запасание гликогена (5%)
Синтез липидов при
Расходование глюкозы клеткой
Гликолиз и тканевое дыхание (65%)
Запасание гликогена (5%)
Синтез липидов при

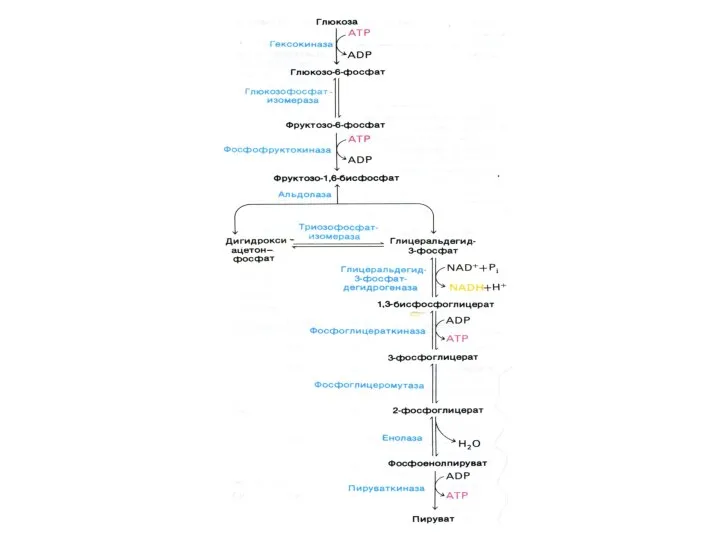
ГЛИКОЛИЗ
Центральный путь метаболизма глюкозы
Самостоятельный способ получения энергии клеткой (в анаэробных условиях)
ГЛИКОЛИЗ
Центральный путь метаболизма глюкозы
Самостоятельный способ получения энергии клеткой (в анаэробных условиях)
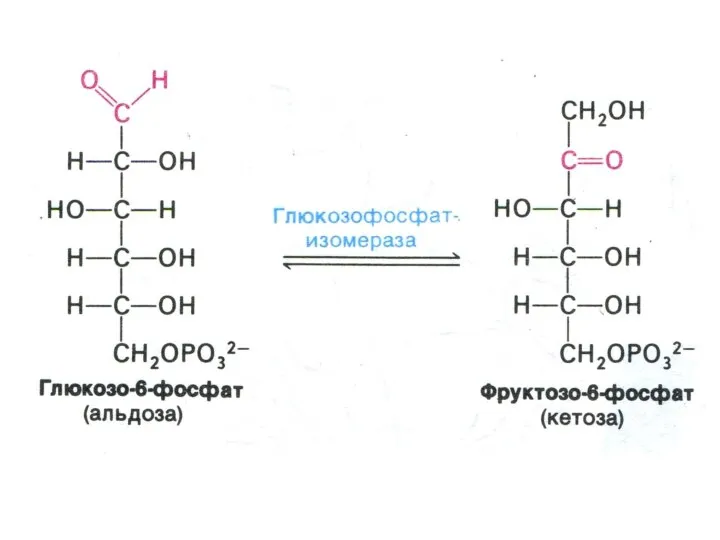



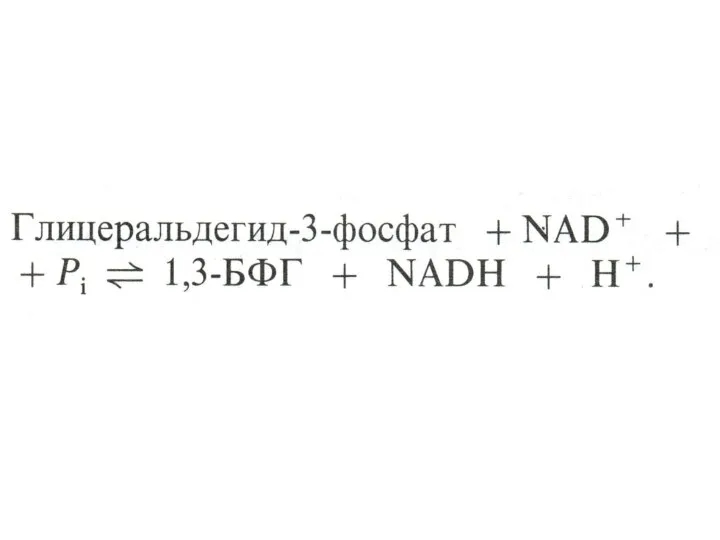

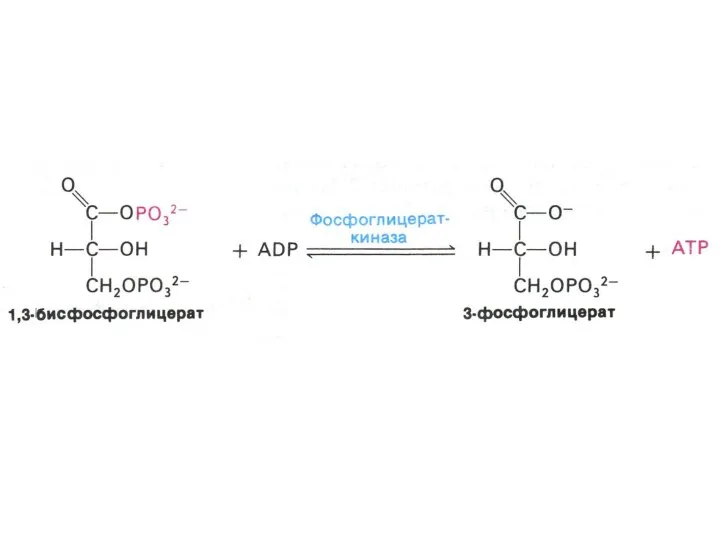

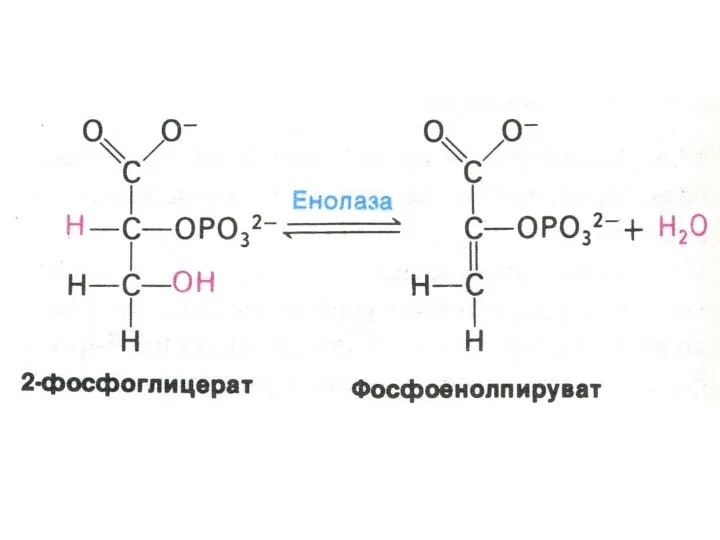
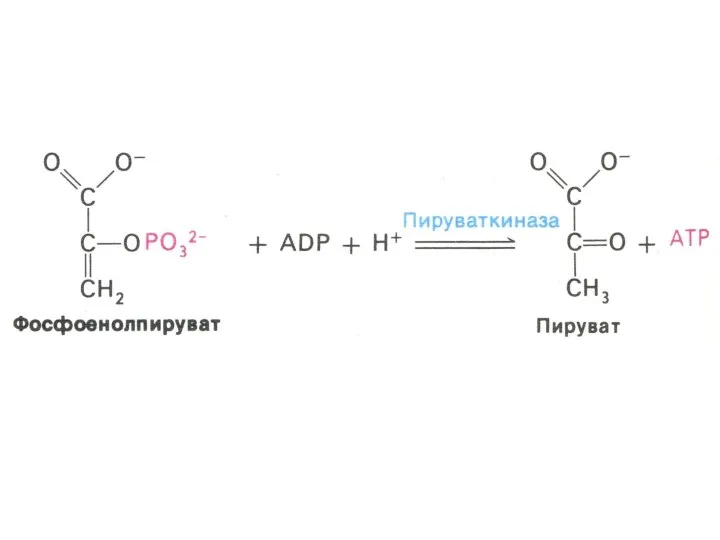

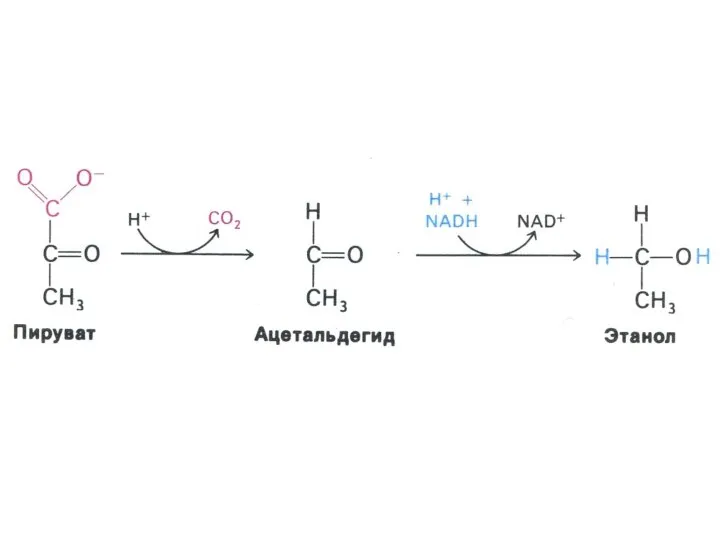
Гликолиз, конечные продукты
Гликолиз – окисление глюкозы, не требующее присутствия кислорода (брожение).
Гликолиз, конечные продукты
Гликолиз – окисление глюкозы, не требующее присутствия кислорода (брожение).

Особенности гликолиза в разных тканях
Наиболее распространенный шунт окисления глюкозы – пентозофосфатные
Особенности гликолиза в разных тканях
Наиболее распространенный шунт окисления глюкозы – пентозофосфатные

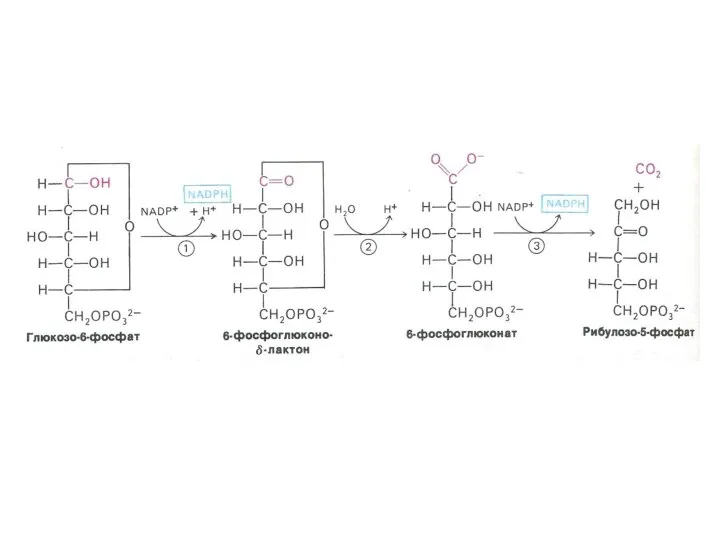
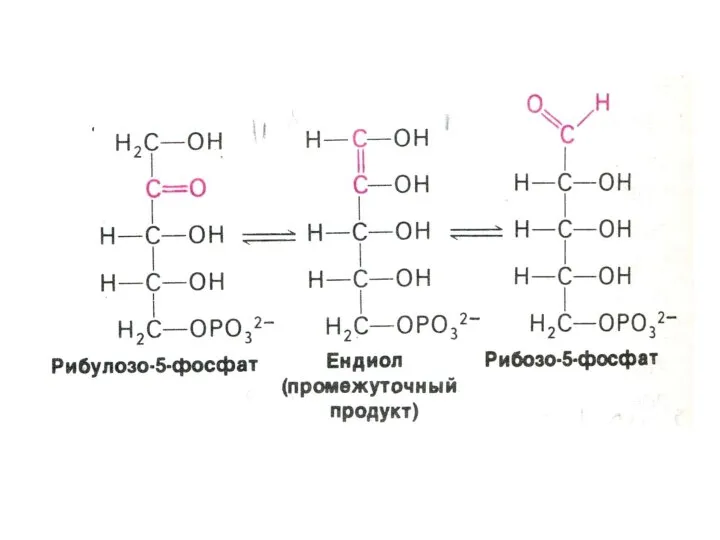
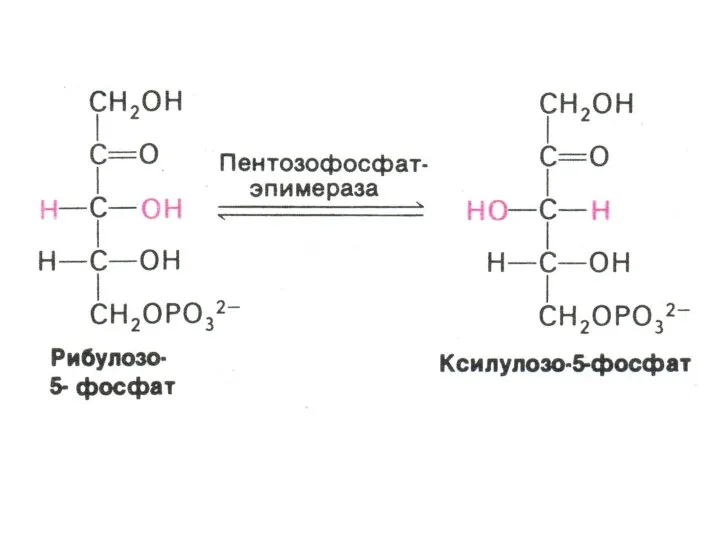
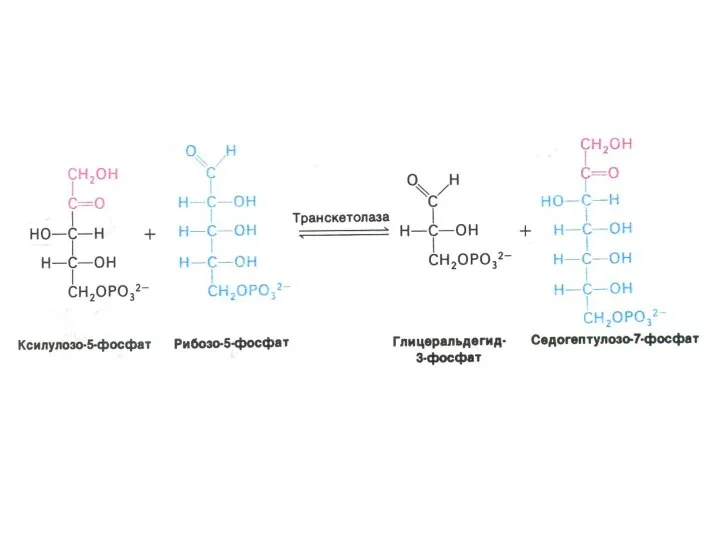
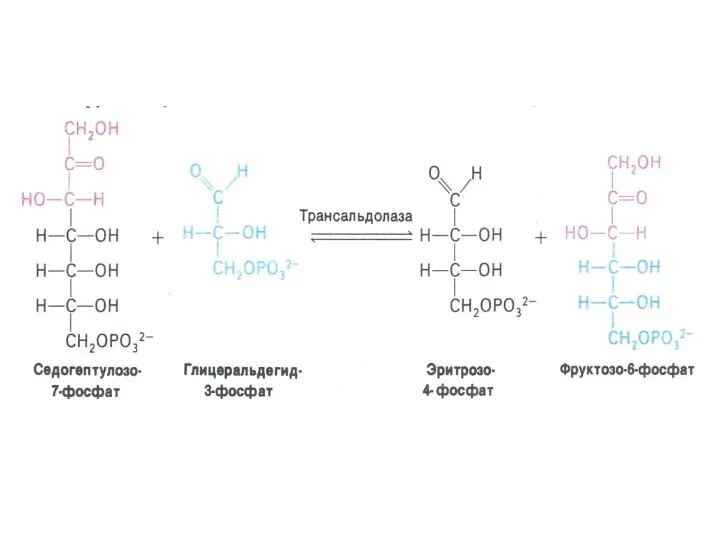
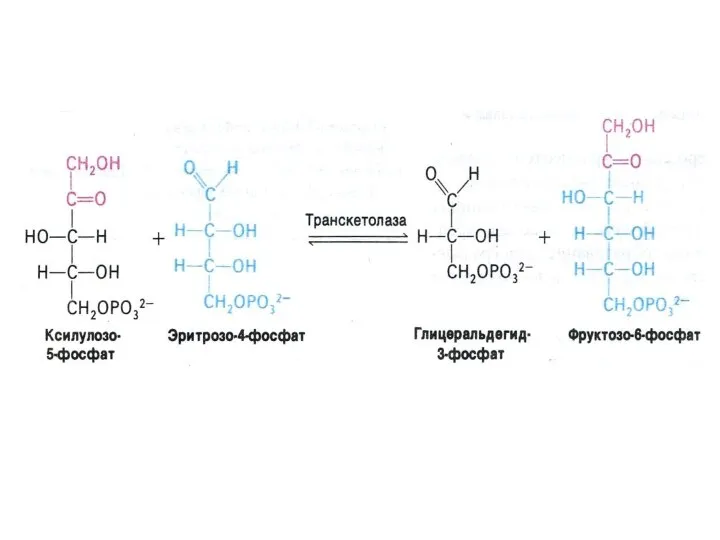
ПЕНТОЗОФОСФАТНЫЙ ПУТЬ окисления глюкозы
Три ферментные системы, локализованные в цитоплазме:
Окислительно-декарбоксилирующая (образование пентозы
ПЕНТОЗОФОСФАТНЫЙ ПУТЬ окисления глюкозы
Три ферментные системы, локализованные в цитоплазме:
Окислительно-декарбоксилирующая (образование пентозы


Шунт Раппопорта в эритроцитах
Особенностью гликолиза в эритроцитах является образование 2,3-глицерофосфата.(особенно усиливающегося
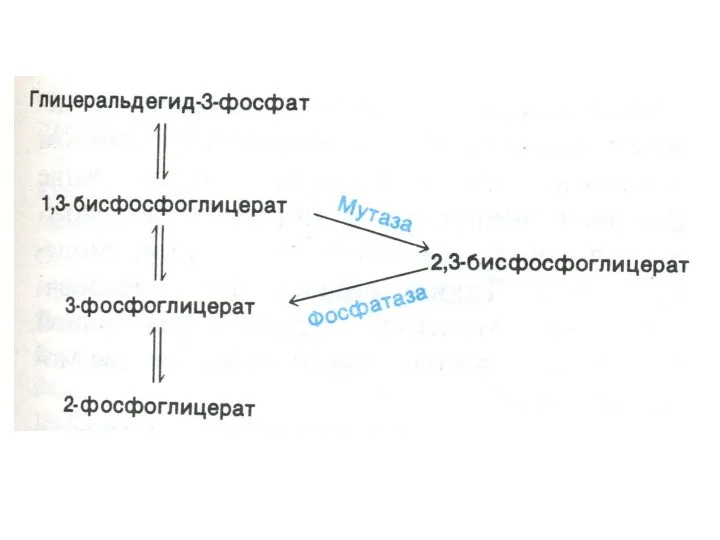
Шунт Раппопорта в эритроцитах
Особенностью гликолиза в эритроцитах является образование 2,3-глицерофосфата.(особенно усиливающегося

Аэробный метаболизм глюкозы
Окисление глюкозы продолжается в матриксе митохондрий, куда пируват поступает
Аэробный метаболизм глюкозы
Окисление глюкозы продолжается в матриксе митохондрий, куда пируват поступает


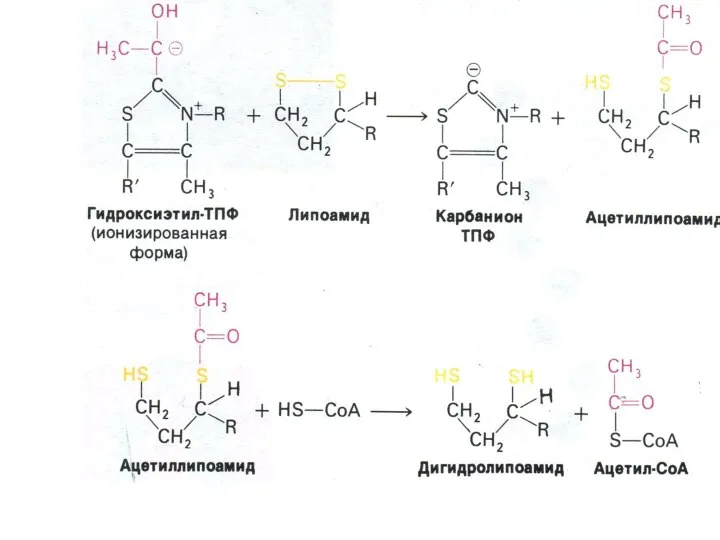
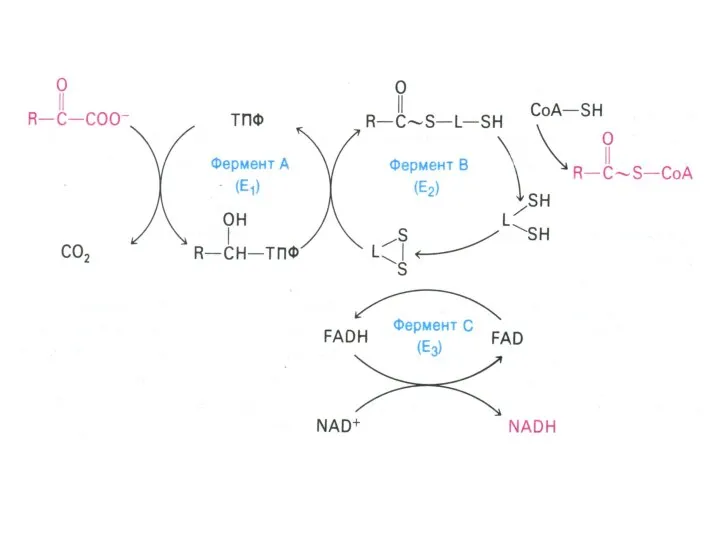

Окисление пирувата
Необратимая реакция.
Продукт окисления и декарбоксилирования – ацетил-КоА.
Протон фиксируется в НАДН.
2-х
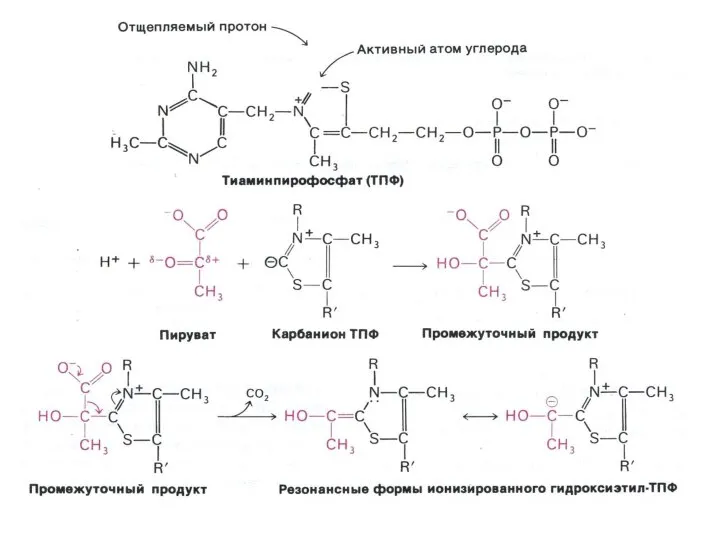
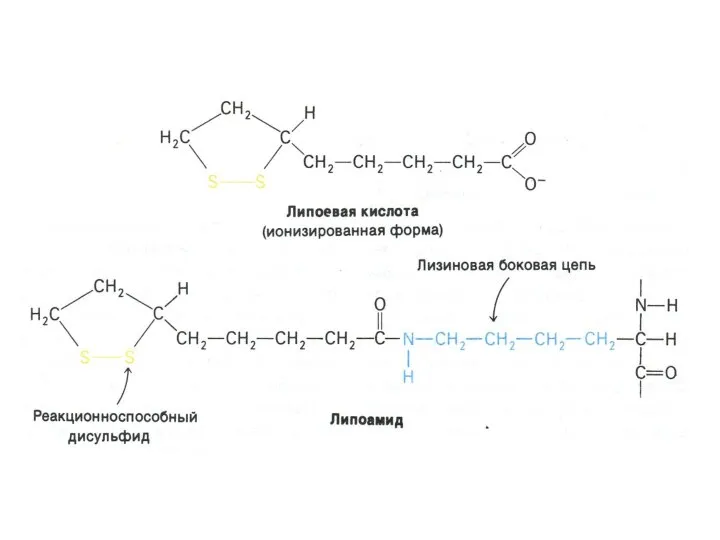
Окисление пирувата
Необратимая реакция.
Продукт окисления и декарбоксилирования – ацетил-КоА.
Протон фиксируется в НАДН.
2-х

Структура и функции КоА
Низкомолекулярный термостабильный (не связан с белком) фактор –
Структура и функции КоА
Низкомолекулярный термостабильный (не связан с белком) фактор –

Ацетил-КоА
Ацетил - КоА – универсальное клеточное топливо, окисляется до СО2 –
Ацетил-КоА
Ацетил - КоА – универсальное клеточное топливо, окисляется до СО2 –

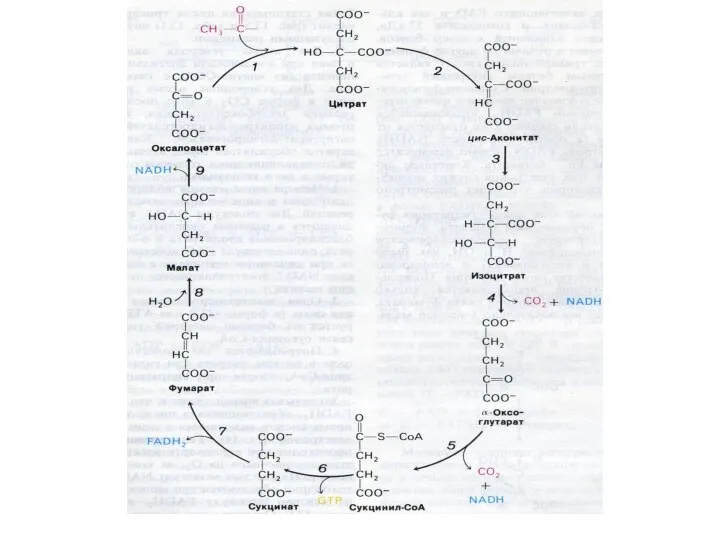
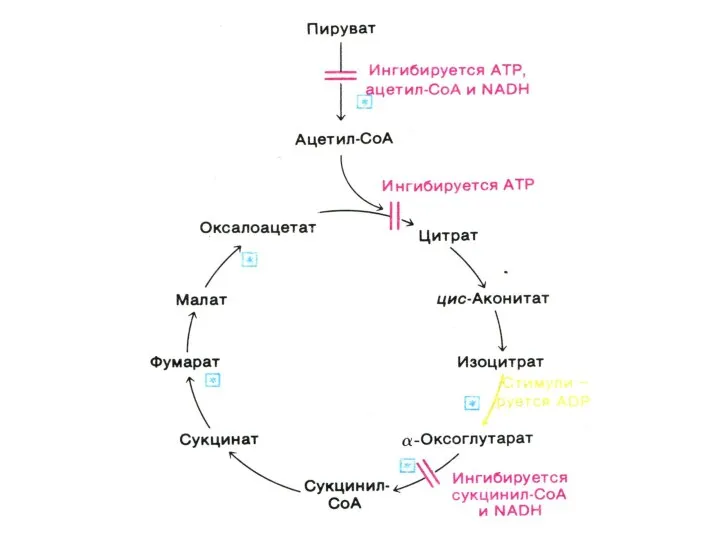
Цикл трикарбоновых кислот (лимонной кислоты), цикл Кребса.
8 реакций, замкнутых в цикл
Цикл трикарбоновых кислот (лимонной кислоты), цикл Кребса.
8 реакций, замкнутых в цикл


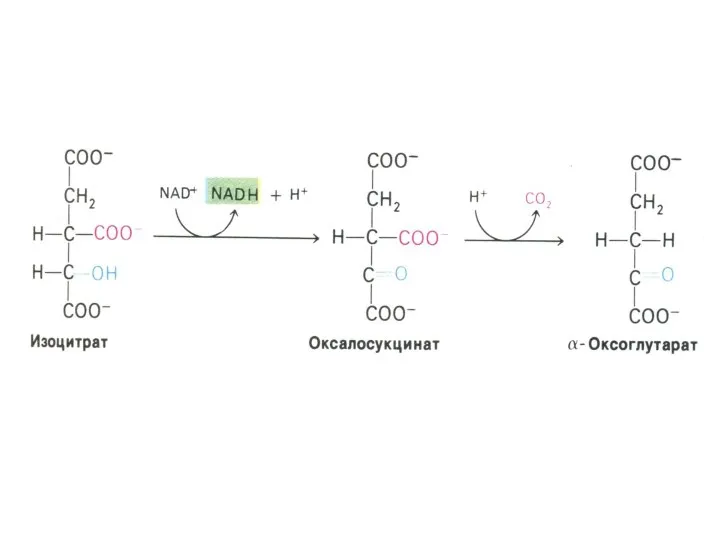

α –кетоглутаратдегидрогеназный комплекс
Мультиферментный комплекс, осуществляющий декарбоксилирование и окисление кетоглутарата, аналогично пируватдегидрогеназному
α –кетоглутаратдегидрогеназный комплекс
Мультиферментный комплекс, осуществляющий декарбоксилирование и окисление кетоглутарата, аналогично пируватдегидрогеназному

Субстратное фосфорилирование
Сукцинил-КоА содержит энергию тиоэфирной связи, достаточную для обеспечения реакции фосфорилирования
Субстратное фосфорилирование
Сукцинил-КоА содержит энергию тиоэфирной связи, достаточную для обеспечения реакции фосфорилирования
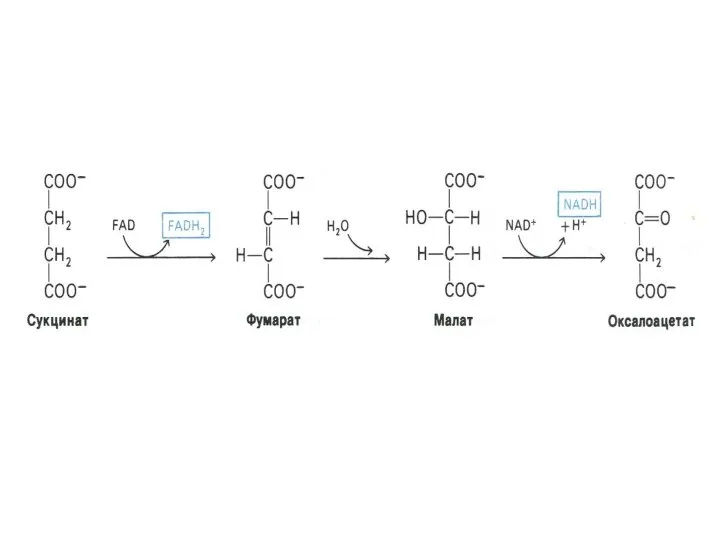

Функции цикла Кребса
Водороддонорная (генерация 4-х пар протонов, поступающих в цепь переноса
Функции цикла Кребса
Водороддонорная (генерация 4-х пар протонов, поступающих в цепь переноса

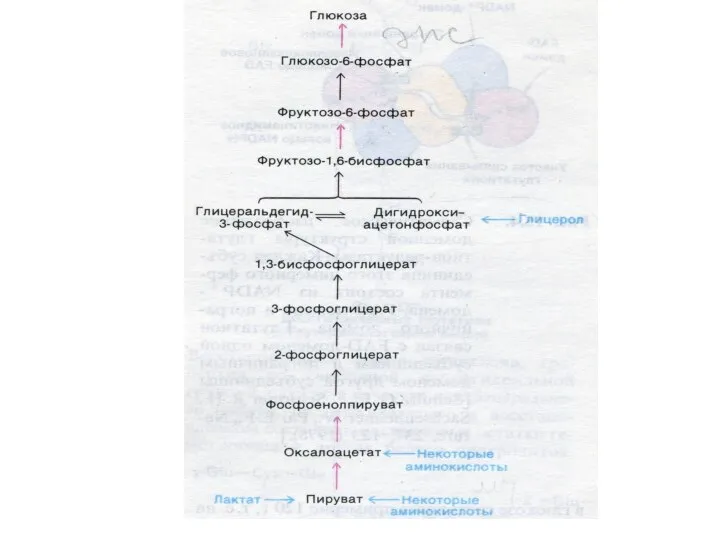
Глюконеогенез
Образование глюкозы из неуглеводных предшественников (лактата, глицерина, аминокислот, метаболитов ЦТК).
4 адаптивных
Глюконеогенез
Образование глюкозы из неуглеводных предшественников (лактата, глицерина, аминокислот, метаболитов ЦТК).
4 адаптивных
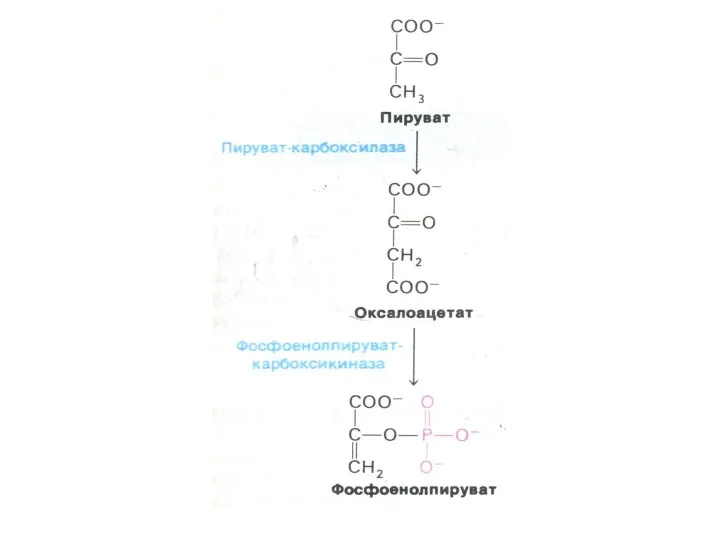
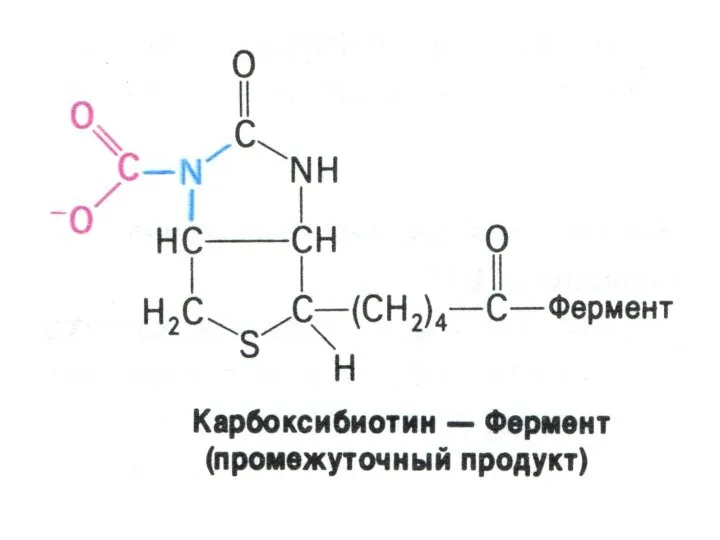


Перемещение оксалоацетата из митохондрий в цитозоль
Пируват, проникнув в митохондрии, карбоксилируется до
Перемещение оксалоацетата из митохондрий в цитозоль
Пируват, проникнув в митохондрии, карбоксилируется до

Ферменты глюконеогенеза
Активны в печени, мозге, почках. Нет в скелетных мышцах.
ФЭП- карбоксикиназа;
Ферменты глюконеогенеза
Активны в печени, мозге, почках. Нет в скелетных мышцах.
ФЭП- карбоксикиназа;


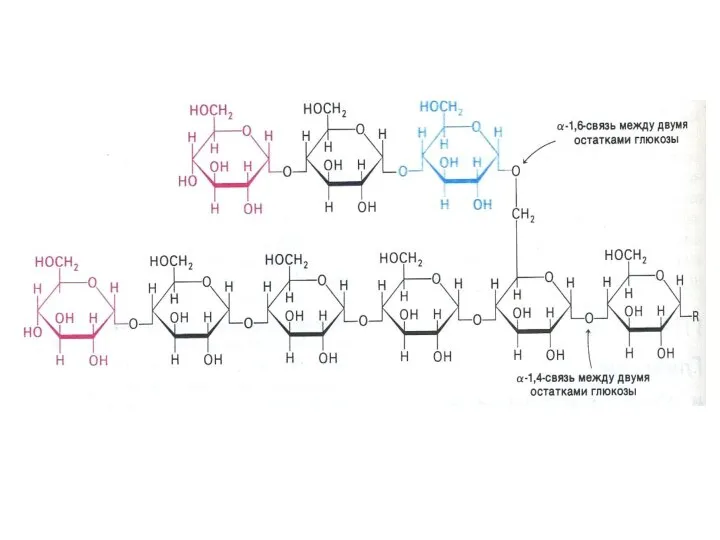
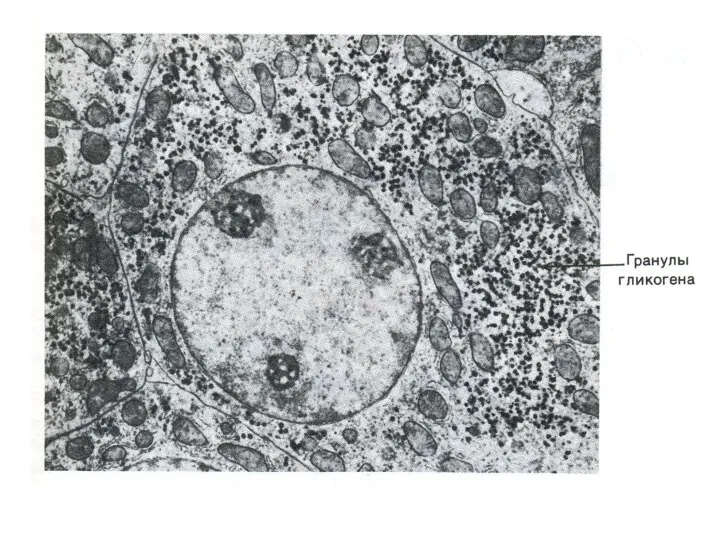
Метаболизм гликогена
Запасы гликогена в клетках разных тканей. Значение. Структура гликогена.
Синтез гликогена.
Метаболизм гликогена
Запасы гликогена в клетках разных тканей. Значение. Структура гликогена.
Синтез гликогена.


Фосфоролиз гликогена
В отличие от внешнего обмена (ЖКТ) , в клетке гликоген
Фосфоролиз гликогена
В отличие от внешнего обмена (ЖКТ) , в клетке гликоген
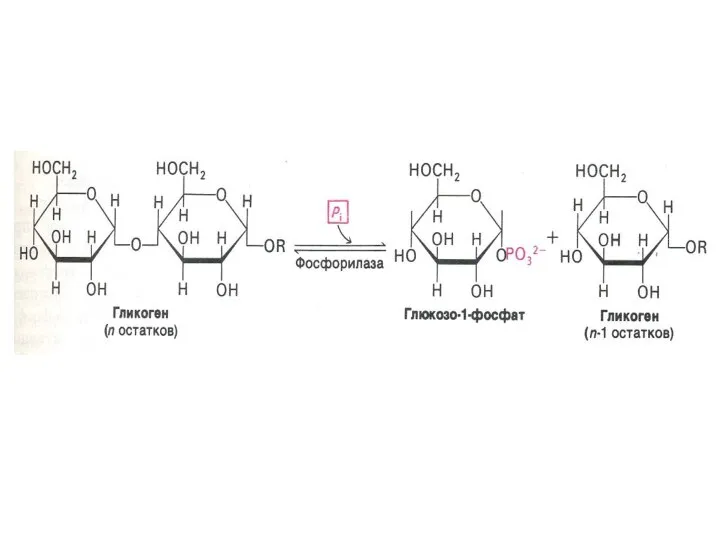



Синтез гликогена
Субстратом для синтеза гликогена является активированная форма глюкозы УДФ -1
Синтез гликогена
Субстратом для синтеза гликогена является активированная форма глюкозы УДФ -1

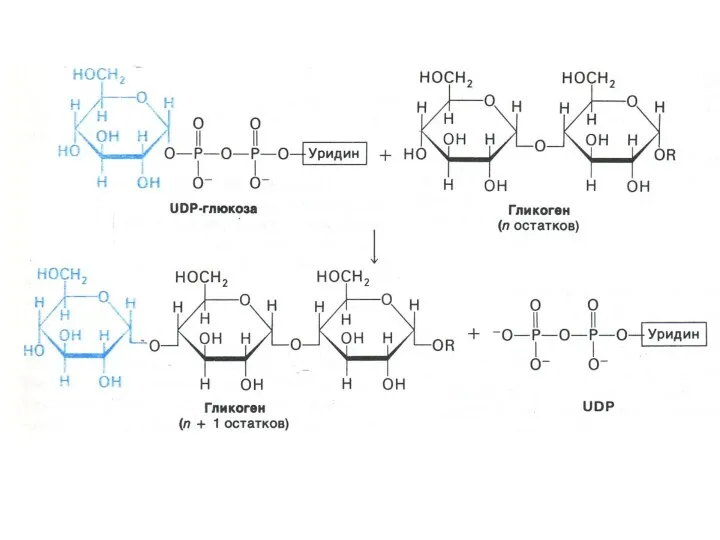


Синтез гликогена
Синтез гликогена осуществляется ферментами: ГЛИКОГЕНСИНТАЗОЙ и амило -1,4 - 1,6
Синтез гликогена
Синтез гликогена осуществляется ферментами: ГЛИКОГЕНСИНТАЗОЙ и амило -1,4 - 1,6

Регуляция метаболизма гликогена
Синтез и распад гликогена регулируются по одним и тем
Регуляция метаболизма гликогена
Синтез и распад гликогена регулируются по одним и тем

Регуляция метаболизма гликогена
Гликогенсинтаза ингибируется фосфорилированием при участии фосфопротеинкиназы и активируется
Регуляция метаболизма гликогена
Гликогенсинтаза ингибируется фосфорилированием при участии фосфопротеинкиназы и активируется

Регуляция метаболизма гликогена
В мышцах:
В состоянии покоя гликогенфосфорилаза аллостерически активируется АМФ;
При слабых
Регуляция метаболизма гликогена
В мышцах:
В состоянии покоя гликогенфосфорилаза аллостерически активируется АМФ;
При слабых

 Кровеносная система. Юля и Настя
Кровеносная система. Юля и Настя My idol Viktor Axelsen
My idol Viktor Axelsen Спасо-Преображенская церковь Евфросиниевского монастыря. Полоцк. XII в. Мастер Иоанн
Спасо-Преображенская церковь Евфросиниевского монастыря. Полоцк. XII в. Мастер Иоанн Технология и схемотехника средств управления в технических системах
Технология и схемотехника средств управления в технических системах Уравнение Лагранжа второго рода
Уравнение Лагранжа второго рода Отчего зависит продуктивность инженеров? Внешние факторы. //А. Орлов 2009. - презентация
Отчего зависит продуктивность инженеров? Внешние факторы. //А. Орлов 2009. - презентация Автоматизация парковок и паркингов. Альянс-Т. Г. Тула
Автоматизация парковок и паркингов. Альянс-Т. Г. Тула Виды: основные, дополнительные, местные
Виды: основные, дополнительные, местные Корпускулярно-волновая природа света
Корпускулярно-волновая природа света  Презентация к уроку по окружающему миру по теме: «Полезные ископаемые» Учитель начальных классов МОУ СОШ №36 г. Тамбова Забавников
Презентация к уроку по окружающему миру по теме: «Полезные ископаемые» Учитель начальных классов МОУ СОШ №36 г. Тамбова Забавников Разработка модели дистанционно управляемого однопалубного лоцманского судна
Разработка модели дистанционно управляемого однопалубного лоцманского судна живопись византии
живопись византии Сибирский Федеральный Округ Подготовили студенты IV курса ФТД группы Т096 Дементьев , Джанвелян и Ефремов.
Сибирский Федеральный Округ Подготовили студенты IV курса ФТД группы Т096 Дементьев , Джанвелян и Ефремов. Базовые основания личности человека как фактор духовно-нравственного воспитания По Янушкявичене О.Л.
Базовые основания личности человека как фактор духовно-нравственного воспитания По Янушкявичене О.Л. Набор Здоровый ребенок
Набор Здоровый ребенок Христианство. Часть III. Таинства. Молитва
Христианство. Часть III. Таинства. Молитва Презентация на тему "Портфоліо клас" - скачать презентации по Педагогике
Презентация на тему "Портфоліо клас" - скачать презентации по Педагогике Внедрение систем мониторинга и защиты информации Информация – это своеобразный товар, как правило, весьма ценный. - презентация
Внедрение систем мониторинга и защиты информации Информация – это своеобразный товар, как правило, весьма ценный. - презентация Австрийская (психологическая) школа маржинализма Выполнили: Студенты 1 курса Экономического факультета Группы М 122 Б Зелепукин
Австрийская (психологическая) школа маржинализма Выполнили: Студенты 1 курса Экономического факультета Группы М 122 Б Зелепукин  Основы Swing
Основы Swing Школа человеческих отношений Выполнила студентка 2 курса ФТД группы Т-1202 Жукова Алёна
Школа человеческих отношений Выполнила студентка 2 курса ФТД группы Т-1202 Жукова Алёна  Видеонаблюдение от Уфанет
Видеонаблюдение от Уфанет Решение задач, примеров, уравнений. Закрепление - презентация для начальной школы_
Решение задач, примеров, уравнений. Закрепление - презентация для начальной школы_ ISO/IEC 17011:2017 Conformity Assessment - Requirements for accreditation bodies accrediting conformity assessment bodies
ISO/IEC 17011:2017 Conformity Assessment - Requirements for accreditation bodies accrediting conformity assessment bodies Браузер. Кем и когда создан
Браузер. Кем и когда создан Полномочия прокурора по надзору за исполнением законов органами, осуществляющими оперативно-розыскную деятельность
Полномочия прокурора по надзору за исполнением законов органами, осуществляющими оперативно-розыскную деятельность Облачные технологии (вычисления)
Облачные технологии (вычисления) 22 © 2010 Informatica. Company Confidential. Forward-looking information is based upon multiple assumptions and uncertainties and does not necessarily. - презентация
22 © 2010 Informatica. Company Confidential. Forward-looking information is based upon multiple assumptions and uncertainties and does not necessarily. - презентация