- Характеристика возбудимых тканей

Содержание
- 2. ПЛАН: Физиологические основы функций. Возбудимые ткани и их общие свойства. Раздражимость. Возбудимость как высокодифференцированная специализированная форма
- 3. В основе физиологических функций лежат общие свойства возбудимых тканей (мышечной, нервной, железистой) – раздражимость, возбудимость, проводимость,
- 4. Общие свойства возбудимых тканей: 1. Раздражимость – свойство всех тканей отвечать на действие слабых раздражителей кратковременной,
- 5. 4. Лабильность (функциональная подвижность) – определяется максимальным количеством импульсов возбуждения, которое способна провести ткань в единицу
- 6. Основные состояния возбудимых тканей: 1. Оперативный покой – состояние готовности клетки ответить на действие раздражителя. 2.
- 7. Раздражитель – любое, всякое изменение внешней или внутренней среды, возникающее достаточно быстро, действующее достаточно долго и
- 8. а – схема установки для фиксации и стимуляции мышцы: 1 - вертикальный миограф; 2 - икроножная
- 9. Порог раздражения (S) – минимальная интенсивность раздражителя, вызывающая минимальный специфический ответ. Как и лабильность, порог раздражения
- 10. амплитуда сокращения мышцы пороговый подпороговые раздражители сила раздражителей максимальный 40
- 11. При действии подпороговых раздражителей нет специфической реакции, но есть неспецифическая. Порог раздражения – это тот предел,
- 12. СОВРЕМЕННЫЕ ПРЕДСТАВЛЕНИЯ О СТРОЕНИИ И ФУНКЦИИ БИОЛОГИЧЕСКИХ МЕМБРАН Толщина мембраны 6 - 12 нм. Состоит из
- 13. Белки: интегральные (И) – погружены в мембрану (некоторые из них свободно перемещаются в мембране, а другие
- 14. каналы – трансмембранные пути, по которым между цитозолем и межклеточным пространством (и в обратном направлении) перемещаются
- 15. Один и тот же белок может быть одновременно рецептором и каналом (ацетилхолин-стимулируемые каналы постсинаптической мембраны нервно-мышечного
- 16. АНТИПОРТ ТРАНСПОРТ ВЕЩЕСТВ ЧЕРЕЗ МЕМБРАНУ Транспортёры специфичны: переносят через липидный бислой, как правило, одно вещество. Различают:
- 17. ВЗАИМОДЕЙСТВИЕ ПЕРЕНОСЧИКА И ТРАНСПОРТИРУЕМОЙ МОЛЕКУЛЫ Белки переносчика формируют канал, на внутренней поверхности которого имеется участок связывания
- 18. ИОННЫЕ КАНАЛЫ МЕМБРАН Это трансмембранные порообразующие белки, плотно упакованные в липидном бислое мембраны 48
- 19. ПОРА – ее канал всегда открыт. Поры формируют белки порины, перфорины, аквапорины, коннексины и др. В
- 20. Ионные каналы состоят из связанных между собой белковых субъединиц, образующих структуру со сложной пространственной конфигурацией, пронизывающих
- 21. Схема воротного механизма ионного канала. А. Ворота канала закрыты, ион не может пройти через мембрану. Б.
- 22. КЛАССИФИКАЦИЯ ИОННЫХ КАНАЛОВ МЕМБРАН По структуре (строению) и происхождению от однотипных генов – например, различают три
- 23. Наиболее часто встречаются два типа каналов ионные каналы с лиганд-зависимыми воротами – расположены на постсинаптической мембране
- 24. СВОЙСТВА КАНАЛОВ электровозбудимой мембраны: 1) селективность – это избирательно повышенная проницаемость ионного канала для определённых ионов
- 25. Способность чувствовать разность потенциалов определяется наличием потенциал-чувствительного домена (ПЧД), четвертый ТМ-сегмент (S4) которого несет большой положительный
- 26. ПОТЕНЦИАЛ-ЗАВИСИМЫЙ НАТРИЕВЫЙ КАНАЛ А – двумерная модель. I - IV – 4 домена, каждый из которых
- 27. m ворота – активационные h ворота – инактивационные, работают более медленно, расположены у внутреннего отверстия канала
- 28. МОДЕЛЬ КАЛИЕВОГО КАНАЛА В отличие от натриевых каналов в калиевых нет инактивационных ворот. Кинетика активационных ворот
- 29. Схема работы потенциал-чувствительного K+-канала: (1) Реполяризация (восстановление потенциала покоя −70 мВ) приводит к «вдвиганию» вольт-сенсора S4
- 30. УЧЕНИЕ О БИОПОТЕНЦИАЛАХ связано с Алессандро Вольта и Луиджи Гальвани – оппонентами в знаменитом в истории
- 31. Гальвани Л. «Трактат о силах электричества при мышечном движении» (1791): «Я разрезал и препарировал лягушку… и,
- 32. Схема опыта по изучению атмосферного электричества. Детектором служит лягушачья лапка, нерв которой соединен с громоотводом, а
- 33. В классическом опыте Луиджи Гальвани препарат задних конечностей лягушки подвешивался на цинковой стойке с помощью медного
- 34. Во втором опыте Гальвани (1794) наблюдал сокращение мышцы, если к ней прикладывались одновременно неповрежденный продольный участок
- 35. В 1840 году Маттеуччи показал, что сокращение мышцы нервно-мышечного препарата может наступить, если нерв этого препарата
- 36. МЕМБРАННЫЙ ПОТЕНЦИАЛ (МП) или ПОТЕНЦИАЛ ПОКОЯ – разность зарядов между наружной и внутренней поверхностями мембраны. В
- 37. ТЕОРИИ ЭЛЕКТРОГЕНЕЗА Классические: 1) диффузионно-ионная (Чаговец, 1896 ) – объясняет формирование биопотенциалов простой диффузией ионов; 2)
- 38. МЕМБРАННО-ИОННАЯ ТЕОРИЯ Условия для возникновения МП: 1. Избирательная проницаемость мембраны – пропускает одни вещества и не
- 39. В 1949 г. Алан Ходжкин и Бернард Катц рассчитали соотношение проницаемости (g) мембраны для различных ионов
- 40. 2. Ионная асимметрия – внутри клетки больше калия (в 30 - 50 раз), снаружи – натрия
- 41. ИОННАЯ АСИММЕТРИЯ Концентрация ионов во внутриклеточной жидкости Концентрация ионов во внеклеточной жидкости 71
- 42. 3. Работа ионного насоса – Na+,К+-АТФ-азы. Насос нагнетает калий в клетку, а натрий – во внеклеточную
- 43. Na+,К+-насос – интегральный мембранный белок. Состоит из 4 субъединиц (2 каталитические (α) – формируют канал и
- 44. Процессы при формировании МП: 1. Пассивные – диффузия по концентрационному и электрохимическому градиентам. 2. Активные: 1)
- 45. ФОРМИРОВАНИЕ МП По концентрационному градиенту ионы калия выходят из клетки и заряжают наружную поверхность мембраны положительно.
- 46. При перемещении калия из клетки по концентрационному градиенту совершается Аосм. Ионами калия, частично возвращающимися в клетку
- 47. Если подставить в уравнение Нернста константы, то при температуре тела EK+ равен: EK+ = –61 мВ·ln([K+вн]/[K+сн])
- 48. Измерения мембранного потенциала покоя [Электронный ресурс].- Режим доступа: http://bse.sci-lib.com/particle010601.html .- Дата доступа 20.02.2012. ИЗМЕРЕНИЕ ВЕЛИЧИНЫ МП
- 49. Физиология. Физиология человека. Передача информации посредством электрического возбуждения [Электронный ресурс].- Режим доступа: http://meduniver.com/Medical/Physiology/39.html.- Дата доступа 20.02.2012.
- 50. Метод локальной фиксации потенциала (patch-clamp): электрофизиологическая методика для изучения свойств ионных каналов, состоящая в том, что
- 51. Изменение величины МП: увеличение – гиперполяризация, уменьшение – деполяризация. ДЕПОЛЯРИЗАЦИЯ ГИПЕРПОЛЯРИЗАЦИЯ 81
- 52. ВЕЛИЧИНА МП ЗАВИСИТ ОТ ТИПА КЛЕТОК: МП плазмолеммы нервных клеток и кардиомиоцитов от –60 мВ до
- 53. ФУНКЦИИ МП: 1. Электрическое поле, рождаемое разностью зарядов, придает заряженным группам макромолекул мембраны определенную пространственную ориентацию.
- 54. ПОТЕНЦИАЛ ДЕЙСТВИЯ (ПД) Это колебание МП при возбуждении (в нервной клетке от -70 мВ до +50
- 55. m ворота –активационные h ворота – инактивационные Na Na Na ПОТЕНЦИАЛ ДЕЙСТВИЯ (ПД) При возбуждении резко
- 56. Натрий движется в клетку по концентрационному и электрохимическому («+» к «-») градиентам и нейтрализует отрицательный заряд
- 57. ЗАКРЫТЫ ИНАКТИВИРОВАНЫ СОСТОЯНИЕ НАТРИЕВЫХ КАНАЛОВ 87 Физиология. Физиология человека. Передача информации посредством электрического возбуждения [Электронный ресурс].-
- 58. ПД БЛИЗОК К НАТРИЕВОМУ РАВНОВЕСНОМУ ПОТЕНЦИАЛУ 88
- 59. Затем происходит восстановление МП – реполяризация. 1. Натриевая инактивация – резкое снижение проницаемости для натрия (закрываются
- 60. Помимо тока покоя и тока действия существуют биопотенциалы: 1. Локальный ответ. 2. Следовые потенциалы: отрицательный (замедление
- 61. Открытие натриевых каналов приводит к потоку ионов Na+ внутрь клетки. Это увеличивает начальную деполяризацию, что ведет
- 62. ВСЕ ЭЛЕКТРОВОЗБУДИМЫЕ МЕМБРАНЫ ЯВЛЯЮТСЯ МЕМБРАНАМИ РЕГЕНЕРАТОРНОГО ТИПА 92
- 63. Сначала проницаемость для натрия ненамного превосходит проницаемость для калия, который выходит из клетки и мешает деполяризации,
- 64. Если деполяризация не достигнет критического уровня, будет формироваться только локальный ответ. 94
- 65. КРИТИЧЕСКИЙ УРОВЕНЬ ДЕПОЛЯРИЗАЦИИ (КУД) ЗАВИСИТ ИСКЛЮЧИТЕЛЬНО ОТ СВОЙСТВ МЕМБРАНЫ: от количества инактивированных натриевых каналов (чем оно
- 66. Местное возбуждение Распространяющееся возбуждение ОСОБЕННОСТИ МЕСТНОГО И РАСПРОСТРАНЯЮЩЕГОСЯ ВОЗБУЖДЕНИЯ 96
- 67. При изменении МП изменяется возбудимость. Это связано с изменением порога деполяризации (Vt) – разницы между Ек
- 68. СООТНОШЕНИЕ ФАЗ ВОЗБУДИМОСТИ С ФАЗАМИ ПОТЕНЦИАЛА ДЕЙСТВИЯ. В момент локального ответа возбудимость повышается, в момент восходящей
- 69. РЕФРАКТЕРНЫЙ ПЕРИОД – ВРЕМЯ ПОСЛЕ ВОЗНИКНОВЕНИЯ ПОТЕНЦИАЛА ДЕЙСТВИЯ, В ХОДЕ КОТОРОГО ВОЗБУДИМОСТЬ МЕМБРАНЫ СНИЖАЕТСЯ, А ЗАТЕМ
- 71. Скачать презентацию

ПЛАН:
Физиологические основы функций. Возбудимые ткани и их общие свойства. Раздражимость. Возбудимость
ПЛАН: Физиологические основы функций. Возбудимые ткани и их общие свойства. Раздражимость. Возбудимость

В основе физиологических функций лежат общие свойства возбудимых тканей (мышечной, нервной,
В основе физиологических функций лежат общие свойства возбудимых тканей (мышечной, нервной,

Общие свойства возбудимых тканей:
1. Раздражимость – свойство всех тканей отвечать на
Общие свойства возбудимых тканей: 1. Раздражимость – свойство всех тканей отвечать на

4. Лабильность (функциональная подвижность) – определяется максимальным количеством импульсов возбуждения, которое
4. Лабильность (функциональная подвижность) – определяется максимальным количеством импульсов возбуждения, которое

Основные состояния возбудимых тканей:
1. Оперативный покой – состояние готовности клетки ответить
Основные состояния возбудимых тканей: 1. Оперативный покой – состояние готовности клетки ответить

Раздражитель – любое, всякое изменение внешней или внутренней среды, возникающее достаточно
Раздражитель – любое, всякое изменение внешней или внутренней среды, возникающее достаточно
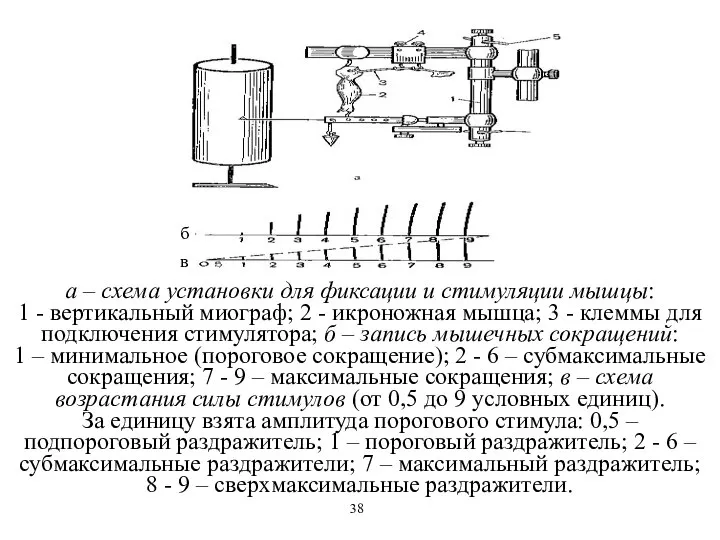
а – схема установки для фиксации и стимуляции мышцы:
1 - вертикальный
а – схема установки для фиксации и стимуляции мышцы: 1 - вертикальный
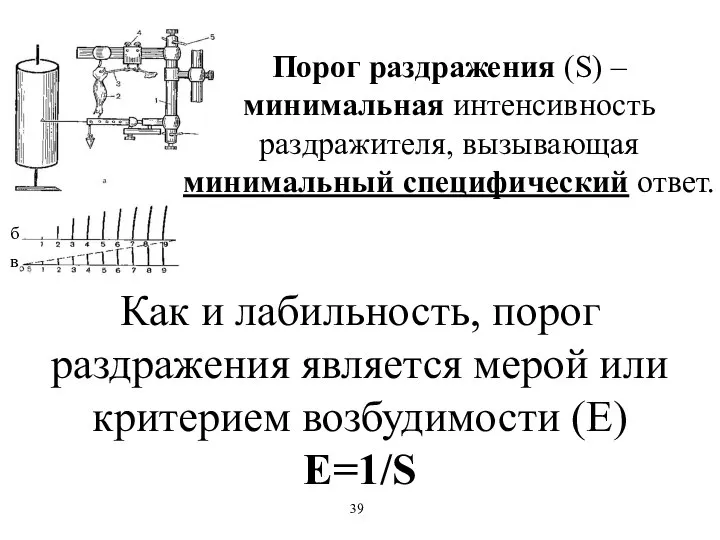
Порог раздражения (S) – минимальная интенсивность раздражителя, вызывающая минимальный специфический ответ.
Как
Порог раздражения (S) – минимальная интенсивность раздражителя, вызывающая минимальный специфический ответ.
Как
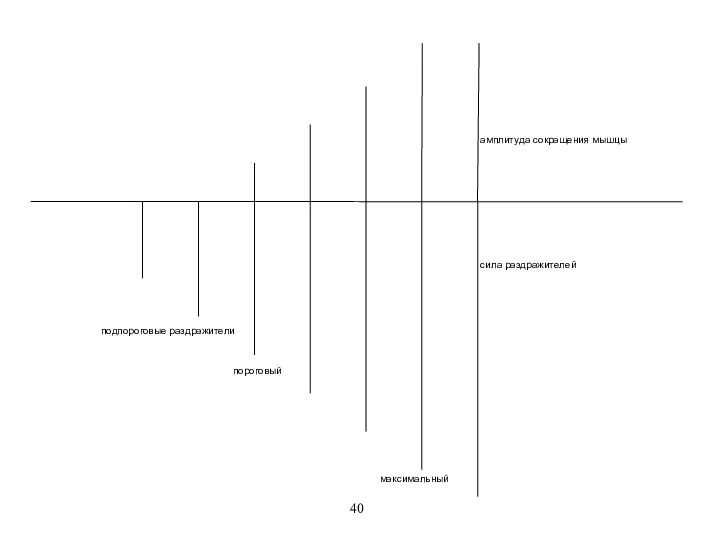
амплитуда сокращения мышцы
пороговый
подпороговые раздражители
сила раздражителей
максимальный
40
амплитуда сокращения мышцы
пороговый
подпороговые раздражители
сила раздражителей
максимальный
40

При действии подпороговых раздражителей нет специфической реакции, но есть неспецифическая.
Порог раздражения
При действии подпороговых раздражителей нет специфической реакции, но есть неспецифическая. Порог раздражения
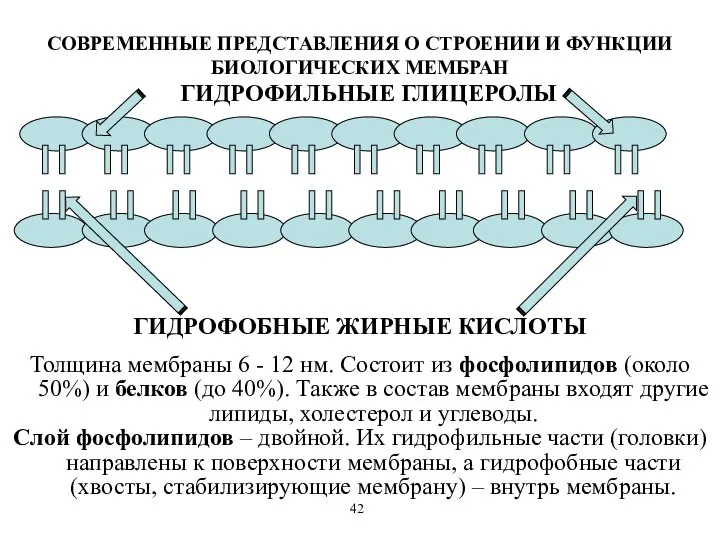
СОВРЕМЕННЫЕ ПРЕДСТАВЛЕНИЯ О СТРОЕНИИ И ФУНКЦИИ БИОЛОГИЧЕСКИХ МЕМБРАН
Толщина мембраны 6 -
СОВРЕМЕННЫЕ ПРЕДСТАВЛЕНИЯ О СТРОЕНИИ И ФУНКЦИИ БИОЛОГИЧЕСКИХ МЕМБРАН
Толщина мембраны 6 -
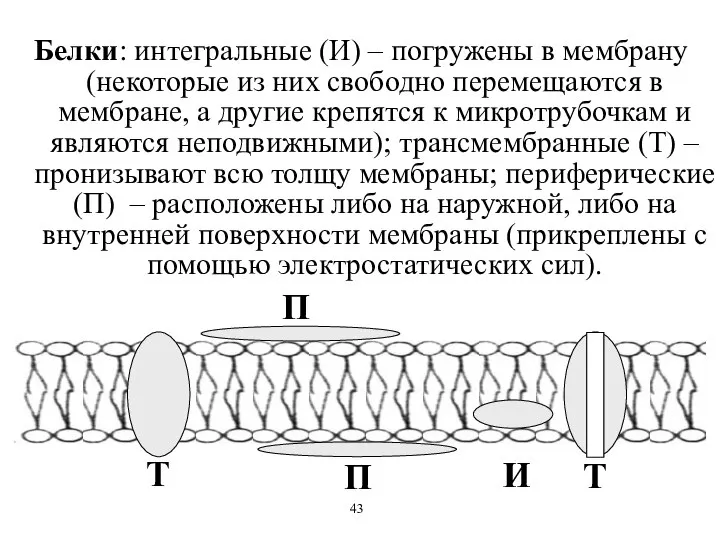
Белки: интегральные (И) – погружены в мембрану (некоторые из них свободно
Белки: интегральные (И) – погружены в мембрану (некоторые из них свободно
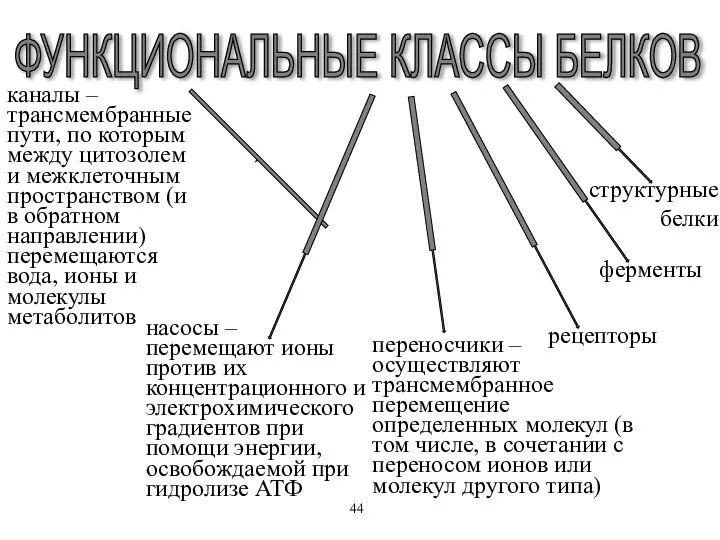
каналы – трансмембранные пути, по которым между цитозолем и межклеточным пространством
каналы – трансмембранные пути, по которым между цитозолем и межклеточным пространством
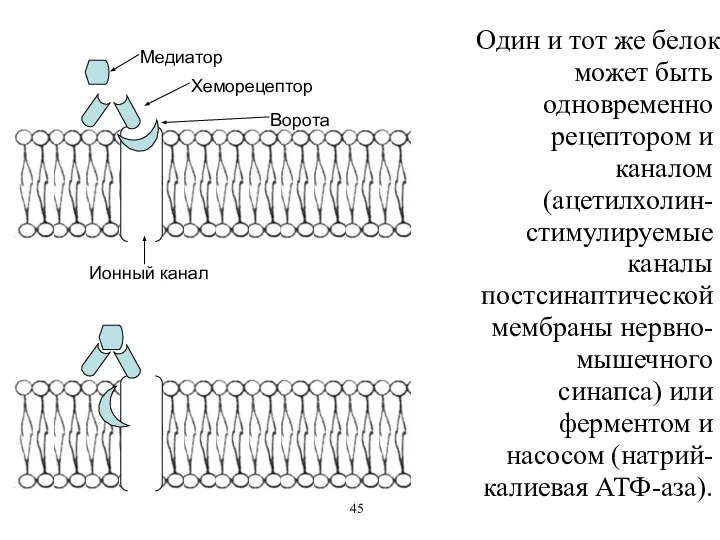
Один и тот же белок может быть одновременно рецептором и каналом
Один и тот же белок может быть одновременно рецептором и каналом
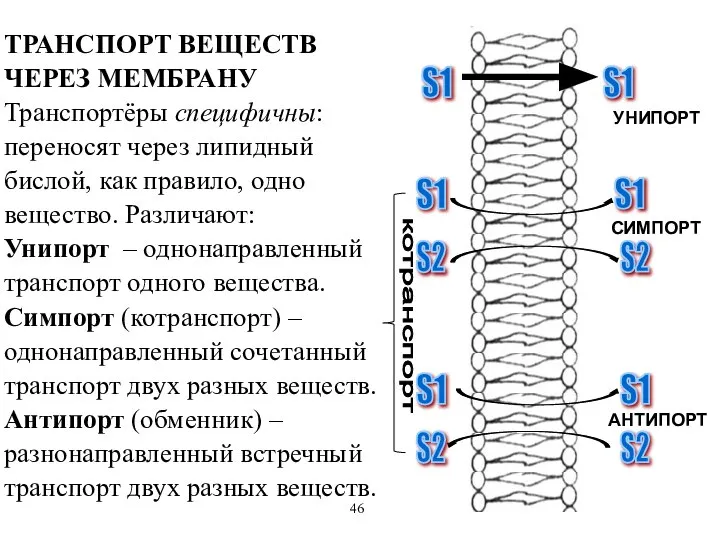
АНТИПОРТ
ТРАНСПОРТ ВЕЩЕСТВ ЧЕРЕЗ МЕМБРАНУ Транспортёры специфичны: переносят через липидный бислой, как
АНТИПОРТ
ТРАНСПОРТ ВЕЩЕСТВ ЧЕРЕЗ МЕМБРАНУ Транспортёры специфичны: переносят через липидный бислой, как
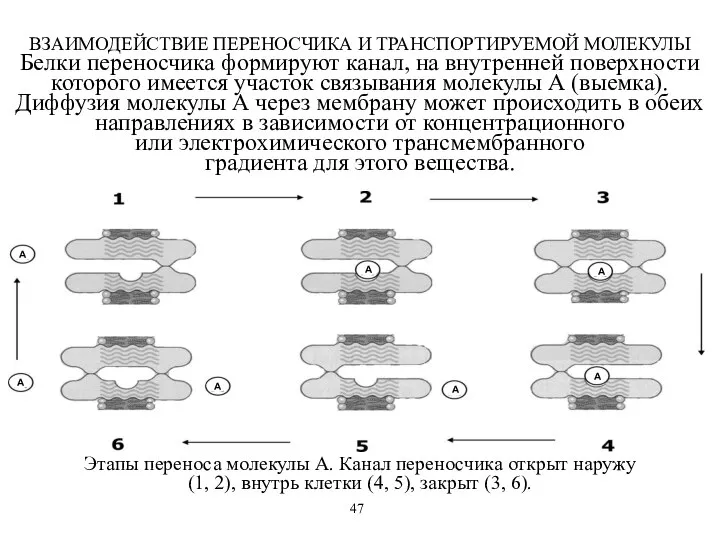
ВЗАИМОДЕЙСТВИЕ ПЕРЕНОСЧИКА И ТРАНСПОРТИРУЕМОЙ МОЛЕКУЛЫ
Белки переносчика формируют канал, на внутренней поверхности
ВЗАИМОДЕЙСТВИЕ ПЕРЕНОСЧИКА И ТРАНСПОРТИРУЕМОЙ МОЛЕКУЛЫ Белки переносчика формируют канал, на внутренней поверхности

ИОННЫЕ КАНАЛЫ МЕМБРАН
Это трансмембранные порообразующие белки,
плотно упакованные
в липидном бислое
ИОННЫЕ КАНАЛЫ МЕМБРАН
Это трансмембранные порообразующие белки, плотно упакованные в липидном бислое
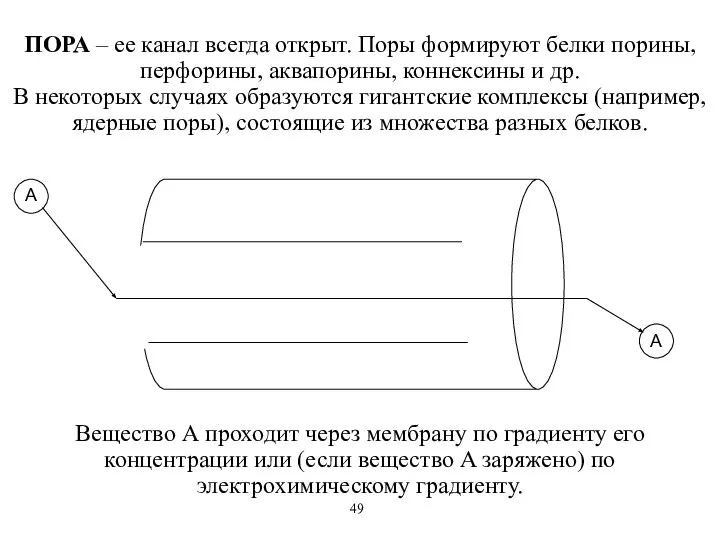
ПОРА – ее канал всегда открыт. Поры формируют белки порины, перфорины,
ПОРА – ее канал всегда открыт. Поры формируют белки порины, перфорины,
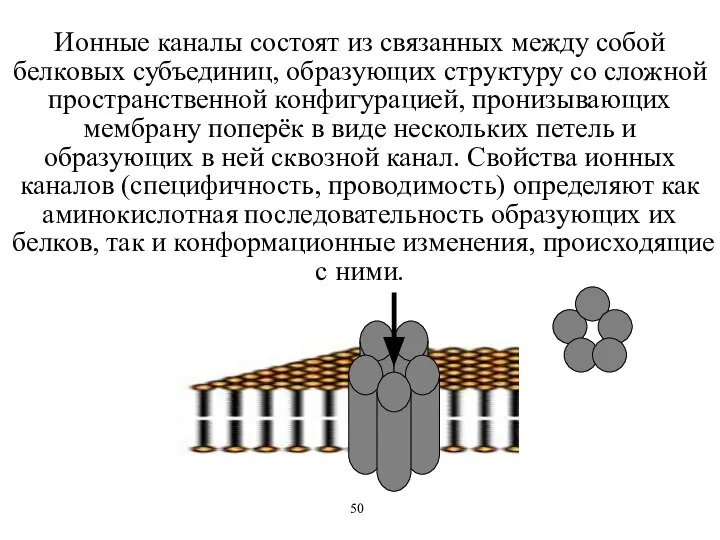
Ионные каналы состоят из связанных между собой белковых субъединиц, образующих структуру
Ионные каналы состоят из связанных между собой белковых субъединиц, образующих структуру
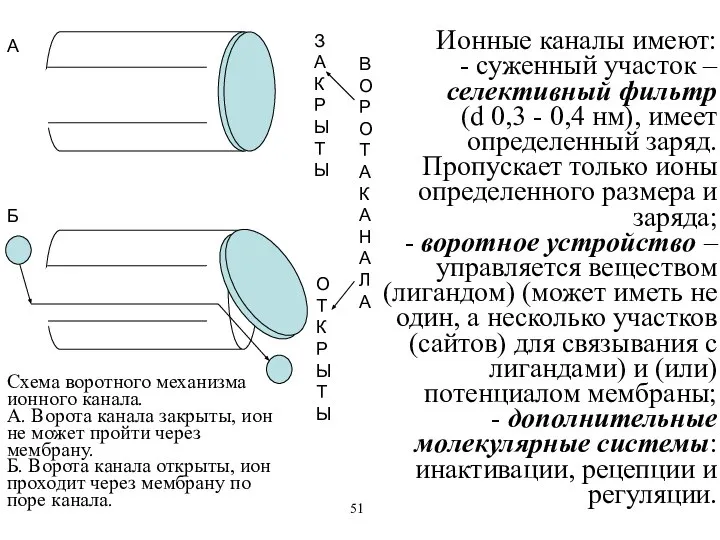
Схема воротного механизма ионного канала.
А. Ворота канала закрыты, ион не может
Схема воротного механизма ионного канала.
А. Ворота канала закрыты, ион не может

КЛАССИФИКАЦИЯ ИОННЫХ КАНАЛОВ МЕМБРАН
По структуре (строению) и происхождению от однотипных генов
КЛАССИФИКАЦИЯ ИОННЫХ КАНАЛОВ МЕМБРАН
По структуре (строению) и происхождению от однотипных генов
Наиболее часто встречаются два типа каналов
ионные каналы с лиганд-зависимыми воротами –
Наиболее часто встречаются два типа каналов
ионные каналы с лиганд-зависимыми воротами –
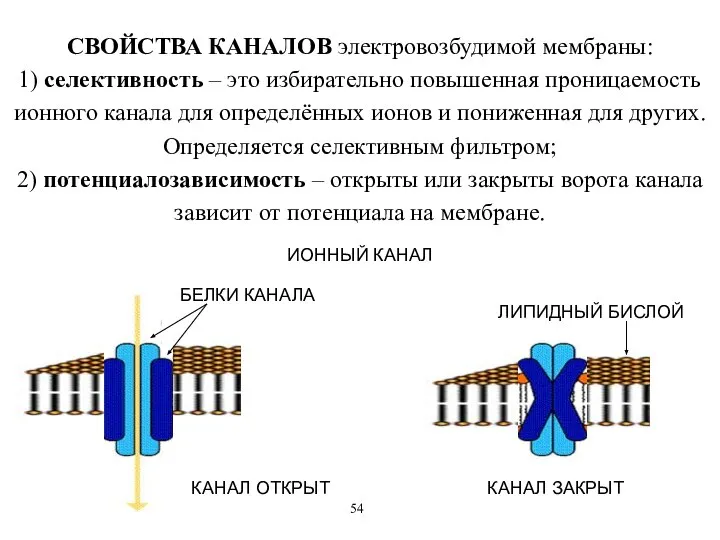
СВОЙСТВА КАНАЛОВ электровозбудимой мембраны:
1) селективность – это избирательно повышенная проницаемость ионного
СВОЙСТВА КАНАЛОВ электровозбудимой мембраны: 1) селективность – это избирательно повышенная проницаемость ионного
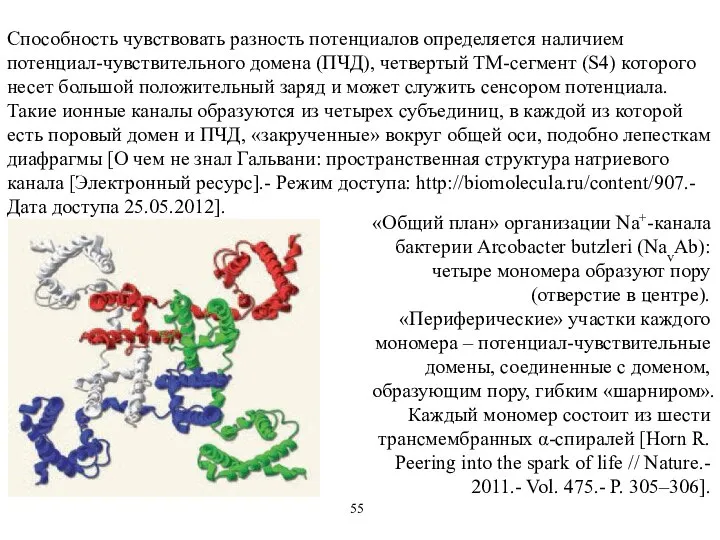
Способность чувствовать разность потенциалов определяется наличием потенциал-чувствительного домена (ПЧД), четвертый ТМ-сегмент
Способность чувствовать разность потенциалов определяется наличием потенциал-чувствительного домена (ПЧД), четвертый ТМ-сегмент
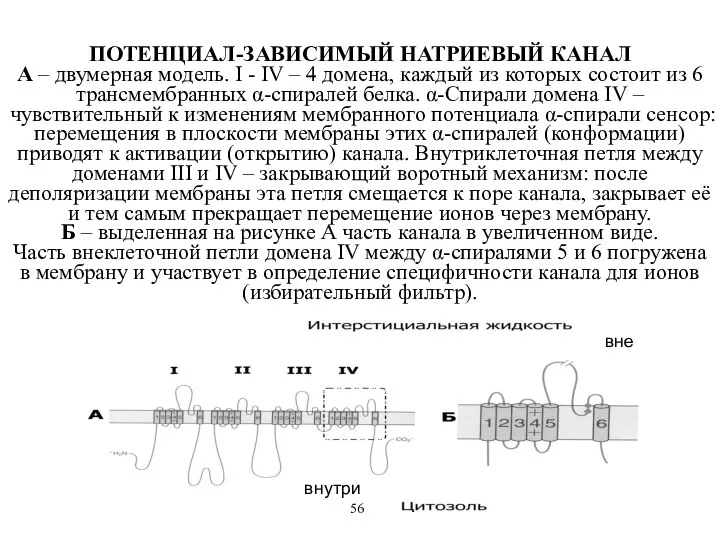
ПОТЕНЦИАЛ-ЗАВИСИМЫЙ НАТРИЕВЫЙ КАНАЛ
А – двумерная модель. I - IV – 4
ПОТЕНЦИАЛ-ЗАВИСИМЫЙ НАТРИЕВЫЙ КАНАЛ А – двумерная модель. I - IV – 4
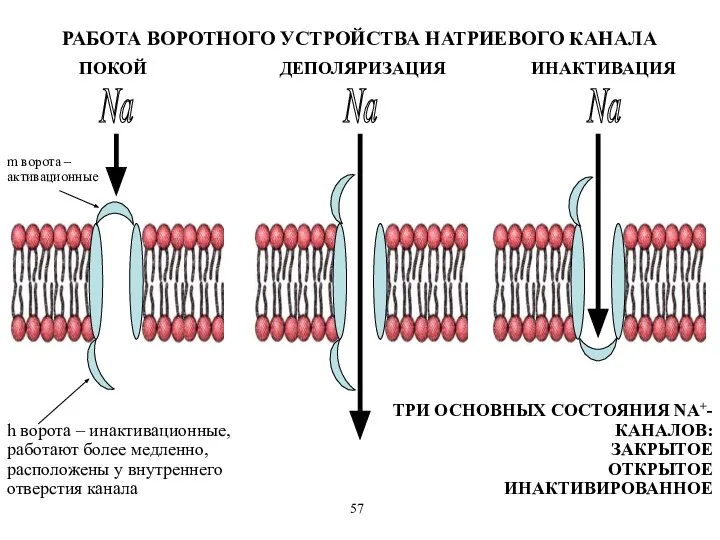
m ворота – активационные
h ворота – инактивационные, работают более медленно, расположены
m ворота – активационные
h ворота – инактивационные, работают более медленно, расположены
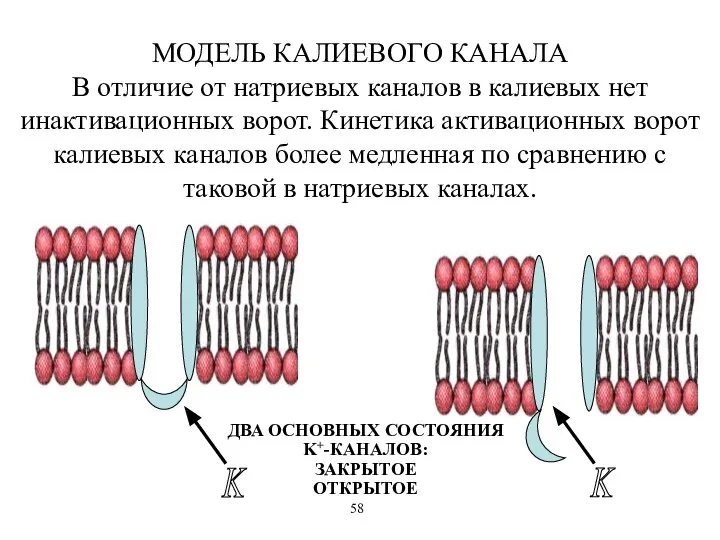
МОДЕЛЬ КАЛИЕВОГО КАНАЛА
В отличие от натриевых каналов в калиевых нет инактивационных
МОДЕЛЬ КАЛИЕВОГО КАНАЛА
В отличие от натриевых каналов в калиевых нет инактивационных
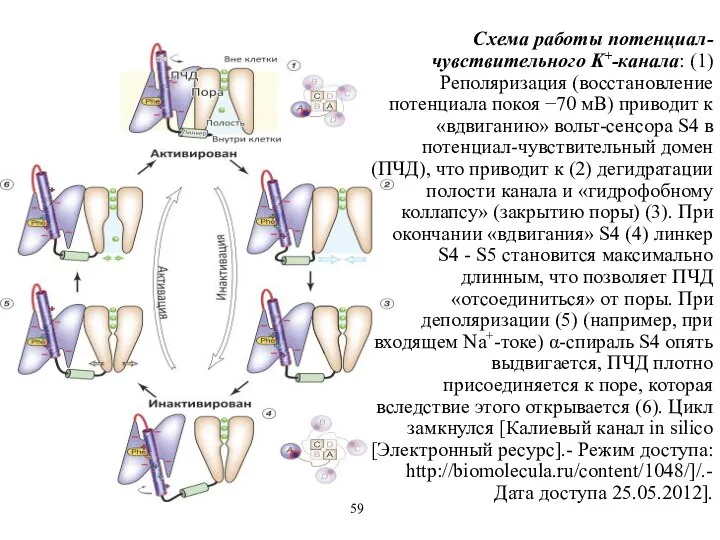
Схема работы потенциал-чувствительного K+-канала: (1) Реполяризация (восстановление потенциала покоя −70 мВ)
Схема работы потенциал-чувствительного K+-канала: (1) Реполяризация (восстановление потенциала покоя −70 мВ)

УЧЕНИЕ О БИОПОТЕНЦИАЛАХ
связано с Алессандро Вольта и Луиджи Гальвани – оппонентами
УЧЕНИЕ О БИОПОТЕНЦИАЛАХ связано с Алессандро Вольта и Луиджи Гальвани – оппонентами

Гальвани Л.
«Трактат о силах электричества при мышечном движении» (1791):
«Я разрезал и
Гальвани Л. «Трактат о силах электричества при мышечном движении» (1791): «Я разрезал и

Схема опыта по изучению атмосферного электричества. Детектором служит лягушачья лапка, нерв
Схема опыта по изучению атмосферного электричества. Детектором служит лягушачья лапка, нерв

В классическом опыте Луиджи Гальвани
препарат задних конечностей лягушки
подвешивался на цинковой стойке
В классическом опыте Луиджи Гальвани препарат задних конечностей лягушки подвешивался на цинковой стойке

Во втором опыте Гальвани (1794) наблюдал сокращение мышцы, если к ней
Во втором опыте Гальвани (1794) наблюдал сокращение мышцы, если к ней

В 1840 году Маттеуччи показал, что сокращение мышцы нервно-мышечного препарата может
В 1840 году Маттеуччи показал, что сокращение мышцы нервно-мышечного препарата может
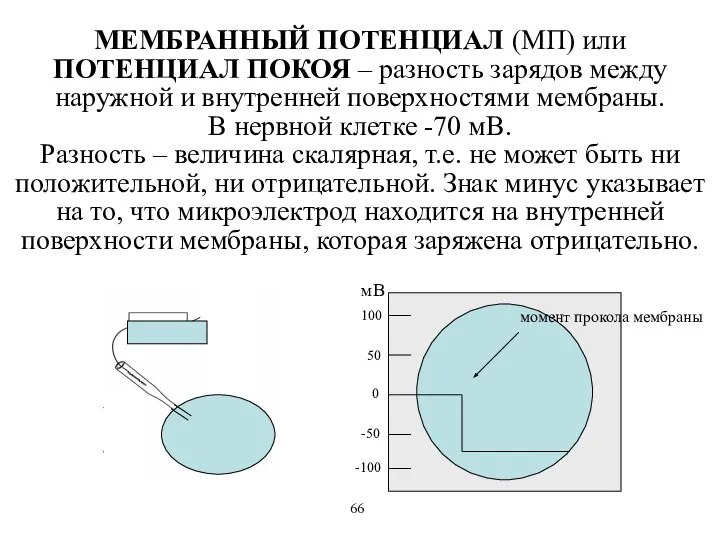
МЕМБРАННЫЙ ПОТЕНЦИАЛ (МП) или ПОТЕНЦИАЛ ПОКОЯ – разность зарядов между наружной
МЕМБРАННЫЙ ПОТЕНЦИАЛ (МП) или ПОТЕНЦИАЛ ПОКОЯ – разность зарядов между наружной

ТЕОРИИ ЭЛЕКТРОГЕНЕЗА
Классические:
1) диффузионно-ионная (Чаговец, 1896 ) – объясняет формирование биопотенциалов простой
диффузией
ТЕОРИИ ЭЛЕКТРОГЕНЕЗА
Классические: 1) диффузионно-ионная (Чаговец, 1896 ) – объясняет формирование биопотенциалов простой диффузией
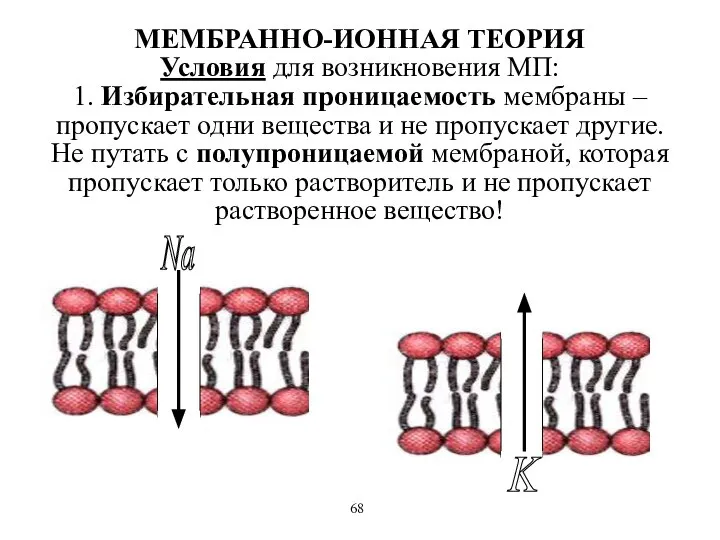
МЕМБРАННО-ИОННАЯ ТЕОРИЯ
Условия для возникновения МП:
1. Избирательная проницаемость мембраны – пропускает одни
МЕМБРАННО-ИОННАЯ ТЕОРИЯ Условия для возникновения МП: 1. Избирательная проницаемость мембраны – пропускает одни

В 1949 г. Алан Ходжкин и Бернард Катц рассчитали соотношение проницаемости (g)
В 1949 г. Алан Ходжкин и Бернард Катц рассчитали соотношение проницаемости (g)

2. Ионная асимметрия – внутри клетки больше калия (в 30 -
2. Ионная асимметрия – внутри клетки больше калия (в 30 -
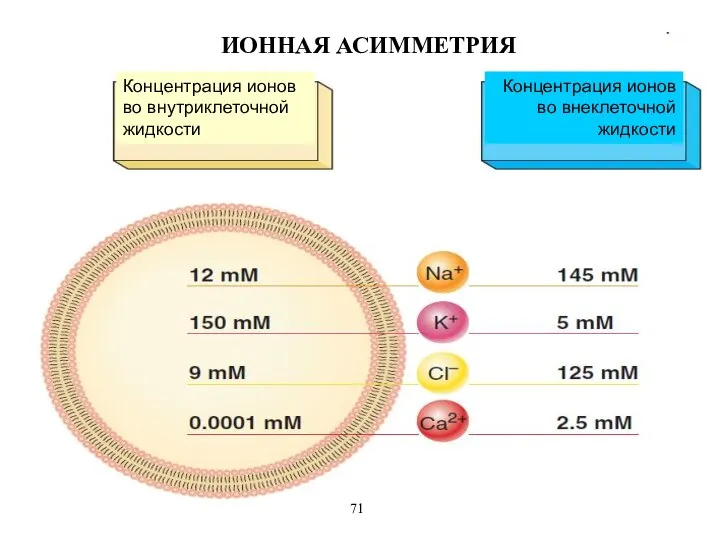
ИОННАЯ АСИММЕТРИЯ
Концентрация ионов во внутриклеточной жидкости
Концентрация ионов во внеклеточной жидкости
71
ИОННАЯ АСИММЕТРИЯ
Концентрация ионов во внутриклеточной жидкости
Концентрация ионов во внеклеточной жидкости
71
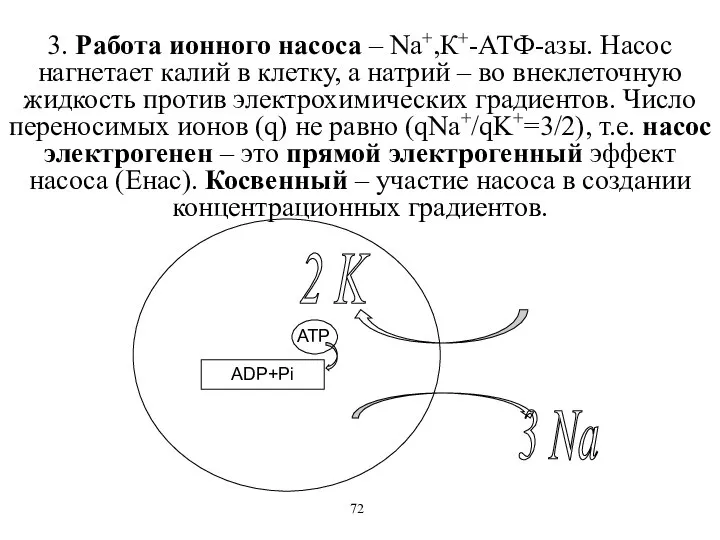
3. Работа ионного насоса – Na+,К+-АТФ-азы. Насос нагнетает калий в клетку,
3. Работа ионного насоса – Na+,К+-АТФ-азы. Насос нагнетает калий в клетку,
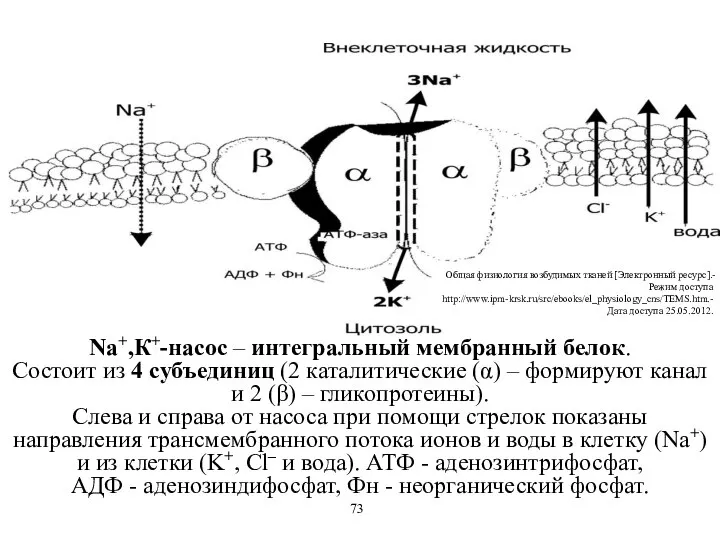
Na+,К+-насос – интегральный мембранный белок.
Состоит из 4 субъединиц (2 каталитические (α)
Na+,К+-насос – интегральный мембранный белок. Состоит из 4 субъединиц (2 каталитические (α)
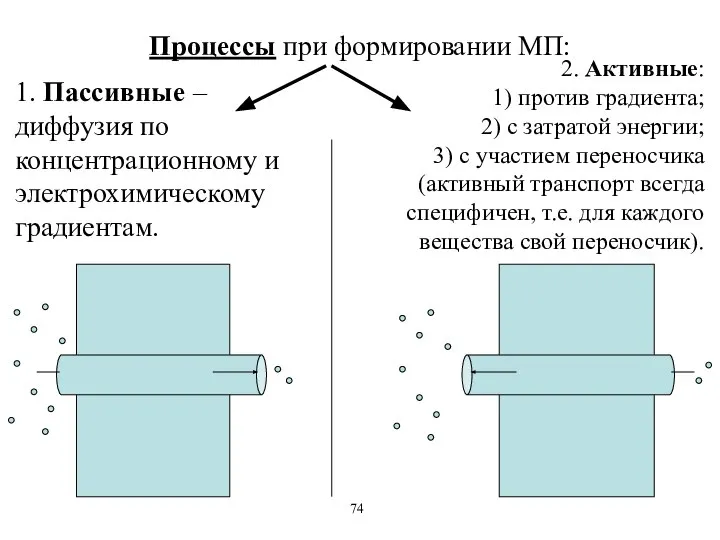
Процессы при формировании МП:
1. Пассивные – диффузия по концентрационному и электрохимическому
Процессы при формировании МП:
1. Пассивные – диффузия по концентрационному и электрохимическому
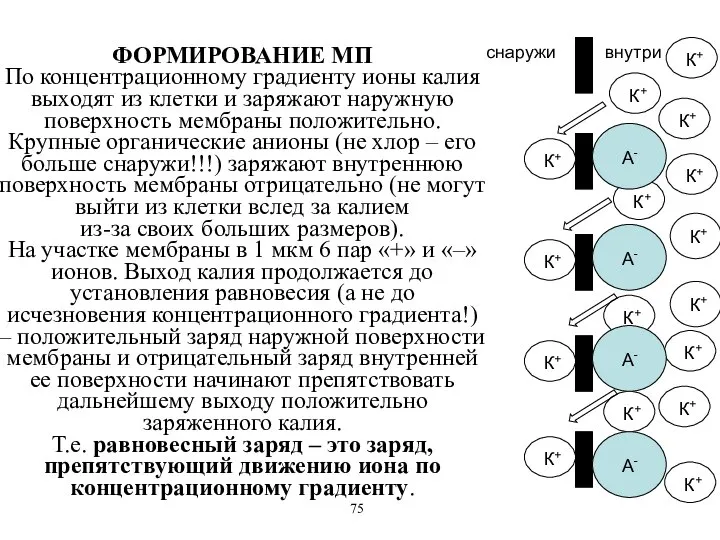
ФОРМИРОВАНИЕ МП
По концентрационному градиенту ионы калия выходят из клетки и заряжают
ФОРМИРОВАНИЕ МП По концентрационному градиенту ионы калия выходят из клетки и заряжают
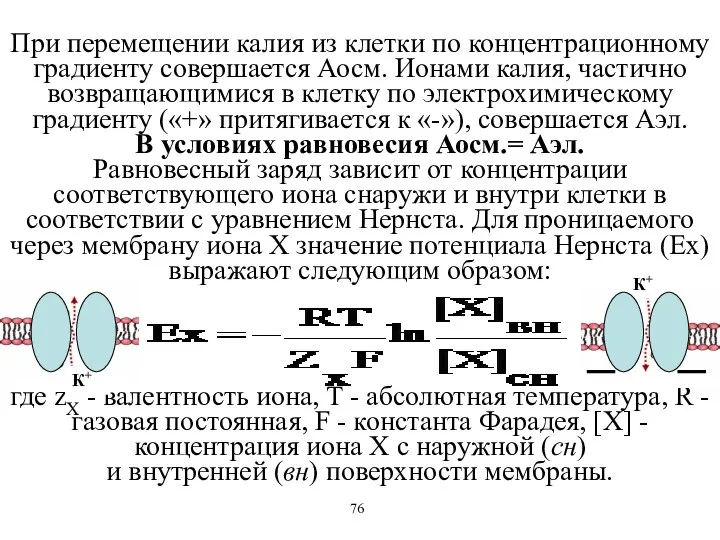
При перемещении калия из клетки по концентрационному градиенту совершается Аосм. Ионами
При перемещении калия из клетки по концентрационному градиенту совершается Аосм. Ионами

Если подставить в уравнение Нернста константы, то при температуре тела EK+
Если подставить в уравнение Нернста константы, то при температуре тела EK+
![Измерения мембранного потенциала покоя [Электронный ресурс].- Режим доступа: http://bse.sci-lib.com/particle010601.html .- Дата](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1290528/slide-47.jpg)
Измерения мембранного потенциала покоя [Электронный ресурс].- Режим доступа: http://bse.sci-lib.com/particle010601.html .- Дата
Измерения мембранного потенциала покоя [Электронный ресурс].- Режим доступа: http://bse.sci-lib.com/particle010601.html .- Дата
![Физиология. Физиология человека. Передача информации посредством электрического возбуждения [Электронный ресурс].- Режим](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1290528/slide-48.jpg)
Физиология. Физиология человека. Передача информации посредством электрического возбуждения [Электронный ресурс].- Режим
Физиология. Физиология человека. Передача информации посредством электрического возбуждения [Электронный ресурс].- Режим

Метод локальной фиксации потенциала (patch-clamp): электрофизиологическая методика для изучения свойств ионных
Метод локальной фиксации потенциала (patch-clamp): электрофизиологическая методика для изучения свойств ионных
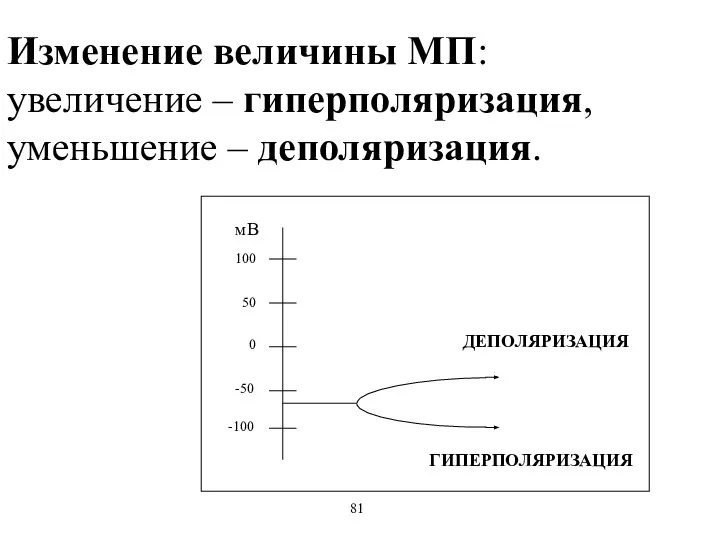
Изменение величины МП:
увеличение – гиперполяризация, уменьшение – деполяризация.
ДЕПОЛЯРИЗАЦИЯ
ГИПЕРПОЛЯРИЗАЦИЯ
81
Изменение величины МП:
увеличение – гиперполяризация, уменьшение – деполяризация.
ДЕПОЛЯРИЗАЦИЯ
ГИПЕРПОЛЯРИЗАЦИЯ
81
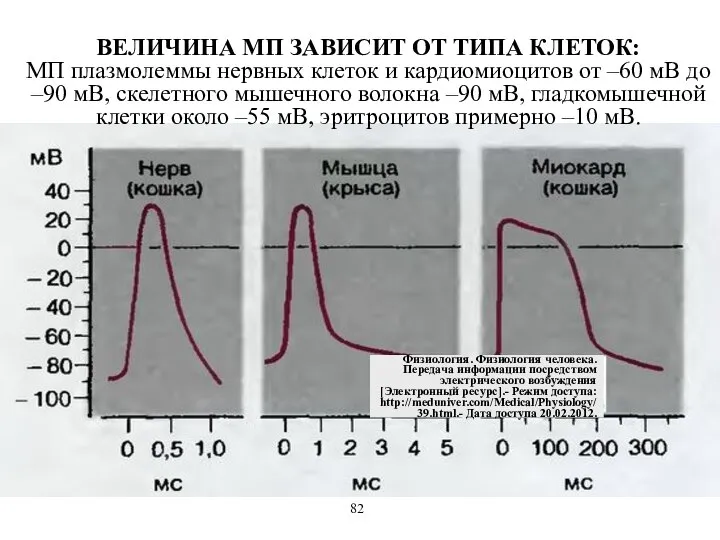
ВЕЛИЧИНА МП ЗАВИСИТ ОТ ТИПА КЛЕТОК:
МП плазмолеммы нервных клеток и кардиомиоцитов
ВЕЛИЧИНА МП ЗАВИСИТ ОТ ТИПА КЛЕТОК: МП плазмолеммы нервных клеток и кардиомиоцитов
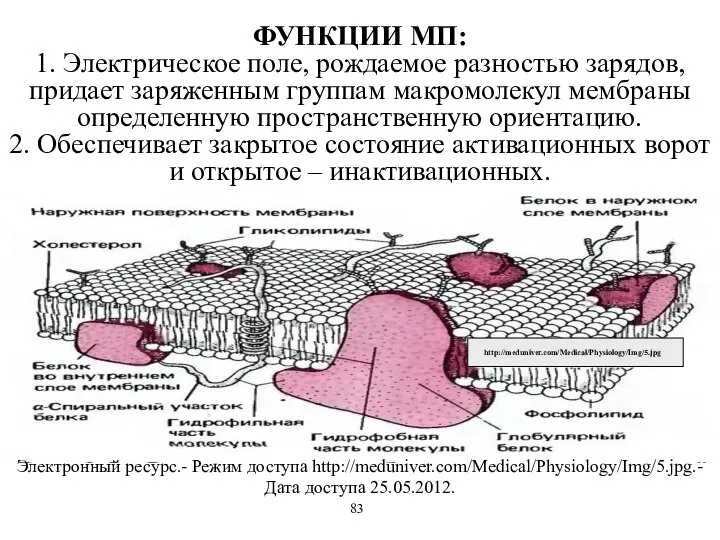
ФУНКЦИИ МП:
1. Электрическое поле, рождаемое разностью зарядов, придает заряженным группам макромолекул
ФУНКЦИИ МП: 1. Электрическое поле, рождаемое разностью зарядов, придает заряженным группам макромолекул
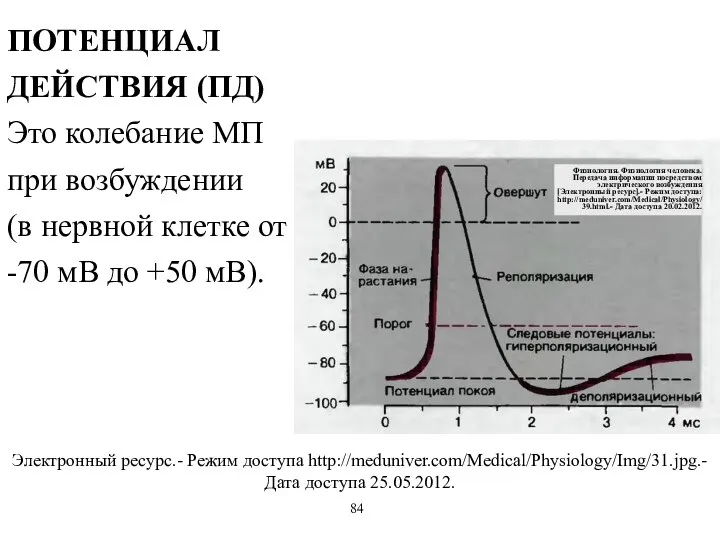
ПОТЕНЦИАЛ ДЕЙСТВИЯ (ПД)
Это колебание МП при возбуждении
(в нервной клетке от -70
ПОТЕНЦИАЛ ДЕЙСТВИЯ (ПД) Это колебание МП при возбуждении (в нервной клетке от -70
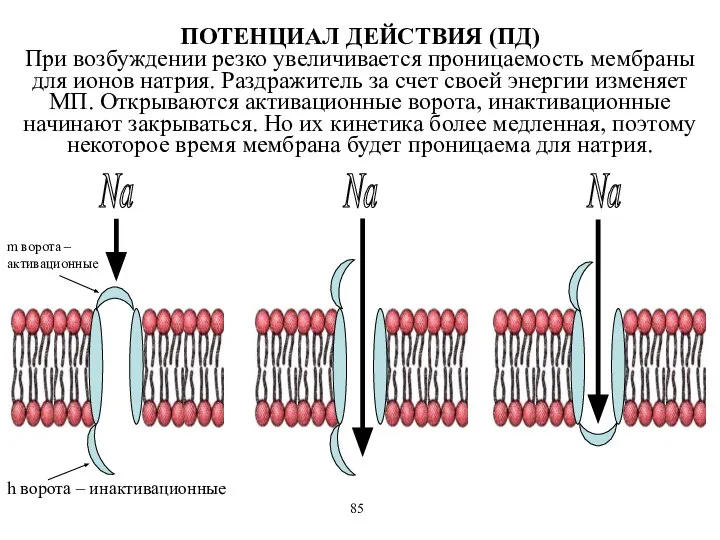
m ворота –активационные
h ворота – инактивационные
Na
Na
Na
ПОТЕНЦИАЛ ДЕЙСТВИЯ (ПД)
При возбуждении
m ворота –активационные
h ворота – инактивационные
Na
Na
Na
ПОТЕНЦИАЛ ДЕЙСТВИЯ (ПД) При возбуждении
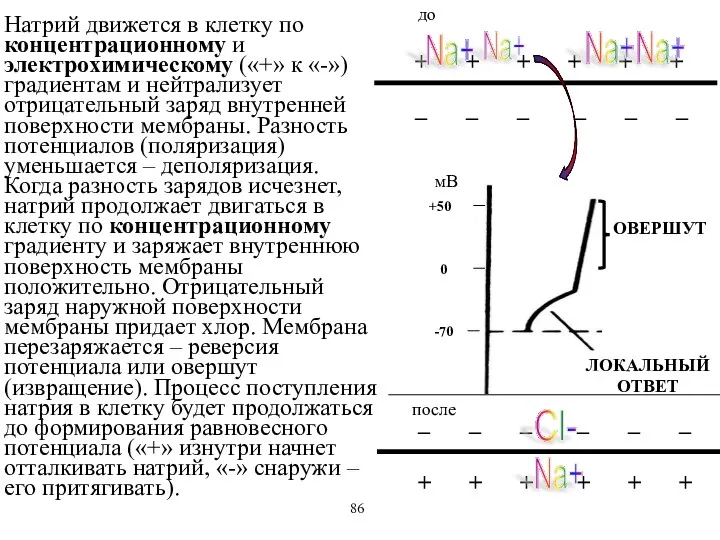
Натрий движется в клетку по концентрационному и электрохимическому («+» к «-»)
Натрий движется в клетку по концентрационному и электрохимическому («+» к «-»)
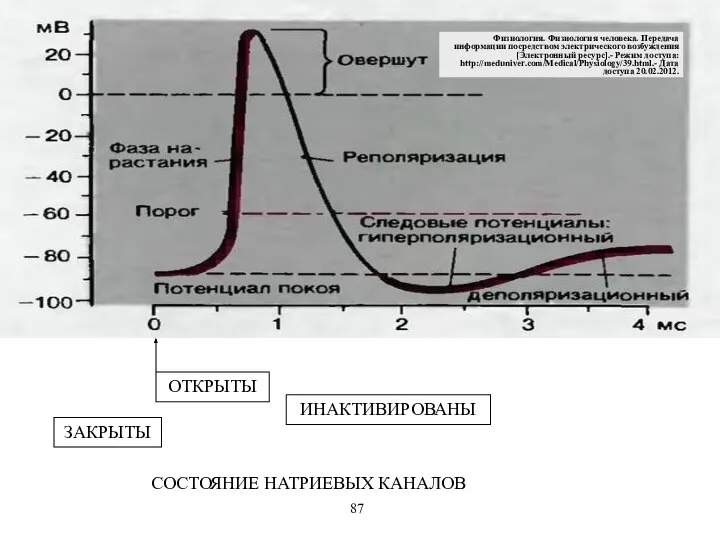
ЗАКРЫТЫ
ИНАКТИВИРОВАНЫ
СОСТОЯНИЕ НАТРИЕВЫХ КАНАЛОВ
87
Физиология. Физиология человека. Передача информации посредством электрического возбуждения [Электронный
ЗАКРЫТЫ
ИНАКТИВИРОВАНЫ
СОСТОЯНИЕ НАТРИЕВЫХ КАНАЛОВ
87
Физиология. Физиология человека. Передача информации посредством электрического возбуждения [Электронный

ПД БЛИЗОК К НАТРИЕВОМУ
РАВНОВЕСНОМУ ПОТЕНЦИАЛУ
88
ПД БЛИЗОК К НАТРИЕВОМУ
РАВНОВЕСНОМУ ПОТЕНЦИАЛУ
88
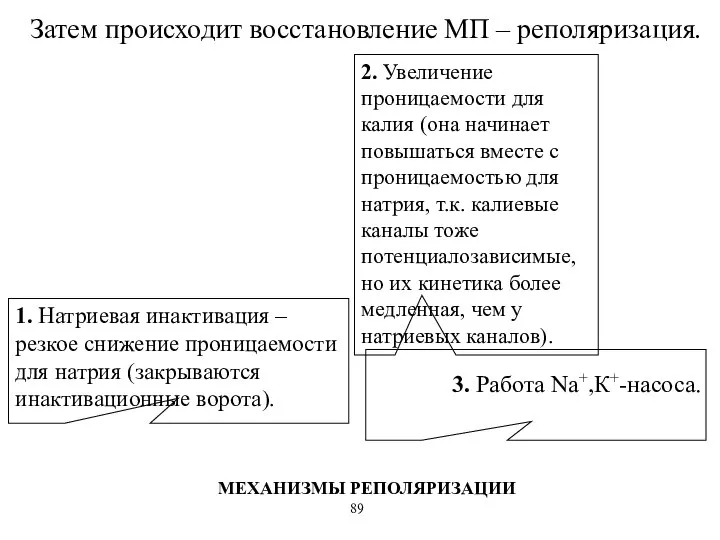
Затем происходит восстановление МП – реполяризация.
1. Натриевая инактивация – резкое снижение
Затем происходит восстановление МП – реполяризация.
1. Натриевая инактивация – резкое снижение
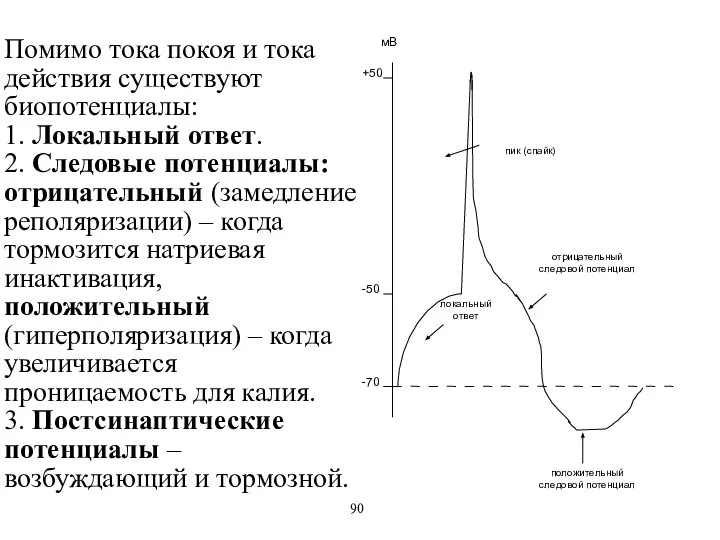
Помимо тока покоя и тока действия существуют биопотенциалы:
1. Локальный ответ.
2. Следовые
Помимо тока покоя и тока действия существуют биопотенциалы: 1. Локальный ответ. 2. Следовые

Открытие натриевых каналов приводит к потоку ионов Na+ внутрь клетки. Это
Открытие натриевых каналов приводит к потоку ионов Na+ внутрь клетки. Это

ВСЕ ЭЛЕКТРОВОЗБУДИМЫЕ МЕМБРАНЫ ЯВЛЯЮТСЯ МЕМБРАНАМИ РЕГЕНЕРАТОРНОГО ТИПА
92
ВСЕ ЭЛЕКТРОВОЗБУДИМЫЕ МЕМБРАНЫ ЯВЛЯЮТСЯ МЕМБРАНАМИ РЕГЕНЕРАТОРНОГО ТИПА
92
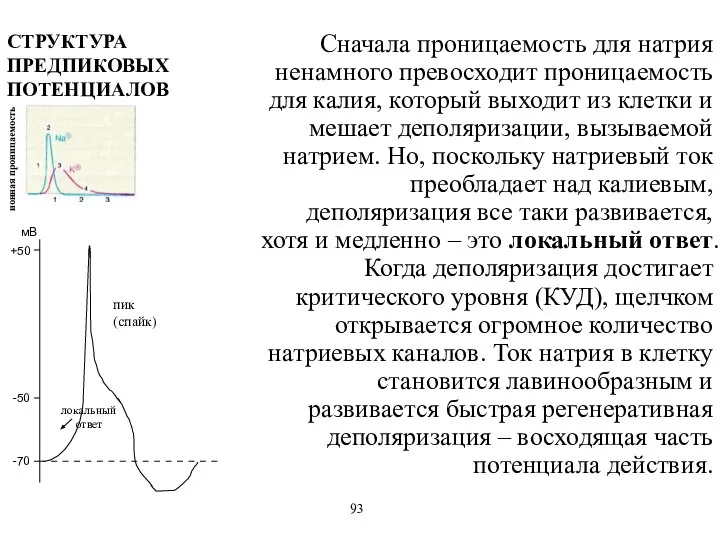
Сначала проницаемость для натрия ненамного превосходит проницаемость для калия, который выходит
Сначала проницаемость для натрия ненамного превосходит проницаемость для калия, который выходит
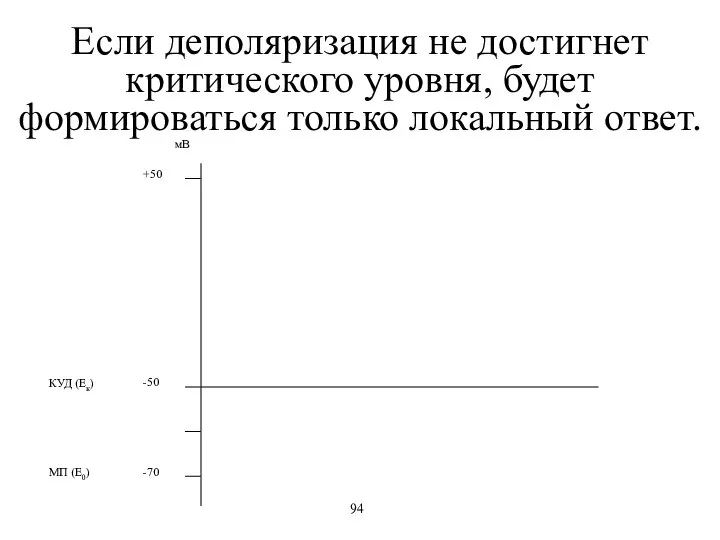
Если деполяризация не достигнет критического уровня, будет формироваться только локальный ответ.
94
Если деполяризация не достигнет критического уровня, будет формироваться только локальный ответ.
94

КРИТИЧЕСКИЙ УРОВЕНЬ ДЕПОЛЯРИЗАЦИИ (КУД) ЗАВИСИТ ИСКЛЮЧИТЕЛЬНО ОТ СВОЙСТВ МЕМБРАНЫ:
от количества инактивированных
КРИТИЧЕСКИЙ УРОВЕНЬ ДЕПОЛЯРИЗАЦИИ (КУД) ЗАВИСИТ ИСКЛЮЧИТЕЛЬНО ОТ СВОЙСТВ МЕМБРАНЫ:
от количества инактивированных

Местное возбуждение
Распространяющееся возбуждение
ОСОБЕННОСТИ МЕСТНОГО И РАСПРОСТРАНЯЮЩЕГОСЯ ВОЗБУЖДЕНИЯ
96
Местное возбуждение
Распространяющееся возбуждение
ОСОБЕННОСТИ МЕСТНОГО И РАСПРОСТРАНЯЮЩЕГОСЯ ВОЗБУЖДЕНИЯ
96
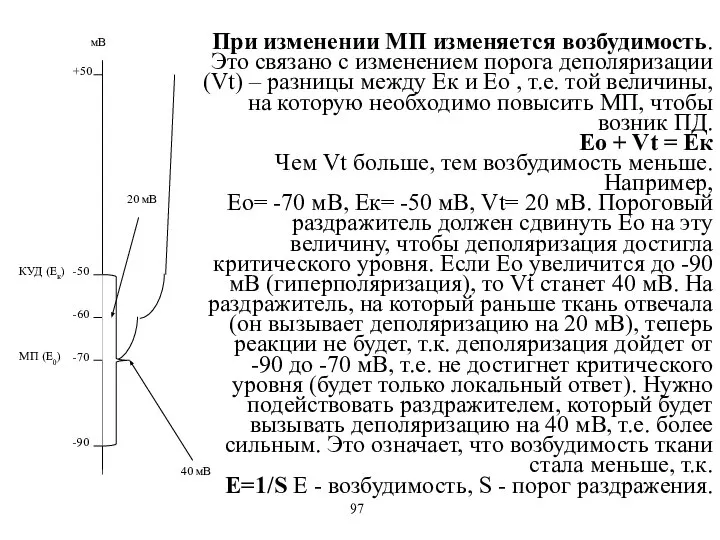
При изменении МП изменяется возбудимость. Это связано с изменением порога деполяризации
При изменении МП изменяется возбудимость. Это связано с изменением порога деполяризации
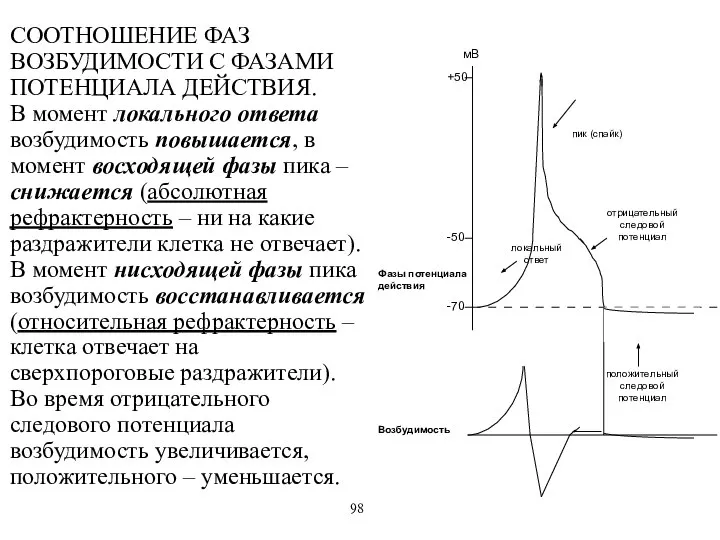
СООТНОШЕНИЕ ФАЗ ВОЗБУДИМОСТИ С ФАЗАМИ ПОТЕНЦИАЛА ДЕЙСТВИЯ.
В момент локального ответа возбудимость
СООТНОШЕНИЕ ФАЗ ВОЗБУДИМОСТИ С ФАЗАМИ ПОТЕНЦИАЛА ДЕЙСТВИЯ. В момент локального ответа возбудимость
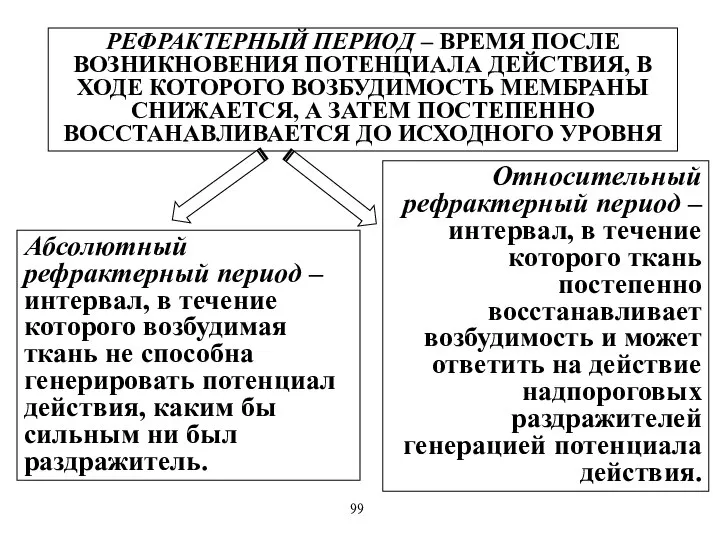
РЕФРАКТЕРНЫЙ ПЕРИОД – ВРЕМЯ ПОСЛЕ ВОЗНИКНОВЕНИЯ ПОТЕНЦИАЛА ДЕЙСТВИЯ, В ХОДЕ КОТОРОГО
РЕФРАКТЕРНЫЙ ПЕРИОД – ВРЕМЯ ПОСЛЕ ВОЗНИКНОВЕНИЯ ПОТЕНЦИАЛА ДЕЙСТВИЯ, В ХОДЕ КОТОРОГО
 Моделирование как технология формирования безопасного поведения детей МБОУ ВМР «Кипеловская средняя общеобразовательная шк
Моделирование как технология формирования безопасного поведения детей МБОУ ВМР «Кипеловская средняя общеобразовательная шк Правовой режим различных категорий земель РФ
Правовой режим различных категорий земель РФ Как оформить исследовательский проект
Как оформить исследовательский проект Практика увеличения продаж при помощи персональных email-рекомендаций Владислав Флакс Электронная торговля Москва, 24 мая 2011. - презе
Практика увеличения продаж при помощи персональных email-рекомендаций Владислав Флакс Электронная торговля Москва, 24 мая 2011. - презе Искусство XX века Авангардизм
Искусство XX века Авангардизм .09.19
.09.19 Основные свойства и применение проводниковых материалов
Основные свойства и применение проводниковых материалов Презентация Исковое заявление к Владивостокской таможне
Презентация Исковое заявление к Владивостокской таможне  Лекция 2 Система проектной деятельности в РФ 2018
Лекция 2 Система проектной деятельности в РФ 2018 Практическая часть работы супервайзера
Практическая часть работы супервайзера Базы данных Язык запросов SQL. Команда SELECT
Базы данных Язык запросов SQL. Команда SELECT Игровые технологии
Игровые технологии КЕСТЕ ЭКРАНЫНЫҢ ӨЗГЕРУІ ЖӘНЕ БОС ОРЫН
КЕСТЕ ЭКРАНЫНЫҢ ӨЗГЕРУІ ЖӘНЕ БОС ОРЫН Каргопольская глиняная игрушка
Каргопольская глиняная игрушка Презентация АНАЛИЗ РЫНКА РЫБЫ И РЫБНОЙ ПРОДУКЦИИ
Презентация АНАЛИЗ РЫНКА РЫБЫ И РЫБНОЙ ПРОДУКЦИИ  Занятие №2
Занятие №2 Программирование на языке Паскаль
Программирование на языке Паскаль 25 января - Татьянин день
25 января - Татьянин день Текстильные материалы трикотаж
Текстильные материалы трикотаж  DeVita Ritm base - цифровое устройство оздоровления
DeVita Ritm base - цифровое устройство оздоровления Религии Древнего Востока
Религии Древнего Востока Организация ЭВМ и вычислительных систем. История развития ВТ и классификация ЭВМ
Организация ЭВМ и вычислительных систем. История развития ВТ и классификация ЭВМ Инновационные образовательные программы Заседание методического Совета РЭА 8.02.2007
Инновационные образовательные программы Заседание методического Совета РЭА 8.02.2007 Один код, две платформы: миф или реальность
Один код, две платформы: миф или реальность Дистанционный семинар-тренинг «Разработка проектов и заявок на гранты. Фандрайзинг»
Дистанционный семинар-тренинг «Разработка проектов и заявок на гранты. Фандрайзинг» Документы как объект получения информации
Документы как объект получения информации Числовые промежутки. Алгебра 8 класс
Числовые промежутки. Алгебра 8 класс Серия терактов в Париже
Серия терактов в Париже