- Анализаторы. Органы чувств человека

Содержание
- 2. Орган зрения включает: 1. глазное яблоко, соединённое через зрительный нерв с мозгом, 2. защитный аппарат (веки
- 3. Оптика глаза Глаз имеет систему линз с различной кривизной и различными показателями преломления световых лучей, включающую
- 4. Преломляющая сила Для расчетов преломляющей силы глаза используют понятие о так называемом «редуцированном глазе», когда все
- 5. Аккомодация - это приспособление глаза к чёткому видению предметов, расположенных на различном расстоянии. Основная роль в
- 6. ПСНС обеспечивает фокусировку при приближении предмета к глазу. СНС незначительно расслабляет ресничную мышцу, но это практически
- 7. Зрачковый рефлекс Просвет зрачка может изменяться от 1 мм до 8 мм. Это придаёт зрачку свойства
- 8. Регуляция зрачкового рефлекса: 1. ПСНС (ацетилхолин и эзерин) вызывают сужение зрачка, а блокада холинорецепторов сфинктера радужки
- 9. Сфинктер иннервируется парасимпатическими нервными волокнами, выходящими из цилиарного (ресничного) ганглия, расположенного позади глаза. Преганглионарные волокна отходят
- 10. Зрачковый рефлекс меняется при заболеваниях ЦНС: а. Торможение возникает при нарушении передачи импульсов от сетчатки к
- 11. Содружественная реакция зрачков У здоровых людей зрачки обоих глаз одинакового размера. Освещение одного глаза ведет к
- 12. Рефракция Эмметропия (нормальное зрение) - параллельные лучи от отдалённых предметов фокусируются на сетчатке, когда ресничная мышца
- 13. Аномалии рефракции Миопия (близорукость) - длинное глазное яблоко либо большая преломляющая сила хрусталика (фокус впереди сетчатки).
- 14. Астигматизм — неодинаковое преломление лучей в разных направлениях, вызванное различной кривизной сферической поверхности роговицы. Аккомодация глаза
- 15. Поля зрения Зрительное поле каждого глаза — часть внешнего пространства, видимого глазом (теоретически оно должно быть
- 16. Бинокулярное зрение Центральная часть зрительных полей двух глаз полностью совпадает; следовательно, любой участок в этом зрительном
- 17. Точки на сетчатке обоих глаз, куда должно попадать изображение, чтобы оно воспринималось бинокулярно как единый предмет,
- 18. ФОТОРЕЦЕПЦИЯ
- 19. Строение сетчатки
- 20. Строение сетчатки А – сосудистая оболочка, В - пигментный эпителий, С – слой палочек и колбочек,
- 21. Строение фоторецепторов
- 22. В состав дисков фоторецепторных клеток входят зрительные пигменты, в том числе родопсин палочек. Родопсин состоит из
- 23. Ионные основы фоторецепторных потенциалов В темноте Na+‑каналы мембраны наружных сегментов палочек и колбочек открыты, и течёт
- 24. При попадании квантов света: 1. активация родопсина в результате фотоизомеризации, 2. каталитическая активация родопсином G‑белка (трансдуцин),
- 25. При попадании квантов света: 5. закрытие цГМФ–зависимых Na+‑каналов, 6. гиперполяризация (гиперполяризационный рецепторный потенциал), 7. потенциал распространяется
- 26. Возврат к исходному состоянию Свет, вызывающий понижение концентрации цГМФ и приводящий к закрытию Na+-каналов, уменьшает содержание
- 27. Световая адаптация Если человек длительное время находится в условиях яркого освещения: - в палочках и колбочках
- 28. Темновая адаптация Если человек продолжительное время остаётся в темноте: - витамин A вновь превращается в ретиналь,
- 29. Темновая адаптация При нахождении в темноте световая чувствительность палочек нарастает неравномерно: - в первые минуты она
- 30. Другие механизмы адаптации 1. Изменение размеров зрачка в течение долей секунды может в 30 раз уменьшить
- 31. Различные клетки сетчатки генерируют локальные потенциалы, но не ПД Из всех клеток сетчатки ПД возникают только
- 32. Особенности локальных потенциалов колбочек и палочек Рецепторные потенциалы колбочек и палочек возникают одинаково быстро, но скорость
- 33. Проведение сигналов от палочек и колбочек отличаются: Нейроны и аксоны ганглиозных клеток от колбочек, значительно толще,
- 34. ЦВЕТОВОЕ ЗРЕНИЕ
- 35. Теории цветового зрения: Теория Эмпедокла (V век до н.э) Любой предмет излучает некую «субстанцию»; ощущение цвета
- 36. Теория света и цвета Ньютона В 1672 году Ньютон разложил свет в спектр (белый цвет всегда
- 37. Гипотеза М. В. Ломоносова, XVIII в. (биофизическое восприятие цвета) Основные цвета: красный, зелёный, жёлтый из которых
- 38. Трехкомпонентная теория (предложил Т.Янг (1802) и развил Г. Гельмгольц) В сетчатке должны быть три вида колбочек,
- 39. Теория Геринга, 1870 год (оппонентная гипотеза, теория обратного процесса) Есть три системы рецепторов: красно-зеленые, желто-голубые и
- 40. Нарушение цветового восприятия Трихромазия (нормальное зрение) — возможность различать любые цвета — определяется присутствием в сетчатке
- 41. Дихромазии (цветовая слепота, или дальтонизм)— дефекты цветового восприятия (преимущественно у мужчин) по одному из первичных цветов
- 42. Передача цветовых сигналов Каждая ганглиозная клетка может стимулироваться как отдельными, так и многими колбочками. Когда все
- 43. Если ганглиозная клетка возбуждается колбочками только одного цвета, то она будет тормозиться возбуждением колбочки другого типа.
- 44. Зрительные пути подразделяют на: 1. старую систему, куда относятся средний мозг и основание переднего мозга, 2.
- 45. Основной путь к зрительной коре (новая система) Аксоны ганглиозных клеток в составе зрительных нервов и (после
- 46. Основной путь к зрительной коре (новая система) В левом ЛКТ (ипсилатеральном глазу) волокна от носовой половины
- 47. Зрительная кора Первичная зрительная воспринимающая область располагается на соответствующей стороне шпорной борозды. В первичной зрительной коре
- 48. Удаление первичной зрительной коры у человека вызывает потерю восприятия осознаваемых зрительных образов, то есть слепоту. Однако
- 50. Физиология слуха и равновесия
- 54. Полость преддверия, барабанная и вестибулярная лестницы улитки заполнены перилимфой, а находящиеся в перилимфе полукружные каналы, маточка,
- 55. Концентрация K+ в эндолимфе в 100 раз больше, чем в ликворе и перилимфе; Концентрация Na+ в
- 56. Перепончатый канал улитки заряжен положительно (60–80 мВ) относительно двух других лестниц. Источник эндокохлеарного потенциала — сосудистая
- 57. Последовательность сенсорного преобразования в органе слуха: Собственно рецепторами являются внутренние волосковые клетки. Наружные волосковые клетки, обладая
- 59. Сканирующая электронная микрофотография волосковых клеток кортиева органа Покровная мембрана удалена, а микроскоп направлен на базиллярную мембрану.
- 60. Волосковые клетки
- 64. Высота звука - это субъективное восприятие частоты звуковых колебаний. Человеческое ухо воспринимает частоты в диапазоне от
- 65. 2. Временным способом (20 – 200 Гц) Информация кодируется разной частотой импульсации в одних и тех
- 66. Острота слуха Абсолютный порог слуховой чувствительности — минимальная сила звука, которую слышит человек в 50% случаев
- 67. Сила звука Диапазон силы звука, воспринимаемый человеческим ухом, огромен (болевой порог в 1013 раз выше порога
- 68. Слуховые пути и центры кохлеарное ядро (нейроны 2-го порядка), комплекс ядер верхней оливы и трапецевидного тела
- 69. Направление источника звука определяется 2 путями: При звуке высокой частоты (звук отражается от головы) источник оценивается
- 70. Слуховая кора Первичная слуховая кора непосредственно получает сигналы от медиального коленчатого тела, в то время как
- 71. Чувство равновесия
- 73. Структуры вестибулярного аппарата
- 74. Полукружные каналы работают в парах (билатерально) При вращение головы вправо: Волосковые клетки с правой стороны гиперполяризуются
- 75. Определение углового ускорения Полукружные каналы Примерно ортогональны друг другу Оба конца заканчиваются в овальном мешочке (место
- 76. Определение линейного ускорения Отолитовые органы: Активный участок – макула – представлена волосковыми клетками, отолитовой мембраной и
- 78. Волосковые клетки При возбуждении в волосковых клетках генерируется рецепторный потенциал и происходит выброс ацетилхолина, который и
- 79. Гребень волосков каждого рецептора настроен на «свое» направление ускорения Темная точка - киноцилия. Какой рецептор из
- 80. Передача сигнала в ЦНС Сигнал от каждого рецептора топически («поточечно») передается в вестибулярные ядра, нейроны которых
- 81. Дальнейшая передача вестибулярной информации (прежде всего, для коррекции движений): в спинной мозг в мозжечок в средний
- 82. Вестибулярная информация необходима: Спинному мозгу (от ядра Дейтерса и медиального ядер по латеральному и медиальному вестибулоспинальным
- 84. Укачивание Укачивание - иррадиация возбуждения по центрам ромбовидной ямки. Субъективные ощущения: головокружение, тошнота и другие реакции,
- 85. Вестибулярная система как источник эмоций (новизна ощущений!)
- 86. Обонятельная сенсорная система
- 87. Классификация запахов по Эймуэру Первичные или основные: - камфорный - (камфора, 1,8-цинеол) - острый или едкий
- 88. Воспринимающие обонятельные структуры образуют в слизистой оболочке носа специализированную область — парную обонятельную выстилку (обонятельное поле),
- 89. Втягивание воздуха Область, содержащая обонятельные рецепторы, плохо вентилируется. Улучшение их вентиляции обеспечивается принюхиванием. Втягивание воздуха —
- 90. Рецепторный аппарат Обонятельный эпителий содержит опорные эпителиальные клетки и расположенные между ними примерно 100 млн рецепторных
- 91. От булавы отходит от 8 до 40 тонких обонятельных волосков — ресничек. В волоски встроены обонятельные
- 92. ОБОНЯТЕЛЬНАЯ ЛУКОВИЦА В обонятельной луковице аксоны рецепторных клеток образуют синапсы с дендритами митральных и пучковых клеток,
- 93. В следующем слое обонятельной луковицы дендриты митральных, зернистых и пучковых клеток образуют реципрокные синапсы. Эти синаптические
- 94. Регистрация обонятельного сигнала взаимодействие одоранта с рецептором в плазмолемме обонятельных волосков активация G‑белка повышение активности аденилатциклазы
- 95. Через цАМФ–зависимые воротные ионные каналы внутрь клетки проходят не только одновалентные катионы, но и Ca2+, связывающийся
- 96. Концепция комбинаторного кодирования Каждый из миллионов одорантов имеет уникальный код; кодирование происходит на уровне органа обоняния,
- 97. Пути передачи обонятельной информации Аксоны митральных клеток в составе обонятельного тракта поднимаются в первичные обонятельные центры
- 98. Пути передачи обонятельной информации Аксоны нервных клеток латеральной обонятельной области, расположенных в грушевидной коре и миндалевидном
- 100. Вкусовая сенсорная система
- 101. Основные вкусы Человек различает четыре первичных вкуса: сладкий, кислый, горький солёный, а также «умами» (от японского
- 102. Рецепторы вкуса — контактные, а пути проведения от них проходят через ствол мозга к таламусу и
- 103. Вкусовые области языка Вкус сладкого регистрируется преимущественно на кончике языка, солёного — ближе к кончику языка,
- 104. Вкусовая почка состоит из удлинённых светлых клеток различных типов. - На апикальной части микроворсинки, занимающие вкусовой
- 105. Различие вкусовых клеток При низких концентрациях деполяризуются и формируют рецепторный потенциал лишь отдельные вкусовые клетки. Некоторые
- 106. Восприятие соленого и кислого Соленый стимул взаимодействует с эпителиальными натриевыми каналами (ENaC), открывая их для натрия.
- 107. Восприятие сладкого Сладкий вкус возникает за счет взаимодействия сладкого стимула с чувствительным к нему рецептором, связанным
- 108. Восприятие горького Горький стимул может деполяризовать мембрану тремя путями: закрытием калиевых каналов, путем взаимодействия с G-белком
- 109. Восприятие умами Глютамат связывается с глютаматрегулируемыми неселективными ионными каналами и открывает их. Это сопровождается деполяризацией и
- 110. ВКУСОВОЙ ПОРОГ Наиболее значительные различия наблюдают между веществами, создающими ощущение горького и всех остальных первичных вкусов.
- 111. АДАПТАЦИЯ Адаптация к воздействию вкусового вещества развивается медленно (минуты) и пропорциональна его концентрации. К сладкому и
- 112. Восходящие пути вкусовой чувствительности состоят из трёх нейронов: 1. первичного чувствительного нейрона - от передних двух
- 113. Восходящие пути вкусовой чувствительности состоят из трёх нейронов: 2. нейроны ядра одиночного пути расположены в ростральной
- 114. Восходящие пути вкусовой чувствительности состоят из трёх нейронов: 3. Нейроны вентромедиального ядра таламуса направляются к вентральной
- 115. Тактильная сенсорная система
- 116. Тактильные ощущения прикосновения, давления и вибрации относятся к раздельным видам ощущений, но воспринимаются одними и теми
- 117. Рецепторы кожи Свободные нервные окончания образуют механорецепторы (прикосновение и давление), терморецепторы и рецепторы болевой чувствительности. Тельца
- 118. Количество и плотность размещения рецепторов - Общее количество тактильных рецепторов оценивается в 10 миллионов штук, сгруппированных
- 119. Разрешающая способность и пороги ощущений: - Порог ощущения кожных рецепторов при надавливании: 10 мкм - Одновременный
- 120. Адаптация
- 121. Передача импульсации Передача тактильных ощущений тонкой дифференцировки происходит по миелинизированным нервным волокнам со скоростью 30-70 м/с.
- 122. Пути передачи соматосенсорных сигналов Практически вся сенсорная информация от сегментов тела поступает в спинной мозг через
- 123. Войдя в спинной мозг они направляются: - либо прямо к продолговатому мозгу (лемнисковая система: тонкий, или
- 125. Скачать презентацию

Орган зрения включает:
1. глазное яблоко, соединённое через зрительный нерв с мозгом,
Орган зрения включает:
1. глазное яблоко, соединённое через зрительный нерв с мозгом,

Оптика глаза
Глаз имеет систему линз с различной кривизной и различными показателями
Оптика глаза
Глаз имеет систему линз с различной кривизной и различными показателями

Преломляющая сила
Для расчетов преломляющей силы глаза используют понятие о так называемом
Преломляющая сила
Для расчетов преломляющей силы глаза используют понятие о так называемом
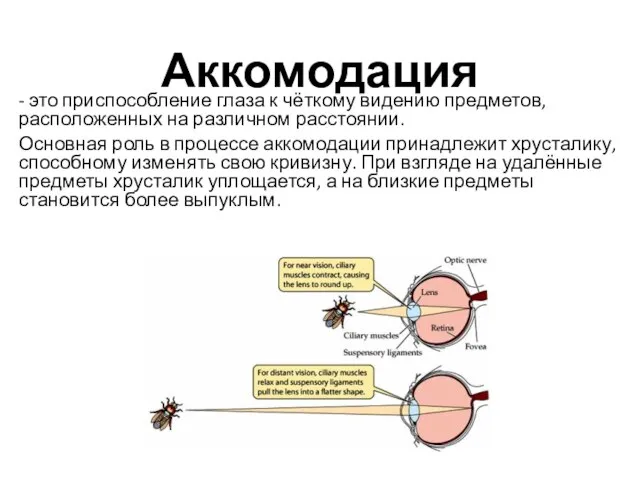
Аккомодация
- это приспособление глаза к чёткому видению предметов, расположенных на различном
Аккомодация
- это приспособление глаза к чёткому видению предметов, расположенных на различном

ПСНС обеспечивает фокусировку при приближении предмета к глазу.
СНС незначительно расслабляет ресничную
ПСНС обеспечивает фокусировку при приближении предмета к глазу.
СНС незначительно расслабляет ресничную

Зрачковый рефлекс
Просвет зрачка может изменяться от 1 мм до 8 мм. Это придаёт
Зрачковый рефлекс
Просвет зрачка может изменяться от 1 мм до 8 мм. Это придаёт

Регуляция зрачкового рефлекса:
1. ПСНС (ацетилхолин и эзерин) вызывают сужение зрачка, а
Регуляция зрачкового рефлекса:
1. ПСНС (ацетилхолин и эзерин) вызывают сужение зрачка, а
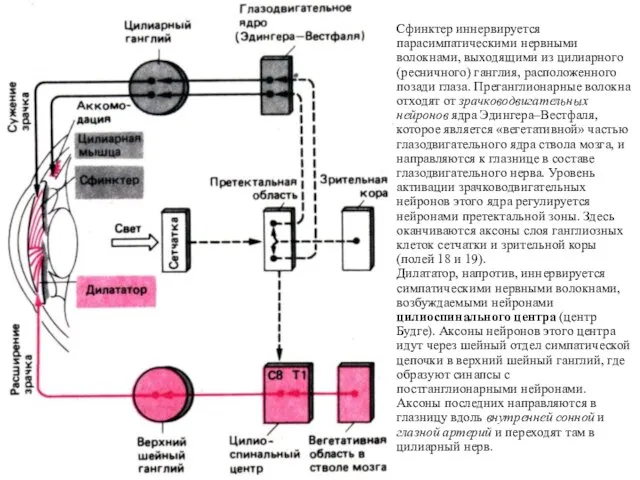
Сфинктер иннервируется парасимпатическими нервными волокнами, выходящими из цилиарного (ресничного) ганглия, расположенного
Сфинктер иннервируется парасимпатическими нервными волокнами, выходящими из цилиарного (ресничного) ганглия, расположенного

Зрачковый рефлекс меняется при заболеваниях ЦНС:
а. Торможение возникает при нарушении передачи
Зрачковый рефлекс меняется при заболеваниях ЦНС:
а. Торможение возникает при нарушении передачи

Содружественная реакция зрачков
У здоровых людей зрачки обоих глаз одинакового размера. Освещение
Содружественная реакция зрачков
У здоровых людей зрачки обоих глаз одинакового размера. Освещение
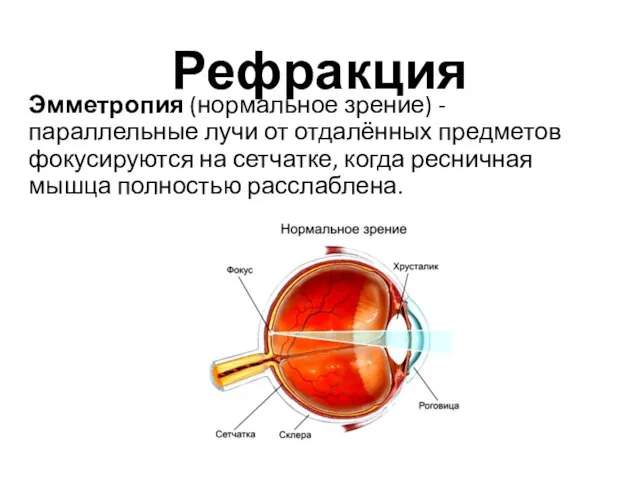
Рефракция
Эмметропия (нормальное зрение) - параллельные лучи от отдалённых предметов фокусируются на
Рефракция
Эмметропия (нормальное зрение) - параллельные лучи от отдалённых предметов фокусируются на
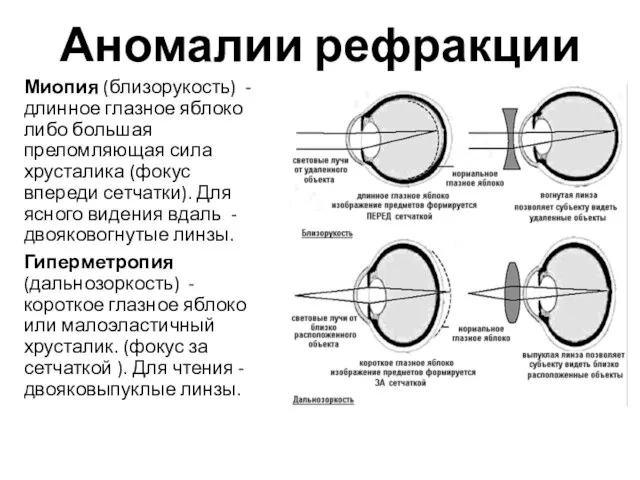
Аномалии рефракции
Миопия (близорукость) - длинное глазное яблоко либо большая преломляющая сила
Аномалии рефракции
Миопия (близорукость) - длинное глазное яблоко либо большая преломляющая сила
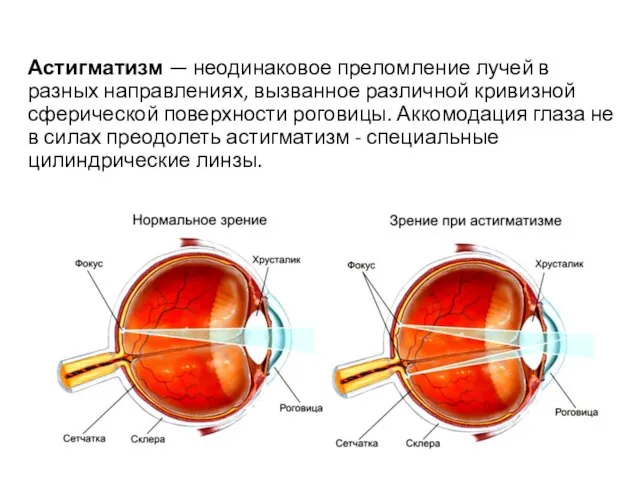
Астигматизм — неодинаковое преломление лучей в разных направлениях, вызванное различной кривизной сферической
Астигматизм — неодинаковое преломление лучей в разных направлениях, вызванное различной кривизной сферической

Поля зрения
Зрительное поле каждого глаза — часть внешнего пространства, видимого глазом
Поля зрения
Зрительное поле каждого глаза — часть внешнего пространства, видимого глазом

Бинокулярное зрение
Центральная часть зрительных полей двух глаз полностью совпадает; следовательно, любой
Бинокулярное зрение
Центральная часть зрительных полей двух глаз полностью совпадает; следовательно, любой
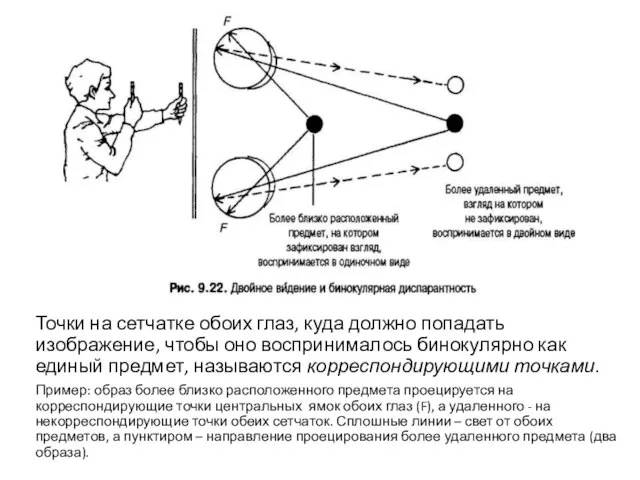
Точки на сетчатке обоих глаз, куда должно попадать изображение, чтобы оно
Точки на сетчатке обоих глаз, куда должно попадать изображение, чтобы оно

ФОТОРЕЦЕПЦИЯ
ФОТОРЕЦЕПЦИЯ
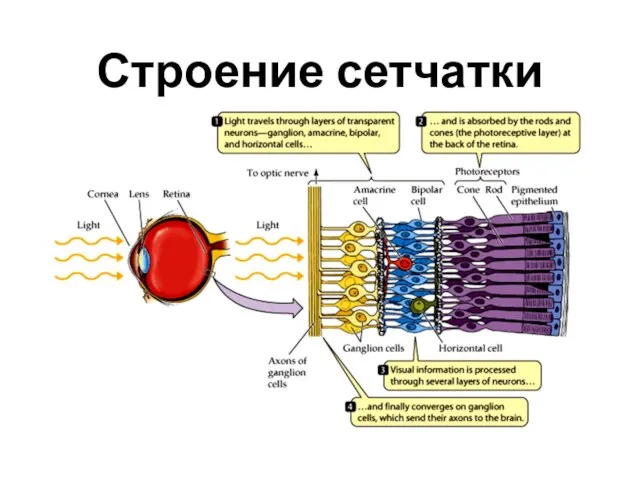
Строение сетчатки
Строение сетчатки
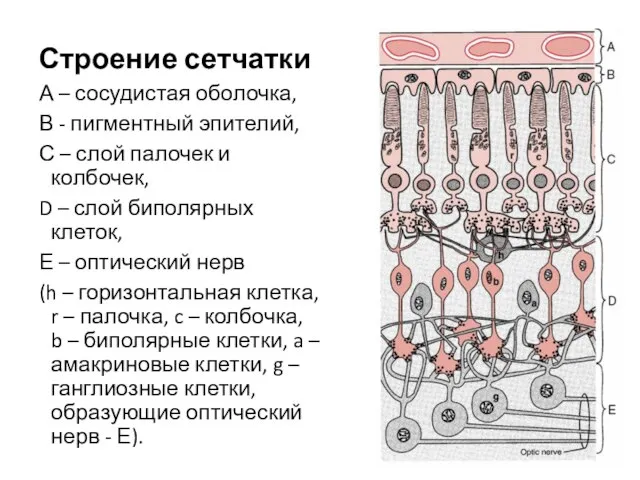
Строение сетчатки
А – сосудистая оболочка,
В - пигментный эпителий,
С –
Строение сетчатки
А – сосудистая оболочка,
В - пигментный эпителий,
С –
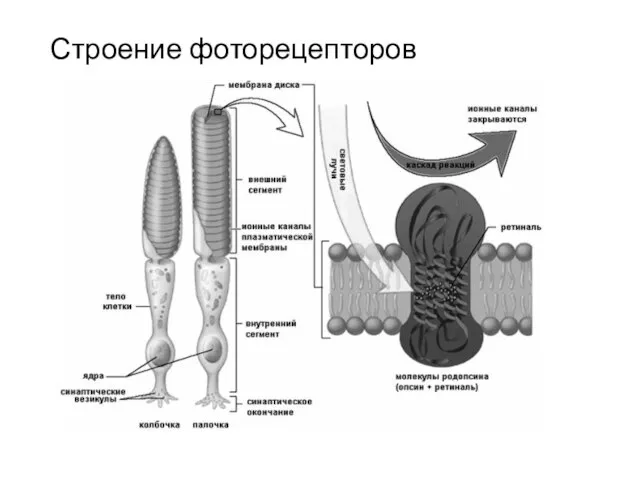
Строение фоторецепторов
Строение фоторецепторов
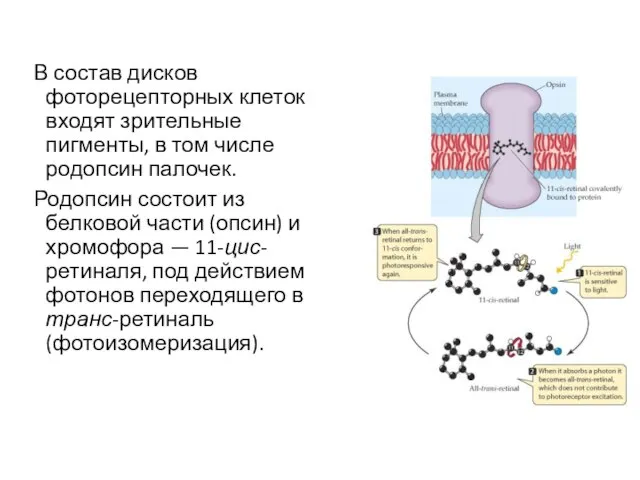
В состав дисков фоторецепторных клеток входят зрительные пигменты, в том числе
В состав дисков фоторецепторных клеток входят зрительные пигменты, в том числе
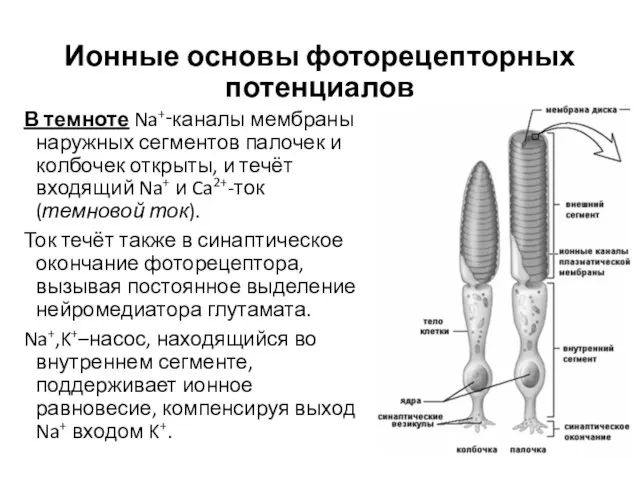
Ионные основы фоторецепторных потенциалов
В темноте Na+‑каналы мембраны наружных сегментов палочек и
Ионные основы фоторецепторных потенциалов
В темноте Na+‑каналы мембраны наружных сегментов палочек и
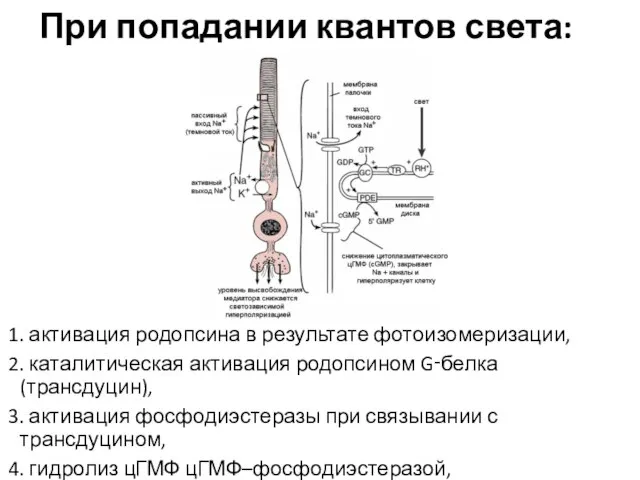
При попадании квантов света:
1. активация родопсина в результате фотоизомеризации,
2.
При попадании квантов света:
1. активация родопсина в результате фотоизомеризации,
2.
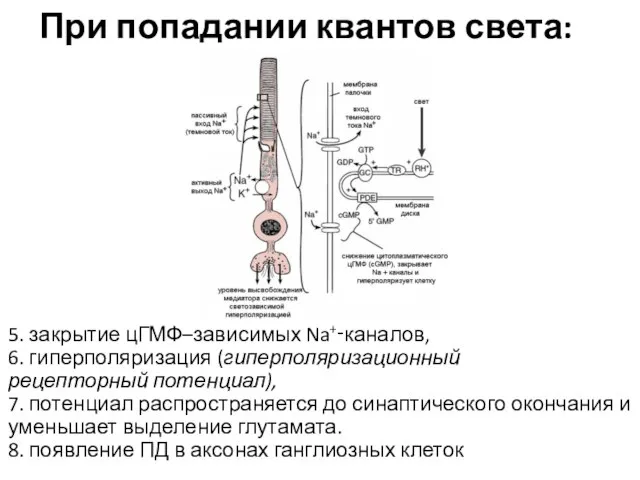
При попадании квантов света:
5. закрытие цГМФ–зависимых Na+‑каналов,
6. гиперполяризация (гиперполяризационный
рецепторный
При попадании квантов света:
5. закрытие цГМФ–зависимых Na+‑каналов,
6. гиперполяризация (гиперполяризационный
рецепторный

Возврат к исходному состоянию
Свет, вызывающий понижение концентрации цГМФ и приводящий к
Возврат к исходному состоянию
Свет, вызывающий понижение концентрации цГМФ и приводящий к

Световая адаптация
Если человек длительное время находится в условиях яркого освещения:
- в
Световая адаптация
Если человек длительное время находится в условиях яркого освещения:
- в

Темновая адаптация
Если человек продолжительное время остаётся в темноте:
- витамин A вновь
Темновая адаптация
Если человек продолжительное время остаётся в темноте:
- витамин A вновь

Темновая адаптация
При нахождении в темноте световая чувствительность палочек нарастает неравномерно:
-
Темновая адаптация
При нахождении в темноте световая чувствительность палочек нарастает неравномерно:
-

Другие механизмы адаптации
1. Изменение размеров зрачка в течение долей секунды может
Другие механизмы адаптации
1. Изменение размеров зрачка в течение долей секунды может

Различные клетки сетчатки генерируют локальные потенциалы, но не ПД
Из всех клеток
Различные клетки сетчатки генерируют локальные потенциалы, но не ПД
Из всех клеток

Особенности локальных потенциалов колбочек и палочек
Рецепторные потенциалы колбочек и палочек возникают
Особенности локальных потенциалов колбочек и палочек
Рецепторные потенциалы колбочек и палочек возникают

Проведение сигналов от палочек и колбочек отличаются:
Нейроны и аксоны ганглиозных клеток
Проведение сигналов от палочек и колбочек отличаются:
Нейроны и аксоны ганглиозных клеток

ЦВЕТОВОЕ ЗРЕНИЕ
ЦВЕТОВОЕ ЗРЕНИЕ

Теории цветового зрения:
Теория Эмпедокла (V век до н.э)
Любой предмет излучает некую
Теории цветового зрения:
Теория Эмпедокла (V век до н.э)
Любой предмет излучает некую

Теория света и цвета Ньютона
В 1672 году Ньютон разложил свет
Теория света и цвета Ньютона
В 1672 году Ньютон разложил свет

Гипотеза М. В. Ломоносова, XVIII в.
(биофизическое восприятие цвета)
Основные цвета: красный, зелёный, жёлтый из
Гипотеза М. В. Ломоносова, XVIII в.
(биофизическое восприятие цвета)
Основные цвета: красный, зелёный, жёлтый из
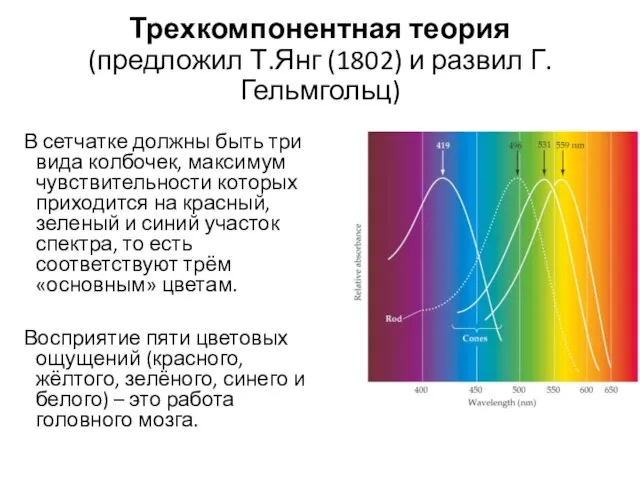
Трехкомпонентная теория
(предложил Т.Янг (1802) и развил Г. Гельмгольц)
В сетчатке должны
Трехкомпонентная теория
(предложил Т.Янг (1802) и развил Г. Гельмгольц)
В сетчатке должны

Теория Геринга, 1870 год
(оппонентная гипотеза, теория обратного процесса)
Есть три системы рецепторов:
Теория Геринга, 1870 год
(оппонентная гипотеза, теория обратного процесса)
Есть три системы рецепторов:

Нарушение цветового восприятия
Трихромазия (нормальное зрение) — возможность различать любые цвета — определяется присутствием
Нарушение цветового восприятия
Трихромазия (нормальное зрение) — возможность различать любые цвета — определяется присутствием

Дихромазии (цветовая слепота, или дальтонизм)— дефекты цветового восприятия (преимущественно у мужчин)

Передача цветовых сигналов
Каждая ганглиозная клетка может стимулироваться как отдельными, так и
Передача цветовых сигналов
Каждая ганглиозная клетка может стимулироваться как отдельными, так и

Если ганглиозная клетка возбуждается колбочками только одного цвета, то она будет
Если ганглиозная клетка возбуждается колбочками только одного цвета, то она будет

Зрительные пути подразделяют на:
1. старую систему, куда относятся средний мозг и
Зрительные пути подразделяют на:
1. старую систему, куда относятся средний мозг и
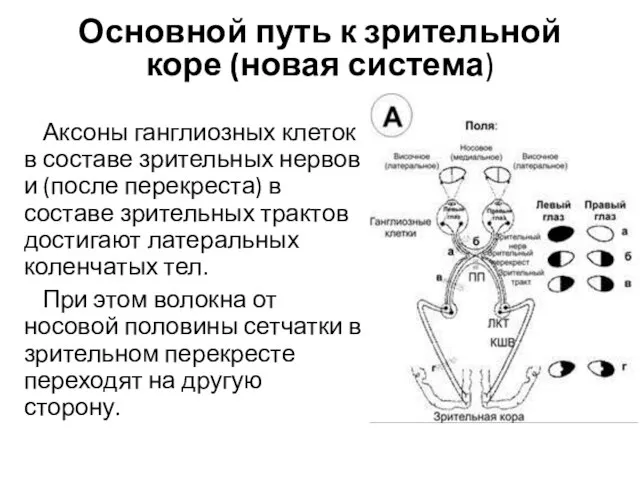
Основной путь к зрительной коре (новая система)
Аксоны ганглиозных клеток в составе
Основной путь к зрительной коре (новая система)
Аксоны ганглиозных клеток в составе
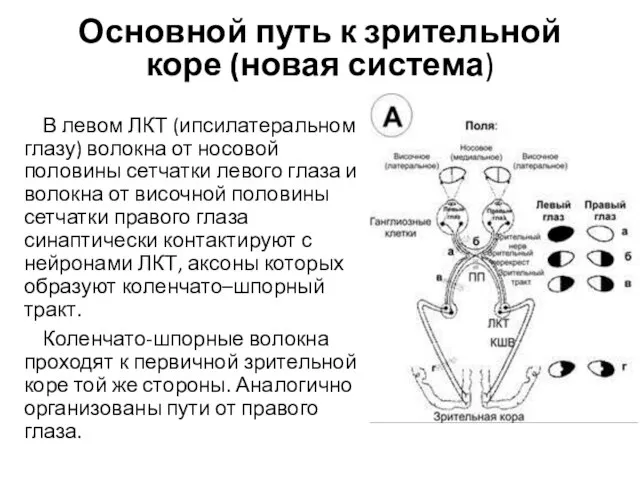
Основной путь к зрительной коре (новая система)
В левом ЛКТ (ипсилатеральном глазу)
Основной путь к зрительной коре (новая система)
В левом ЛКТ (ипсилатеральном глазу)

Зрительная кора
Первичная зрительная воспринимающая область располагается на соответствующей стороне шпорной борозды.
Зрительная кора
Первичная зрительная воспринимающая область располагается на соответствующей стороне шпорной борозды.

Удаление первичной зрительной коры
у человека вызывает потерю восприятия осознаваемых зрительных образов,
Удаление первичной зрительной коры
у человека вызывает потерю восприятия осознаваемых зрительных образов,
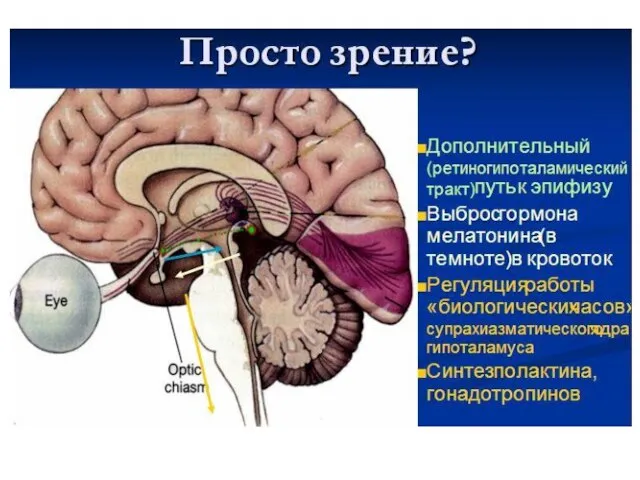

Физиология
слуха и равновесия
Физиология
слуха и равновесия
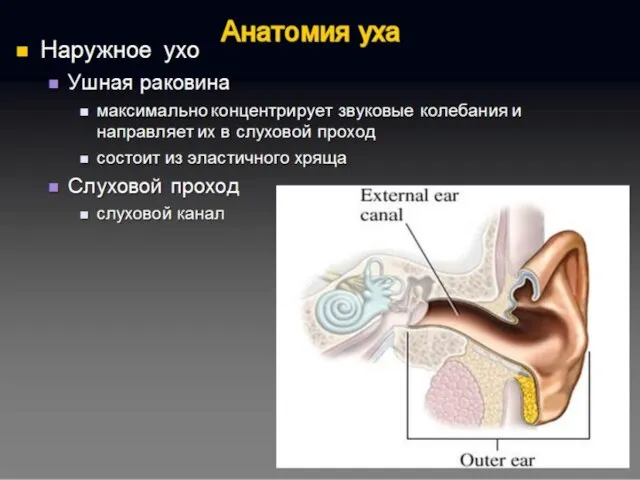
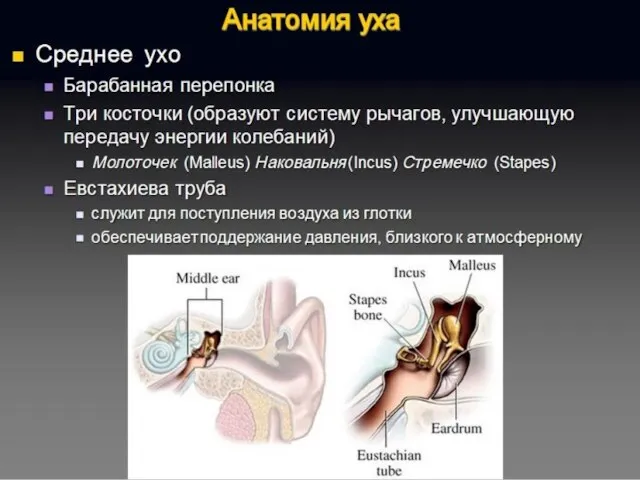
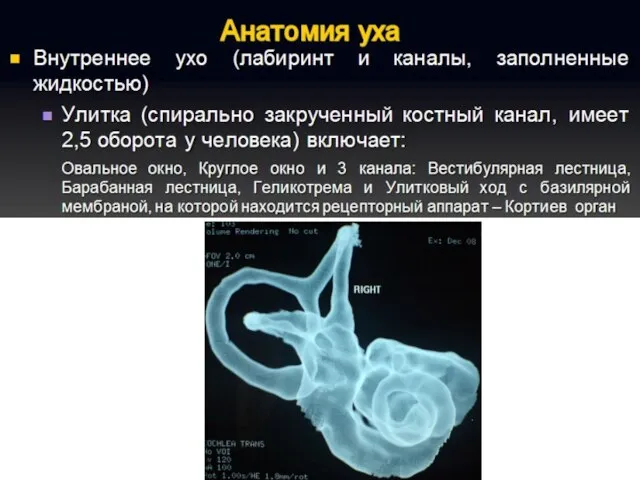
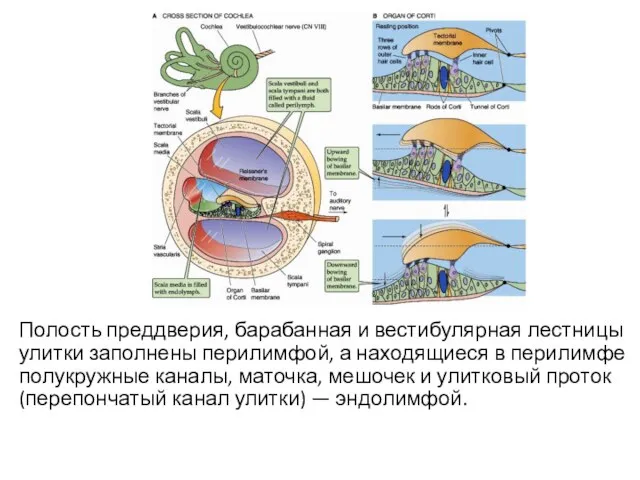
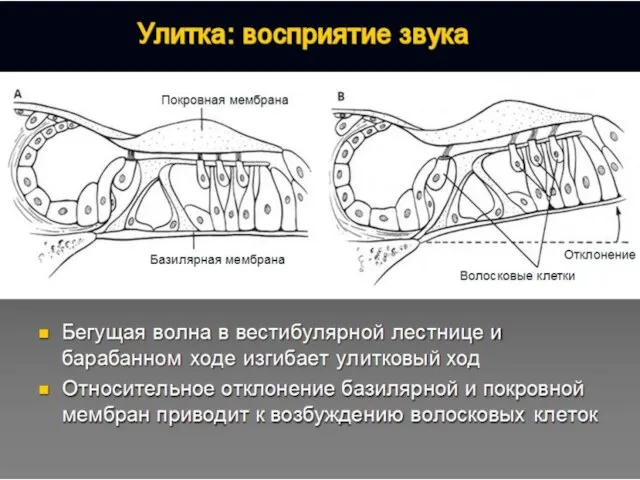
Полость преддверия, барабанная и вестибулярная лестницы улитки заполнены перилимфой, а находящиеся
Полость преддверия, барабанная и вестибулярная лестницы улитки заполнены перилимфой, а находящиеся
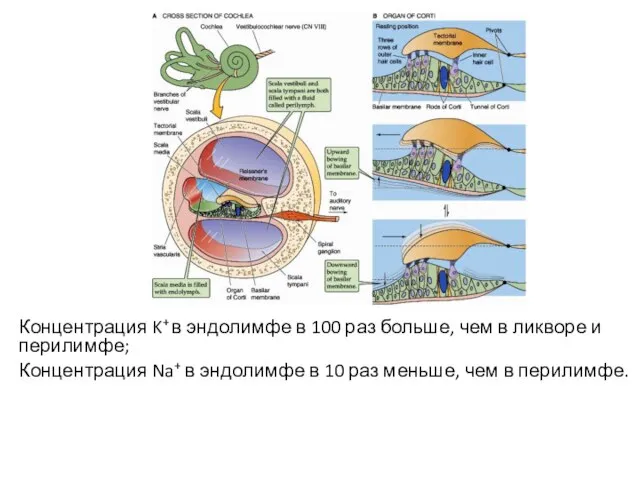
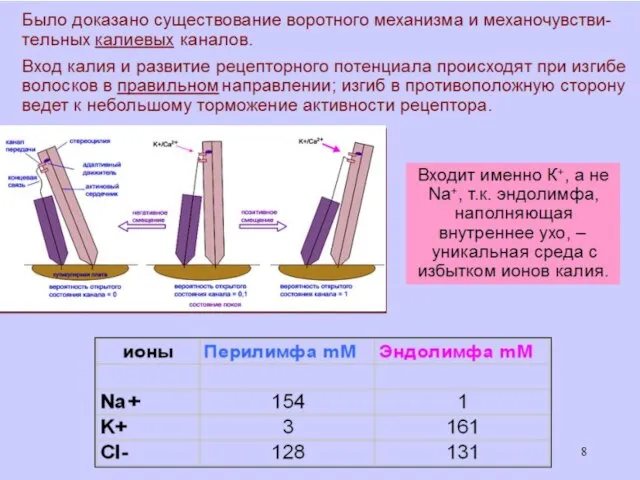
Концентрация K+ в эндолимфе в 100 раз больше, чем в ликворе
Концентрация K+ в эндолимфе в 100 раз больше, чем в ликворе
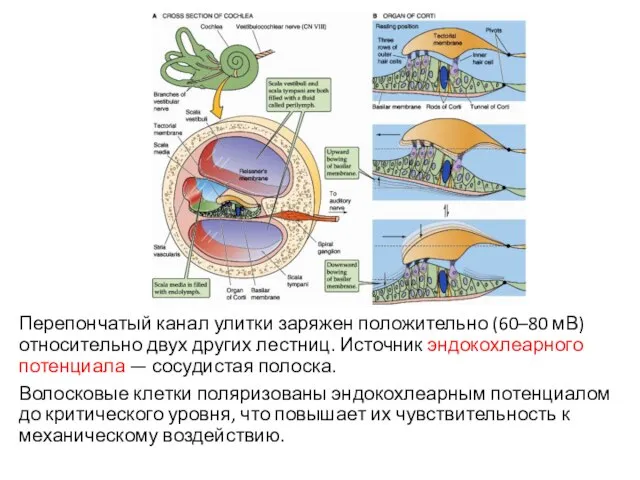
Перепончатый канал улитки заряжен положительно (60–80 мВ) относительно двух других лестниц.
Перепончатый канал улитки заряжен положительно (60–80 мВ) относительно двух других лестниц.
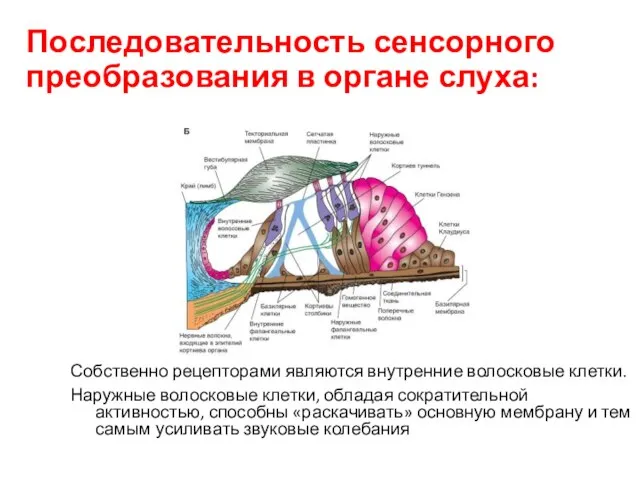
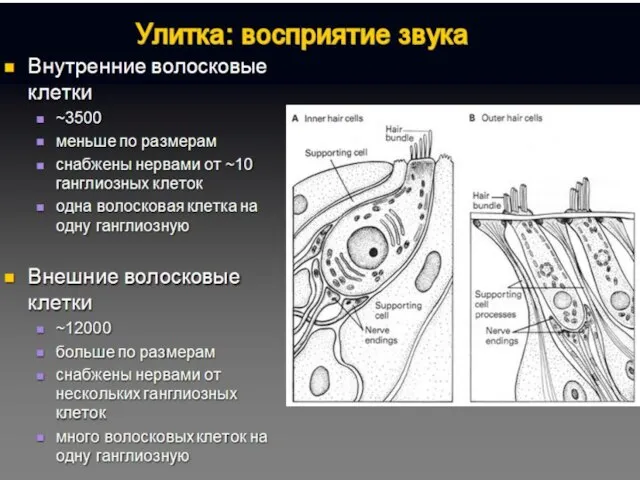
Последовательность сенсорного преобразования в органе слуха:
Собственно рецепторами являются внутренние волосковые клетки.
Наружные волосковые
Последовательность сенсорного преобразования в органе слуха:
Собственно рецепторами являются внутренние волосковые клетки.
Наружные волосковые
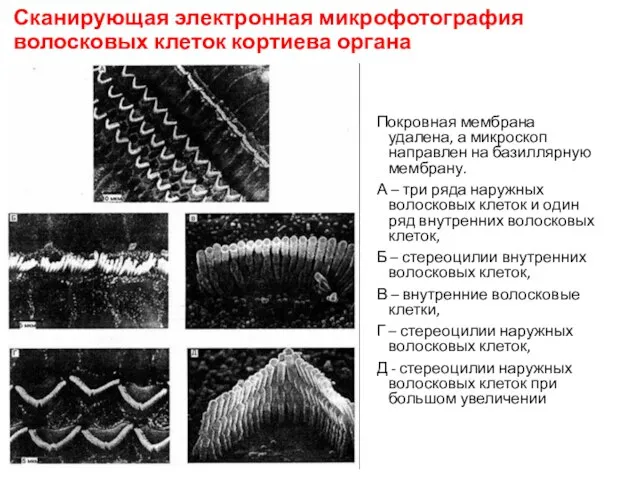
Сканирующая электронная микрофотография волосковых клеток кортиева органа
Покровная мембрана удалена, а микроскоп
Сканирующая электронная микрофотография волосковых клеток кортиева органа
Покровная мембрана удалена, а микроскоп

Волосковые клетки
Волосковые клетки
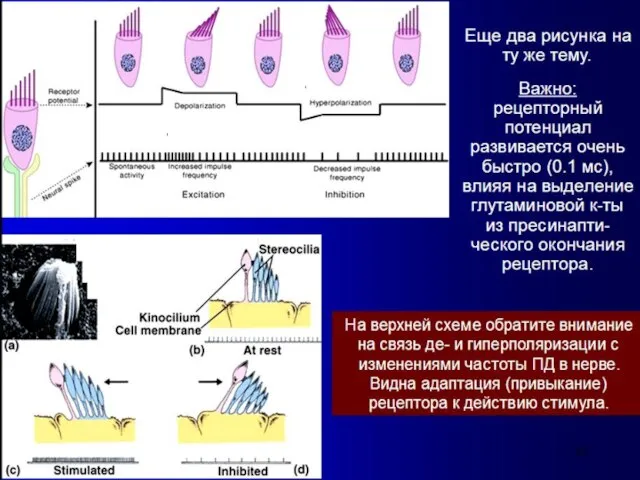
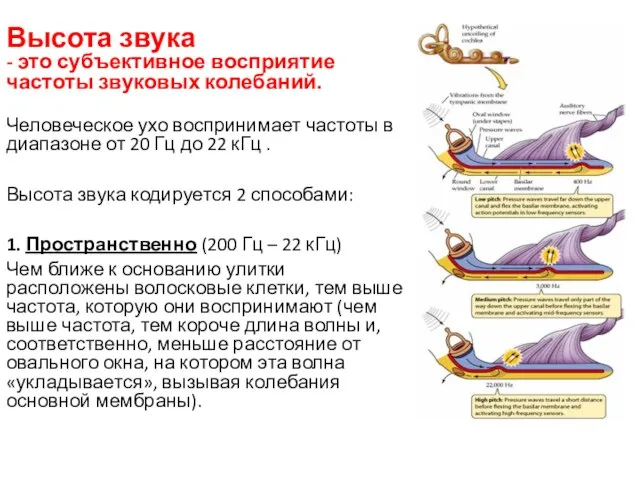
Высота звука
- это субъективное восприятие частоты звуковых колебаний.
Человеческое ухо
Высота звука
- это субъективное восприятие частоты звуковых колебаний.
Человеческое ухо
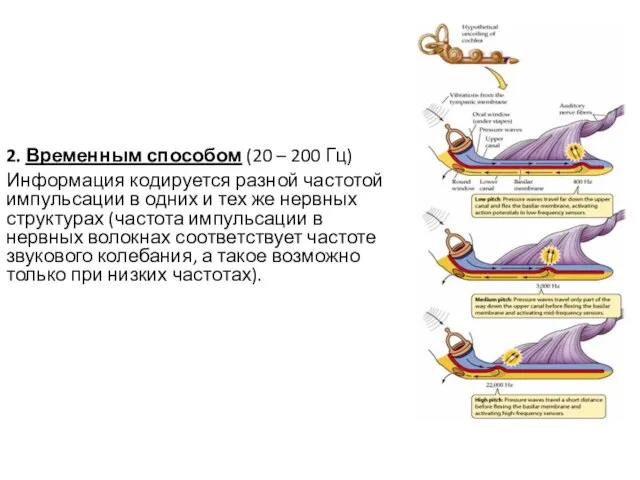
2. Временным способом (20 – 200 Гц)
Информация кодируется разной частотой импульсации в
2. Временным способом (20 – 200 Гц)
Информация кодируется разной частотой импульсации в

Острота слуха
Абсолютный порог слуховой чувствительности — минимальная сила звука, которую слышит человек
Острота слуха
Абсолютный порог слуховой чувствительности — минимальная сила звука, которую слышит человек

Сила звука
Диапазон силы звука, воспринимаемый человеческим ухом, огромен (болевой порог в 1013 раз
Сила звука
Диапазон силы звука, воспринимаемый человеческим ухом, огромен (болевой порог в 1013 раз
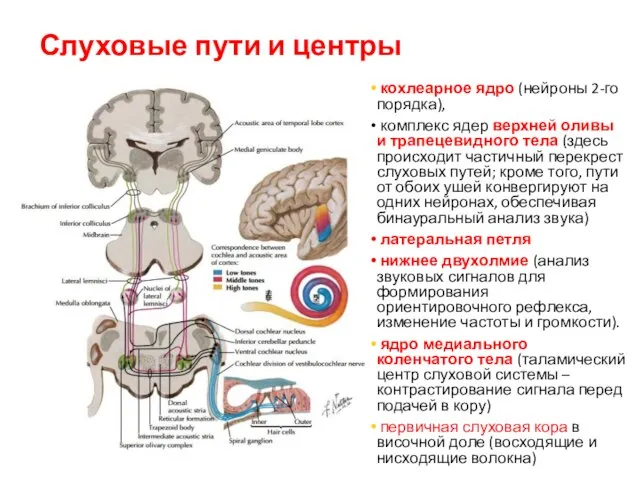
Слуховые пути и центры
кохлеарное ядро (нейроны 2-го порядка),
комплекс
Слуховые пути и центры
кохлеарное ядро (нейроны 2-го порядка),
комплекс

Направление источника звука определяется 2 путями:
При звуке высокой частоты (звук отражается
Направление источника звука определяется 2 путями:
При звуке высокой частоты (звук отражается
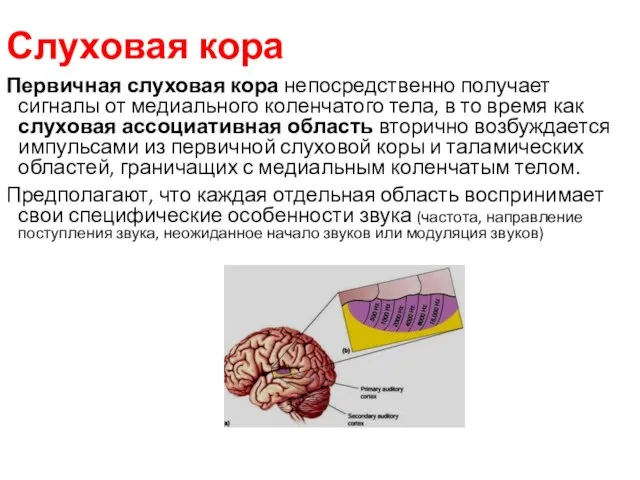
Слуховая кора
Первичная слуховая кора непосредственно получает сигналы от медиального коленчатого тела,
Слуховая кора
Первичная слуховая кора непосредственно получает сигналы от медиального коленчатого тела,

Чувство равновесия
Чувство равновесия

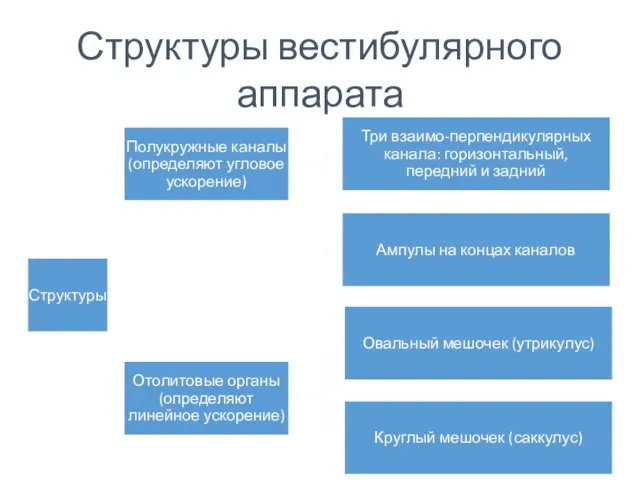
Структуры вестибулярного аппарата
Структуры вестибулярного аппарата

Полукружные каналы работают в парах (билатерально)
При вращение головы вправо:
Волосковые клетки с
Полукружные каналы работают в парах (билатерально)
При вращение головы вправо:
Волосковые клетки с

Определение углового ускорения
Полукружные каналы
Примерно ортогональны друг другу
Оба конца заканчиваются в овальном
Определение углового ускорения
Полукружные каналы
Примерно ортогональны друг другу
Оба конца заканчиваются в овальном
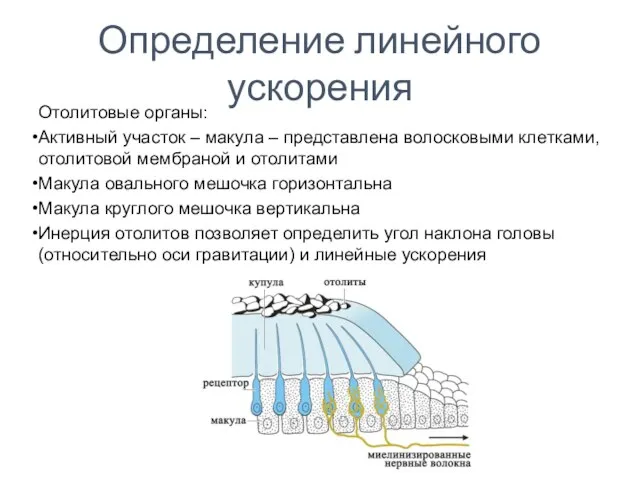
Определение линейного ускорения
Отолитовые органы:
Активный участок – макула – представлена волосковыми клетками,
Определение линейного ускорения
Отолитовые органы:
Активный участок – макула – представлена волосковыми клетками,
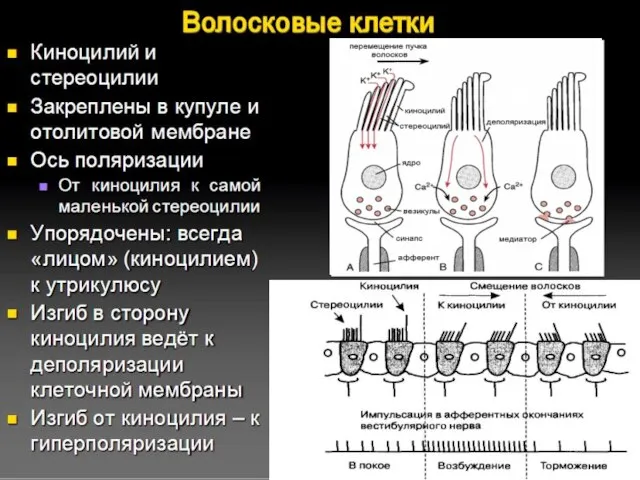
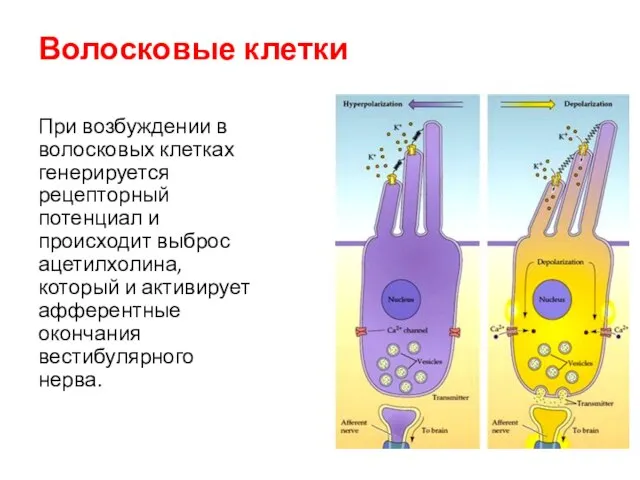
Волосковые клетки
При возбуждении в волосковых клетках генерируется рецепторный потенциал и происходит
Волосковые клетки
При возбуждении в волосковых клетках генерируется рецепторный потенциал и происходит
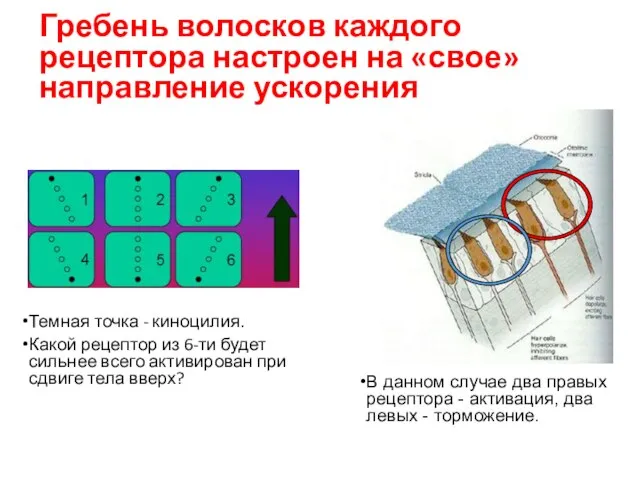
Гребень волосков каждого рецептора настроен на «свое» направление ускорения
Темная точка -
Гребень волосков каждого рецептора настроен на «свое» направление ускорения
Темная точка -
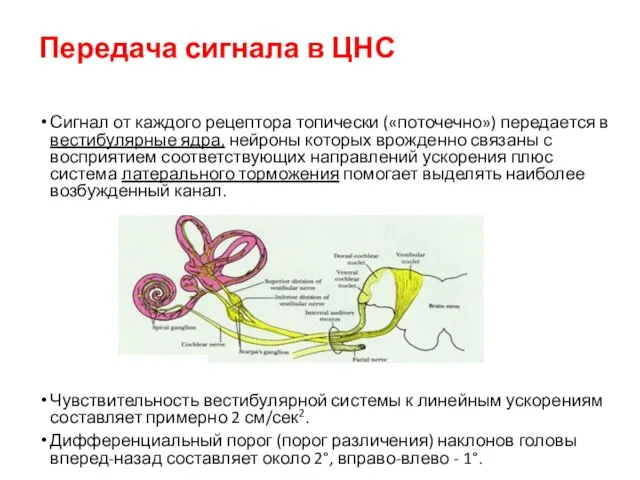
Передача сигнала в ЦНС
Сигнал от каждого рецептора топически («поточечно») передается в
Передача сигнала в ЦНС
Сигнал от каждого рецептора топически («поточечно») передается в
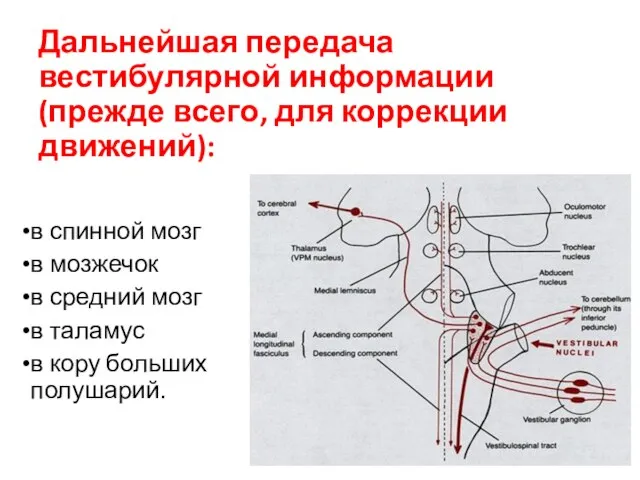
Дальнейшая передача вестибулярной информации (прежде всего, для коррекции движений):
в спинной мозг
Дальнейшая передача вестибулярной информации (прежде всего, для коррекции движений):
в спинной мозг

Вестибулярная информация необходима:
Спинному мозгу (от ядра Дейтерса и медиального ядер по
Вестибулярная информация необходима:
Спинному мозгу (от ядра Дейтерса и медиального ядер по
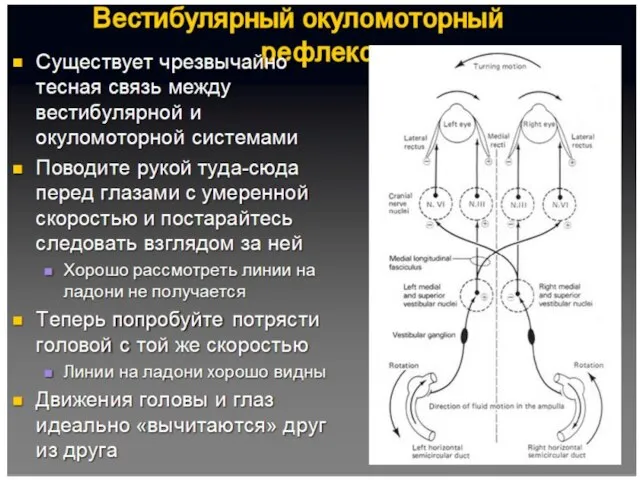
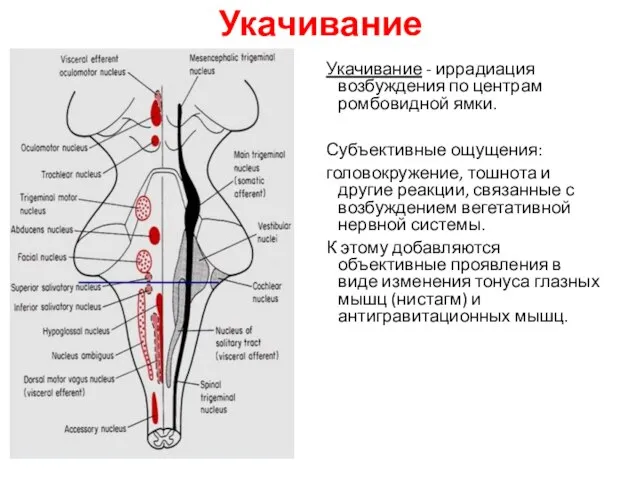
Укачивание
Укачивание - иррадиация возбуждения по центрам ромбовидной ямки.
Субъективные ощущения:
головокружение, тошнота и
Укачивание
Укачивание - иррадиация возбуждения по центрам ромбовидной ямки.
Субъективные ощущения:
головокружение, тошнота и

Вестибулярная система как источник эмоций (новизна ощущений!)
Вестибулярная система как источник эмоций (новизна ощущений!)

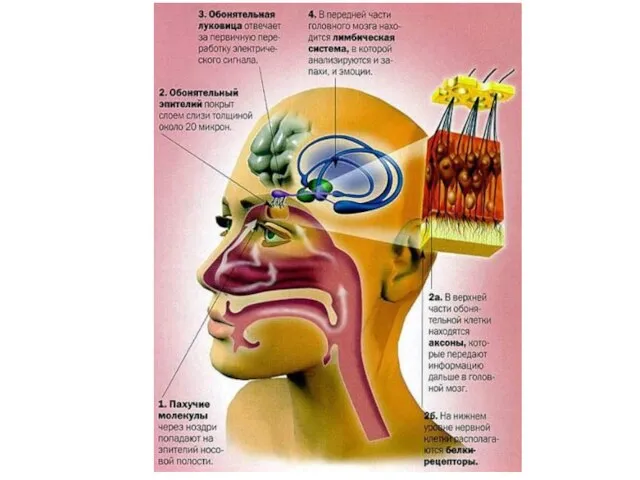
Обонятельная сенсорная система
Обонятельная сенсорная система

Классификация запахов по Эймуэру
Первичные или основные:
- камфорный - (камфора, 1,8-цинеол)
Классификация запахов по Эймуэру
Первичные или основные:
- камфорный - (камфора, 1,8-цинеол)
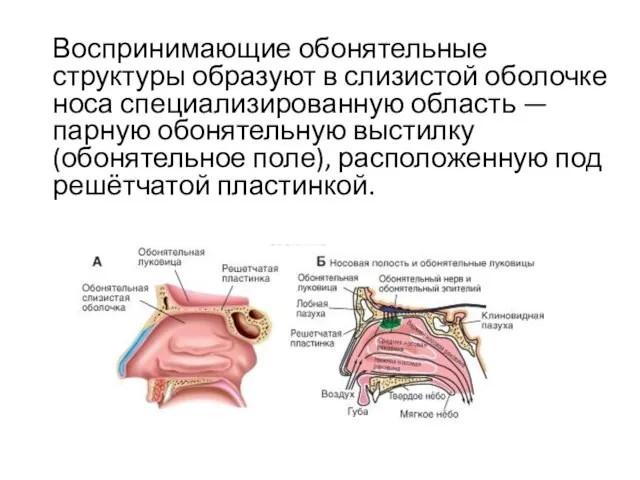
Воспринимающие обонятельные структуры образуют в слизистой оболочке носа специализированную область — парную
Воспринимающие обонятельные структуры образуют в слизистой оболочке носа специализированную область — парную

Втягивание воздуха
Область, содержащая обонятельные рецепторы, плохо вентилируется. Улучшение их вентиляции обеспечивается
Втягивание воздуха
Область, содержащая обонятельные рецепторы, плохо вентилируется. Улучшение их вентиляции обеспечивается
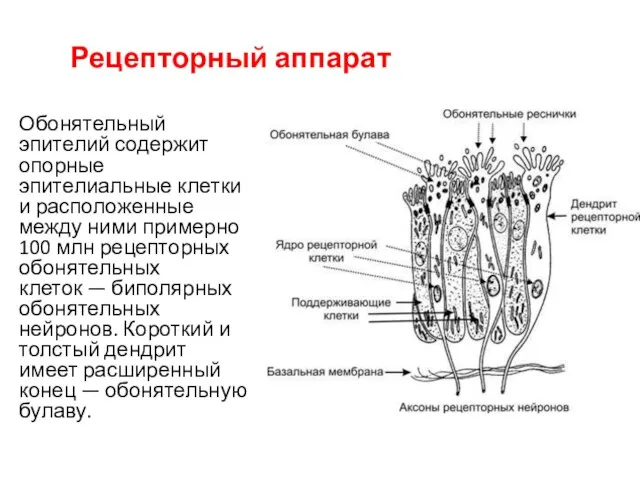
Рецепторный аппарат
Обонятельный эпителий содержит опорные эпителиальные клетки и расположенные между ними
Рецепторный аппарат
Обонятельный эпителий содержит опорные эпителиальные клетки и расположенные между ними
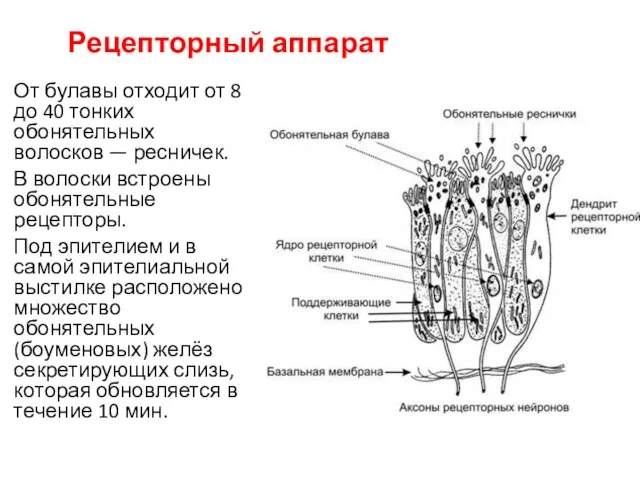
От булавы отходит от 8 до 40 тонких обонятельных волосков — ресничек.
От булавы отходит от 8 до 40 тонких обонятельных волосков — ресничек.
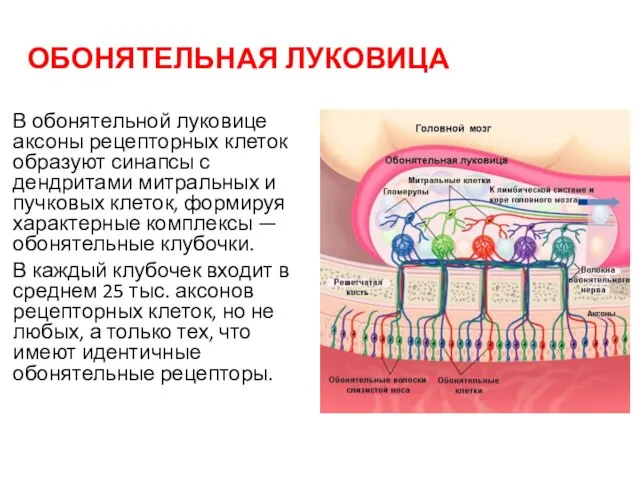
ОБОНЯТЕЛЬНАЯ ЛУКОВИЦА
В обонятельной луковице аксоны рецепторных клеток образуют синапсы с дендритами
ОБОНЯТЕЛЬНАЯ ЛУКОВИЦА
В обонятельной луковице аксоны рецепторных клеток образуют синапсы с дендритами
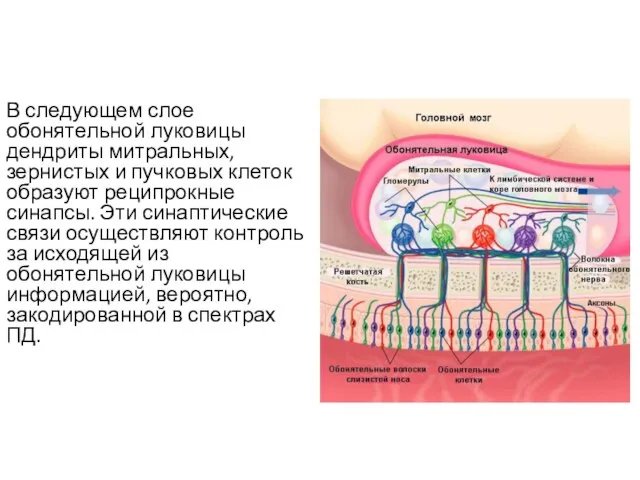
В следующем слое обонятельной луковицы дендриты митральных, зернистых и пучковых клеток
В следующем слое обонятельной луковицы дендриты митральных, зернистых и пучковых клеток
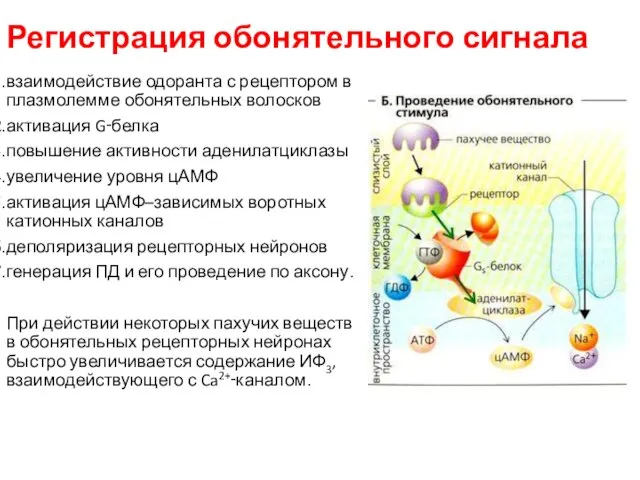
Регистрация обонятельного сигнала
взаимодействие одоранта с рецептором в плазмолемме обонятельных волосков
активация
Регистрация обонятельного сигнала
взаимодействие одоранта с рецептором в плазмолемме обонятельных волосков
активация

Через цАМФ–зависимые воротные ионные каналы внутрь клетки проходят не только одновалентные
Через цАМФ–зависимые воротные ионные каналы внутрь клетки проходят не только одновалентные

Концепция комбинаторного кодирования
Каждый из миллионов одорантов имеет уникальный код; кодирование происходит
Концепция комбинаторного кодирования
Каждый из миллионов одорантов имеет уникальный код; кодирование происходит
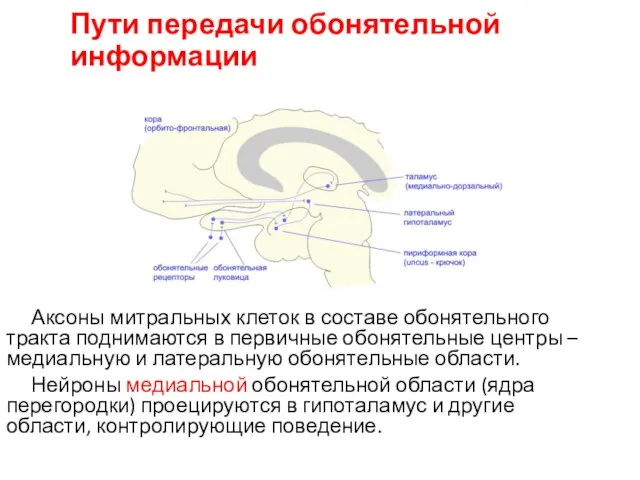
Пути передачи обонятельной информации
Аксоны митральных клеток в составе обонятельного тракта поднимаются
Пути передачи обонятельной информации
Аксоны митральных клеток в составе обонятельного тракта поднимаются
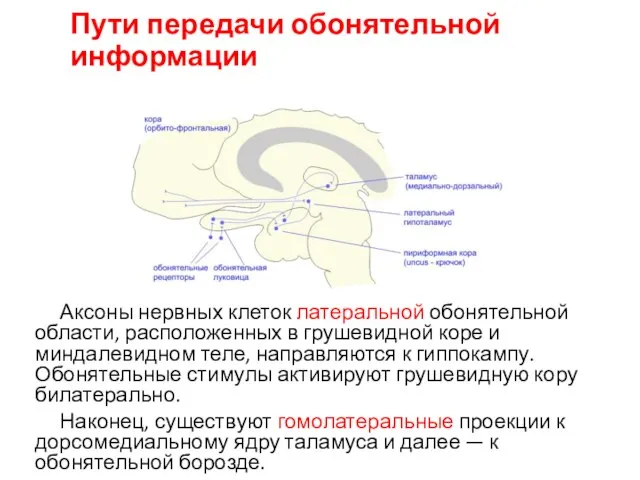
Пути передачи обонятельной информации
Аксоны нервных клеток латеральной обонятельной области, расположенных в
Пути передачи обонятельной информации
Аксоны нервных клеток латеральной обонятельной области, расположенных в
Вкусовая сенсорная система
Вкусовая сенсорная система

Основные вкусы
Человек различает четыре первичных вкуса:
сладкий,
кислый,
горький
солёный,
а также
Основные вкусы
Человек различает четыре первичных вкуса:
сладкий,
кислый,
горький
солёный,
а также
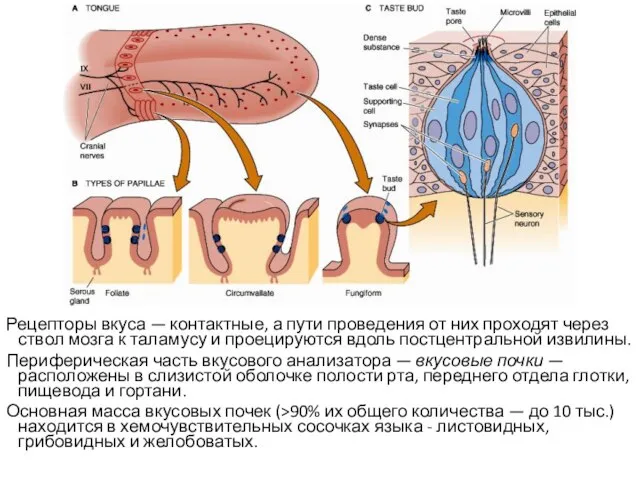
Рецепторы вкуса — контактные, а пути проведения от них проходят через ствол
Рецепторы вкуса — контактные, а пути проведения от них проходят через ствол
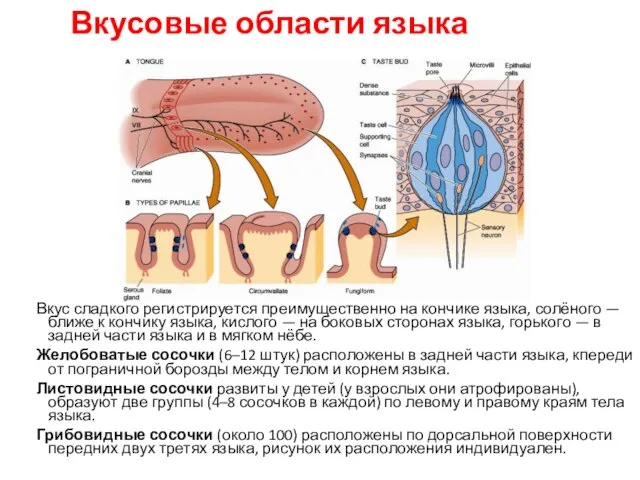
Вкусовые области языка
Вкус сладкого регистрируется преимущественно на кончике языка, солёного — ближе
Вкусовые области языка
Вкус сладкого регистрируется преимущественно на кончике языка, солёного — ближе
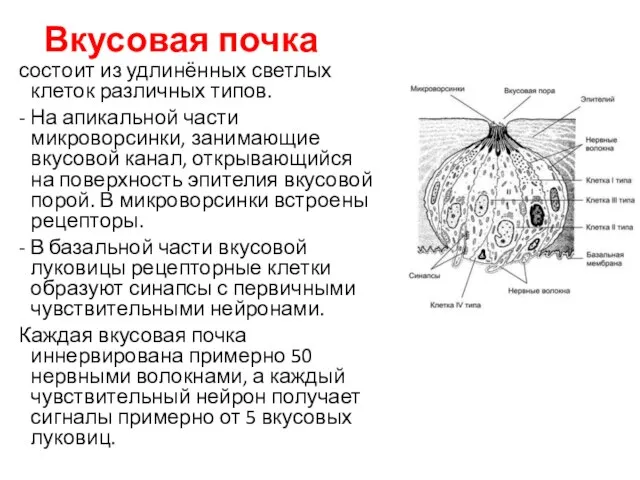
Вкусовая почка
состоит из удлинённых светлых клеток различных типов.
- На апикальной
Вкусовая почка
состоит из удлинённых светлых клеток различных типов.
- На апикальной

Различие вкусовых клеток
При низких концентрациях деполяризуются и формируют рецепторный потенциал лишь
Различие вкусовых клеток
При низких концентрациях деполяризуются и формируют рецепторный потенциал лишь
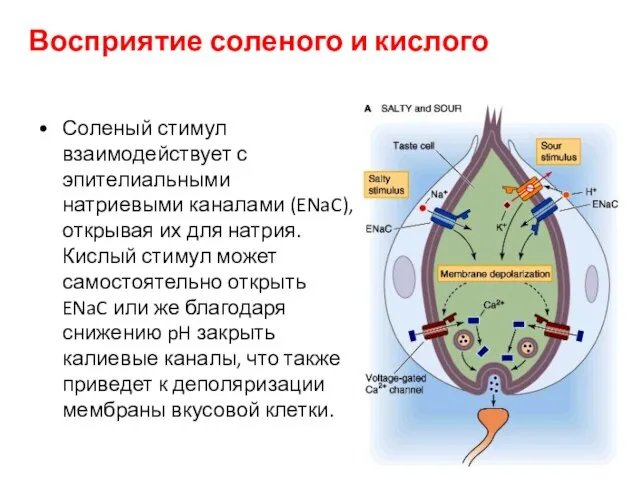
Восприятие соленого и кислого
Соленый стимул взаимодействует с эпителиальными натриевыми каналами (ENaC),
Восприятие соленого и кислого
Соленый стимул взаимодействует с эпителиальными натриевыми каналами (ENaC),
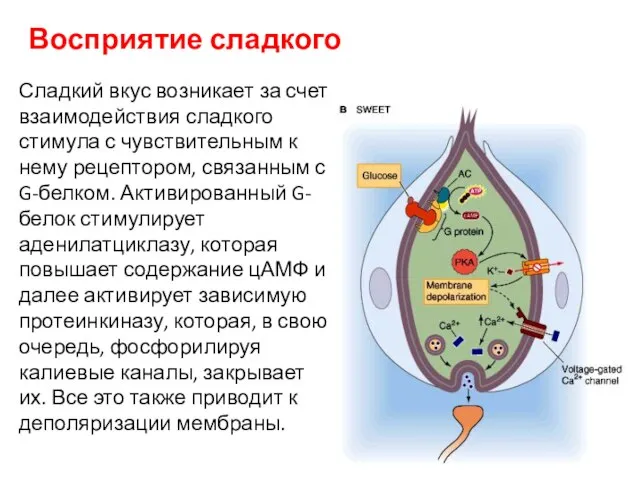
Восприятие сладкого
Сладкий вкус возникает за счет взаимодействия сладкого стимула с чувствительным
Восприятие сладкого
Сладкий вкус возникает за счет взаимодействия сладкого стимула с чувствительным
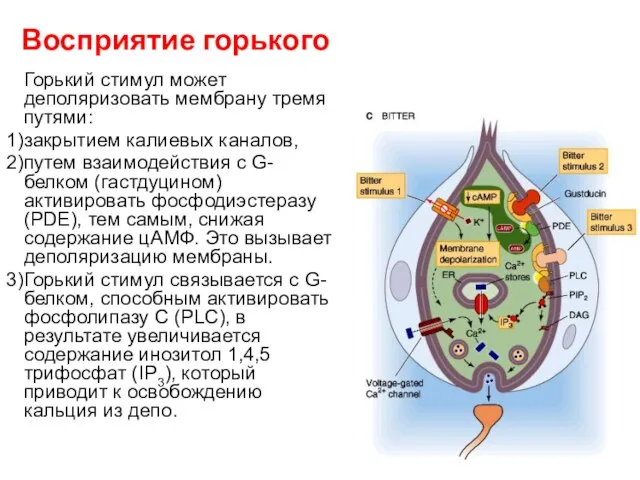
Восприятие горького
Горький стимул может деполяризовать мембрану тремя путями:
закрытием калиевых каналов,
Восприятие горького
Горький стимул может деполяризовать мембрану тремя путями:
закрытием калиевых каналов,
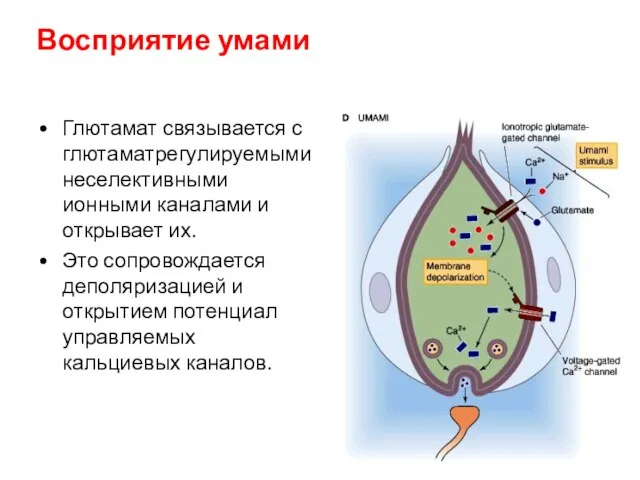
Восприятие умами
Глютамат связывается с глютаматрегулируемыми неселективными ионными каналами и открывает их.
Восприятие умами
Глютамат связывается с глютаматрегулируемыми неселективными ионными каналами и открывает их.

ВКУСОВОЙ ПОРОГ
Наиболее значительные различия наблюдают между веществами, создающими ощущение горького и
ВКУСОВОЙ ПОРОГ
Наиболее значительные различия наблюдают между веществами, создающими ощущение горького и

АДАПТАЦИЯ
Адаптация к воздействию вкусового вещества развивается медленно (минуты) и пропорциональна его
АДАПТАЦИЯ
Адаптация к воздействию вкусового вещества развивается медленно (минуты) и пропорциональна его
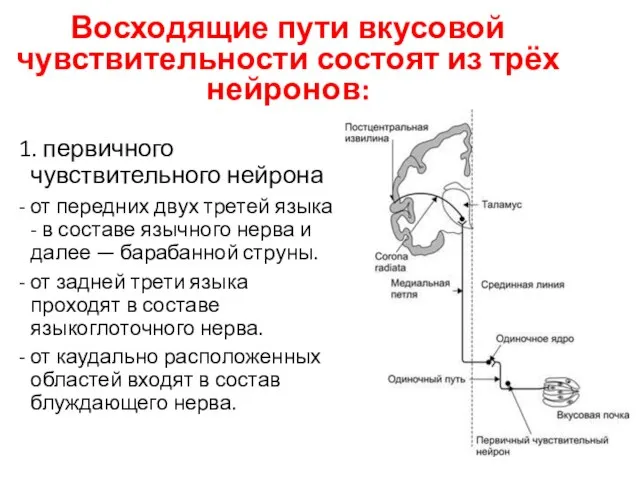
Восходящие пути вкусовой чувствительности состоят из трёх нейронов:
1. первичного чувствительного нейрона
-
Восходящие пути вкусовой чувствительности состоят из трёх нейронов:
1. первичного чувствительного нейрона
-
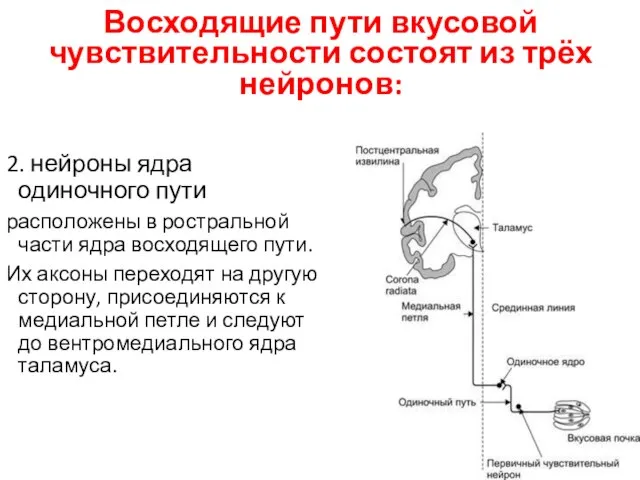
Восходящие пути вкусовой чувствительности состоят из трёх нейронов:
2. нейроны ядра
Восходящие пути вкусовой чувствительности состоят из трёх нейронов:
2. нейроны ядра
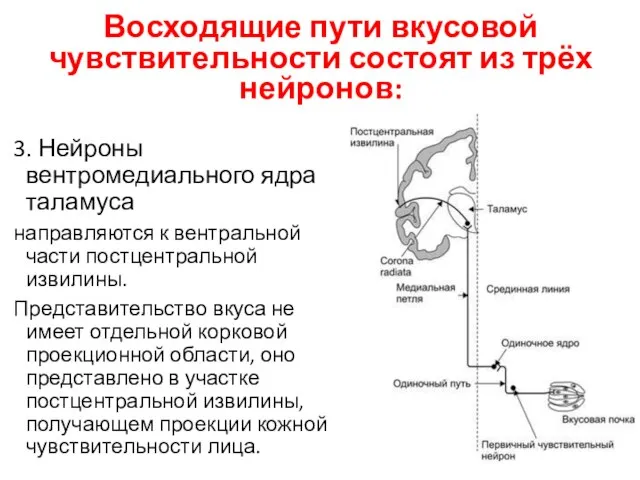
Восходящие пути вкусовой чувствительности состоят из трёх нейронов:
3. Нейроны вентромедиального ядра
Восходящие пути вкусовой чувствительности состоят из трёх нейронов:
3. Нейроны вентромедиального ядра

Тактильная сенсорная система
Тактильная сенсорная система

Тактильные ощущения прикосновения, давления и вибрации относятся к раздельным видам ощущений,
Тактильные ощущения прикосновения, давления и вибрации относятся к раздельным видам ощущений,
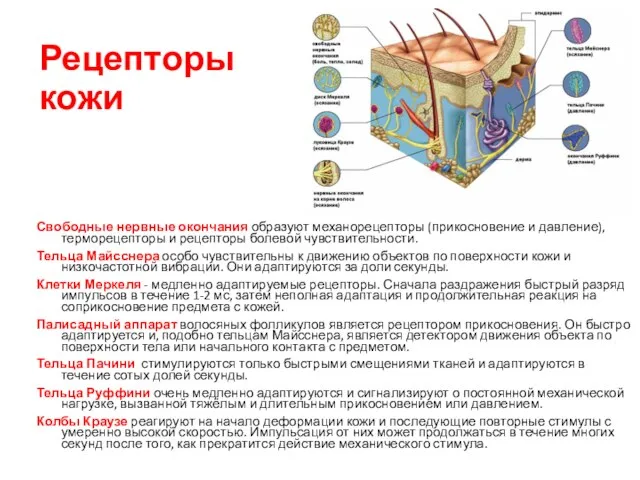
Рецепторы кожи
Свободные нервные окончания образуют механорецепторы (прикосновение и давление), терморецепторы и рецепторы
Рецепторы кожи
Свободные нервные окончания образуют механорецепторы (прикосновение и давление), терморецепторы и рецепторы

Количество и плотность размещения рецепторов
- Общее количество тактильных рецепторов оценивается в
Количество и плотность размещения рецепторов
- Общее количество тактильных рецепторов оценивается в
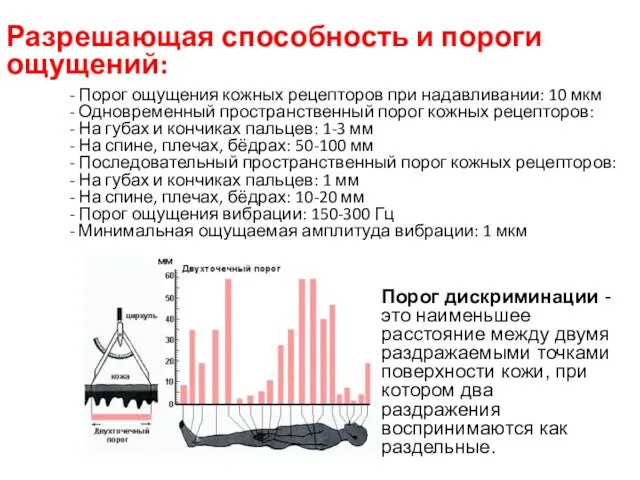
Разрешающая способность и пороги ощущений:
- Порог ощущения кожных рецепторов при надавливании:
Разрешающая способность и пороги ощущений:
- Порог ощущения кожных рецепторов при надавливании:
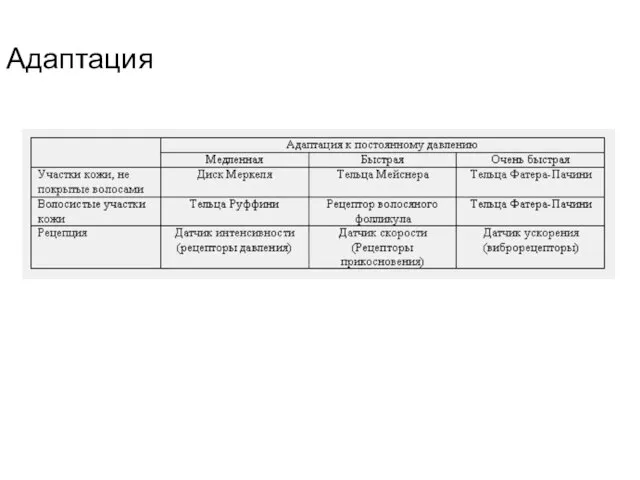
Адаптация
Адаптация

Передача импульсации
Передача тактильных ощущений тонкой дифференцировки происходит по миелинизированным нервным волокнам
Передача импульсации
Передача тактильных ощущений тонкой дифференцировки происходит по миелинизированным нервным волокнам
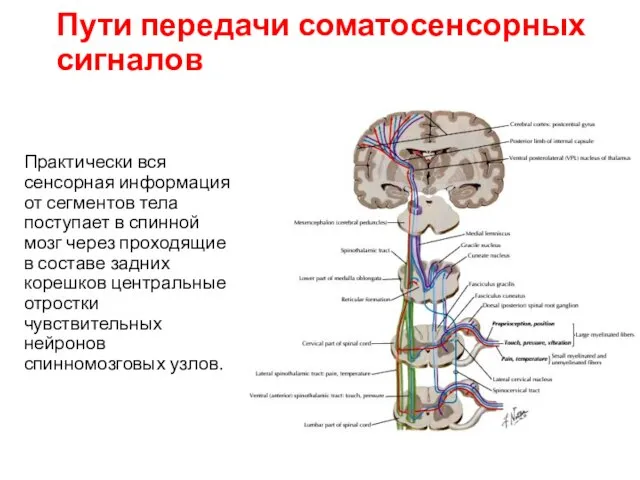
Пути передачи соматосенсорных сигналов
Практически вся сенсорная информация от сегментов тела поступает
Пути передачи соматосенсорных сигналов
Практически вся сенсорная информация от сегментов тела поступает
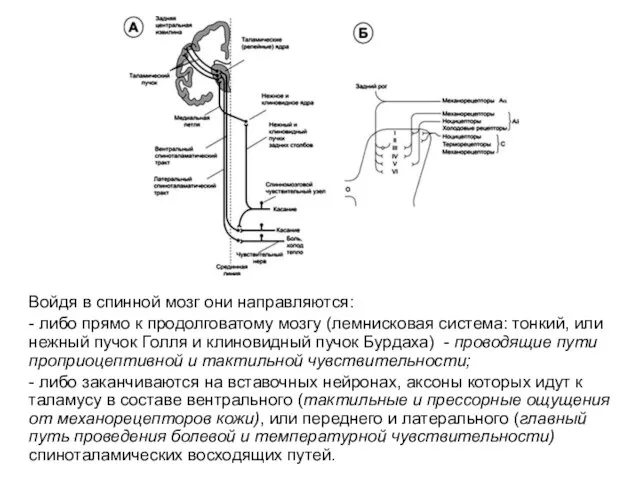
Войдя в спинной мозг они направляются:
- либо прямо к продолговатому мозгу
Войдя в спинной мозг они направляются:
- либо прямо к продолговатому мозгу
 Вирусы. Дмитрий Иосифович Ивановский
Вирусы. Дмитрий Иосифович Ивановский Синтетическая теория эволюции
Синтетическая теория эволюции Биоритмы человека: сова или жаворонок. Тест
Биоритмы человека: сова или жаворонок. Тест Ткани организма человека
Ткани организма человека Растения-хищники
Растения-хищники Животные Южной Америки
Животные Южной Америки Белки. Строение белков. Роль белков в клетке и организме человека
Белки. Строение белков. Роль белков в клетке и организме человека Круглые черви. Признаки круглых червей
Круглые черви. Признаки круглых червей Презентация Пищеварение в ротовой полости
Презентация Пищеварение в ротовой полости ОСНОВНЫЕ ПОНЯТИЯ ГЕНЕТИКИ
ОСНОВНЫЕ ПОНЯТИЯ ГЕНЕТИКИ Организм побеждает болезни УМК «Школа 2100» Урок окружающего мира
Организм побеждает болезни УМК «Школа 2100» Урок окружающего мира  Пищевод. Желудок. Кишечник. Печень и поджелудочная железа
Пищевод. Желудок. Кишечник. Печень и поджелудочная железа Veterinärmedizin. Milomir Kovac
Veterinärmedizin. Milomir Kovac Ткани, их строение, местонахождение и функции
Ткани, их строение, местонахождение и функции Индивидуальное развитие организмов. Биогенетический закон. Задачи: 1.Познакомитесь с индивидуальным развитием организмов. 2.Узна
Индивидуальное развитие организмов. Биогенетический закон. Задачи: 1.Познакомитесь с индивидуальным развитием организмов. 2.Узна Презентация на тему "Зачёт по теме «Пищеварение»" - скачать презентации по Биологии
Презентация на тему "Зачёт по теме «Пищеварение»" - скачать презентации по Биологии Насекомые. Средняя группа
Насекомые. Средняя группа Рефлекторная регуляция Урок биологии в 8 классе
Рефлекторная регуляция Урок биологии в 8 классе  Морфофункциональная характеристика аппарата движения. Исследование двигательных функций методом активных и пассивных движений
Морфофункциональная характеристика аппарата движения. Исследование двигательных функций методом активных и пассивных движений Сенсорные системы с волосковыми рецепторами
Сенсорные системы с волосковыми рецепторами Кошки в жизни человека
Кошки в жизни человека Место человека в живой природе
Место человека в живой природе Приспособления животных к среде обитания
Приспособления животных к среде обитания Плоды.Строение.Разнообразие и распространение
Плоды.Строение.Разнообразие и распространение Геологический календарь Земли
Геологический календарь Земли Презентация на тему ПОЛЕЗНАЯ АЗБУКА ПИТАНИЯ
Презентация на тему ПОЛЕЗНАЯ АЗБУКА ПИТАНИЯ Аттестационная работа. Проектная и исследовательская работа на уроках окружающего мира
Аттестационная работа. Проектная и исследовательская работа на уроках окружающего мира Ткани человека. 5
Ткани человека. 5