- Биохимия минерализованных тканей полости рта

Содержание
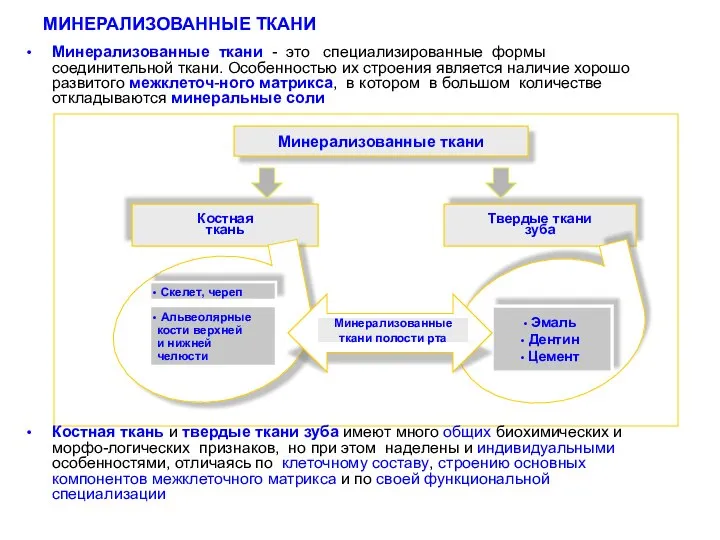
- 2. МИНЕРАЛИЗОВАННЫЕ ТКАНИ Минерализованные ткани - это специализированные формы соединительной ткани. Особенностью их строения является наличие хорошо
- 3. ФУНКЦИИ И ОСОБЕННОСТИ СТРОЕНИЯ КОСТНОЙ ТКАНИ Костная ткань (textus osseus) является сложно организованным и активно функционирующим
- 4. ПРОЦЕНТНОЕ СООТНОШЕНИЕ КОМПОНЕНТОВ КОСТНОЙ ТКАНИ - остеобласты - органические вещества - 30% - остеоциты - минеральные
- 5. СОСТАВ МЕЖКЛЕТОЧНОГО ВЕЩЕСТВА Особенности организации межклеточного матрикса, структура входящих в него органических и минеральных компонентов, а
- 6. ОСОБЕННОСТИ СОСТАВА МИНЕРАЛИЗОВАННЫХ ТКАНЕЙ ЗУБА Минерализованные ткани зуба, кроме зрелой эмали, содержат небольшое количество специфических клеток:
- 7. СТРОЕНИЕ И СВОЙСТВА МЕЖКЛЕТОЧНОГО МАТРИКСА Межклеточный матрикс (substantia intercellularis) минерализованных тканей состоит из волокнистых структур и
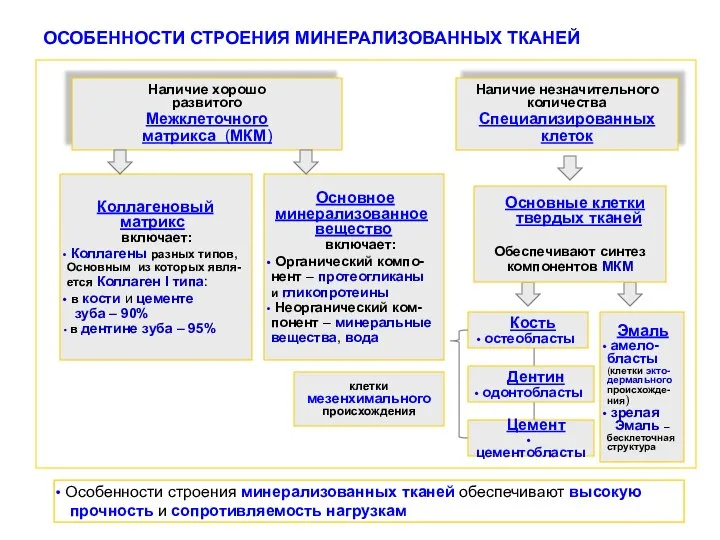
- 8. ОСОБЕННОСТИ СТРОЕНИЯ МИНЕРАЛИЗОВАННЫХ ТКАНЕЙ Особенности строения минерализованных тканей обеспечивают высокую прочность и сопротивляемость нагрузкам Наличие хорошо
- 9. КОМПОНЕНТЫ МЕЖКЛЕТОЧНОГО МАТРИКСА Компоненты межклеточного матрикса Гидроксиапатит костей является основным хранилищем: кальция – 99% фосфора –
- 10. Процентное соотношение органического и неорганического компонентов межклеточного матрикса минерализованных тканей Вода Органическое вещество Минеральное вещество Эмаль
- 11. ОСОБЕННОСТИ СОСТАВА МЕЖКЛЕТОЧНОГО ВЕЩЕСТВА РАЗНЫХ МИНЕРАЛИЗОВАННЫХ ТКАНЕЙ Основное отличие межклеточного матрикса отдельных минерализованных тканей заключается в
- 12. БЕЛКИ МКМ ТВЕРДЫХ ТКАНЕЙ Коллагены разных типов - структурная функция Неколлагеновые белки Фибронектин, ламинин - адгезивная
- 13. Костная ткань и твердые ткани зуба отличаются по составу и количеству коллагенов и неколлагеновых белков межклеточного
- 14. КОЛЛАГЕНЫ. СТРУКТУРА И ФУНКЦИИ Коллагены являются самыми распространенными белками в человеческом теле и входят в состав
- 15. ОСОБЕННОСТИ СТРОЕНИЯ КОЛЛАГЕНОВ МИНЕРАЛИЗОВАННЫХ ТКАНЕЙ Коллагены являются гликопротеинами с разным аминокислотным составом и количеством углеводов Молекула
- 16. СТРОЕНИЕ КОЛЛАГЕНА I ТИПА Коллаген I типа: [α1(І)]2α2(І) является гетерополимером, состоящим из разных α-полипептидных цепей римская
- 17. СОСТАВ И ДЛИНА ВОЛОКНА КОЛЛАГЕНОВ РАЗНЫХ ТИПОВ Тип коллагена Состав Длина волокна, нм
- 18. ОСОБЕННОСТИ КОЛЛАГЕНА I ТИПА КОСТНОЙ ТКАНИ Особенностью коллагена І типа костной ткани (остеоколлагена) является: высокое содержание
- 19. ДЕСМОЗИН И ИЗОДЕСМОЗИН В коллагене І типа кости присутствуют такие специфические структуры, как десмозин и изодесмозин,
- 20. ДЕСМОЗИН И ИЗОДЕСМОЗИН - В КЛИНИЧЕСКОЙ ПРАКТИКЕ Десмозин (пиридинолин) и изодесмозин (изопиридинолин) обеспечивают стабильность коллагенового матрикса
- 21. КОЛЛАГЕНОВЫЙ СОСТАВ МИНЕРАЛИЗОВАННЫХ ТКАНЕЙ Минерализованная ткань Типы коллагенов Костная ткань I, V, XI, XII Цемент зуба
- 22. Коллагены II, III,V, XI типов, как и коллаген І, относятся к фибриллообразующим коллагенам. Коллаген ІІ -
- 23. Коллаген образуется из секретируемого остеобластами предшественника – про-коллагена, представляющего собой тройную суперспираль, состоящую из 3-х левозакрученных
- 24. ОБРАЗОВАНИЕ ТРОПОКОЛЛАГЕНА 3-и α-спирали объединяются и образуют правозакрученную суперспиральную мо-лекулу тропоколлагена, имеющего форму стержня длиной 300
- 25. Из сформировавшихся молекул тропоколлагена происходит сборка коллагено-вых фибрилл, в которых одна нить молекул тропоколлагена сдвинута по
- 26. Коллагеновые микрофибриллы содержат 5 рядов молекул тропоколлагена, коллагеновые фибриллы включают 9 – 12 микрофибрилл Фибриллы представляют
- 27. БИОСИНТЕЗ И СОЗРЕВАНИЕ КОЛЛАГЕНА Синтез и созревание коллагена представляют сложный процесс, состоящий из целого ряда последовательных
- 28. Гидроксилирование остатков пролина и лизина является результатом котран-сляционного превращения соответствующих аминокислот в составе пептидных цепей проколлагена.
- 29. НАРУШЕНИЯ СИНТЕЗА И СОЗРЕВАНИЯ КОЛЛАГЕНА Оксипролин и оксилизин являются маркерными аминокислотами зрелого кол-лагена. Гидроксилирование остатков пролина
- 30. ГЛИКОЗИЛИРОВАНИЕ ОСТАТКОВ ГИДРОКСИЛИЗИНА В ПРОКОЛЛАГЕНЕ Гликозилтрнсфераза УДФ-галактоза УДФ УДФ-глюкоза УДФ Гликозилированный гидроксилизин Гидроксилизин О || -NН-СН-С-
- 31. ВНЕКЛЕТОЧНЫЙ ЭТАП СИНТЕЗА КОЛЛАГЕНА При участии специфических проколлагенпептидаз от проколлагена І, ІІ, ІІІ типов происходит отщепление
- 32. Остаток лизина Альдегидное производное лизина Остаток лизина Лизилнорлейцин Альдегидные производные лизина Альдольная межмолекулярная сшивка ЛО ОБРАЗОВАНИЕ
- 33. ЭТАПЫ ФОРМИРОВАНИЯ КОЛЛАГЕНОВОГО ВОЛОКНА Глицин Пролин Глицин Глицин Глицин 64 нм 3 α–цепи коллагена Тропоколлагеновые нити
- 34. ЛОКАЛИЗАЦИЯ И ПОСЛЕДОВАТЕЛЬНОСТЬ ЭТАПОВ ПРОЦЕССИНГА КОЛЛАГЕНА
- 35. ГЕНЕТИЧЕСКИ ОБУСЛОВЛЕННЫЕ НАРУШЕНИЯ СИНТЕЗА И СОЗРЕВАНИЯ КОЛЛАГЕНА Генетически обусловленные нарушения синтеза и созревания коллагена приво-дят к
- 36. Период полужизни коллагена составляет несколько недель - месяцев. Распад коллагена происходит при участии Са2+, Zn2+-зависимой коллагеназы,
- 38. Скачать презентацию
МИНЕРАЛИЗОВАННЫЕ ТКАНИ
Минерализованные ткани - это специализированные формы соединительной ткани. Особенностью их
МИНЕРАЛИЗОВАННЫЕ ТКАНИ
Минерализованные ткани - это специализированные формы соединительной ткани. Особенностью их

ФУНКЦИИ И ОСОБЕННОСТИ СТРОЕНИЯ КОСТНОЙ ТКАНИ
Костная ткань (textus osseus) является сложно
ФУНКЦИИ И ОСОБЕННОСТИ СТРОЕНИЯ КОСТНОЙ ТКАНИ
Костная ткань (textus osseus) является сложно

ПРОЦЕНТНОЕ СООТНОШЕНИЕ КОМПОНЕНТОВ КОСТНОЙ ТКАНИ
- остеобласты - органические вещества - 30%
ПРОЦЕНТНОЕ СООТНОШЕНИЕ КОМПОНЕНТОВ КОСТНОЙ ТКАНИ
- остеобласты - органические вещества - 30%

СОСТАВ МЕЖКЛЕТОЧНОГО ВЕЩЕСТВА
Особенности организации межклеточного матрикса, структура входящих в него органических
СОСТАВ МЕЖКЛЕТОЧНОГО ВЕЩЕСТВА
Особенности организации межклеточного матрикса, структура входящих в него органических

ОСОБЕННОСТИ СОСТАВА МИНЕРАЛИЗОВАННЫХ ТКАНЕЙ ЗУБА
Минерализованные ткани зуба, кроме зрелой эмали, содержат
ОСОБЕННОСТИ СОСТАВА МИНЕРАЛИЗОВАННЫХ ТКАНЕЙ ЗУБА
Минерализованные ткани зуба, кроме зрелой эмали, содержат

СТРОЕНИЕ И СВОЙСТВА МЕЖКЛЕТОЧНОГО МАТРИКСА
Межклеточный матрикс (substantia intercellularis) минерализованных тканей состоит
СТРОЕНИЕ И СВОЙСТВА МЕЖКЛЕТОЧНОГО МАТРИКСА
Межклеточный матрикс (substantia intercellularis) минерализованных тканей состоит
ОСОБЕННОСТИ СТРОЕНИЯ МИНЕРАЛИЗОВАННЫХ ТКАНЕЙ
Особенности строения минерализованных тканей обеспечивают высокую
ОСОБЕННОСТИ СТРОЕНИЯ МИНЕРАЛИЗОВАННЫХ ТКАНЕЙ
Особенности строения минерализованных тканей обеспечивают высокую

КОМПОНЕНТЫ МЕЖКЛЕТОЧНОГО МАТРИКСА
Компоненты межклеточного матрикса
Гидроксиапатит костей
является основным
хранилищем:
КОМПОНЕНТЫ МЕЖКЛЕТОЧНОГО МАТРИКСА
Компоненты межклеточного матрикса
Гидроксиапатит костей
является основным
хранилищем:
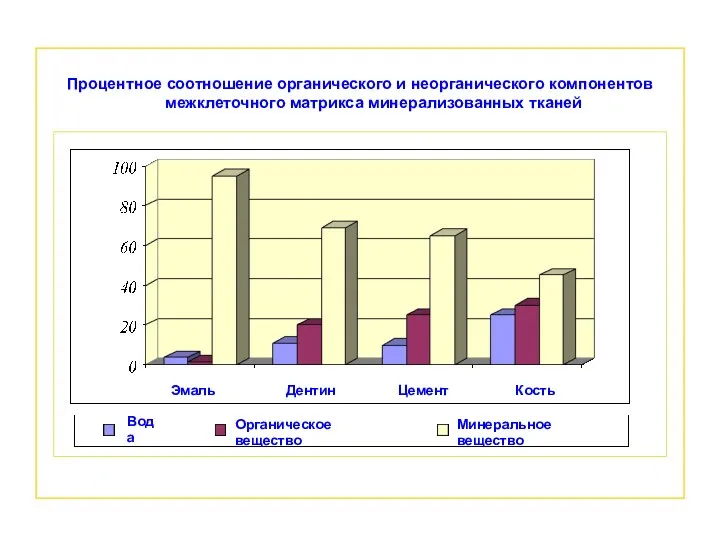
Процентное соотношение органического и неорганического компонентов межклеточного матрикса минерализованных тканей
Вода
Органическое вещество
Минеральное
Вода
Органическое вещество
Минеральное

ОСОБЕННОСТИ СОСТАВА МЕЖКЛЕТОЧНОГО ВЕЩЕСТВА РАЗНЫХ
МИНЕРАЛИЗОВАННЫХ ТКАНЕЙ
Основное отличие межклеточного матрикса отдельных минерализованных
ОСОБЕННОСТИ СОСТАВА МЕЖКЛЕТОЧНОГО ВЕЩЕСТВА РАЗНЫХ
МИНЕРАЛИЗОВАННЫХ ТКАНЕЙ
Основное отличие межклеточного матрикса отдельных минерализованных
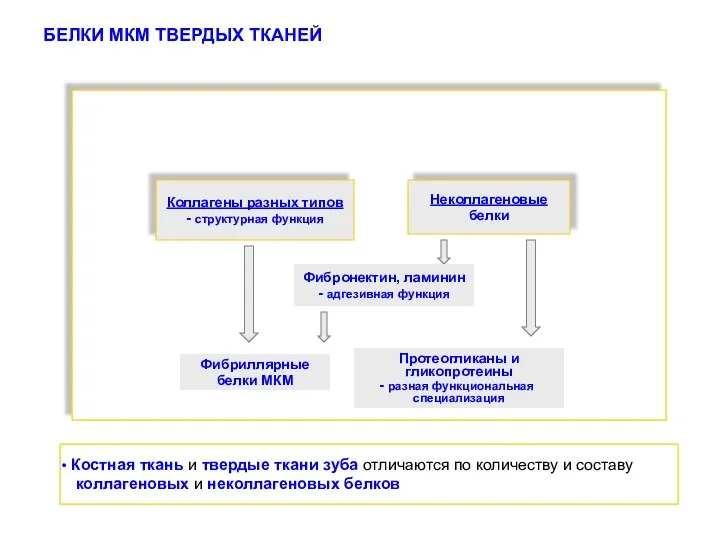
БЕЛКИ МКМ ТВЕРДЫХ ТКАНЕЙ
Коллагены разных типов
- структурная функция
Неколлагеновые
белки
Фибронектин, ламинин
- адгезивная функция
Протеогликаны
БЕЛКИ МКМ ТВЕРДЫХ ТКАНЕЙ
Коллагены разных типов
- структурная функция
Неколлагеновые
белки
Фибронектин, ламинин
- адгезивная функция
Протеогликаны
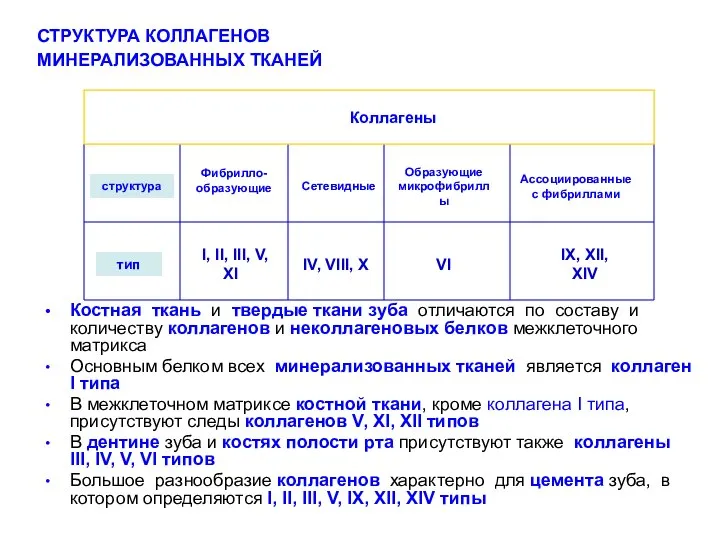
Костная ткань и твердые ткани зуба отличаются по составу и количеству
Костная ткань и твердые ткани зуба отличаются по составу и количеству

КОЛЛАГЕНЫ. СТРУКТУРА И ФУНКЦИИ
Коллагены являются самыми распространенными белками в человеческом теле
КОЛЛАГЕНЫ. СТРУКТУРА И ФУНКЦИИ
Коллагены являются самыми распространенными белками в человеческом теле

ОСОБЕННОСТИ СТРОЕНИЯ КОЛЛАГЕНОВ
МИНЕРАЛИЗОВАННЫХ ТКАНЕЙ
Коллагены являются гликопротеинами с разным аминокислотным составом
ОСОБЕННОСТИ СТРОЕНИЯ КОЛЛАГЕНОВ
МИНЕРАЛИЗОВАННЫХ ТКАНЕЙ
Коллагены являются гликопротеинами с разным аминокислотным составом
![СТРОЕНИЕ КОЛЛАГЕНА I ТИПА Коллаген I типа: [α1(І)]2α2(І) является гетерополимером, состоящим](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/662851/slide-15.jpg)
СТРОЕНИЕ КОЛЛАГЕНА I ТИПА
Коллаген I типа: [α1(І)]2α2(І) является гетерополимером, состоящим из
СТРОЕНИЕ КОЛЛАГЕНА I ТИПА
Коллаген I типа: [α1(І)]2α2(І) является гетерополимером, состоящим из
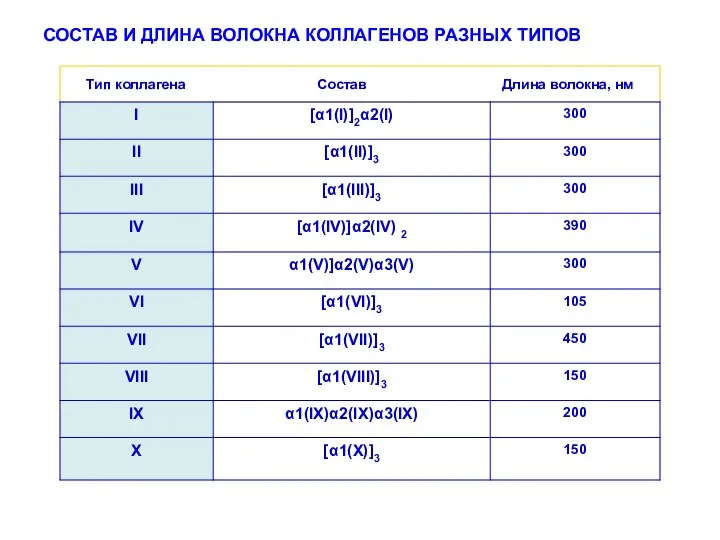
СОСТАВ И ДЛИНА ВОЛОКНА КОЛЛАГЕНОВ РАЗНЫХ ТИПОВ
Тип коллагена Состав Длина
СОСТАВ И ДЛИНА ВОЛОКНА КОЛЛАГЕНОВ РАЗНЫХ ТИПОВ
Тип коллагена Состав Длина
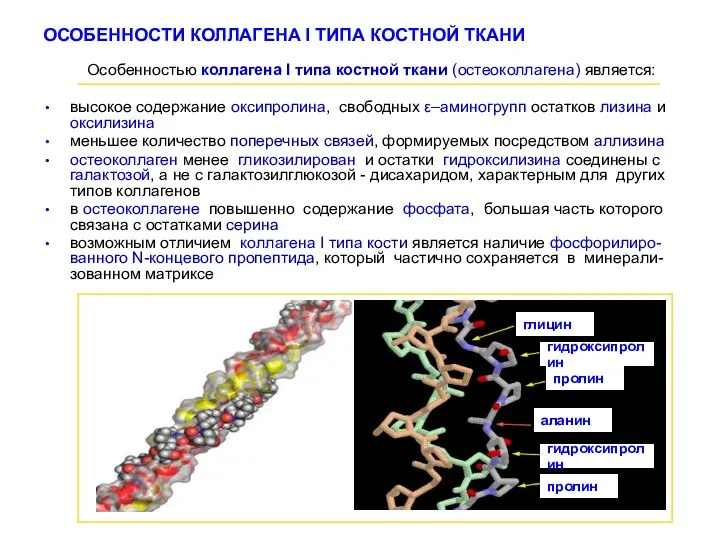
ОСОБЕННОСТИ КОЛЛАГЕНА I ТИПА КОСТНОЙ ТКАНИ
Особенностью коллагена І типа костной
ОСОБЕННОСТИ КОЛЛАГЕНА I ТИПА КОСТНОЙ ТКАНИ
Особенностью коллагена І типа костной
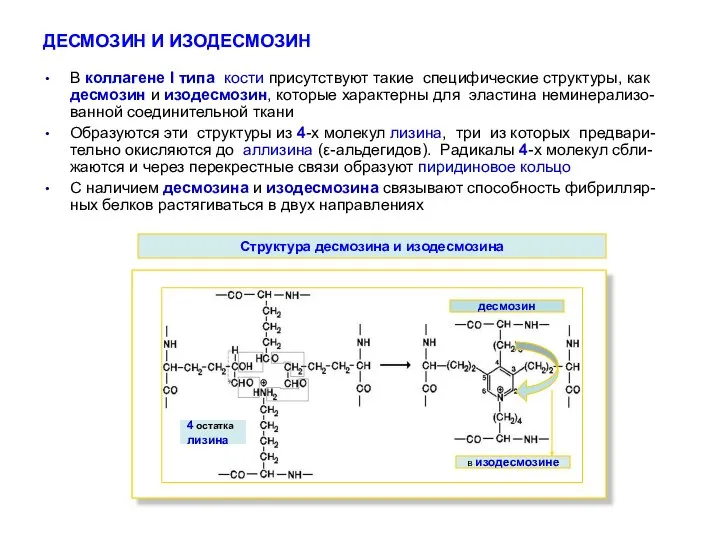
ДЕСМОЗИН И ИЗОДЕСМОЗИН
В коллагене І типа кости присутствуют такие специфические структуры,
ДЕСМОЗИН И ИЗОДЕСМОЗИН
В коллагене І типа кости присутствуют такие специфические структуры,

ДЕСМОЗИН И ИЗОДЕСМОЗИН - В КЛИНИЧЕСКОЙ ПРАКТИКЕ
Десмозин (пиридинолин) и изодесмозин (изопиридинолин)
ДЕСМОЗИН И ИЗОДЕСМОЗИН - В КЛИНИЧЕСКОЙ ПРАКТИКЕ
Десмозин (пиридинолин) и изодесмозин (изопиридинолин)

КОЛЛАГЕНОВЫЙ СОСТАВ МИНЕРАЛИЗОВАННЫХ ТКАНЕЙ
Минерализованная ткань Типы коллагенов
Костная ткань I, V, XI,
КОЛЛАГЕНОВЫЙ СОСТАВ МИНЕРАЛИЗОВАННЫХ ТКАНЕЙ
Минерализованная ткань Типы коллагенов
Костная ткань I, V, XI,

Коллагены II, III,V, XI типов, как и коллаген І, относятся к
Коллагены II, III,V, XI типов, как и коллаген І, относятся к

Коллаген образуется из секретируемого остеобластами предшественника – про-коллагена, представляющего собой тройную
Коллаген образуется из секретируемого остеобластами предшественника – про-коллагена, представляющего собой тройную

ОБРАЗОВАНИЕ ТРОПОКОЛЛАГЕНА
3-и α-спирали объединяются и образуют правозакрученную суперспиральную мо-лекулу тропоколлагена, имеющего
ОБРАЗОВАНИЕ ТРОПОКОЛЛАГЕНА
3-и α-спирали объединяются и образуют правозакрученную суперспиральную мо-лекулу тропоколлагена, имеющего

Из сформировавшихся молекул тропоколлагена происходит сборка коллагено-вых фибрилл, в которых одна
Из сформировавшихся молекул тропоколлагена происходит сборка коллагено-вых фибрилл, в которых одна

Коллагеновые микрофибриллы содержат 5 рядов молекул тропоколлагена, коллагеновые фибриллы включают 9
Коллагеновые микрофибриллы содержат 5 рядов молекул тропоколлагена, коллагеновые фибриллы включают 9

БИОСИНТЕЗ И СОЗРЕВАНИЕ КОЛЛАГЕНА
Синтез и созревание коллагена представляют сложный процесс, состоящий
БИОСИНТЕЗ И СОЗРЕВАНИЕ КОЛЛАГЕНА
Синтез и созревание коллагена представляют сложный процесс, состоящий

Гидроксилирование остатков пролина и лизина является результатом котран-сляционного превращения соответствующих аминокислот
Гидроксилирование остатков пролина и лизина является результатом котран-сляционного превращения соответствующих аминокислот

НАРУШЕНИЯ СИНТЕЗА И
СОЗРЕВАНИЯ КОЛЛАГЕНА
Оксипролин и оксилизин являются маркерными аминокислотами зрелого
НАРУШЕНИЯ СИНТЕЗА И
СОЗРЕВАНИЯ КОЛЛАГЕНА
Оксипролин и оксилизин являются маркерными аминокислотами зрелого
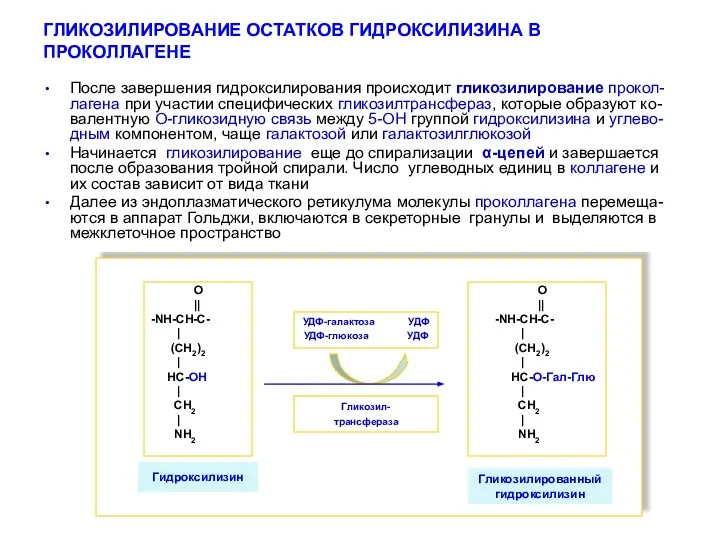
ГЛИКОЗИЛИРОВАНИЕ ОСТАТКОВ ГИДРОКСИЛИЗИНА В ПРОКОЛЛАГЕНЕ
Гликозилтрнсфераза
УДФ-галактоза УДФ
УДФ-глюкоза УДФ
Гликозилированный
гидроксилизин
Гидроксилизин
О
||
-NН-СН-С-
|
ГЛИКОЗИЛИРОВАНИЕ ОСТАТКОВ ГИДРОКСИЛИЗИНА В ПРОКОЛЛАГЕНЕ
Гликозилтрнсфераза
УДФ-галактоза УДФ
УДФ-глюкоза УДФ
Гликозилированный
гидроксилизин
Гидроксилизин
О
||
-NН-СН-С-
|

ВНЕКЛЕТОЧНЫЙ ЭТАП СИНТЕЗА КОЛЛАГЕНА
При участии специфических проколлагенпептидаз от проколлагена І, ІІ,
ВНЕКЛЕТОЧНЫЙ ЭТАП СИНТЕЗА КОЛЛАГЕНА
При участии специфических проколлагенпептидаз от проколлагена І, ІІ,

Остаток лизина
Альдегидное
производное
лизина
Остаток лизина
Лизилнорлейцин
Альдегидные производные лизина
Альдольная межмолекулярная сшивка
ЛО
ОБРАЗОВАНИЕ АЛЛИЗИНА И ЛИЗИНОВЫХ СШИВОК
Остаток лизина
Альдегидное
производное
лизина
Остаток лизина
Лизилнорлейцин
Альдегидные производные лизина
Альдольная межмолекулярная сшивка
ЛО
ОБРАЗОВАНИЕ АЛЛИЗИНА И ЛИЗИНОВЫХ СШИВОК
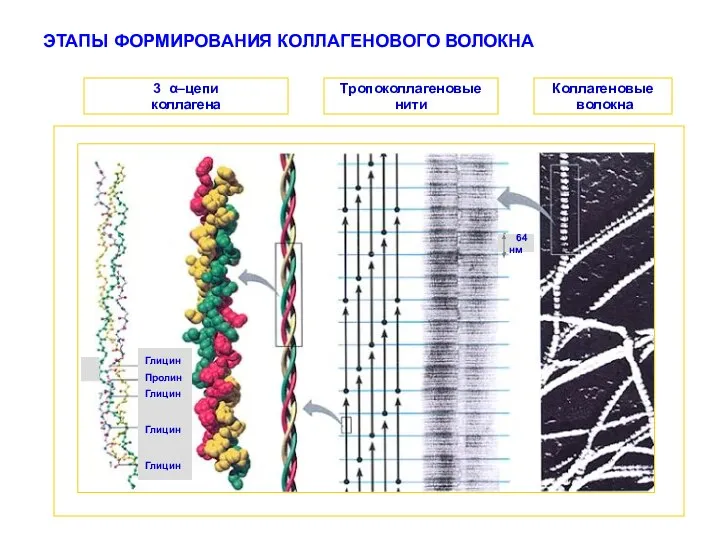
ЭТАПЫ ФОРМИРОВАНИЯ КОЛЛАГЕНОВОГО ВОЛОКНА
Глицин
Пролин
Глицин
Глицин
Глицин
64 нм
3 α–цепи
коллагена
Тропоколлагеновые
нити
Коллагеновые
волокна
ЭТАПЫ ФОРМИРОВАНИЯ КОЛЛАГЕНОВОГО ВОЛОКНА
Глицин
Пролин
Глицин
Глицин
Глицин
64 нм
3 α–цепи
коллагена
Тропоколлагеновые
нити
Коллагеновые
волокна
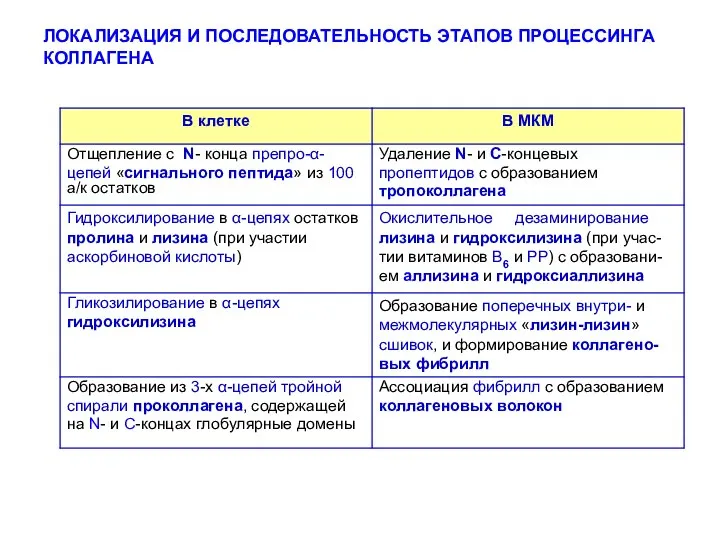
ЛОКАЛИЗАЦИЯ И ПОСЛЕДОВАТЕЛЬНОСТЬ ЭТАПОВ ПРОЦЕССИНГА КОЛЛАГЕНА
ЛОКАЛИЗАЦИЯ И ПОСЛЕДОВАТЕЛЬНОСТЬ ЭТАПОВ ПРОЦЕССИНГА КОЛЛАГЕНА
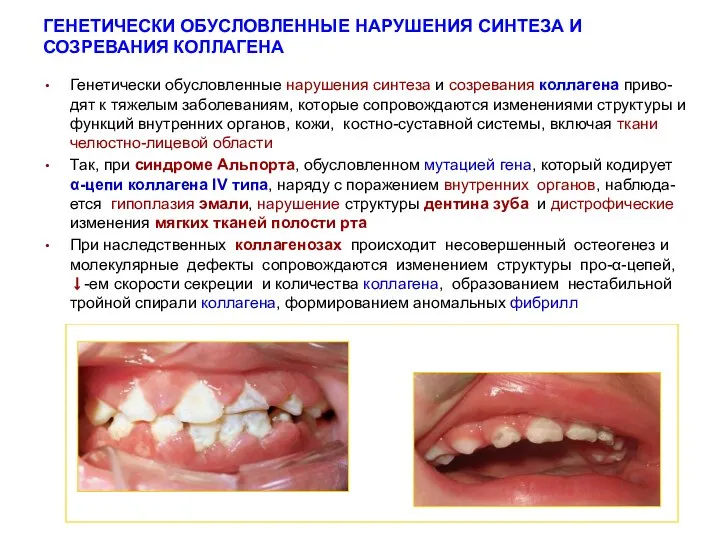
ГЕНЕТИЧЕСКИ ОБУСЛОВЛЕННЫЕ НАРУШЕНИЯ СИНТЕЗА И СОЗРЕВАНИЯ КОЛЛАГЕНА
Генетически обусловленные нарушения синтеза
ГЕНЕТИЧЕСКИ ОБУСЛОВЛЕННЫЕ НАРУШЕНИЯ СИНТЕЗА И СОЗРЕВАНИЯ КОЛЛАГЕНА
Генетически обусловленные нарушения синтеза

Период полужизни коллагена составляет несколько недель - месяцев. Распад коллагена происходит
Период полужизни коллагена составляет несколько недель - месяцев. Распад коллагена происходит
 Строение, распространение и практическое применение мхов
Строение, распространение и практическое применение мхов Презентация по биологии Почвенное питание растений
Презентация по биологии Почвенное питание растений  Строение и значение нервной системы
Строение и значение нервной системы Презентация на тему "Волки" - скачать презентации по Биологии
Презентация на тему "Волки" - скачать презентации по Биологии Отряд сумчатые
Отряд сумчатые  Империя эукариоты
Империя эукариоты Растения – живой организм Автор: учитель биологии ГБОУ ЦО №1456 Водопьянова Марина Александровна
Растения – живой организм Автор: учитель биологии ГБОУ ЦО №1456 Водопьянова Марина Александровна Мозг необъятен как Вселенная
Мозг необъятен как Вселенная Значение воды в жизнедеятельности растений
Значение воды в жизнедеятельности растений Вермикультура: технологии и перспективы
Вермикультура: технологии и перспективы Первый русский агроном
Первый русский агроном  Ботаника. Лист
Ботаника. Лист Строение эукариотической клетки Клетка
Строение эукариотической клетки Клетка  Многоклеточные (Metazoa)
Многоклеточные (Metazoa) Дыхательная система
Дыхательная система Сверлящие субстрат беспозвоночные животные
Сверлящие субстрат беспозвоночные животные Весна
Весна Взаимодействие генов
Взаимодействие генов Что происходит с нашей кожей зимой
Что происходит с нашей кожей зимой « Особенности организации нервных окончаний».
« Особенности организации нервных окончаний».  Водоросли. Строение и значение. Многообразие водорослей. 5 класс
Водоросли. Строение и значение. Многообразие водорослей. 5 класс Вклад отечественных ученых в разработку учения о высшей нервной деятельности
Вклад отечественных ученых в разработку учения о высшей нервной деятельности  Что такое старение
Что такое старение Уровни организации живого мира (10 класс)
Уровни организации живого мира (10 класс) Организмы, способные жить при крайних значениях температуры
Организмы, способные жить при крайних значениях температуры Интродукция сельскохозяйственных растений
Интродукция сельскохозяйственных растений О чем поет морская канарейка
О чем поет морская канарейка Древние животные
Древние животные