- Эквивалентная электрическая модель нейрона

Содержание
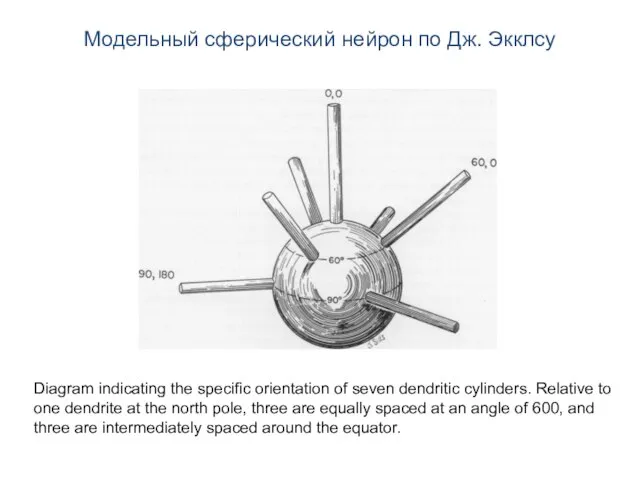
- 2. Модельный сферический нейрон по Дж. Экклсу Diagram indicating the specific orientation of seven dendritic cylinders. Relative
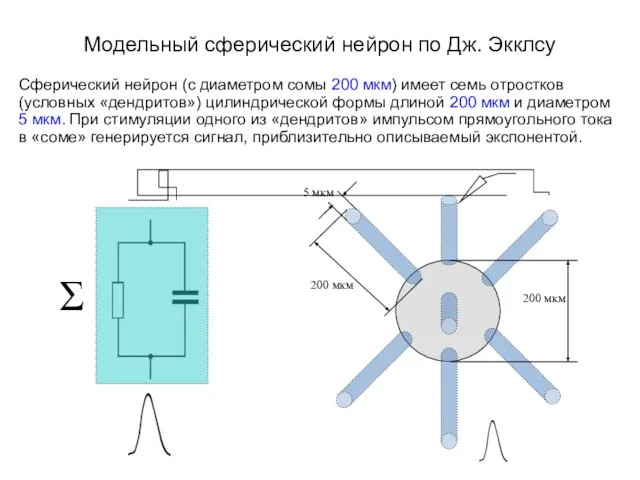
- 3. Модельный сферический нейрон по Дж. Экклсу Сферический нейрон (с диаметром сомы 200 мкм) имеет семь отростков
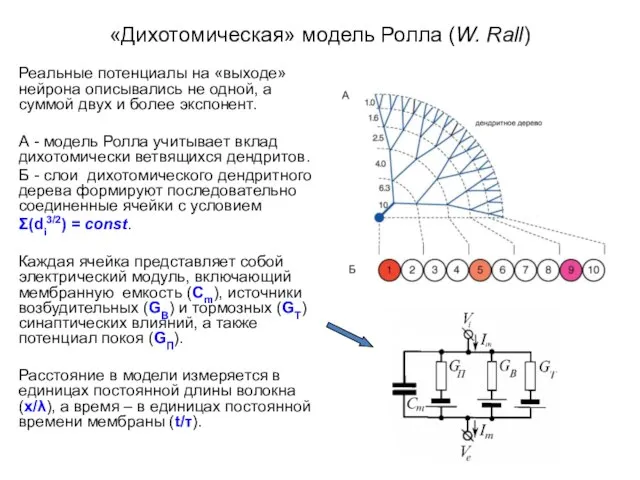
- 4. «Дихотомическая» модель Ролла (W. Rall) Реальные потенциалы на «выходе» нейрона описывались не одной, а суммой двух
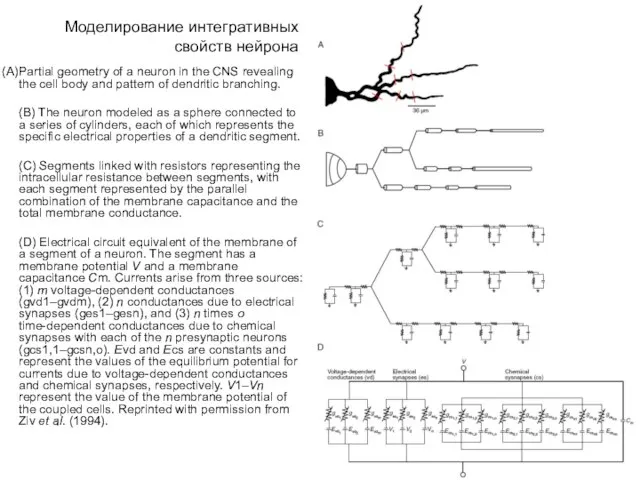
- 5. Моделирование интегративных свойств нейрона Partial geometry of a neuron in the CNS revealing the cell body
- 6. Моделирование интегративных свойств нейрона Electrical circuit equivalent of the membrane of a segment of a neuron.
- 7. «Дихотомическая» модель Ролла (W. Rall) http://www.scholarpedia.org/article/Rall_model (А) Модель нейрона с дендритами (модель Ролла), числа слева -
- 8. «Дихотомическая» модель Ролла (W. Rall) Форма синаптических потенциалов, рассчитанных по модели Ролла Оценивают два параметра –
- 9. «Дихотомическая» модель Ролла (W. Rall) Форма синаптических потенциалов, рассчитанных по модели Ролла ВПСП при разном удалении
- 10. «Дихотомическая» модель Ролла (W. Rall) Форма ВПСП в мотонейронах спинного мозга Серия ВПСП, зарегистрированных при разном
- 11. Электрические свойства мембраны нейрона Электрические свойства (входное сопротивление, Rвх) различных участков мембраны нейрона (сома, дендриты) значительно
- 12. Электрические свойства мембраны нейрона Зависимость порогового тока (synaptic current) от входного сопротивления (input resistance). Виртуальная гиперболическая
- 13. Генерация ПД в нейроне Потенциалы действия мотонейрона при различных способах стимуляции: А - антидромный; Б -
- 14. Генерация ПД в нейроне При антидромной стимуляции при различных уровнях фиксации тока выделяются три отдельных компонента:
- 15. Генерация ПД в нейроне При антидромной стимуляции при различных уровнях фиксации тока выделяются три отдельных компонента:
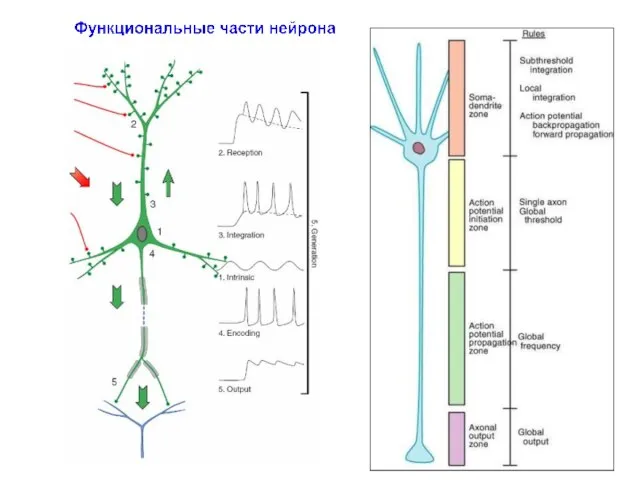
- 16. 1. Генерация собственной активности (в любом фрагменте нейрона за счет вольт-зависимых свойств мембраны и внутренних путей
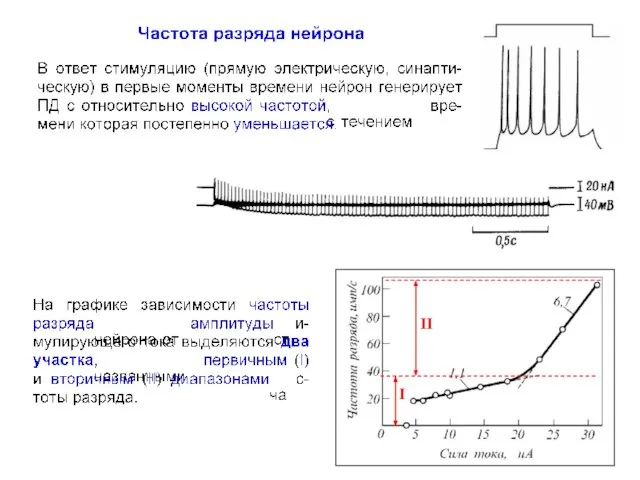
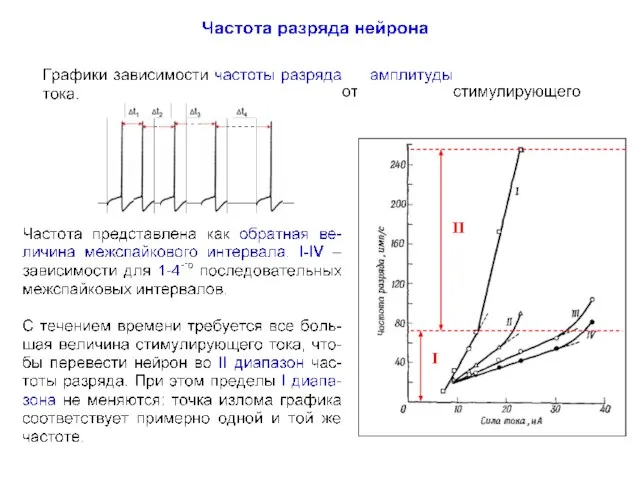
- 18. I II
- 19. Fi = 1/∆ti
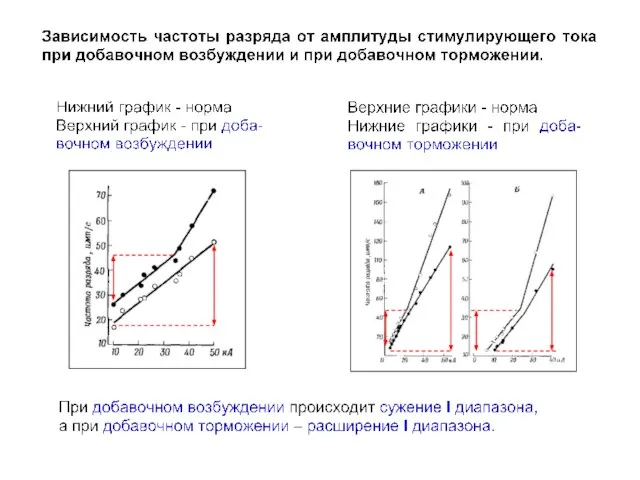
- 20. I II
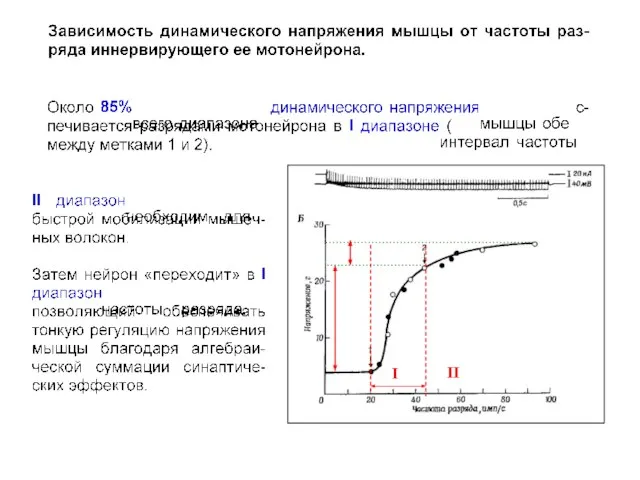
- 23. I II
- 27. Суммарная активность мозга (ЭЭГ) При активации синапсов поверхностные слои коры заряжены отрицательно, и электрод, расположенный на
- 28. Суммарная активность мозга (ЭЭГ) Полярность потенциалов в зависимости от локализации активирующих входов на функциональных сегментах нейрона.
- 29. Суммарная активность мозга (ЭЭГ) Суммарная электрическая активность представляет собой синхронизированные постсинаптические потенциалы.
- 30. Синхронизация ЭЭГ (слева от линии) сменяется десинхронизацией ЭЭГ (справа от линии) при сенсорном воздействии
- 34. Распределение потенциала плоскопараллельного диполя Открытое дендритное поле (сома нейронов возбуждена)
- 35. Распределение потенциала сферического диполя Закрытое дендритное поле в соме торможение в соме возбуждение
- 36. Распределение потенциала полусферического диполя Полузакрытое дендритное поле
- 37. Структура сетчатки позвоночных
- 40. Сложная пространственная конфигурация коры мозга затрудняет интерпретацию потенциалов, регистрируемых с поверхности головы Срез затылочного полюса мозга
- 41. Слоистое строение зрительной коры мозга (как и строение сетчатки лягушки) надо учитывать при интерпретации потенциалов, регистрируемых
- 43. Скачать презентацию
Модельный сферический нейрон по Дж. Экклсу
Diagram indicating the specific orientation of
Модельный сферический нейрон по Дж. Экклсу
Diagram indicating the specific orientation of
Модельный сферический нейрон по Дж. Экклсу
Сферический нейрон (с диаметром сомы 200
Модельный сферический нейрон по Дж. Экклсу
Сферический нейрон (с диаметром сомы 200
«Дихотомическая» модель Ролла (W. Rall)
Реальные потенциалы на «выходе» нейрона описывались не
«Дихотомическая» модель Ролла (W. Rall)
Реальные потенциалы на «выходе» нейрона описывались не
Моделирование интегративных свойств нейрона
Partial geometry of a neuron in the CNS
Моделирование интегративных свойств нейрона
Partial geometry of a neuron in the CNS
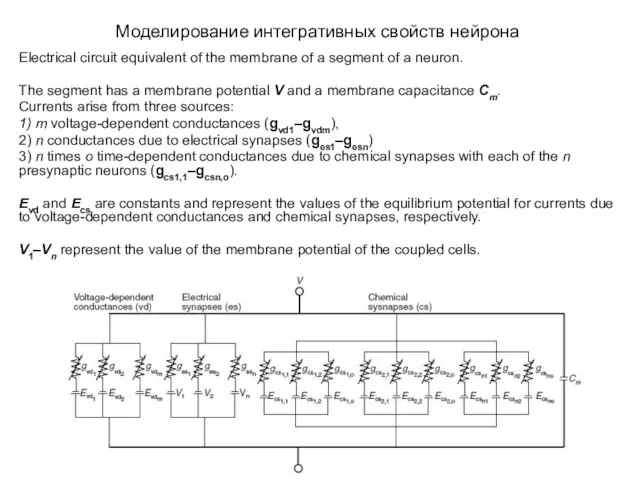
Моделирование интегративных свойств нейрона
Electrical circuit equivalent of the membrane of a
Моделирование интегративных свойств нейрона
Electrical circuit equivalent of the membrane of a
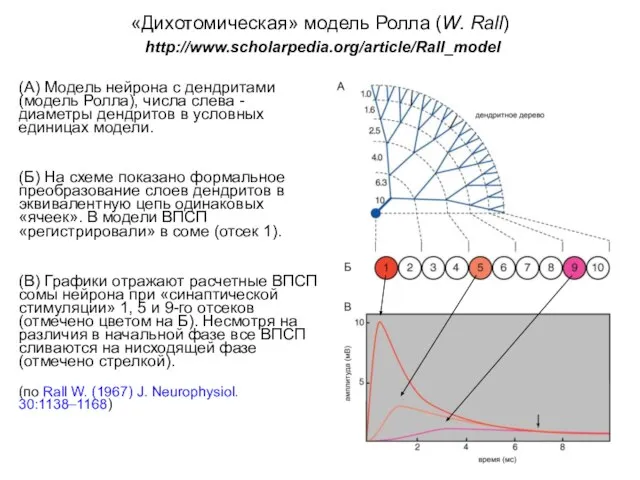
«Дихотомическая» модель Ролла (W. Rall)
http://www.scholarpedia.org/article/Rall_model
(А) Модель нейрона с дендритами (модель
«Дихотомическая» модель Ролла (W. Rall)
http://www.scholarpedia.org/article/Rall_model
(А) Модель нейрона с дендритами (модель
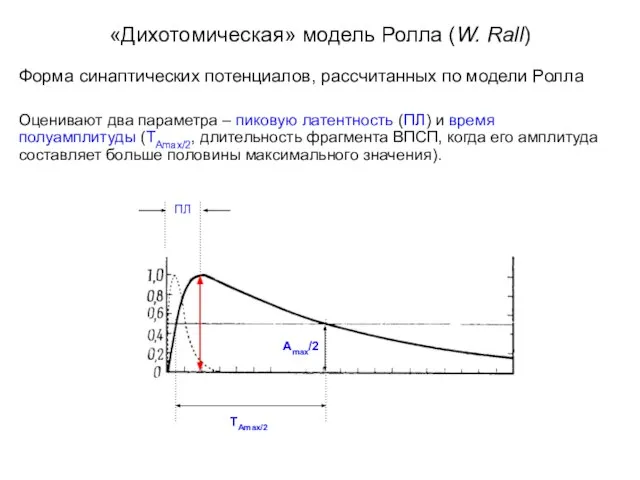
«Дихотомическая» модель Ролла (W. Rall)
Форма синаптических потенциалов, рассчитанных по модели Ролла
Оценивают
«Дихотомическая» модель Ролла (W. Rall)
Форма синаптических потенциалов, рассчитанных по модели Ролла
Оценивают
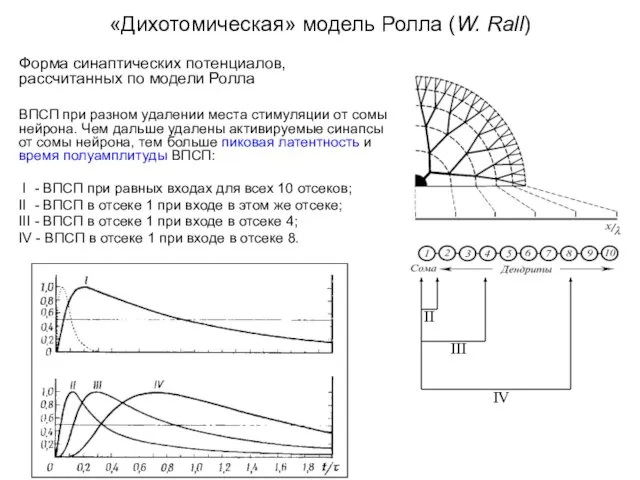
«Дихотомическая» модель Ролла (W. Rall)
Форма синаптических потенциалов, рассчитанных по модели Ролла
ВПСП
«Дихотомическая» модель Ролла (W. Rall)
Форма синаптических потенциалов, рассчитанных по модели Ролла
ВПСП
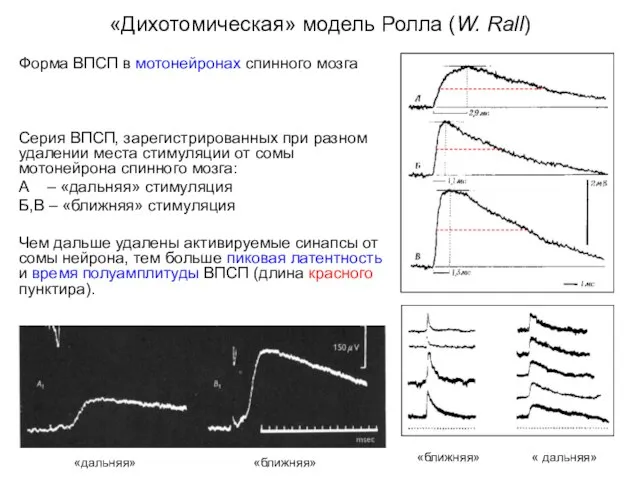
«Дихотомическая» модель Ролла (W. Rall)
Форма ВПСП в мотонейронах спинного мозга
Серия ВПСП,
«Дихотомическая» модель Ролла (W. Rall)
Форма ВПСП в мотонейронах спинного мозга
Серия ВПСП,
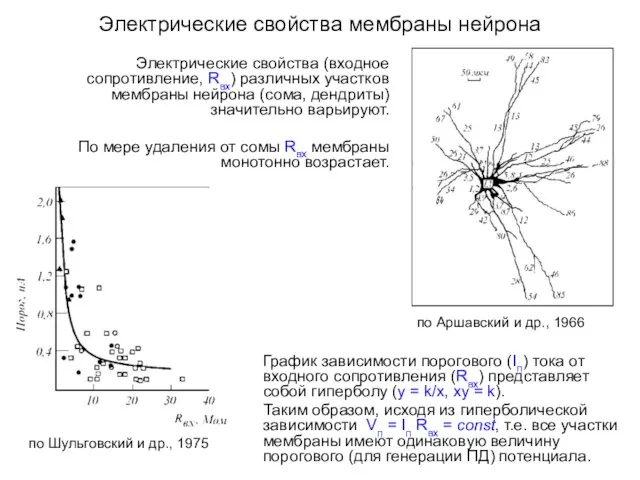
Электрические свойства мембраны нейрона
Электрические свойства (входное сопротивление, Rвх) различных участков мембраны
Электрические свойства мембраны нейрона
Электрические свойства (входное сопротивление, Rвх) различных участков мембраны
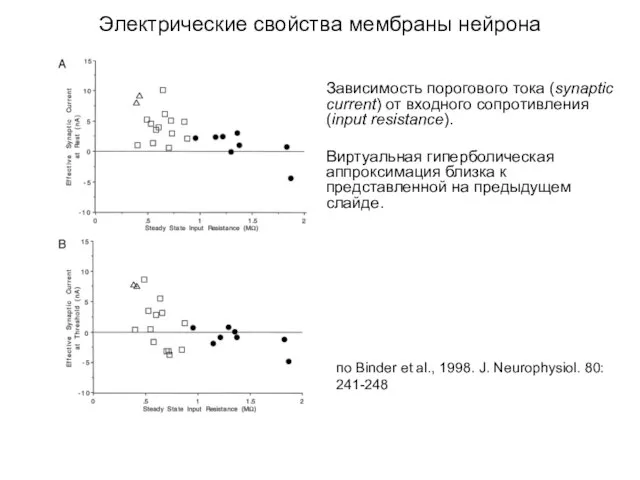
Электрические свойства мембраны нейрона
Зависимость порогового тока (synaptic current) от входного сопротивления
Электрические свойства мембраны нейрона
Зависимость порогового тока (synaptic current) от входного сопротивления

Генерация ПД в нейроне
Потенциалы действия мотонейрона при различных способах стимуляции:
А -
Генерация ПД в нейроне
Потенциалы действия мотонейрона при различных способах стимуляции:
А -
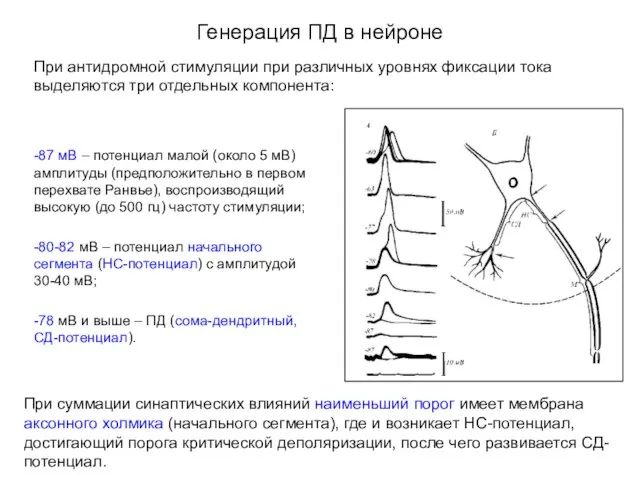
Генерация ПД в нейроне
При антидромной стимуляции при различных уровнях фиксации тока
Генерация ПД в нейроне
При антидромной стимуляции при различных уровнях фиксации тока
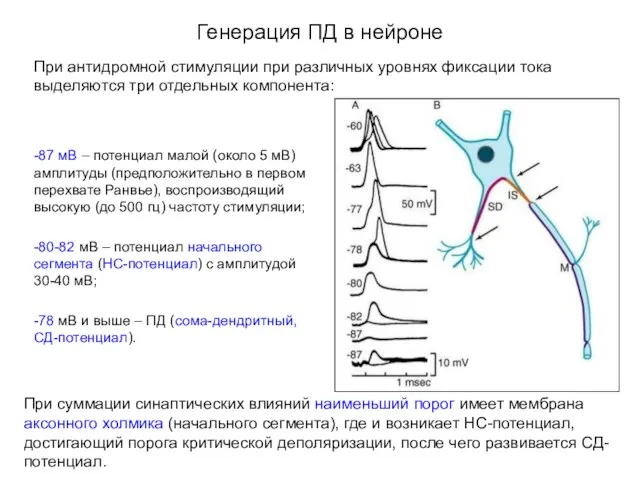
Генерация ПД в нейроне
При антидромной стимуляции при различных уровнях фиксации тока
Генерация ПД в нейроне
При антидромной стимуляции при различных уровнях фиксации тока
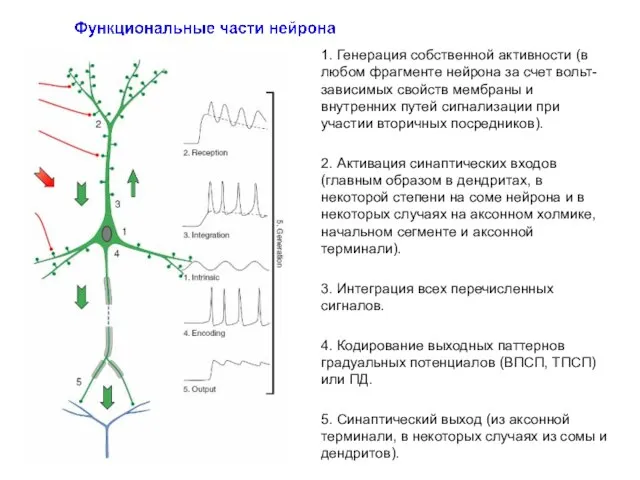
1. Генерация собственной активности (в любом фрагменте нейрона за счет вольт-зависимых
1. Генерация собственной активности (в любом фрагменте нейрона за счет вольт-зависимых
I
II
I
II

Fi = 1/∆ti
Fi = 1/∆ti
I
II
I
II

I
II
I
II


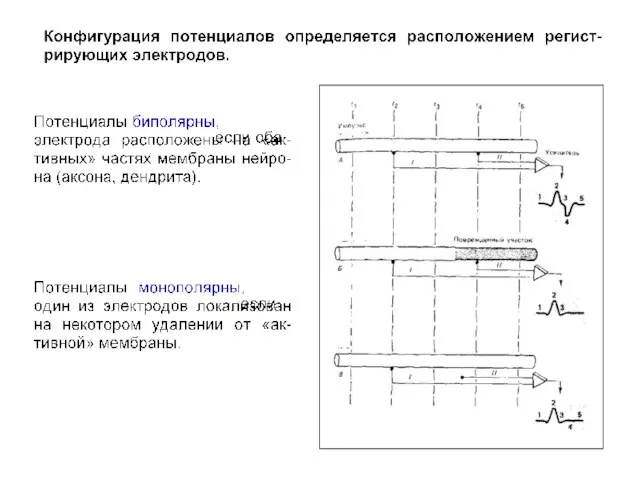
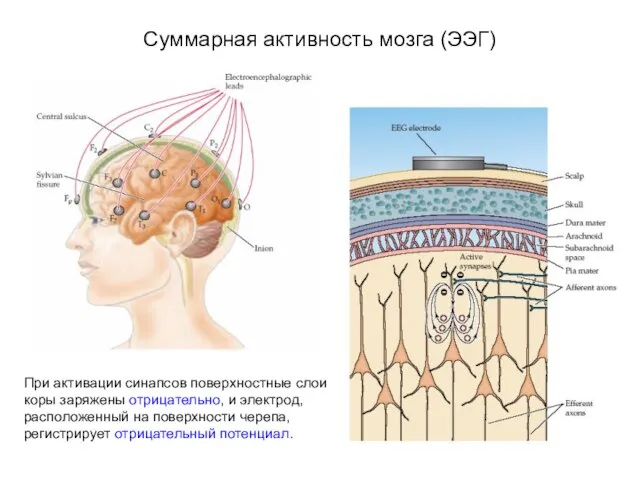
Суммарная активность мозга (ЭЭГ)
При активации синапсов поверхностные слои коры заряжены отрицательно,
Суммарная активность мозга (ЭЭГ)
При активации синапсов поверхностные слои коры заряжены отрицательно,
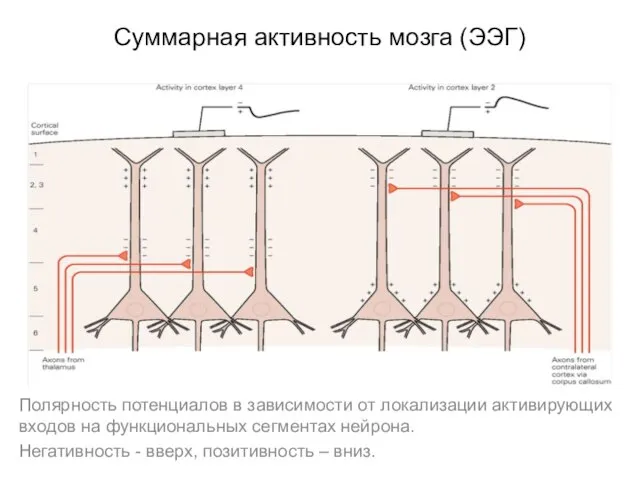
Суммарная активность мозга (ЭЭГ)
Полярность потенциалов в зависимости от локализации активирующих входов
Суммарная активность мозга (ЭЭГ)
Полярность потенциалов в зависимости от локализации активирующих входов
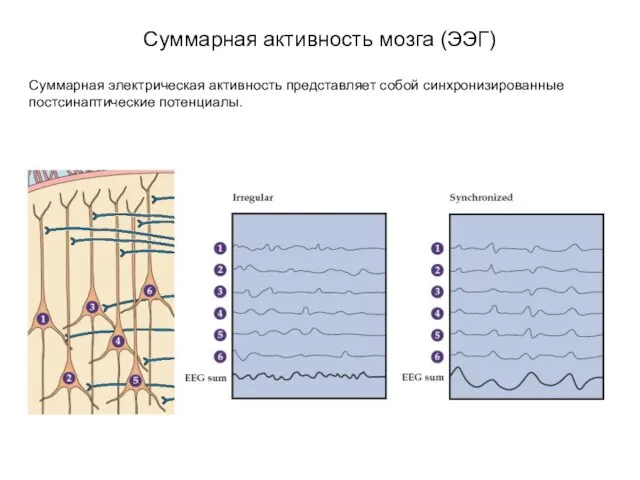
Суммарная активность мозга (ЭЭГ)
Суммарная электрическая активность представляет собой синхронизированные постсинаптические потенциалы.
Суммарная активность мозга (ЭЭГ)
Суммарная электрическая активность представляет собой синхронизированные постсинаптические потенциалы.
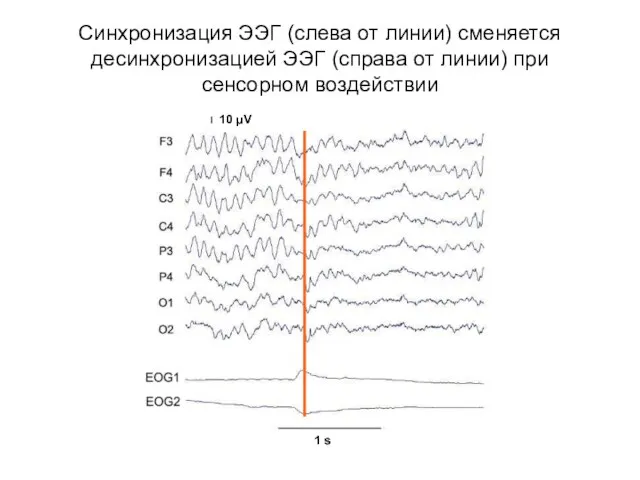
Синхронизация ЭЭГ (слева от линии) сменяется десинхронизацией ЭЭГ (справа от линии)
Синхронизация ЭЭГ (слева от линии) сменяется десинхронизацией ЭЭГ (справа от линии)


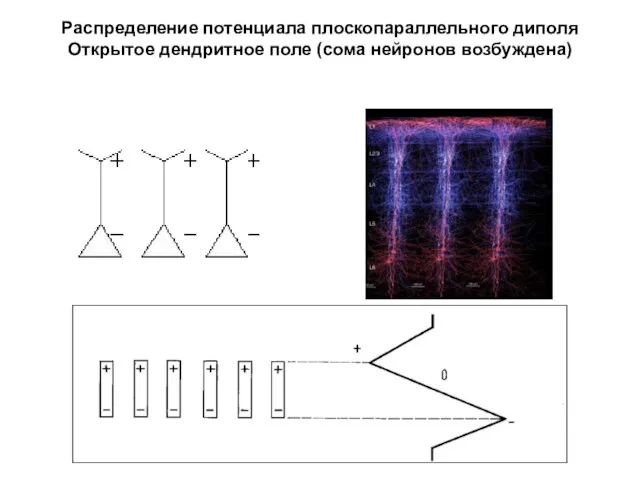
Распределение потенциала плоскопараллельного диполя
Открытое дендритное поле (сома нейронов возбуждена)
Распределение потенциала плоскопараллельного диполя
Открытое дендритное поле (сома нейронов возбуждена)
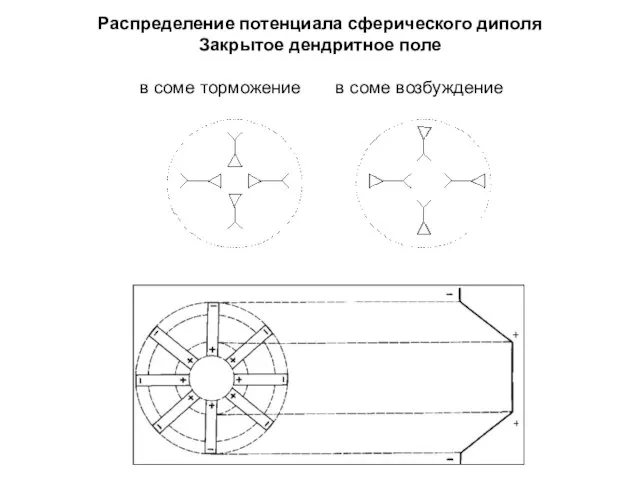
Распределение потенциала сферического диполя
Закрытое дендритное поле
в соме торможение в соме возбуждение
Распределение потенциала сферического диполя
Закрытое дендритное поле
в соме торможение в соме возбуждение
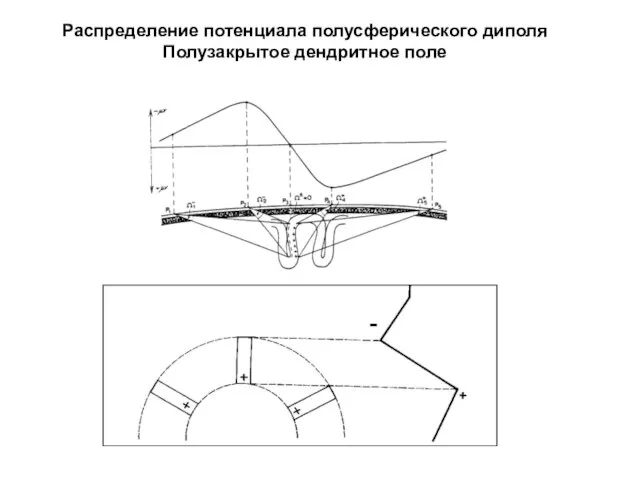
Распределение потенциала полусферического диполя
Полузакрытое дендритное поле
Распределение потенциала полусферического диполя
Полузакрытое дендритное поле
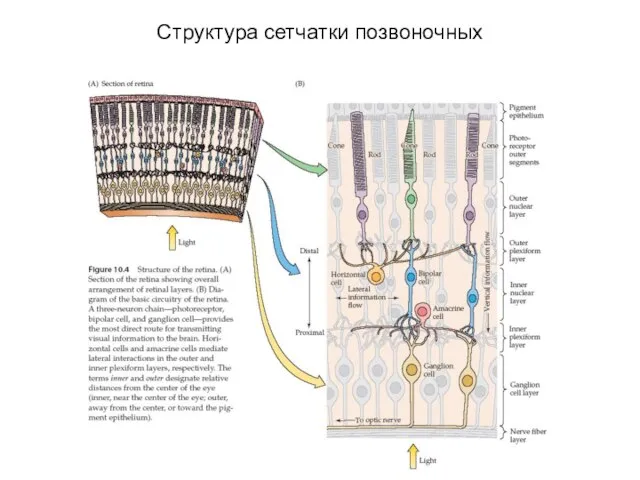
Структура сетчатки позвоночных
Структура сетчатки позвоночных
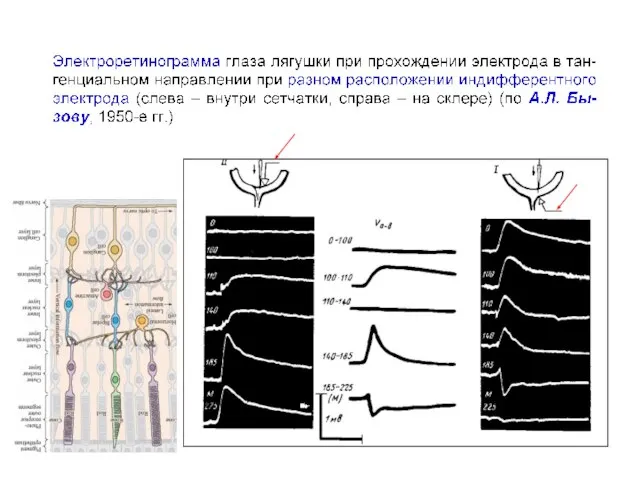

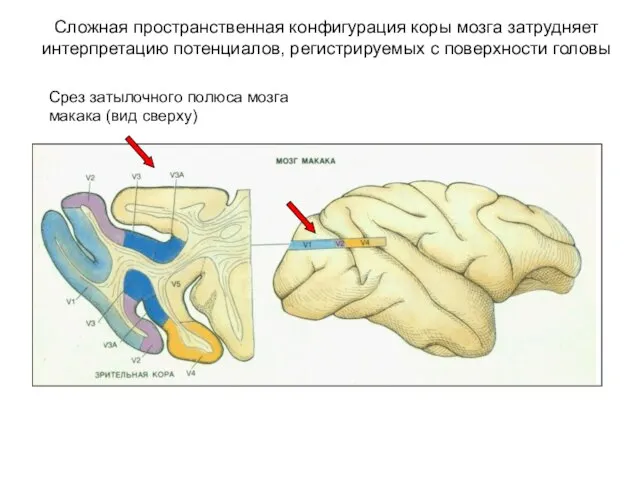
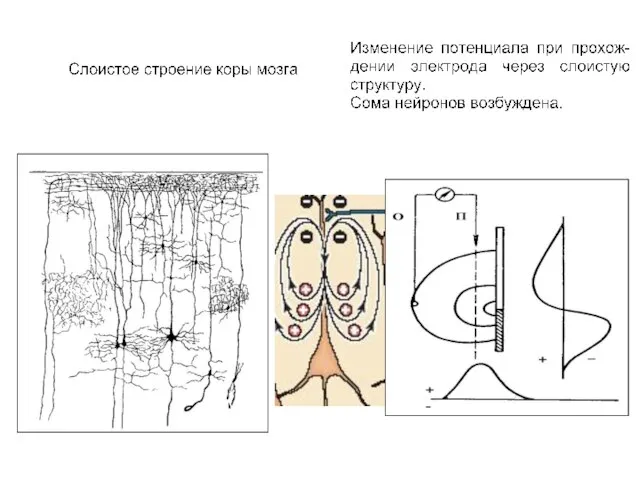
Сложная пространственная конфигурация коры мозга затрудняет интерпретацию потенциалов, регистрируемых с поверхности
Сложная пространственная конфигурация коры мозга затрудняет интерпретацию потенциалов, регистрируемых с поверхности
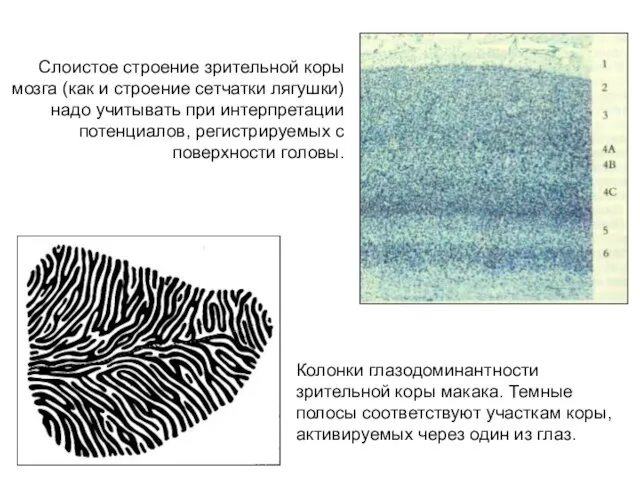
Слоистое строение зрительной коры мозга (как и строение сетчатки лягушки) надо
Слоистое строение зрительной коры мозга (как и строение сетчатки лягушки) надо
 Презентация на тему "Генетические опыты Менделя" - скачать бесплатно презентации по Биологии
Презентация на тему "Генетические опыты Менделя" - скачать бесплатно презентации по Биологии Жизнь хомячков в неволе
Жизнь хомячков в неволе Решу ОГЭ
Решу ОГЭ Аксолотль Земноводное животное
Аксолотль Земноводное животное  Происхождение растений. Основные этапы развития растительного мира
Происхождение растений. Основные этапы развития растительного мира Проптеридофиты. Моховидные. Псилотовидные. Плауновидные. Хвощевидные. Папоротниковидные
Проптеридофиты. Моховидные. Псилотовидные. Плауновидные. Хвощевидные. Папоротниковидные Филогенез скелета позвоночных животных
Филогенез скелета позвоночных животных Наследственность и окружающая среда
Наследственность и окружающая среда Лотос – цветок самый древний на земле 7 чудес Хабаровского края Выполнили: Уч-ся 4 класса МБОУ ООШ №1п.Ванино Прокопенко Наталь
Лотос – цветок самый древний на земле 7 чудес Хабаровского края Выполнили: Уч-ся 4 класса МБОУ ООШ №1п.Ванино Прокопенко Наталь Презентация на тему Жиры
Презентация на тему Жиры Передача наследственной информации от ДНК к и-РНК и к белку
Передача наследственной информации от ДНК к и-РНК и к белку Мутационная изменчивость
Мутационная изменчивость Реакции глюконеогенеза
Реакции глюконеогенеза Обмен и роль белков, жиров, углеводов
Обмен и роль белков, жиров, углеводов Жизненный цикл клетки. Митоз
Жизненный цикл клетки. Митоз Цветоведение
Цветоведение Мир животных
Мир животных Презентация на тему "Дрозофіла – модельний організм генетичних експериментів" - скачать бесплатно презентации по Биологии
Презентация на тему "Дрозофіла – модельний організм генетичних експериментів" - скачать бесплатно презентации по Биологии Человек как объект генетики. Методы изучения генетики человека. (Лекции 1-2)
Человек как объект генетики. Методы изучения генетики человека. (Лекции 1-2) Презентация Основные факторы неорганических соединений
Презентация Основные факторы неорганических соединений ТИП ЧЛЕНИСТОНОГИЕ
ТИП ЧЛЕНИСТОНОГИЕ Приспособленность живых организмов к среде обитания Творческое название: Способы выживания организмов
Приспособленность живых организмов к среде обитания Творческое название: Способы выживания организмов Цікаве про птахів Цікаве про птахів Птахи “жують” шлунком. Більшість птахів не мають зубів. Зазвичай вони ковтають гальку або гравій, які енергійно переміщуються в шлунку і сприяють перемелюванню їжі. Пташеняті може знадобитися два дні, щоб вибра
Цікаве про птахів Цікаве про птахів Птахи “жують” шлунком. Більшість птахів не мають зубів. Зазвичай вони ковтають гальку або гравій, які енергійно переміщуються в шлунку і сприяють перемелюванню їжі. Пташеняті може знадобитися два дні, щоб вибра Репродукция вирусов
Репродукция вирусов Выявление приспособлений организмов к среде обитания
Выявление приспособлений организмов к среде обитания Система кровообращения человека
Система кровообращения человека Презентация на тему "Как близки к нам биоматериалы «Аллоплант»" - скачать презентации по Биологии
Презентация на тему "Как близки к нам биоматериалы «Аллоплант»" - скачать презентации по Биологии Вопросы для олимпиады
Вопросы для олимпиады