- Еволюція міжклітинного сигналінгу

Содержание
- 2. Склад сигнальної системи
- 3. Загальні принципи еволюції сигналінгу
- 4. Еволюція механізмів сигналінгу Вторинні месенджери
- 5. 1 етап
- 6. Рання еволюція сигнальної системи Одними із перших стимулів, що одночасно являються і месенджерами могли бути йони
- 7. Іонофори Ще один варіант простих переносників йонів (лише за градієнтом). Наприклад, A23187 – частково специфічний до
- 8. Небілкові кальцієві канали у сучасних бактерій Полі-β-гідроксибутиратні канали відомі у Azotobacter vinelandii, Bacillus subtilis, Haemophilus influenzae,
- 9. Первинні системи сигналінгу без ампліфікації 1. Сигналінг із зовнішнім середовищем 2. Примітивний міжклітинний сигналиінг
- 10. Первинна кальцій-залежна система Кальцієва система зв’язана з іншою. Повноцінний (незалежний від середовища) кальцієвий сигналінг може виникнути
- 11. Можливі функції Груповий гідроліз субстрату. Примітивний хемотаксис за принципом роботи аналогічний прокаріотичному (є індуктор – є
- 12. 2 етап
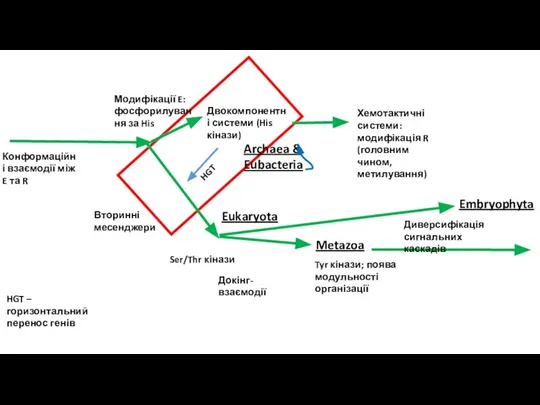
- 13. Поява ампліфікації Модифікації білків – > двокомпонентні системи. Транскрипційна регуляція. Вторинні месcенджери. Додатковий шлях у бактерій
- 14. Еволюція сайтів фосфорилування Першим кроком до вирішення проблеми необхідності ампліфікації сигналу була поява посттрансляційних модифікацій ефекторів
- 15. Двокомпонентні системи Kristin K. Koretke, Andrei N. Lupas, Patrick V. Warren, Martin Rosenberg, and James R.
- 16. Вторинні месенджери
- 17. Поява модифікації рецепторів – хемотаксис бактерій Регуляція за – зв’язком
- 18. Вторинні месенджери Циклічні нуклеотиди – найбільш розповсюджені як серед про- так і еукаріот. cAMP
- 19. 3 етап
- 20. Прокаріоти - + зв’язок
- 21. Регуляція позитивним зворотнім зв’язком Повноцінна ампліфікація у сигнальній системі, що активно підтримується груповим добором. Виникає у
- 22. Кворум сенсинг (відчуття кворуму) Найчастіше ефектор і синтаза автоіндуктора знаходяться в одному опероні. Кворум сенсинг у
- 23. Бувають і складніші системи з декількома автоіндукторами Кворум сенсинг у Pseudomonas aeruginosa
- 24. Еукаріоти – ускладнення рецепторів
- 25. Рецептори з ферментативною активністю Тирозин кіназний рецептор - Metazoa Серин-треонін кіназний рецептор – майже всі еукаріоти
- 26. G-білок залежні рецептори
- 27. Арешт рецепторів – негативна регуляція сигналу Регуляція за – зв’язком
- 28. Регуляторні мономерні ГТФази – «перемикачі»
- 29. Ускладнення регуляції Bhattacharyya et al., 2006
- 30. Диверсифікація сигнальних каскадів Збільшення різноманітності сигнальних каскадів – важлива передумова появи багатоклітинності. Наприклад, у Metazoa з’являються
- 31. Висновки Таким чином, еволюція сигнальних систем йшла в напрямку збільшення “еволюційної здатності” (evolvability) системи, кількості шляхів
- 33. Скачать презентацию

Склад сигнальної системи
Склад сигнальної системи

Загальні принципи еволюції сигналінгу
Загальні принципи еволюції сигналінгу

Еволюція механізмів сигналінгу
Вторинні месенджери
Еволюція механізмів сигналінгу
Вторинні месенджери

1 етап
1 етап

Рання еволюція сигнальної системи
Одними із перших стимулів, що одночасно являються і
Рання еволюція сигнальної системи
Одними із перших стимулів, що одночасно являються і

Іонофори
Ще один варіант простих переносників йонів (лише за градієнтом).
Наприклад, A23187 –
Іонофори
Ще один варіант простих переносників йонів (лише за градієнтом).
Наприклад, A23187 –

Небілкові кальцієві канали у сучасних бактерій
Полі-β-гідроксибутиратні канали відомі у Azotobacter vinelandii,
Небілкові кальцієві канали у сучасних бактерій
Полі-β-гідроксибутиратні канали відомі у Azotobacter vinelandii,
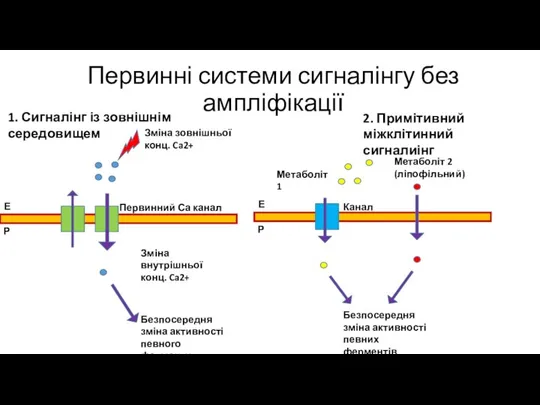
Первинні системи сигналінгу без ампліфікації
1. Сигналінг із зовнішнім середовищем
2. Примітивний міжклітинний
Первинні системи сигналінгу без ампліфікації
1. Сигналінг із зовнішнім середовищем
2. Примітивний міжклітинний

Первинна кальцій-залежна система
Кальцієва система зв’язана з іншою.
Повноцінний (незалежний від середовища) кальцієвий
Первинна кальцій-залежна система
Кальцієва система зв’язана з іншою.
Повноцінний (незалежний від середовища) кальцієвий

Можливі функції
Груповий гідроліз субстрату.
Примітивний хемотаксис за принципом роботи аналогічний прокаріотичному (є
Можливі функції
Груповий гідроліз субстрату.
Примітивний хемотаксис за принципом роботи аналогічний прокаріотичному (є

2 етап
2 етап

Поява ампліфікації
Модифікації білків – > двокомпонентні системи.
Транскрипційна регуляція.
Вторинні месcенджери.
Додатковий шлях у
Поява ампліфікації
Модифікації білків – > двокомпонентні системи.
Транскрипційна регуляція.
Вторинні месcенджери.
Додатковий шлях у

Еволюція сайтів фосфорилування
Першим кроком до вирішення проблеми необхідності ампліфікації сигналу була
Еволюція сайтів фосфорилування
Першим кроком до вирішення проблеми необхідності ампліфікації сигналу була

Двокомпонентні системи
Kristin K. Koretke, Andrei N. Lupas, Patrick V. Warren, Martin
Двокомпонентні системи
Kristin K. Koretke, Andrei N. Lupas, Patrick V. Warren, Martin
Вторинні месенджери
Вторинні месенджери

Поява модифікації рецепторів – хемотаксис бактерій
Регуляція за – зв’язком
Поява модифікації рецепторів – хемотаксис бактерій
Регуляція за – зв’язком
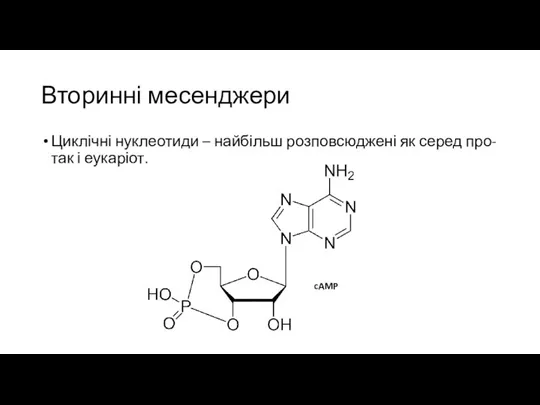
Вторинні месенджери
Циклічні нуклеотиди – найбільш розповсюджені як серед про- так і
Вторинні месенджери
Циклічні нуклеотиди – найбільш розповсюджені як серед про- так і

3 етап
3 етап

Прокаріоти - + зв’язок
Прокаріоти - + зв’язок

Регуляція позитивним зворотнім зв’язком
Повноцінна ампліфікація у сигнальній системі, що активно
Регуляція позитивним зворотнім зв’язком
Повноцінна ампліфікація у сигнальній системі, що активно

Кворум сенсинг (відчуття кворуму)
Найчастіше ефектор і синтаза автоіндуктора знаходяться в одному
Кворум сенсинг (відчуття кворуму)
Найчастіше ефектор і синтаза автоіндуктора знаходяться в одному

Бувають і складніші системи з декількома автоіндукторами
Кворум сенсинг у Pseudomonas aeruginosa
Бувають і складніші системи з декількома автоіндукторами
Кворум сенсинг у Pseudomonas aeruginosa

Еукаріоти – ускладнення рецепторів
Еукаріоти – ускладнення рецепторів

Рецептори з ферментативною активністю
Тирозин кіназний рецептор - Metazoa
Серин-треонін кіназний рецептор –
Рецептори з ферментативною активністю
Тирозин кіназний рецептор - Metazoa
Серин-треонін кіназний рецептор –

G-білок залежні рецептори
G-білок залежні рецептори

Арешт рецепторів – негативна регуляція сигналу
Регуляція за – зв’язком
Арешт рецепторів – негативна регуляція сигналу
Регуляція за – зв’язком

Регуляторні мономерні ГТФази – «перемикачі»
Регуляторні мономерні ГТФази – «перемикачі»
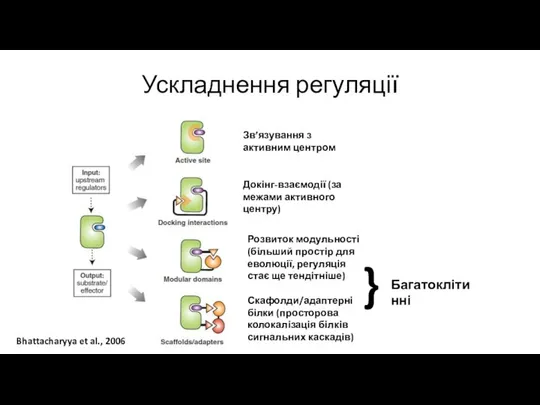
Ускладнення регуляції
Bhattacharyya et al., 2006
Ускладнення регуляції
Bhattacharyya et al., 2006

Диверсифікація сигнальних каскадів
Збільшення різноманітності сигнальних каскадів – важлива передумова появи багатоклітинності.
Наприклад,
Диверсифікація сигнальних каскадів
Збільшення різноманітності сигнальних каскадів – важлива передумова появи багатоклітинності.
Наприклад,

Висновки
Таким чином, еволюція сигнальних систем йшла в напрямку збільшення “еволюційної здатності”
Висновки
Таким чином, еволюція сигнальних систем йшла в напрямку збільшення “еволюційної здатності”
 Центральная нервная система. Спинной мозг
Центральная нервная система. Спинной мозг Общая характеристика бурых водорослей
Общая характеристика бурых водорослей Системы теплообмена в биореакторах. (Лекция 6)
Системы теплообмена в биореакторах. (Лекция 6) Презентация на тему "Селекция микроорганизмов. Биотехнология 9 класс" - скачать презентации по Биологии
Презентация на тему "Селекция микроорганизмов. Биотехнология 9 класс" - скачать презентации по Биологии Хищные растения
Хищные растения Рослини-мандрівники
Рослини-мандрівники Вода и питьевой режим
Вода и питьевой режим Презентация на тему "Значение корней в жизни растений" - скачать презентации по Биологии
Презентация на тему "Значение корней в жизни растений" - скачать презентации по Биологии Рыжие и чёрные тараканы
Рыжие и чёрные тараканы Морж
Морж Интегративная деятельность мозга. Циклы сон-бодрствование
Интегративная деятельность мозга. Циклы сон-бодрствование Гендік маркерлерді өсімдіктер селекциясында қолдану
Гендік маркерлерді өсімдіктер селекциясында қолдану Презентация на тему "Каменный лес" - скачать презентации по Биологии
Презентация на тему "Каменный лес" - скачать презентации по Биологии Зоология – наука о животных 7 класс. 1 урок
Зоология – наука о животных 7 класс. 1 урок 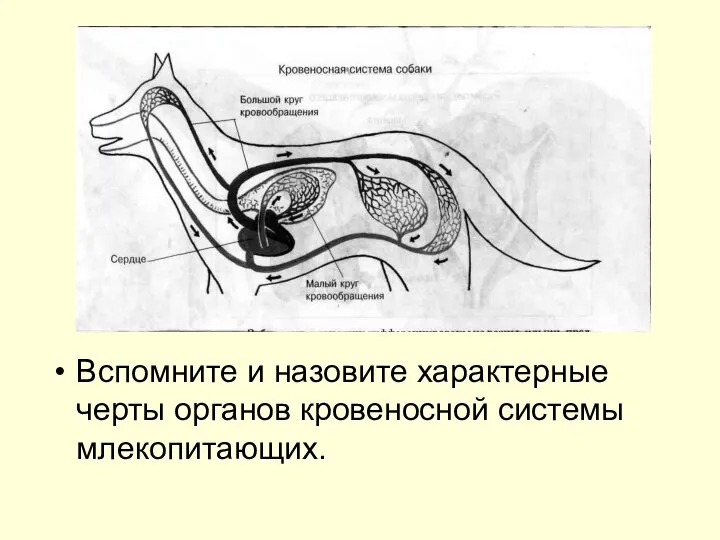 Вспомните и назовите характерные черты органов кровеносной системы млекопитающих.
Вспомните и назовите характерные черты органов кровеносной системы млекопитающих. Стійкість до важких металів
Стійкість до важких металів Исследование активности аланинаминотрансферазы и аспартатаминотрансферазы
Исследование активности аланинаминотрансферазы и аспартатаминотрансферазы Органические вещества живых организмов
Органические вещества живых организмов Уголок природы дошкольников. Цветы
Уголок природы дошкольников. Цветы Ткани животных
Ткани животных Соединительная ткань. Классификация соединительной ткани
Соединительная ткань. Классификация соединительной ткани ДРЕВО ХОРДОВЫХ Лекции по зоологии позвоночных
ДРЕВО ХОРДОВЫХ Лекции по зоологии позвоночных  Презентация Креационизм
Презентация Креационизм Презентация на тему "Классификация углеводов" - скачать презентации по Биологии
Презентация на тему "Классификация углеводов" - скачать презентации по Биологии Общая анатомия развитие скелета. Кость как орган
Общая анатомия развитие скелета. Кость как орган Световые явления в живой и неживой природе
Световые явления в живой и неживой природе Жестоколистные и вечнозеленые леса и кустарники
Жестоколистные и вечнозеленые леса и кустарники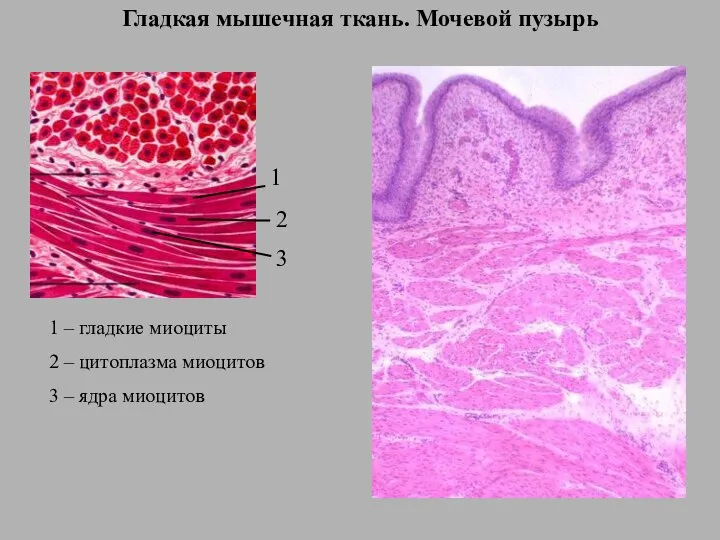 Гладкая мышечная ткань. Мочевой пузырь
Гладкая мышечная ткань. Мочевой пузырь