- Физиология человека и животных

Содержание
- 2. Раздел ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ Раздражимость. Раздражители. Классификация раздражителей Возбудимые ткани Возбудимость и возбуждение. Общие свойства возбудимых
- 3. Основные понятия и термины «Физиологии возбудимых тканей» Раздражимость Все живые клетки обладают раздражимостью, т. е. способностью
- 4. По своей природе 1) химические; 2) физические; 3) механические; 4) термические; 5) биологические. По биологическому соответствию
- 5. Понятие о состоянии покоя и активности возбудимых тканей Состояние покоя в возбудимых тканях -на ткань не
- 6. Возбудимость – это способность возбудимых тканей реагировать на раздражители генерацией процесса возбуждения (др. словами потенциалом действия
- 7. Возбуждение – это проявление свойства тканей - возбудимости Общие признаки возбуждения: 1) изменение уровня обменных процессов,
- 9. Возбудимые ткани: нервная мышечная железистая
- 10. Противоположное возбуждению состояние возбудимых тканей называется торможением. Торможение – это активное состояние возбудимых тканей, сопровождающееся специфическими
- 13. Биоэлектрические явления в живых тканях. История вопроса Луиджи Гальвани (Luigi Aloisio Galvani, 9 сентября 1737 —
- 14. Открытием Гальвани заинтересовались и физики и врачи. Среди физиков был соотечественник Гальвани Алессандро Вольта (1745 -
- 15. Маттеуччи Карло (1811-1868) — итал. физик, профессор в Болонье, Равенне и Пизе, особенно много работал по
- 16. Мембранно-ионная теория возбуждения
- 17. Ионные каналы: Специфические – селективные (натриевые, калиевые, кальциевые, хлорные) - открываются и закрываются в ответ на
- 18. ПРИРОДА ПОТЕНЦИАЛА ПОКОЯ В 1896 г. В. Ю. Чаговец высказал гипотезу об ионном механизме электрических потенциалов
- 19. ОСНОВНЫЕ ПОЛОЖЕНИЯ МЕМБРАННО-ИОННОЙ ТЕОРИИ Бернштейн, 1902 1.В состоянии покоя клеточная мембрана хорошо проницаема для К+, менее
- 21. Природа потенциала покоя . Электрохимический потенциал К+ KCL 0,1 м KCL 0,1 м К+ ТОК=0
- 22. Электрохимический потенциал К+ KCL 0,1 м KCL 0,01 м К+ ТОК + + + + +
- 23. ПОЛЯРИЗАЦИЯ МЕМБРАНЫ К+ KCL 0,1 м KCL 0,01 м + + - - - - -
- 24. ПРИРОДА ПОТЕНЦИАЛА ПОКОЯ Чтобы понять, каким образом возникает этот потенциал, рассмотрим следующий модельный опыт. Представим сосуд,
- 25. Равновесный потенциал – это такой потенциал на мембране, который уравновешивает химический концентрационный градиент для данного иона
- 26. Потенциал покоя
- 28. ПРИРОДА ПОТЕНЦИАЛА ПОКОЯ Предположение о том, что в состоянии покоя мембрана нервных и мышечных покоя, было
- 30. РОЛЬ ОБМЕНА ВЕЩЕСТВ В ГЕНЕЗЕ И ПОДДЕРЖАНИИ ПОТЕНЦИАЛА ПОКОЯ (НАТРИЕВЫЙ НАСОС МЕМБРАНЫ) Несмотря на то что
- 31. ПОТЕНЦИАЛ ДЕЙСТВИЯ (ПД) ПД - называют быстрое колебание МП, возникающее при возбуждении нервных, мышечных и некоторых
- 32. ЗНАЧЕНИЕ ПОТЕНЦИАЛОВ ДЕЙСТВИЯ (ПД) Распространение потенциалов действия по нервным волокнам обеспечивает передачу информации в нервной системе.
- 33. ПОТЕНЦИАЛ ДЕЙСТВИЯ. ЛОКАЛЬНЫЙ ОТВЕТ
- 34. ДЕПОЛЯРИЗАЦИЯ: -70 -50, -70 -25, -70 +30 ГИПЕРПОЛЯРИЗАЦИЯ: - 70 - 100 РЕПОЛЯРИЗАЦИЯ – ВОЗВРАЩЕНИЕ ПОТЕНЦИАЛА
- 35. Потенциал действия (ПД) Фазы: Деполяризация Реполяризация Следовые потенциалы: следовая гиперполяризация следовая деполяризация Различают восходящую (деполяризация) и
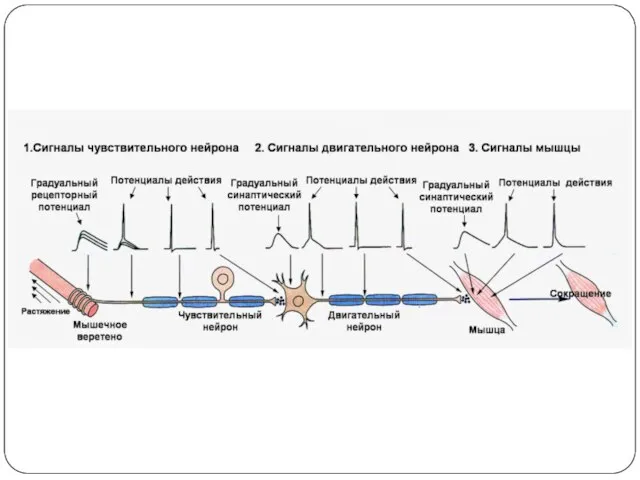
- 36. Потенциал действия. Ионный механизм возникновения ПД В основе ПД лежат последовательно развивающиеся во времени изменения ионной
- 37. Изменение проницаемости мембраны для различных ионов во время ПД
- 38. О ПРИРОДЕ ИОННОЙ ПРОНИЦАЕМОСТИ МЕМБРАНЫ. ИОННЫЕ КАНАЛЫ В основе рассмотренных изменений ионной проницаемости мембраны при генерации
- 39. Состояние ионных каналов в разные фазы потенциала действия Na+ каналы К+ каналы
- 40. МЕХАНИЗМЫ ИЗМЕНЕНИЯ ИОННОЙ ПРОВОДИМОСТИ ВО ВРЕМЯ ГЕНЕРАЦИИ ПОТЕНЦИАЛА ДЕЙСТВИЯ Таким образом, восходящая фаза ПД связана с
- 41. Критический уровень деполяризации Регистрация изменений МП при внутриклеточном раздражении нервного или мышечного волокна показала, что ПД
- 42. Локальный ответ (ЛО) В механизме критической деполяризации мембраны наряду с пассивными существенную роль играют активные подпороговые
- 43. Локальный ответ: Стимулы, величина которых меньше определённого порога, вызывают локальные ответы, затухающие вблизи места раздражения. ЛО
- 44. Изменение возбудимости в фазы потенциала действия Период повышенной возбудимости (ЛО) Фаза абсолютной рефрактерности (пик) Фаза относительной
- 45. ИЗМЕНЕНИЯ ВОЗБУДИМОСТИ ПРИ ВОЗБУЖДЕНИИ Используя тестирующие стимулы в различные фазы развития ПД, можно проследить временной ход
- 46. Рефрактерность Снижение способности клетки отвечать на раздражение в результате временной инактивации натриевых каналов Абсолютная рефрактерность –
- 47. Законы распространения возбуждения по нервному волокну Закон анатомической и физиологической целостности Закон изолированного проведения возбуждения Закон
- 48. Проведение возбуждения вдоль нервных и мышечных волокон осуществляется при помощи так называемых местных токов (Герман, 1899),
- 50. Распространение возбуждения по нервному волокну
- 51. Законы раздражения 1. Закон силы Чтобы возникло возбуждение, раздражитель должен быть достаточно сильным - пороговым или
- 53. Скачать презентацию

Раздел
ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ
Раздражимость. Раздражители. Классификация раздражителей
Возбудимые ткани
Возбудимость и возбуждение.
Общие свойства возбудимых
Раздел
ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ
Раздражимость. Раздражители. Классификация раздражителей
Возбудимые ткани
Возбудимость и возбуждение.
Общие свойства возбудимых

Основные понятия и термины
«Физиологии возбудимых тканей»
Раздражимость
Все живые клетки обладают раздражимостью,
Основные понятия и термины
«Физиологии возбудимых тканей»
Раздражимость
Все живые клетки обладают раздражимостью,

По своей природе
1) химические;
2) физические;
3) механические;
4) термические;
По своей природе
1) химические;
2) физические;
3) механические;
4) термические;

Понятие о состоянии покоя и активности возбудимых тканей
Состояние покоя в
Понятие о состоянии покоя и активности возбудимых тканей
Состояние покоя в

Возбудимость – это способность возбудимых тканей реагировать на раздражители генерацией процесса
Возбудимость – это способность возбудимых тканей реагировать на раздражители генерацией процесса

Возбуждение – это проявление свойства тканей - возбудимости
Общие признаки возбуждения:
1)
Возбуждение – это проявление свойства тканей - возбудимости
Общие признаки возбуждения:
1)
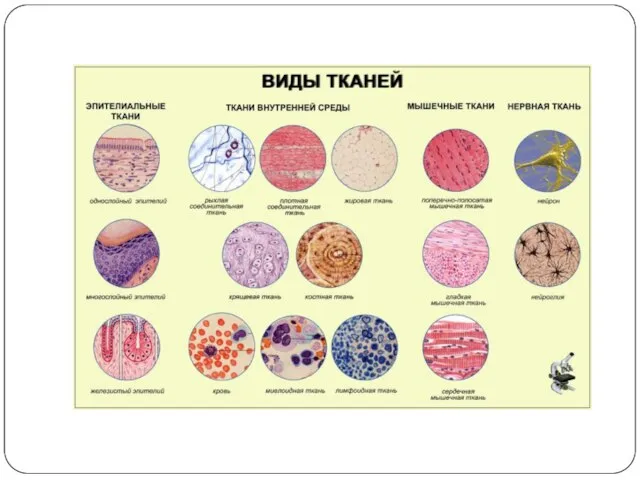
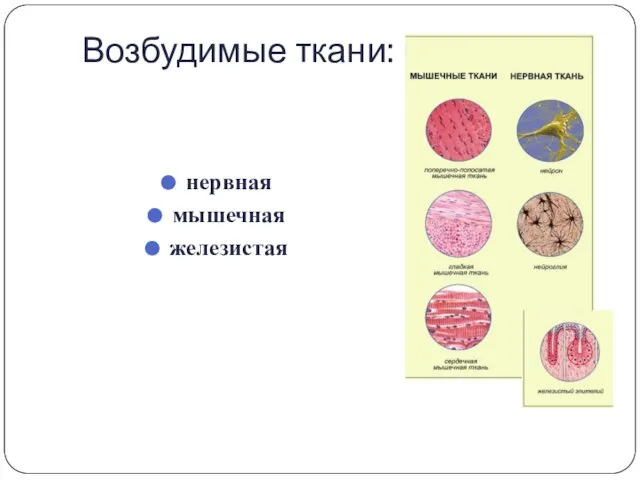
Возбудимые ткани:
нервная
мышечная
железистая
Возбудимые ткани:
нервная
мышечная
железистая

Противоположное возбуждению состояние возбудимых тканей называется торможением.
Торможение – это активное
Противоположное возбуждению состояние возбудимых тканей называется торможением.
Торможение – это активное


Биоэлектрические явления в живых тканях.
История вопроса
Луиджи Гальвани (Luigi Aloisio Galvani,
Биоэлектрические явления в живых тканях.
История вопроса
Луиджи Гальвани (Luigi Aloisio Galvani,

Открытием Гальвани заинтересовались и физики и врачи.
Среди физиков был соотечественник
Открытием Гальвани заинтересовались и физики и врачи.
Среди физиков был соотечественник

Маттеуччи Карло (1811-1868) — итал. физик, профессор в Болонье, Равенне и
Маттеуччи Карло (1811-1868) — итал. физик, профессор в Болонье, Равенне и
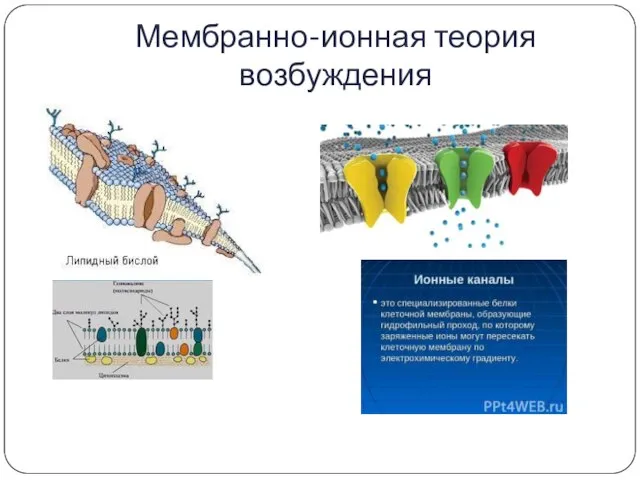
Мембранно-ионная теория возбуждения
Мембранно-ионная теория возбуждения

Ионные каналы:
Специфические – селективные (натриевые, калиевые, кальциевые, хлорные) -
открываются и
Ионные каналы:
Специфические – селективные (натриевые, калиевые, кальциевые, хлорные) -
открываются и

ПРИРОДА ПОТЕНЦИАЛА ПОКОЯ
В 1896 г. В. Ю. Чаговец высказал гипотезу об
ПРИРОДА ПОТЕНЦИАЛА ПОКОЯ
В 1896 г. В. Ю. Чаговец высказал гипотезу об

ОСНОВНЫЕ ПОЛОЖЕНИЯ МЕМБРАННО-ИОННОЙ ТЕОРИИ
Бернштейн, 1902
1.В состоянии покоя клеточная мембрана хорошо
ОСНОВНЫЕ ПОЛОЖЕНИЯ МЕМБРАННО-ИОННОЙ ТЕОРИИ
Бернштейн, 1902
1.В состоянии покоя клеточная мембрана хорошо

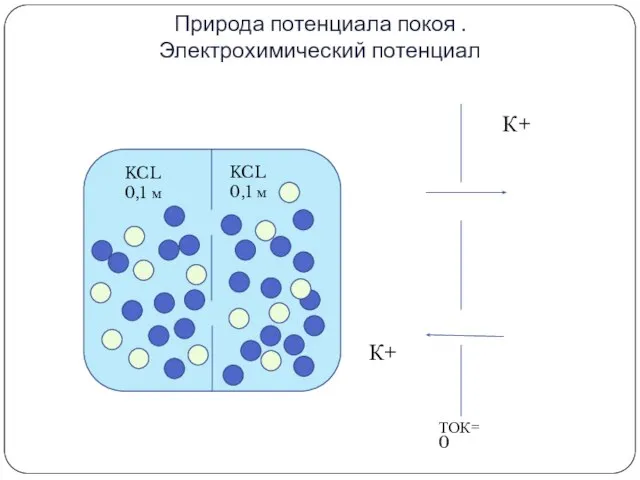
Природа потенциала покоя .
Электрохимический потенциал
К+
KCL
0,1 м
KCL
0,1 м
К+
ТОК=0
Природа потенциала покоя .
Электрохимический потенциал
К+
KCL
0,1 м
KCL
0,1 м
К+
ТОК=0
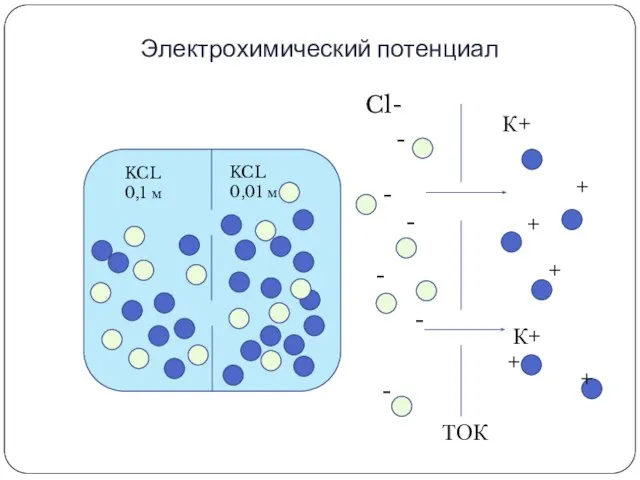
Электрохимический потенциал
К+
KCL
0,1 м
KCL
0,01 м
К+
ТОК
+
+
+
+
+
-
-
-
-
-
-
Cl-
Электрохимический потенциал
К+
KCL
0,1 м
KCL
0,01 м
К+
ТОК
+
+
+
+
+
-
-
-
-
-
-
Cl-
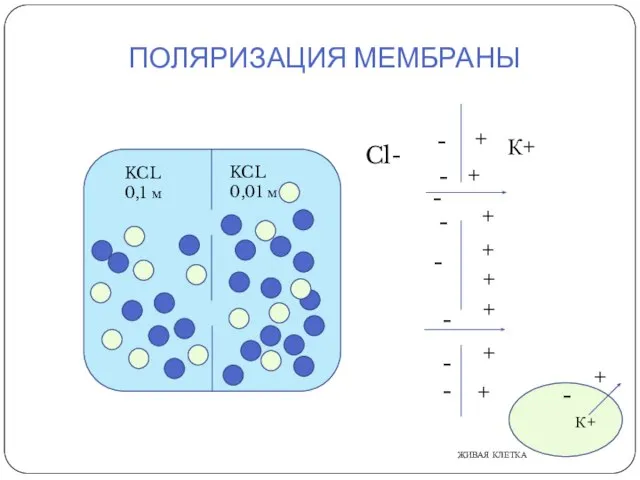
ПОЛЯРИЗАЦИЯ МЕМБРАНЫ
К+
KCL
0,1 м
KCL
0,01 м
+
+
-
-
-
-
-
Cl-
-
-
-
+
+
+
+
+
+
-
+
К+
ЖИВАЯ КЛЕТКА
ПОЛЯРИЗАЦИЯ МЕМБРАНЫ
К+
KCL
0,1 м
KCL
0,01 м
+
+
-
-
-
-
-
Cl-
-
-
-
+
+
+
+
+
+
-
+
К+
ЖИВАЯ КЛЕТКА
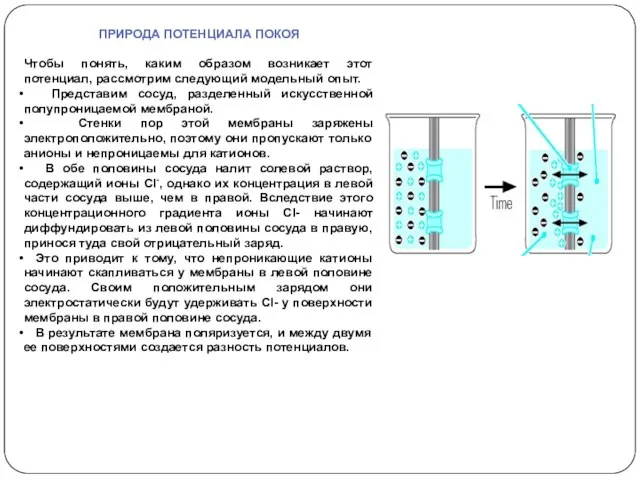
ПРИРОДА ПОТЕНЦИАЛА ПОКОЯ
Чтобы понять, каким образом возникает этот потенциал, рассмотрим следующий
ПРИРОДА ПОТЕНЦИАЛА ПОКОЯ
Чтобы понять, каким образом возникает этот потенциал, рассмотрим следующий
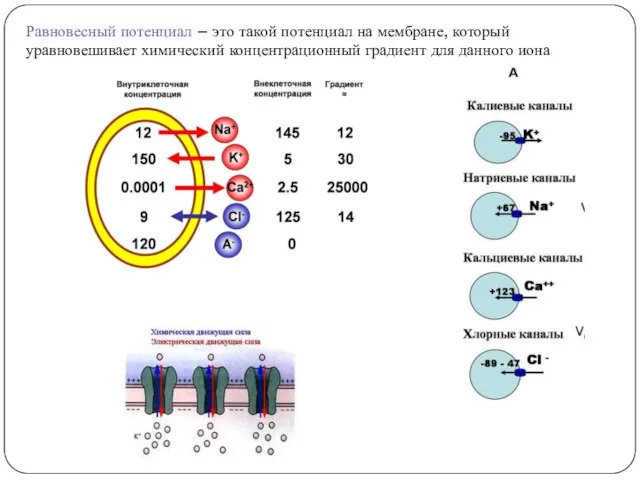
Равновесный потенциал – это такой потенциал на мембране, который уравновешивает химический
Равновесный потенциал – это такой потенциал на мембране, который уравновешивает химический
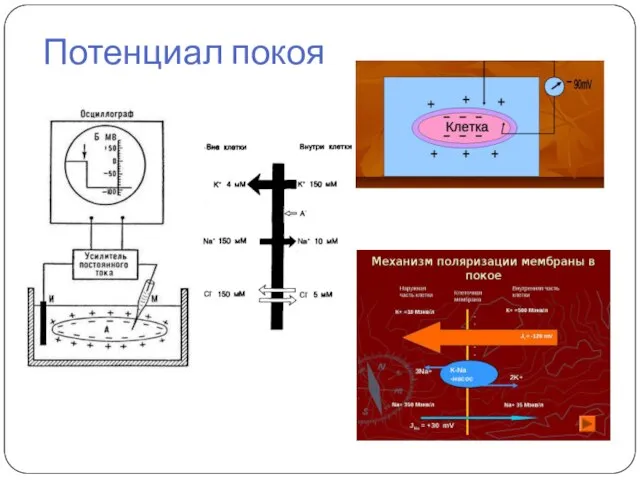
Потенциал покоя
Потенциал покоя
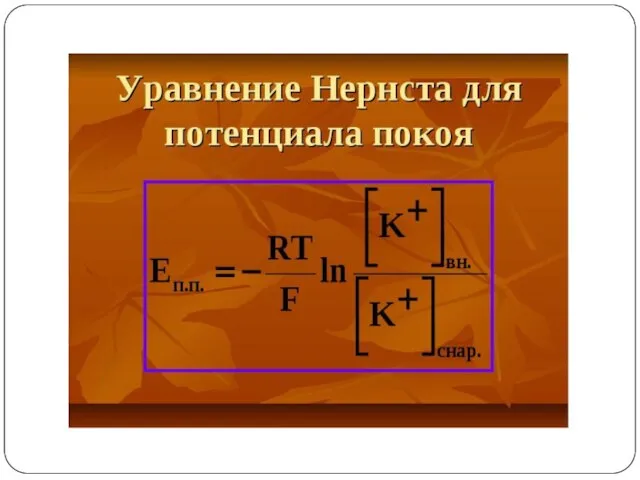

ПРИРОДА ПОТЕНЦИАЛА ПОКОЯ
Предположение о том, что в состоянии покоя мембрана
ПРИРОДА ПОТЕНЦИАЛА ПОКОЯ
Предположение о том, что в состоянии покоя мембрана


РОЛЬ ОБМЕНА ВЕЩЕСТВ В ГЕНЕЗЕ И ПОДДЕРЖАНИИ ПОТЕНЦИАЛА ПОКОЯ (НАТРИЕВЫЙ НАСОС
РОЛЬ ОБМЕНА ВЕЩЕСТВ В ГЕНЕЗЕ И ПОДДЕРЖАНИИ ПОТЕНЦИАЛА ПОКОЯ (НАТРИЕВЫЙ НАСОС

ПОТЕНЦИАЛ ДЕЙСТВИЯ (ПД)
ПД - называют быстрое колебание МП, возникающее при
ПОТЕНЦИАЛ ДЕЙСТВИЯ (ПД)
ПД - называют быстрое колебание МП, возникающее при

ЗНАЧЕНИЕ ПОТЕНЦИАЛОВ ДЕЙСТВИЯ (ПД)
Распространение потенциалов действия по нервным волокнам обеспечивает передачу
ЗНАЧЕНИЕ ПОТЕНЦИАЛОВ ДЕЙСТВИЯ (ПД)
Распространение потенциалов действия по нервным волокнам обеспечивает передачу
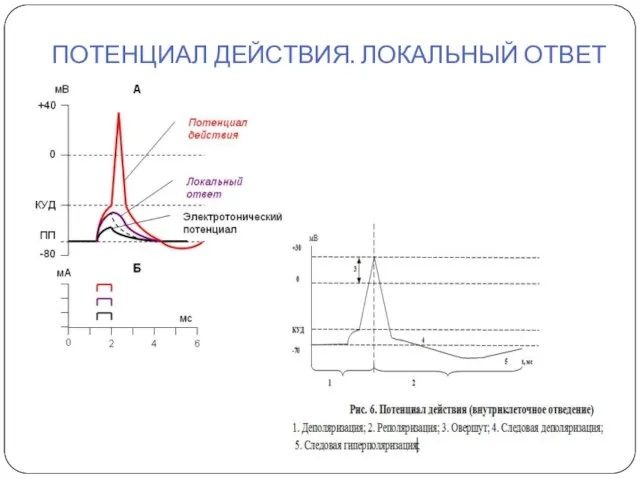
ПОТЕНЦИАЛ ДЕЙСТВИЯ. ЛОКАЛЬНЫЙ ОТВЕТ
ПОТЕНЦИАЛ ДЕЙСТВИЯ. ЛОКАЛЬНЫЙ ОТВЕТ
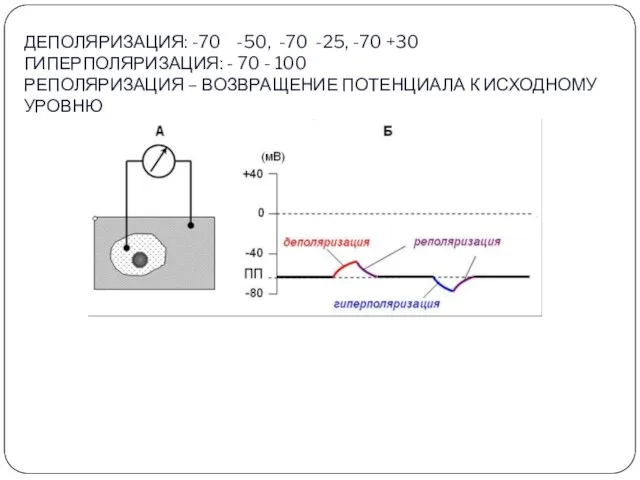
ДЕПОЛЯРИЗАЦИЯ: -70 -50, -70 -25, -70 +30
ГИПЕРПОЛЯРИЗАЦИЯ: - 70 - 100
РЕПОЛЯРИЗАЦИЯ
ДЕПОЛЯРИЗАЦИЯ: -70 -50, -70 -25, -70 +30 ГИПЕРПОЛЯРИЗАЦИЯ: - 70 - 100 РЕПОЛЯРИЗАЦИЯ
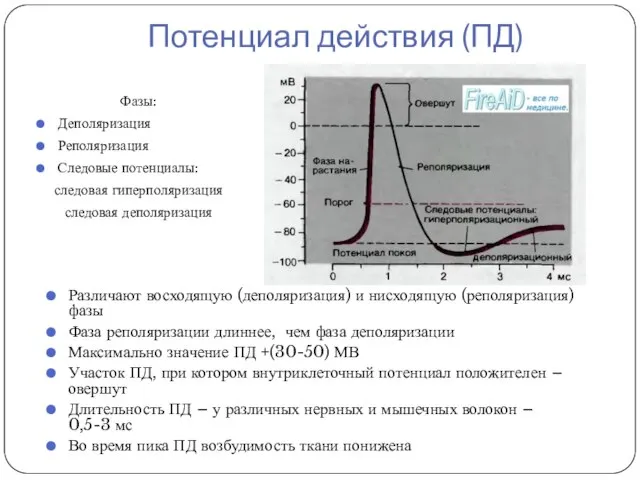
Потенциал действия (ПД)
Фазы:
Деполяризация
Реполяризация
Следовые потенциалы:
следовая гиперполяризация
следовая деполяризация
Различают восходящую (деполяризация) и нисходящую
Потенциал действия (ПД)
Фазы:
Деполяризация
Реполяризация
Следовые потенциалы:
следовая гиперполяризация
следовая деполяризация
Различают восходящую (деполяризация) и нисходящую

Потенциал действия. Ионный механизм возникновения ПД
В основе ПД лежат последовательно развивающиеся
Потенциал действия. Ионный механизм возникновения ПД
В основе ПД лежат последовательно развивающиеся
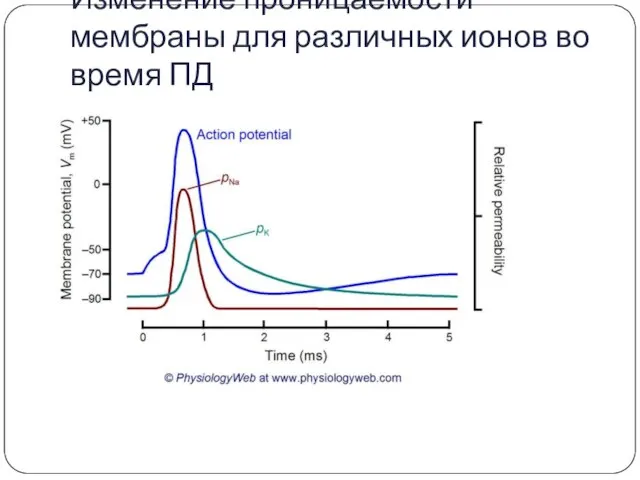
Изменение проницаемости мембраны для различных ионов во время ПД
Изменение проницаемости мембраны для различных ионов во время ПД
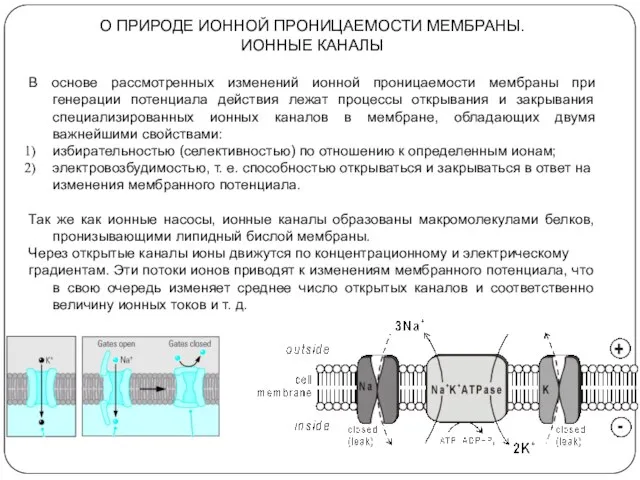
О ПРИРОДЕ ИОННОЙ ПРОНИЦАЕМОСТИ МЕМБРАНЫ.
ИОННЫЕ КАНАЛЫ
В основе рассмотренных изменений ионной
О ПРИРОДЕ ИОННОЙ ПРОНИЦАЕМОСТИ МЕМБРАНЫ.
ИОННЫЕ КАНАЛЫ
В основе рассмотренных изменений ионной
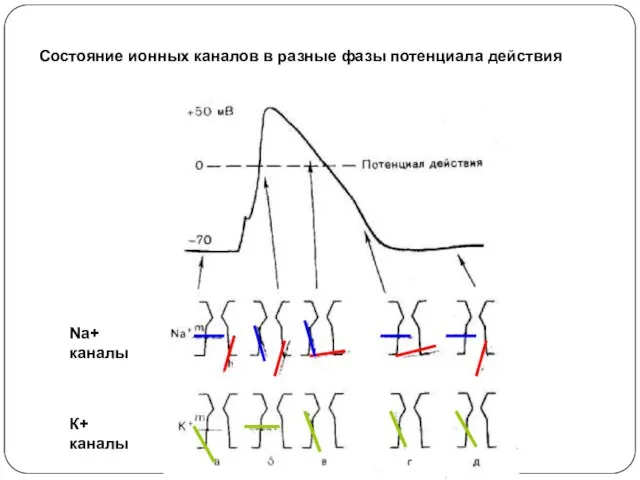
Состояние ионных каналов в разные фазы потенциала действия
Na+ каналы
К+ каналы
Состояние ионных каналов в разные фазы потенциала действия
Na+ каналы
К+ каналы

МЕХАНИЗМЫ ИЗМЕНЕНИЯ ИОННОЙ ПРОВОДИМОСТИ
ВО ВРЕМЯ ГЕНЕРАЦИИ ПОТЕНЦИАЛА ДЕЙСТВИЯ
Таким образом, восходящая фаза
МЕХАНИЗМЫ ИЗМЕНЕНИЯ ИОННОЙ ПРОВОДИМОСТИ
ВО ВРЕМЯ ГЕНЕРАЦИИ ПОТЕНЦИАЛА ДЕЙСТВИЯ
Таким образом, восходящая фаза
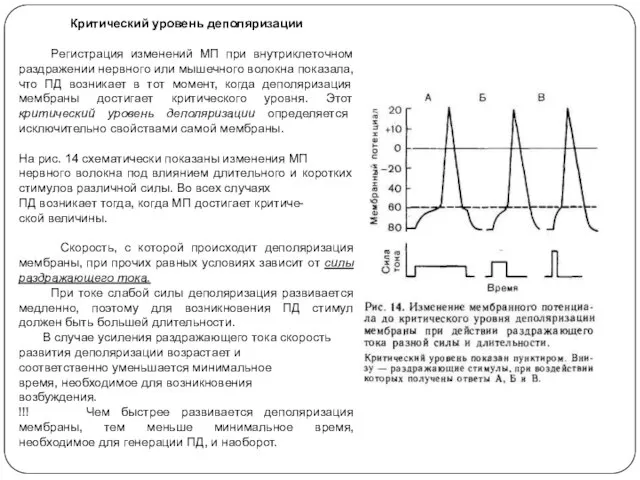
Критический уровень деполяризации
Регистрация изменений МП при внутриклеточном раздражении нервного или
Критический уровень деполяризации
Регистрация изменений МП при внутриклеточном раздражении нервного или
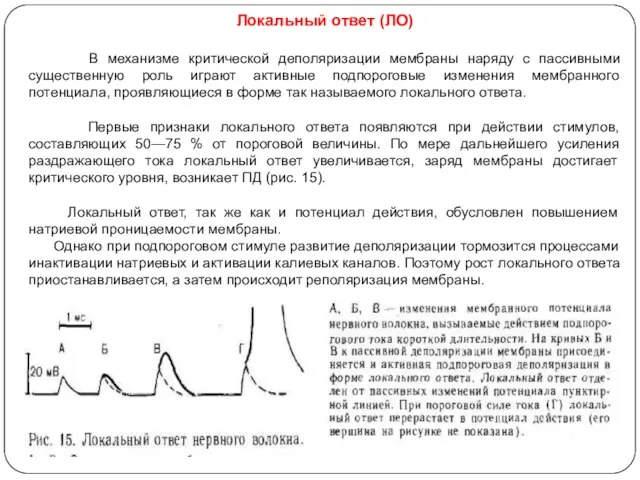
Локальный ответ (ЛО)
В механизме критической деполяризации мембраны наряду с пассивными
Локальный ответ (ЛО)
В механизме критической деполяризации мембраны наряду с пассивными
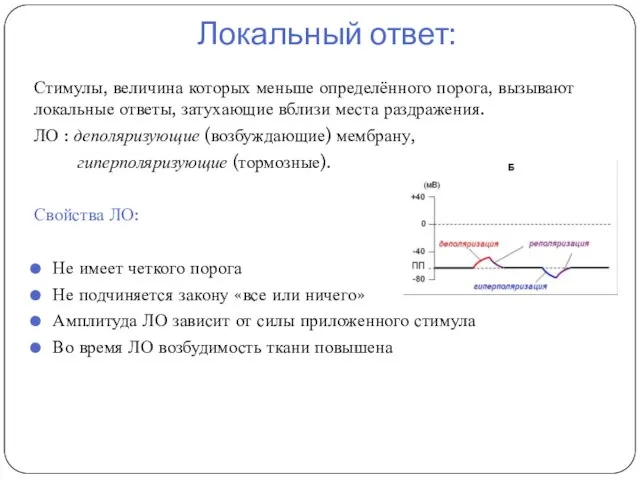
Локальный ответ:
Стимулы, величина которых меньше определённого порога, вызывают локальные ответы, затухающие
Локальный ответ:
Стимулы, величина которых меньше определённого порога, вызывают локальные ответы, затухающие
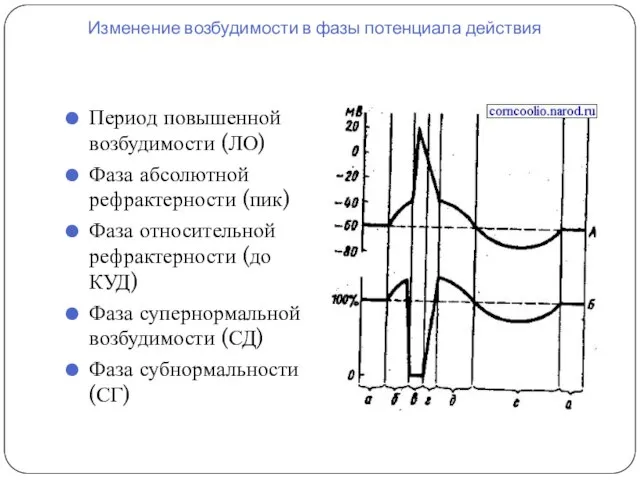
Изменение возбудимости в фазы потенциала действия
Период повышенной возбудимости (ЛО)
Фаза абсолютной рефрактерности
Изменение возбудимости в фазы потенциала действия
Период повышенной возбудимости (ЛО)
Фаза абсолютной рефрактерности

ИЗМЕНЕНИЯ ВОЗБУДИМОСТИ ПРИ ВОЗБУЖДЕНИИ
Используя тестирующие стимулы в различные фазы развития ПД,
ИЗМЕНЕНИЯ ВОЗБУДИМОСТИ ПРИ ВОЗБУЖДЕНИИ
Используя тестирующие стимулы в различные фазы развития ПД,
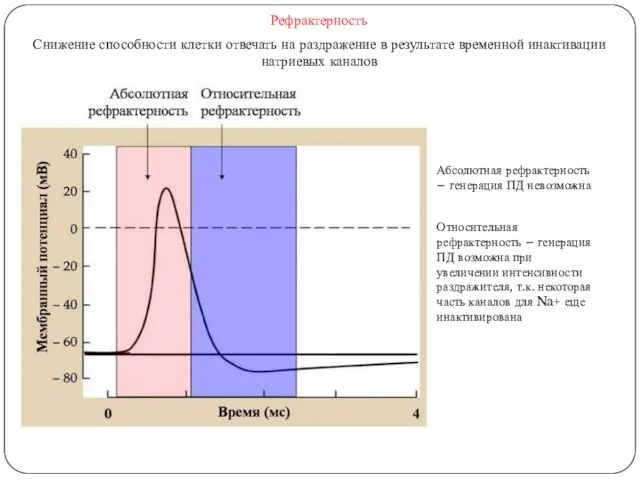
Рефрактерность
Снижение способности клетки отвечать на раздражение в результате временной инактивации
Рефрактерность
Снижение способности клетки отвечать на раздражение в результате временной инактивации
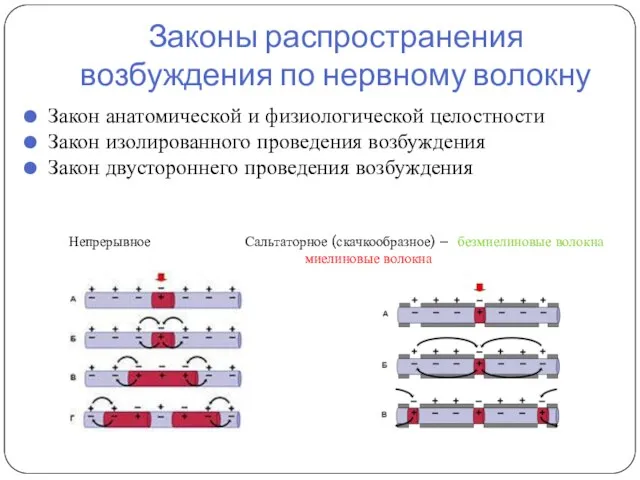
Законы распространения возбуждения по нервному волокну
Закон анатомической и физиологической целостности
Закон изолированного
Законы распространения возбуждения по нервному волокну
Закон анатомической и физиологической целостности
Закон изолированного

Проведение возбуждения вдоль нервных и мышечных волокон осуществляется при помощи так
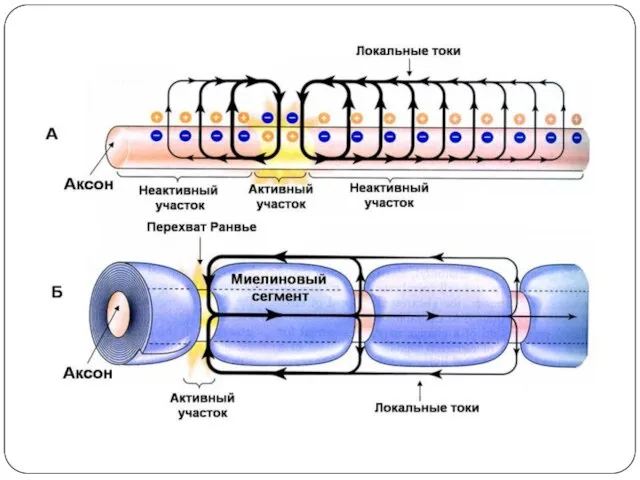
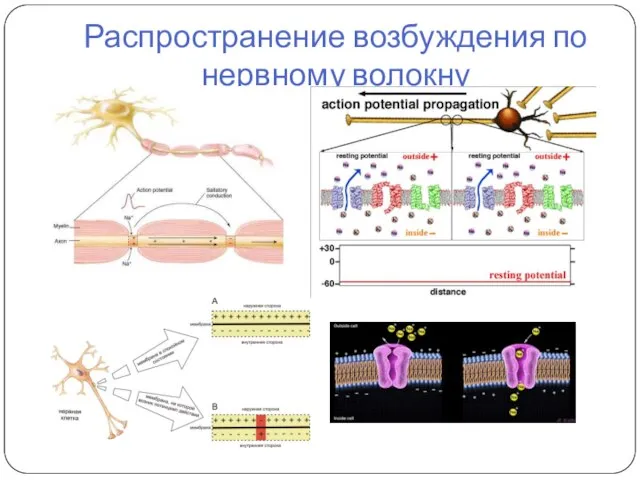
Распространение возбуждения по нервному волокну
Распространение возбуждения по нервному волокну
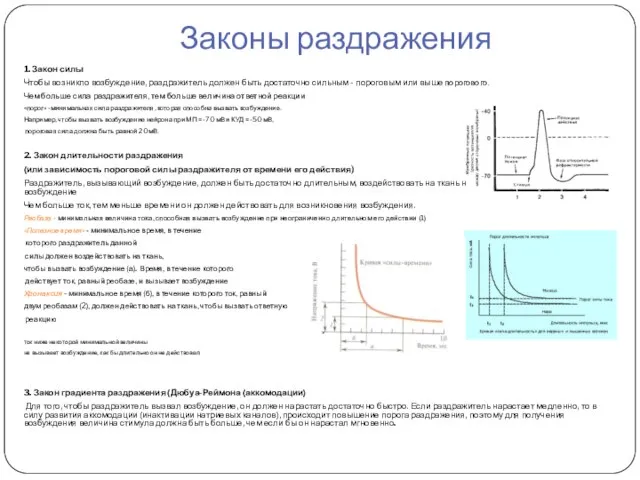
Законы раздражения
1. Закон силы
Чтобы возникло возбуждение, раздражитель должен быть достаточно сильным
Законы раздражения
1. Закон силы
Чтобы возникло возбуждение, раздражитель должен быть достаточно сильным
 Клетка - единица строения и жизнедеятельности организма
Клетка - единица строения и жизнедеятельности организма Животные Карелии
Животные Карелии Презентация на тему "Интересные факты о лягушках" - скачать бесплатно презентации по Биологии
Презентация на тему "Интересные факты о лягушках" - скачать бесплатно презентации по Биологии Периодическое культивирование. Кривая роста
Периодическое культивирование. Кривая роста Өсімдік тіршілігіндегі судың маңызы
Өсімдік тіршілігіндегі судың маңызы Амурский тигр
Амурский тигр (фрагмент урока)
(фрагмент урока)  ОСНОВЫ БИОТЕХНОЛОГИИ ТРАНСГЕНЕЗ. ЖИВОТНЫЕ. Лекция 7
ОСНОВЫ БИОТЕХНОЛОГИИ ТРАНСГЕНЕЗ. ЖИВОТНЫЕ. Лекция 7  Вакуолярная система
Вакуолярная система Органы дыхания и газообмен Завалишина Е.Н. Учитель биологии МОУ СОШ № 33 г. Талнах
Органы дыхания и газообмен Завалишина Е.Н. Учитель биологии МОУ СОШ № 33 г. Талнах  Микроскоп и его строение. Работа с микроскопом
Микроскоп и его строение. Работа с микроскопом Морская вода
Морская вода Презентация на тему "Влияние абиотических факторов на рост и развитие растений" - скачать презентации по Биологии
Презентация на тему "Влияние абиотических факторов на рост и развитие растений" - скачать презентации по Биологии Проблемы увядающей кожи. Серия омолаживающей продукции TS
Проблемы увядающей кожи. Серия омолаживающей продукции TS Первозвери Яйцекладущие и сумчатые Презентация учителя биологии Отряскиной Т.А.
Первозвери Яйцекладущие и сумчатые Презентация учителя биологии Отряскиной Т.А.  Тип Плоские черви Презентация для 7 класса
Тип Плоские черви Презентация для 7 класса GM-foods. Would you like to eat GM tomato?
GM-foods. Would you like to eat GM tomato? Презентация Журавко Анны Аскариды
Презентация Журавко Анны Аскариды Рычаги в природе и технике
Рычаги в природе и технике Человек, общество, природа. Каков же возраст нашей природы?
Человек, общество, природа. Каков же возраст нашей природы? ГМО Генетически модифицированный организм
ГМО Генетически модифицированный организм  Hypericum perforatum
Hypericum perforatum Выращивание шампиньонов Сафрай А.И
Выращивание шампиньонов Сафрай А.И Игра ДА-НЕТ
Игра ДА-НЕТ Радиобиология. История и этапы развития
Радиобиология. История и этапы развития Орган зрения - глаз
Орган зрения - глаз Шуточная биология
Шуточная биология Класс Двудольные, характерные особенности растений семейства Крестоцветные или Капустные
Класс Двудольные, характерные особенности растений семейства Крестоцветные или Капустные