- Физиология микроорганизмов

Содержание
- 2. Микроорганизмам, как и другим биологическим объектам присущи все физиологические функции. Они питаются, растут, дышат, размножаются.
- 3. Совокупность биохимических реакций у микроорганизмов подчиняется принципу биохимического единства, т.е. в биохимическом отношении все живые существа
- 4. Химический состав бактерий. Вода основной компонент бактериальной клетки, она находится в свободном и связанном состоянии. Гидролитические
- 5. В микробной клетке содержатся углерод (45%), азот (8-15%), кислород (30%), водород (6-8%) и минеральные вещества от
- 7. Минеральные вещества стимулируют процессы роста и размножения бактерий, определяют рН среды, окислительно-восстановительный потенциал, поддерживают осмотическое давление,
- 8. В составе бактерий имеются белки, углеводы, липиды, витамины.
- 9. Белки составляют 50-80% сухого вещества микробной клетки. В состав белков прокариот входят 20 аминокислот. Белки входят
- 10. Нуклеиновые кислоты бактерий представлены РНК и ДНК Содержание нуклеиновых кислот в микробной клетке зависит от вида
- 11. РНК содержится преимущественно в цитоплазме, обеспечивает транскрипцию и трансляцию на рибосомах. ДНК- преимущественно в нуклеоиде -
- 12. Углеводы (12-18%) бактерий Представлены: многоатомными спиртами (сорбит, дульцит, манит); полисахаридами (гексозы, пентозы, гликоген, декстрин); моносахаридами (глюкоза,
- 13. Липиды Липиды (истинные жиры)встречаются у риккетсий, дрожжей, микобактерий, грибов и др. Бактериальные липиды играют роль резервных
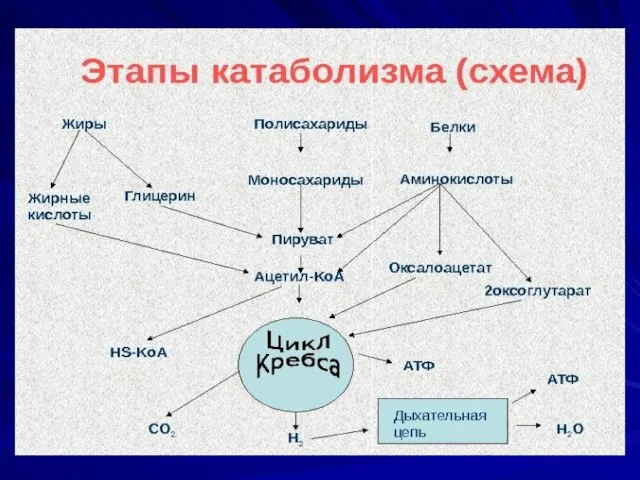
- 14. Метаболизм - совокупность ферментативных реакций направленных на получение энергии и превращение простых соединений в макромолекулы. -
- 15. Этапы ферментативных реакций катаболизма и анаболизма: Начальный (периферический метаболизм)- ферменты оказывают воздействие на исходный субстрат Ферментативное
- 16. Конструктивный метаболизм (анаболизм) Поток реакций, в результате которых за счет поступающих извне веществ строится вещество клетки,
- 17. Питание (бактерий) По способу питания бактерии относятся к голофитным объектам (нет специализированных органов для принятия пищи).
- 18. Механизмы питания у бактерий Пассивная диффузия (по градиенту концентрации) Облегченная диффузия (посредством системы транслоказ и пермеаз
- 19. Пермеазы катализируют присоединение вещества-субстрата к активному центру на своей поверхности и проводят это вещество с наружной
- 20. Выход веществ из микробной клетки осуществляется: фосфотрансферазной реакцией, при фосфорилировании переносимой молекулы; котрансляционной секрецией,
- 21. Типы секреции: I тип секреции Требует наличия 3 белков: Транспортной аденозинфосфатсинтетазы ЦПМ; Белка ЦПМ, формирующего канал
- 22. Данная система осуществляет секрецию субстратов непосредственно из цитоплазмы. Продукты секреции изначально активны.
- 23. II тип секреции «общий секреторный путь» (GSP). Секреторный аппарат формирует 12-14 белков, основная часть которых расположена
- 24. Система секреции состоит из двух частей и осуществляется в две стадии. Sec-система направляет предшественников к транслокационному
- 25. III тип секреции Обеспечивает не только перенос секреторных продуктов через мембранные структуры бактериальной клетки, но и
- 26. Система III типа Транспортируемые молекулы находятся в интактной форме и активируются только после попадания в цитоплазму
- 27. IV тип секреции Обеспечивает выделение белков- аутотранспортёров. У каждой молекулы существует терминальный N-участок с сигнальной последовательностью,
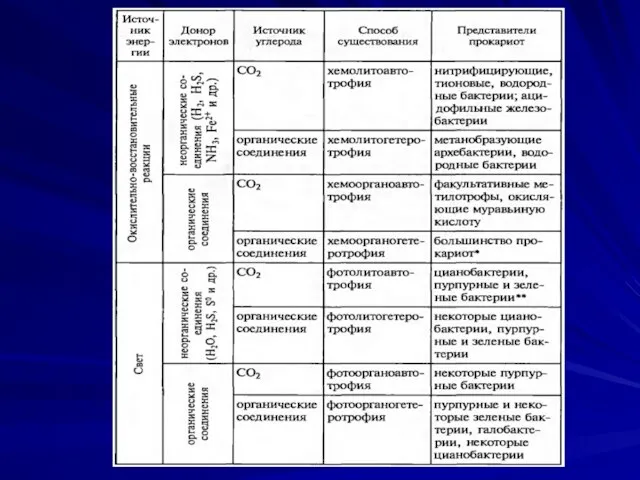
- 28. Типы питания В зависимости от источников углерода микроорганизмы делятся: Автотрофы- синтезируют все компоненты из неорганического углерода
- 29. Олиготрофы- достаточно низкой концентрации органических веществ. Копиотрофы- нуждаются в высокой концентрации органических веществ.
- 30. Строгие (облигатные) паразиты- живут только внутри клетки-хозяина; Факультативные паразиты- существуют как внутри, так и вне клетки-хозяина;
- 31. В зависимости от источников азота микроорганизмы делятся: Прототрофы- способны синтезировать азотсодержащие соединения из солей аммония, нитратов,
- 32. Факторы роста микроорганизмов макроэлементы (Ca, Mg, Fe, К, Mn) микроэлементы (Co, Ni, Cu, Zn, Mo и
- 33. Синтез углеводов Углеводы представлены в виде моно-, ди-, полисахаридов, а также комплексных соединений Автотрофы синтезируют углеводы
- 35. Получение аминокислот прокариотами Осуществляется из: Пирувата, альфакетоглутората, фумарата (из цикла трикарбоновых кислот) в реакциях аминирования (ионами
- 37. Синтез липидов У бактерий преобладают длиноцепочечные (С14-С18) насыщенные жирные кислоты и ненасыщенные жирные кислоты с одной
- 38. Энергетический метаболизм (катаболизм) Поток химических реакций, сопровождающийся мобилизацией энергии и преобразованием ее в форму, которая затем
- 40. В зависимости от источников энергии Микроорганизмы делятся: Фототрофы- способные использовать энергию света (фотосинтезирующие) Хемотрофы- получают энергию
- 41. Донором электронов не может быть предельно окисленное вещество, а акцептором- предельно восстановленное. Поэтому должен существовать внешний
- 42. В зависимости от природы доноров электронов Микроорганизмы делятся: Хемолитотрофы (хемоавтотрофы)- Н2, Fe, NH3, CH3 и др.
- 44. Механизмы получения энергии у бактерий: Окислительный метаболизм (дыхание); Бродильный (ферментативный) метаболизм Смешанный метаболизм фотосинтез
- 45. При окислительном метаболизме Энергия образуется в реакциях окисления-восстановления, при которых донорами электронов могут быть органические и
- 46. Окисление происходит в результате переноса электронов через локализованную в мембране дыхательную электрон-транспортную цепь (мембранное фосфорилирование).
- 47. Дыхательная цепь у бактерий- сложная мультиферментная система, локализована на ЦПМ и мезосомах. Переносчики располагаются в мембране
- 48. Организация дыхательной цепи
- 49. У микроорганизмов существует несколько типов богатых энергией соединений. Самые многочисленные: ацилфосфаты, нуклеотидди- и трифосфаты, аденозинфосфосульфат ацилтиоэфиры
- 50. При анаэробном дыхании происходит перенос высокоэнергетической фосфатной группы от молекулы-донора на АДФ с образованием АТФ. Субстратное
- 51. Ферментативный (бродильный) метаболизм Процесс получения энергии при котором отщепленный от субстрата водород переносится на органические соединения.
- 52. В зависимости от типа конечных продуктов различают: Спиртовое брожение Маслянокислое Молочнокислое Муравьинокислое Уксуснокислое Пропионовокислое и др.
- 54. Сбраживание углеводов Осуществляется: Гликолитическим путем (Эмбдена-Мейергофа- Парнаса); В окислительном пентозофосфатном (Варбурга- Диккенса-Хореккера) пути; 2-кето-3- дезокси-6-фосфоглюконатном пути
- 56. фотосинтез Типы фотосинтеза: Бескислородный фотосинтез, наблюдается у зеленых и пурпурных бактерий и гелиобактерий; Кислородный фотосинтез, зависимый
- 57. По способу дыхания Анаэробы (облигатные –для них кислород токсичен; факультативные - растут и размножаются как при
- 58. Прокариоты способны синтезировать энергию в большем количестве, чем требуется. Они способны консервировать ее путем синтеза восстановленных
- 59. Ферменты бактерий Все метаболические процессы протекающие в микроорганизмах являются ферментозависимыми. Набор ферментов конкретных микроорганизмов определяется их
- 60. По своей природе ферменты-белки. Ферменты распознают соответствующие субстраты, вступают с ними во взаимодействие, ускоряют протекание химических
- 61. Классификация ферментов (International Union of Biochemistry) класс – подкласс- субподкласс.
- 63. По месту действия различают: Эндоферменты, катализируют метаболизм, проходящий внутри клетки; Экзоферменты- выделяются клеткой в окружающую среду,
- 64. Регуляция метаболизма микробной клетки сводится к регуляции активности ферментативных реакций. Скорость ферментативных реакций регулируется: Путем изменения
- 65. активность ферментов Характеристикой активности ферментов является скорость, с которой они катализируют ту или иную реакцию. Она
- 66. Активность ферментов зависит от температуры среды, рН, Присутствия ионов металлов, конечных продуктов метаболизма, электрических импульсов света
- 67. Количество фермента Регулируется: на этапе транскрипции, трансляции, в процессе сборки и разрушения ферментного белка
- 68. Конститутивные ферменты- постоянно синтезируются в микробных клетках в определенных концентрациях (ферменты гликолиза). Индуцибельные (адаптивные) ферменты -
- 69. Аллостерические ферменты – их активность меняется в зависимости от взаимодействия с метаболитами или субстратами. Активатором выступает
- 70. В зависимости от химической природы субстрата различают ферменты: сахаролитические, протеолитические, липолитические.
- 71. Методы определения ферментативной активности Химический метод – количественное определение субстрата или продуктов с помощью химических реагентов
- 72. Манометрический метод – определение количества газа, выделяющегося в процессе реакции (оксидазы – по поглощению О2, декарбоксилазы
- 73. Вещества ауторегуляторы Регулируют межклеточные взаимодействия. Они характеризуются строгой родо- и видо-специфичностью. Большинство таких ауторегуляторов- вещества липидной
- 74. Питательные среды Используют для выращивания микроорганизмов в искусственных условиях. Они могут быть по консистенции: плотными, жидкими,
- 75. Питательные среды По происхождению: Животного или растительного происхождения Синтетические среды - готовят из определенных химически чистых
- 76. По целевому назначению питательные среды Делят: Универсальные (МПА, МПБ); Элективные (ЖСА, желчный бульон); Дифференциально-диагностические (Эндо, Плоскирева);
- 77. Элективные (избирательные) питательные среды Обеспечивают преимущественный рост определенной группы бактерий (желточно-солевой агар, желчный бульон)
- 78. Дифференциально-диагностические среды Позволяют дифференцировать группы или виды бактерий по ферментативной активности Среды Эндо, Левина, Плоскирева –
- 79. Требования к условиям культивирования Среда должна быть полноценной определенной рН Определенного осмотического давления и редокс-потенциала Оптимальная
- 80. Этапы выделения чистой культуры бактерий 1 этап: микроскопия мазков или нативного материала; посев на чашки с
- 82. Рост бактерий Координированное увеличение количества всех компонентов микробной клетки (массы клетки).
- 83. Размножение бактерий Увеличение количества клеток в популяции Поперечное деление: Синтеза поперечной перегородки ( у грам+ бактерий)
- 84. Эффективность размножения микроорганизмов оценивается: - Концентрацией клеток культуры в мл. Временем генерации- промежутком времени за который
- 85. Стадии развития микроорганизмов в жидкой питательной среде: Стационарная фага (отсутствия роста) длится 1 -2 часа. Лаг-фаза-
- 87. В жидкой среде бактерии Образуют: Помутнение; Поверхностную пленку; Осадок; Пристенный или придонный рост
- 88. На плотной среде бактерии образуют колонии, которые различаются по своим свойствам
- 89. Методы оценки состояния микробной культуры Прямые (непосредственный подсчет клеток под микроскопом в счетных камерах или фиксированных
- 90. Косвенные (высев на плотные питательные среды, осаждение на мембранных фильтрах, мутность суспензии, определение биомассы, общего азота,
- 91. Появление метода проточной цитометрии позволило изучать физиологическое состояние отдельных клеток.
- 92. Пигменты бактерий По химическому составу и свойствам неоднородны. растворимые в воде; растворимые в спирте; нерастворимые в
- 93. Каротиноиды- жирорастворимые пигменты красного, желтого и оранжевого цвета (микобактерии) Пирроловые- спирторастворимые: продигиозин (Serratia marcescens); Фенозиновые- водорастворимые
- 94. Функции пигментов защищают бактерии от природной ультрафиолетовой радиации, участвуют в процессах дыхания, реакциях синтеза, обладают антибиотическим
- 96. Скачать презентацию

Микроорганизмам, как и другим биологическим объектам присущи все физиологические функции.
Они
Микроорганизмам, как и другим биологическим объектам присущи все физиологические функции.
Они

Совокупность биохимических реакций у микроорганизмов подчиняется принципу биохимического единства, т.е. в
Совокупность биохимических реакций у микроорганизмов подчиняется принципу биохимического единства, т.е. в

Химический состав бактерий.
Вода основной компонент бактериальной клетки, она находится в
Химический состав бактерий.
Вода основной компонент бактериальной клетки, она находится в

В микробной клетке содержатся
углерод (45%), азот (8-15%), кислород (30%),
В микробной клетке содержатся
углерод (45%), азот (8-15%), кислород (30%),


Минеральные вещества
стимулируют процессы роста и размножения бактерий,
определяют рН среды,
Минеральные вещества
стимулируют процессы роста и размножения бактерий,
определяют рН среды,

В составе бактерий имеются
белки,
углеводы,
липиды,
витамины.
В составе бактерий имеются
белки,
углеводы,
липиды,
витамины.

Белки
составляют 50-80% сухого вещества микробной клетки. В состав белков прокариот входят
Белки
составляют 50-80% сухого вещества микробной клетки. В состав белков прокариот входят

Нуклеиновые кислоты бактерий представлены РНК и ДНК
Содержание нуклеиновых кислот в
Нуклеиновые кислоты бактерий представлены РНК и ДНК
Содержание нуклеиновых кислот в

РНК содержится преимущественно в цитоплазме, обеспечивает транскрипцию и трансляцию на рибосомах.
РНК содержится преимущественно в цитоплазме, обеспечивает транскрипцию и трансляцию на рибосомах.

Углеводы (12-18%) бактерий
Представлены:
многоатомными спиртами (сорбит, дульцит, манит);
полисахаридами (гексозы, пентозы, гликоген,
Углеводы (12-18%) бактерий
Представлены:
многоатомными спиртами (сорбит, дульцит, манит);
полисахаридами (гексозы, пентозы, гликоген,

Липиды
Липиды (истинные жиры)встречаются у риккетсий, дрожжей, микобактерий, грибов и
Липиды
Липиды (истинные жиры)встречаются у риккетсий, дрожжей, микобактерий, грибов и

Метаболизм
- совокупность ферментативных реакций направленных на получение энергии и превращение
Метаболизм
- совокупность ферментативных реакций направленных на получение энергии и превращение

Этапы ферментативных реакций катаболизма и анаболизма:
Начальный (периферический метаболизм)- ферменты оказывают воздействие
Этапы ферментативных реакций катаболизма и анаболизма:
Начальный (периферический метаболизм)- ферменты оказывают воздействие

Конструктивный метаболизм (анаболизм)
Поток реакций, в результате которых за счет поступающих извне
Конструктивный метаболизм (анаболизм)
Поток реакций, в результате которых за счет поступающих извне

Питание (бактерий)
По способу питания бактерии относятся к
голофитным объектам (нет специализированных органов
Питание (бактерий)
По способу питания бактерии относятся к
голофитным объектам (нет специализированных органов
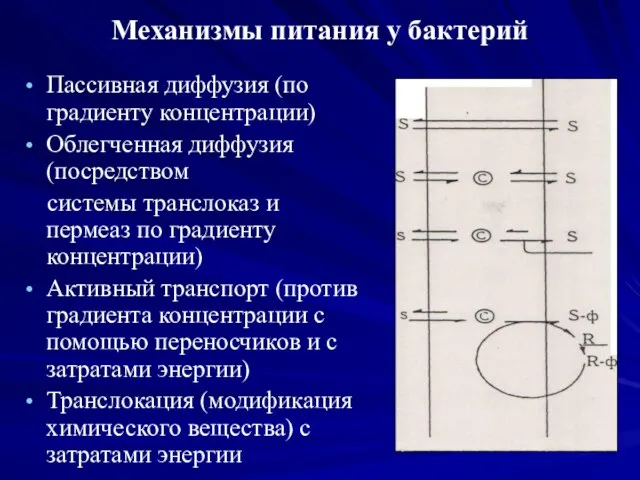
Механизмы питания у бактерий
Пассивная диффузия (по градиенту концентрации)
Облегченная диффузия (посредством
системы
Механизмы питания у бактерий
Пассивная диффузия (по градиенту концентрации)
Облегченная диффузия (посредством
системы

Пермеазы катализируют присоединение вещества-субстрата к активному центру на своей поверхности
Пермеазы катализируют присоединение вещества-субстрата к активному центру на своей поверхности
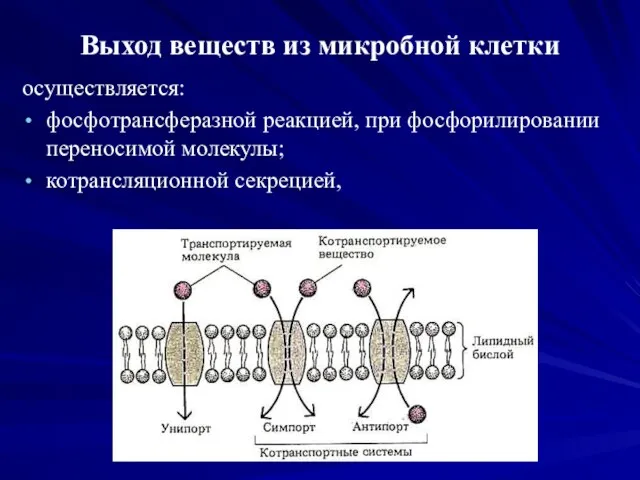
Выход веществ из микробной клетки
осуществляется:
фосфотрансферазной реакцией, при фосфорилировании переносимой молекулы;
котрансляционной
Выход веществ из микробной клетки
осуществляется:
фосфотрансферазной реакцией, при фосфорилировании переносимой молекулы;
котрансляционной

Типы секреции:
I тип секреции
Требует наличия 3 белков:
Транспортной аденозинфосфатсинтетазы ЦПМ;
Белка ЦПМ,
Типы секреции:
I тип секреции
Требует наличия 3 белков:
Транспортной аденозинфосфатсинтетазы ЦПМ;
Белка ЦПМ,

Данная система осуществляет секрецию субстратов непосредственно из цитоплазмы. Продукты секреции изначально
Данная система осуществляет секрецию субстратов непосредственно из цитоплазмы. Продукты секреции изначально

II тип секреции
«общий секреторный путь» (GSP).
Секреторный аппарат формирует 12-14 белков,
II тип секреции
«общий секреторный путь» (GSP).
Секреторный аппарат формирует 12-14 белков,
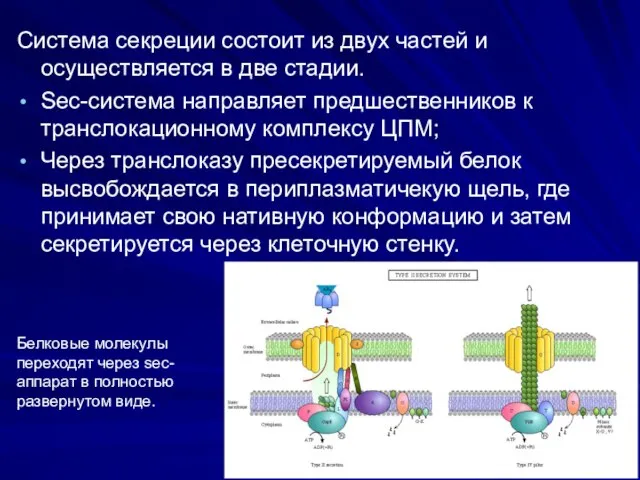
Система секреции состоит из двух частей и осуществляется в две стадии.
Sec-система
Система секреции состоит из двух частей и осуществляется в две стадии.
Sec-система
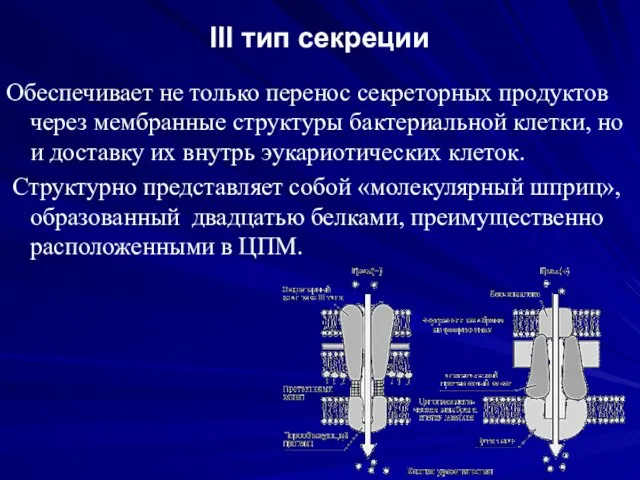
III тип секреции
Обеспечивает не только перенос секреторных продуктов через мембранные структуры
III тип секреции
Обеспечивает не только перенос секреторных продуктов через мембранные структуры
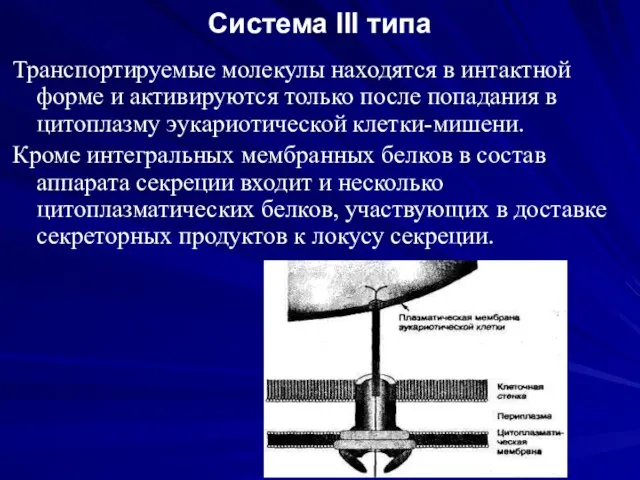
Система III типа
Транспортируемые молекулы находятся в интактной форме и активируются
Система III типа
Транспортируемые молекулы находятся в интактной форме и активируются
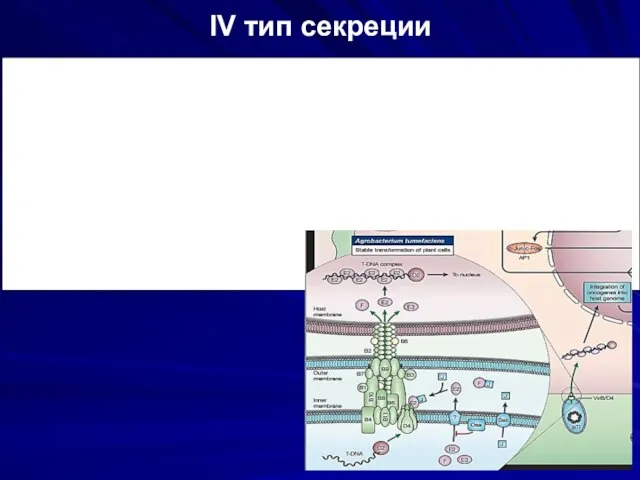
IV тип секреции
Обеспечивает выделение белков- аутотранспортёров.
У каждой молекулы существует терминальный
IV тип секреции
Обеспечивает выделение белков- аутотранспортёров.
У каждой молекулы существует терминальный
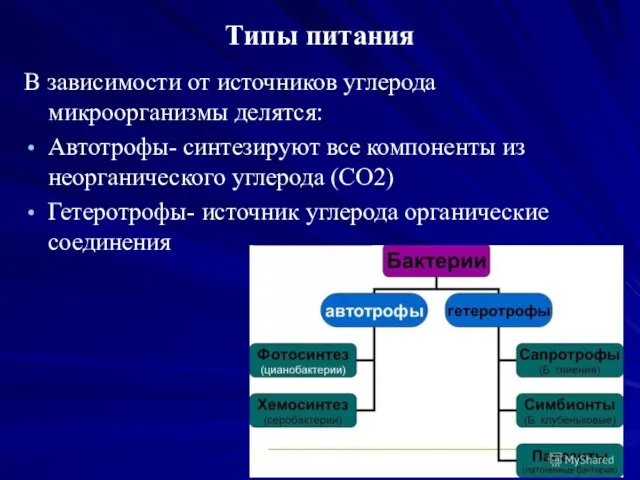
Типы питания
В зависимости от источников углерода
микроорганизмы делятся:
Автотрофы- синтезируют все компоненты из
Типы питания
В зависимости от источников углерода
микроорганизмы делятся:
Автотрофы- синтезируют все компоненты из

Олиготрофы- достаточно низкой концентрации органических веществ.
Копиотрофы- нуждаются в высокой концентрации органических
Олиготрофы- достаточно низкой концентрации органических веществ.
Копиотрофы- нуждаются в высокой концентрации органических

Строгие (облигатные) паразиты- живут только внутри клетки-хозяина;
Факультативные паразиты- существуют как
Строгие (облигатные) паразиты- живут только внутри клетки-хозяина;
Факультативные паразиты- существуют как

В зависимости от источников азота
микроорганизмы делятся:
Прототрофы- способны синтезировать азотсодержащие соединения из
В зависимости от источников азота
микроорганизмы делятся:
Прототрофы- способны синтезировать азотсодержащие соединения из

Факторы роста микроорганизмов
макроэлементы (Ca, Mg, Fe, К, Mn)
микроэлементы (Co,
Факторы роста микроорганизмов
макроэлементы (Ca, Mg, Fe, К, Mn)
микроэлементы (Co,

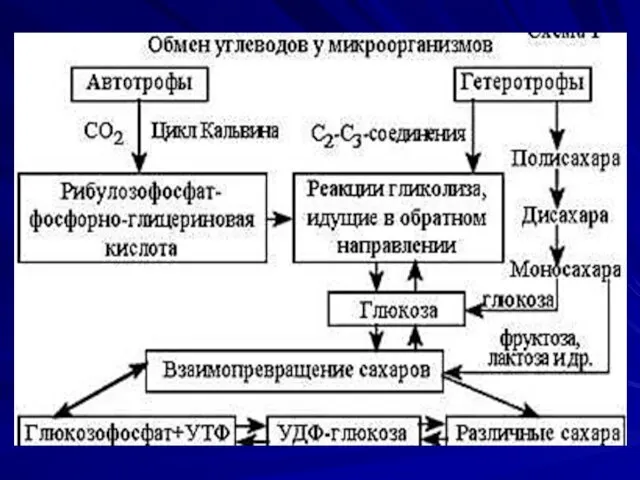
Синтез углеводов
Углеводы представлены в виде моно-, ди-, полисахаридов, а также комплексных
Синтез углеводов
Углеводы представлены в виде моно-, ди-, полисахаридов, а также комплексных

Получение аминокислот прокариотами
Осуществляется из:
Пирувата, альфакетоглутората, фумарата (из цикла трикарбоновых кислот) в
Получение аминокислот прокариотами
Осуществляется из:
Пирувата, альфакетоглутората, фумарата (из цикла трикарбоновых кислот) в

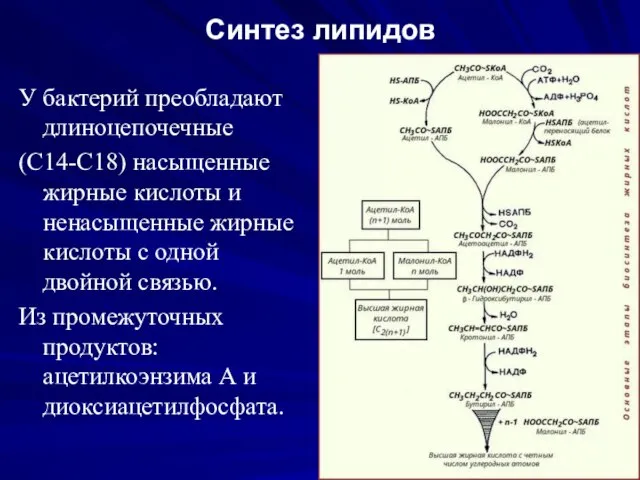
Синтез липидов
У бактерий преобладают длиноцепочечные
(С14-С18) насыщенные жирные кислоты и ненасыщенные
Синтез липидов
У бактерий преобладают длиноцепочечные
(С14-С18) насыщенные жирные кислоты и ненасыщенные

Энергетический метаболизм (катаболизм)
Поток химических реакций, сопровождающийся мобилизацией энергии и преобразованием ее
Энергетический метаболизм (катаболизм)
Поток химических реакций, сопровождающийся мобилизацией энергии и преобразованием ее

В зависимости от источников энергии
Микроорганизмы делятся:
Фототрофы- способные использовать энергию света (фотосинтезирующие)
Хемотрофы-
В зависимости от источников энергии
Микроорганизмы делятся:
Фототрофы- способные использовать энергию света (фотосинтезирующие)
Хемотрофы-
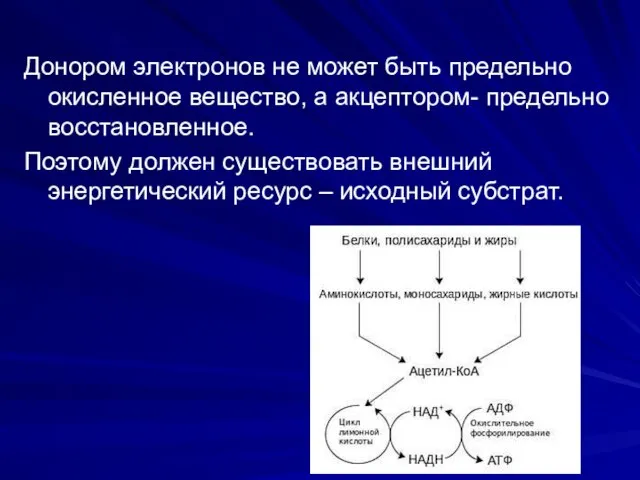
Донором электронов не может быть предельно окисленное вещество, а акцептором- предельно
Донором электронов не может быть предельно окисленное вещество, а акцептором- предельно

В зависимости от природы доноров электронов
Микроорганизмы делятся:
Хемолитотрофы (хемоавтотрофы)- Н2, Fe, NH3,
В зависимости от природы доноров электронов
Микроорганизмы делятся:
Хемолитотрофы (хемоавтотрофы)- Н2, Fe, NH3,
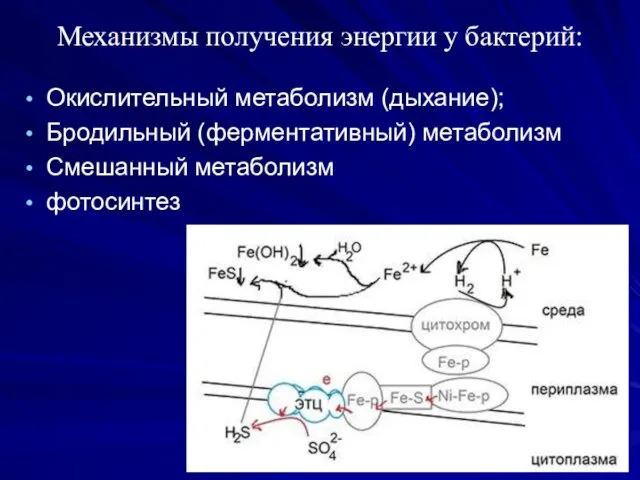
Механизмы получения энергии у бактерий:
Окислительный метаболизм (дыхание);
Бродильный (ферментативный) метаболизм
Смешанный метаболизм
фотосинтез
Механизмы получения энергии у бактерий:
Окислительный метаболизм (дыхание);
Бродильный (ферментативный) метаболизм
Смешанный метаболизм
фотосинтез

При окислительном метаболизме
Энергия образуется в реакциях окисления-восстановления, при которых донорами
При окислительном метаболизме
Энергия образуется в реакциях окисления-восстановления, при которых донорами
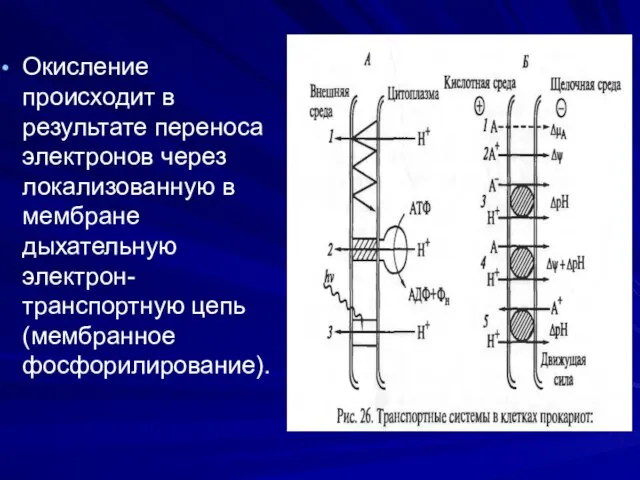
Окисление происходит в результате переноса электронов через локализованную в мембране дыхательную
Окисление происходит в результате переноса электронов через локализованную в мембране дыхательную

Дыхательная цепь у бактерий- сложная мультиферментная система, локализована на ЦПМ и
Дыхательная цепь у бактерий- сложная мультиферментная система, локализована на ЦПМ и
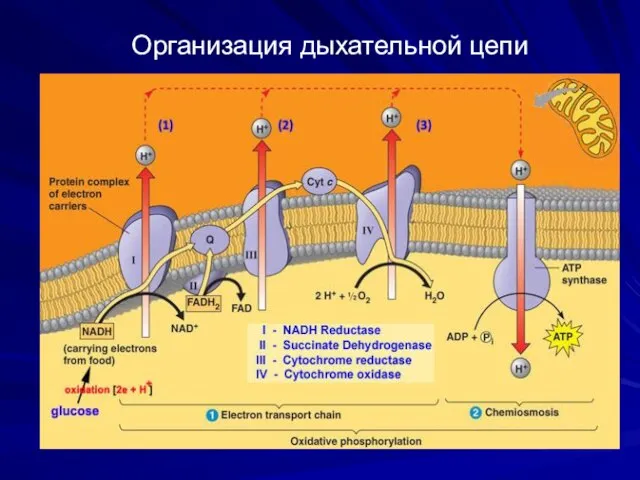
Организация дыхательной цепи
Организация дыхательной цепи

У микроорганизмов существует несколько типов богатых энергией соединений. Самые многочисленные:
ацилфосфаты,
У микроорганизмов существует несколько типов богатых энергией соединений. Самые многочисленные:
ацилфосфаты,

При анаэробном дыхании
происходит перенос высокоэнергетической фосфатной группы от молекулы-донора на
При анаэробном дыхании
происходит перенос высокоэнергетической фосфатной группы от молекулы-донора на
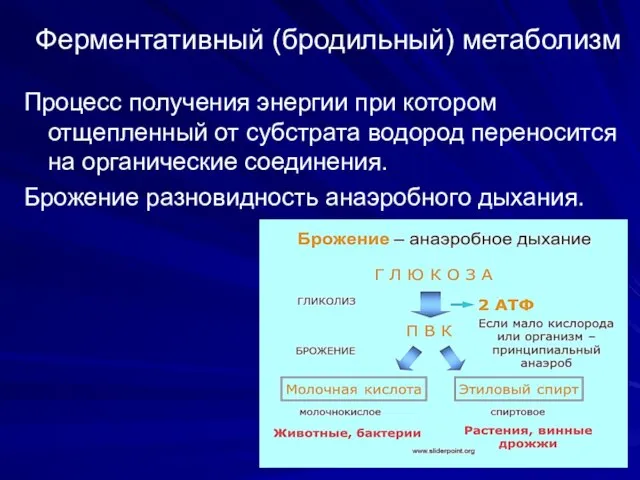
Ферментативный (бродильный) метаболизм
Процесс получения энергии при котором отщепленный от субстрата
Ферментативный (бродильный) метаболизм
Процесс получения энергии при котором отщепленный от субстрата

В зависимости от типа конечных продуктов различают:
Спиртовое брожение
Маслянокислое
Молочнокислое
Муравьинокислое
Уксуснокислое
Пропионовокислое и др.
В зависимости от типа конечных продуктов различают:
Спиртовое брожение
Маслянокислое
Молочнокислое
Муравьинокислое
Уксуснокислое
Пропионовокислое и др.
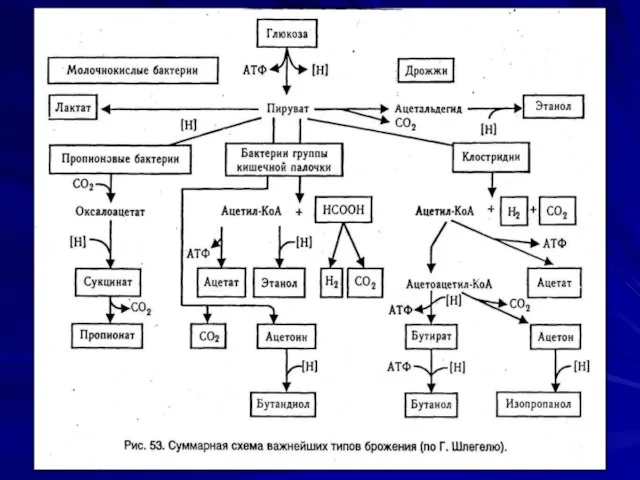

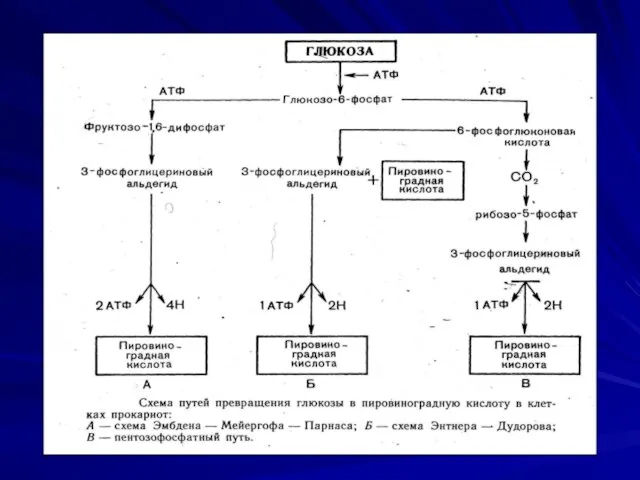
Сбраживание углеводов
Осуществляется:
Гликолитическим путем (Эмбдена-Мейергофа- Парнаса);
В окислительном пентозофосфатном (Варбурга- Диккенса-Хореккера) пути;
2-кето-3- дезокси-6-фосфоглюконатном
Сбраживание углеводов
Осуществляется:
Гликолитическим путем (Эмбдена-Мейергофа- Парнаса);
В окислительном пентозофосфатном (Варбурга- Диккенса-Хореккера) пути;
2-кето-3- дезокси-6-фосфоглюконатном

фотосинтез
Типы фотосинтеза:
Бескислородный фотосинтез, наблюдается у зеленых и пурпурных бактерий и гелиобактерий;
Кислородный
фотосинтез
Типы фотосинтеза:
Бескислородный фотосинтез, наблюдается у зеленых и пурпурных бактерий и гелиобактерий;
Кислородный

По способу дыхания
Анаэробы (облигатные –для них кислород токсичен; факультативные -
По способу дыхания
Анаэробы (облигатные –для них кислород токсичен; факультативные -

Прокариоты способны синтезировать энергию в большем количестве, чем требуется.
Они способны
Прокариоты способны синтезировать энергию в большем количестве, чем требуется.
Они способны

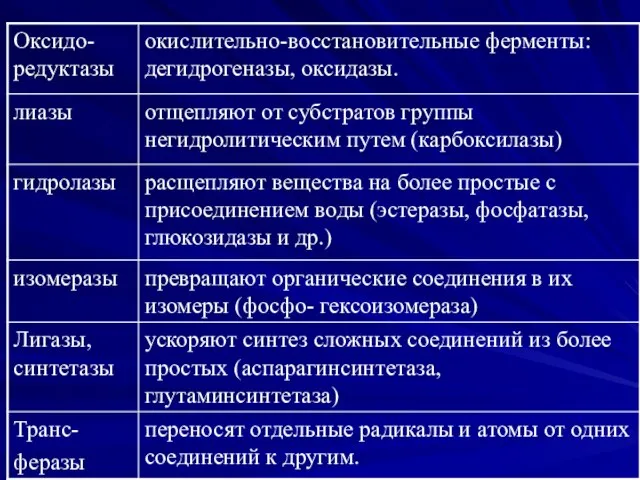
Ферменты бактерий
Все метаболические процессы протекающие в микроорганизмах являются ферментозависимыми.
Набор ферментов конкретных
Ферменты бактерий
Все метаболические процессы протекающие в микроорганизмах являются ферментозависимыми.
Набор ферментов конкретных

По своей природе
ферменты-белки.
Ферменты распознают соответствующие субстраты,
вступают с ними во
По своей природе
ферменты-белки.
Ферменты распознают соответствующие субстраты,
вступают с ними во
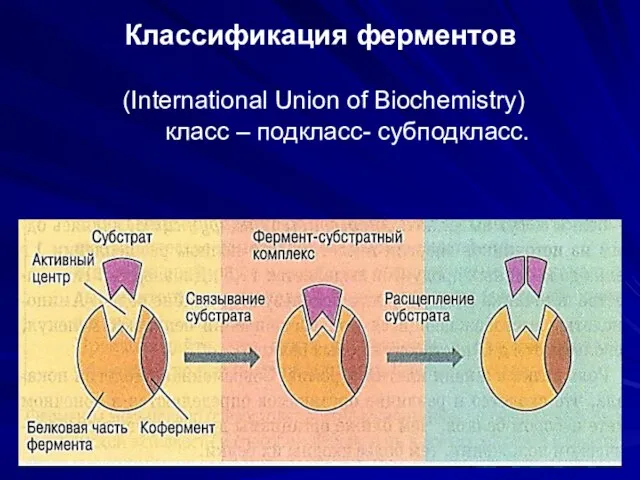
Классификация ферментов
(International Union of Biochemistry)
класс – подкласс- субподкласс.
Классификация ферментов
(International Union of Biochemistry)
класс – подкласс- субподкласс.

По месту действия
различают:
Эндоферменты, катализируют метаболизм, проходящий внутри клетки;
Экзоферменты- выделяются клеткой
По месту действия
различают:
Эндоферменты, катализируют метаболизм, проходящий внутри клетки;
Экзоферменты- выделяются клеткой

Регуляция метаболизма микробной клетки сводится к регуляции активности ферментативных реакций.
Скорость ферментативных
Регуляция метаболизма микробной клетки сводится к регуляции активности ферментативных реакций.
Скорость ферментативных

активность ферментов
Характеристикой активности ферментов является скорость, с которой они катализируют ту
активность ферментов
Характеристикой активности ферментов является скорость, с которой они катализируют ту

Активность ферментов зависит
от температуры среды,
рН,
Присутствия ионов
Активность ферментов зависит
от температуры среды,
рН,
Присутствия ионов
Количество фермента
Регулируется:
на этапе транскрипции,
трансляции,
в процессе сборки и разрушения
Количество фермента
Регулируется:
на этапе транскрипции,
трансляции,
в процессе сборки и разрушения

Конститутивные ферменты- постоянно синтезируются в микробных клетках в определенных концентрациях (ферменты

Аллостерические ферменты – их активность меняется в зависимости от взаимодействия с
Аллостерические ферменты – их активность меняется в зависимости от взаимодействия с

В зависимости от химической природы субстрата различают ферменты:
сахаролитические,
протеолитические,
липолитические.
В зависимости от химической природы субстрата различают ферменты:
сахаролитические,
протеолитические,
липолитические.

Методы определения ферментативной активности
Химический метод – количественное определение субстрата или продуктов
Методы определения ферментативной активности
Химический метод – количественное определение субстрата или продуктов

Манометрический метод – определение количества газа, выделяющегося в процессе реакции (оксидазы
Манометрический метод – определение количества газа, выделяющегося в процессе реакции (оксидазы

Вещества ауторегуляторы
Регулируют межклеточные взаимодействия. Они характеризуются строгой родо- и видо-специфичностью.
Большинство таких
Вещества ауторегуляторы
Регулируют межклеточные взаимодействия. Они характеризуются строгой родо- и видо-специфичностью.
Большинство таких

Питательные среды
Используют для выращивания микроорганизмов в искусственных условиях.
Они могут быть по
Питательные среды
Используют для выращивания микроорганизмов в искусственных условиях.
Они могут быть по
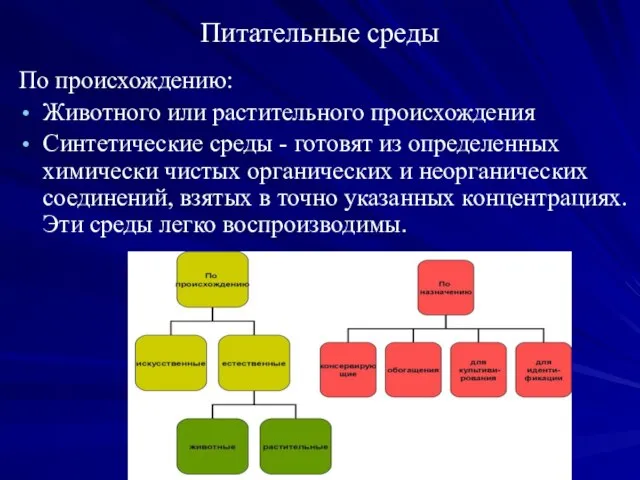
Питательные среды
По происхождению:
Животного или растительного происхождения
Синтетические среды - готовят из
Питательные среды
По происхождению:
Животного или растительного происхождения
Синтетические среды - готовят из

По целевому назначению питательные среды
Делят:
Универсальные (МПА, МПБ);
Элективные (ЖСА, желчный бульон);
Дифференциально-диагностические (Эндо,
По целевому назначению питательные среды
Делят:
Универсальные (МПА, МПБ);
Элективные (ЖСА, желчный бульон);
Дифференциально-диагностические (Эндо,

Элективные (избирательные) питательные среды
Обеспечивают преимущественный рост определенной группы бактерий (желточно-солевой агар,
Элективные (избирательные) питательные среды
Обеспечивают преимущественный рост определенной группы бактерий (желточно-солевой агар,
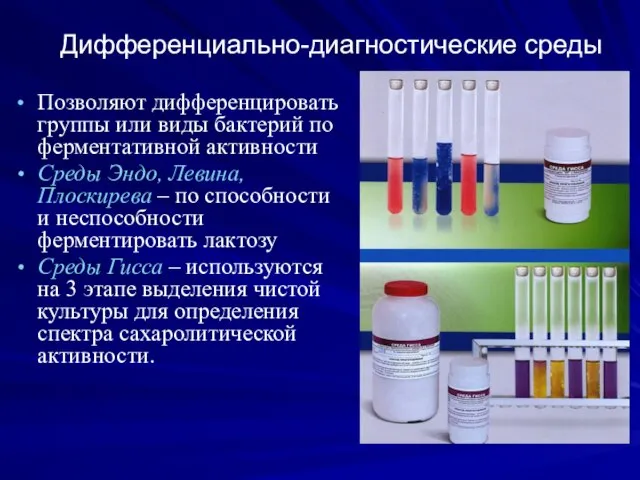
Дифференциально-диагностические среды
Позволяют дифференцировать группы или виды бактерий по ферментативной активности
Среды Эндо,
Дифференциально-диагностические среды
Позволяют дифференцировать группы или виды бактерий по ферментативной активности
Среды Эндо,

Требования к условиям культивирования
Среда должна быть полноценной
определенной рН
Определенного осмотического давления
Требования к условиям культивирования
Среда должна быть полноценной
определенной рН
Определенного осмотического давления

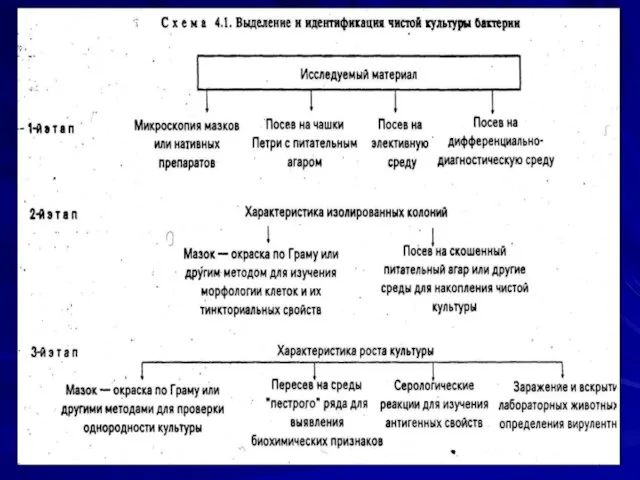
Этапы выделения чистой культуры бактерий
1 этап:
микроскопия мазков или нативного материала;
Этапы выделения чистой культуры бактерий
1 этап:
микроскопия мазков или нативного материала;
Рост бактерий
Координированное увеличение количества всех компонентов микробной клетки (массы клетки).
Рост бактерий
Координированное увеличение количества всех компонентов микробной клетки (массы клетки).
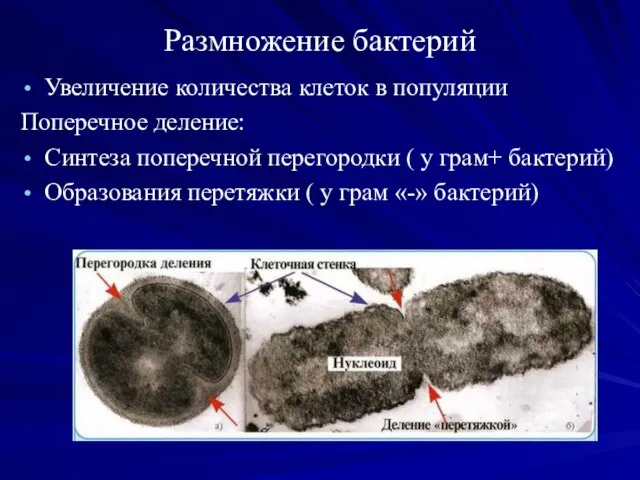
Размножение бактерий
Увеличение количества клеток в популяции
Поперечное деление:
Синтеза поперечной перегородки ( у
Размножение бактерий
Увеличение количества клеток в популяции
Поперечное деление:
Синтеза поперечной перегородки ( у

Эффективность размножения микроорганизмов оценивается:
- Концентрацией клеток культуры в мл.
Временем генерации- промежутком
Эффективность размножения микроорганизмов оценивается:
- Концентрацией клеток культуры в мл.
Временем генерации- промежутком

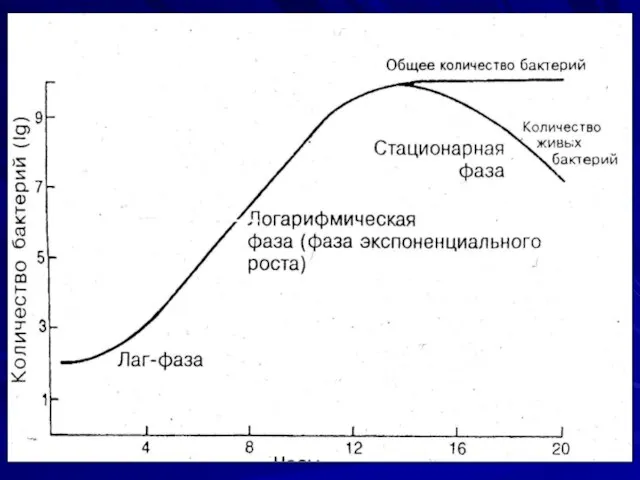
Стадии развития микроорганизмов в жидкой питательной среде:
Стационарная фага (отсутствия роста) длится
Стадии развития микроорганизмов в жидкой питательной среде:
Стационарная фага (отсутствия роста) длится

В жидкой среде бактерии
Образуют:
Помутнение;
Поверхностную пленку;
Осадок;
Пристенный или придонный рост
В жидкой среде бактерии
Образуют:
Помутнение;
Поверхностную пленку;
Осадок;
Пристенный или придонный рост
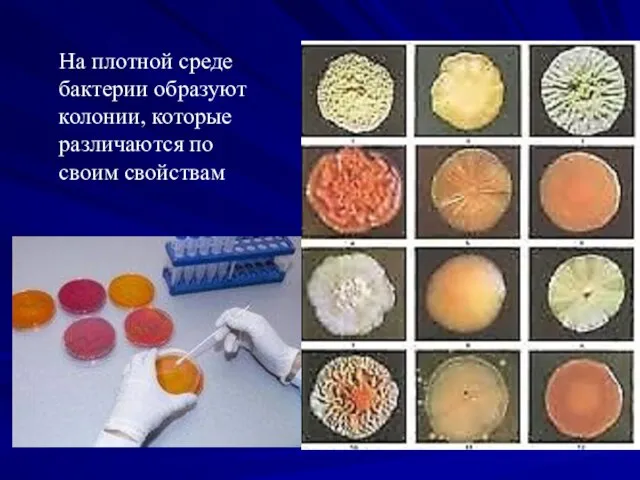
На плотной среде
бактерии образуют
колонии, которые
различаются по
своим свойствам
На плотной среде
бактерии образуют
колонии, которые
различаются по
своим свойствам

Методы оценки состояния микробной культуры
Прямые (непосредственный подсчет клеток под микроскопом в
Методы оценки состояния микробной культуры
Прямые (непосредственный подсчет клеток под микроскопом в
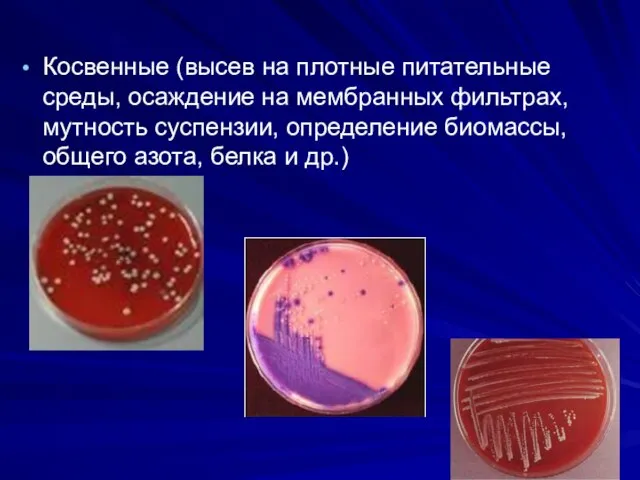
Косвенные (высев на плотные питательные среды, осаждение на мембранных фильтрах, мутность
Косвенные (высев на плотные питательные среды, осаждение на мембранных фильтрах, мутность

Появление метода проточной цитометрии позволило изучать физиологическое состояние отдельных клеток.
Появление метода проточной цитометрии позволило изучать физиологическое состояние отдельных клеток.

Пигменты бактерий
По химическому составу и свойствам неоднородны.
растворимые в воде;
растворимые в
Пигменты бактерий
По химическому составу и свойствам неоднородны.
растворимые в воде;
растворимые в

Каротиноиды- жирорастворимые пигменты красного, желтого и оранжевого цвета (микобактерии)
Пирроловые- спирторастворимые: продигиозин
Каротиноиды- жирорастворимые пигменты красного, желтого и оранжевого цвета (микобактерии)
Пирроловые- спирторастворимые: продигиозин

Функции пигментов
защищают бактерии от природной ультрафиолетовой радиации,
участвуют в процессах дыхания,
Функции пигментов
защищают бактерии от природной ультрафиолетовой радиации,
участвуют в процессах дыхания,
 Архозавры. Разнообразие архозавров
Архозавры. Разнообразие архозавров Proving the variability of species in nature
Proving the variability of species in nature Строение клетки. Клеточная мембрана. Ядро. Разработала: учитель биологии МОУ СОШ №1 Радько О.В.
Строение клетки. Клеточная мембрана. Ядро. Разработала: учитель биологии МОУ СОШ №1 Радько О.В. Происхождение жизни. Источники информации и логика рассуждений
Происхождение жизни. Источники информации и логика рассуждений Тип кишечнополостные Вводная характеристика кишечнополостных. Класс Гидроидные
Тип кишечнополостные Вводная характеристика кишечнополостных. Класс Гидроидные  Цікаві факти з генетики
Цікаві факти з генетики  Цветковые или Покрытосеменные
Цветковые или Покрытосеменные Вегетативное размножение
Вегетативное размножение Цитология, как комплексная наука о клетке
Цитология, как комплексная наука о клетке Krovenosnaya_sistema_Dvizhenie_krovi_i_limfy_v_organizme
Krovenosnaya_sistema_Dvizhenie_krovi_i_limfy_v_organizme Тип Кишечнополостные Гидра
Тип Кишечнополостные Гидра  НАСЛЕДСТВЕННЫЕ БОЛЕЗНИ ЧЕЛОВЕКА НАСЛЕДСТВЕННЫЕ БОЛЕЗНИ ЧЕЛОВЕКА
НАСЛЕДСТВЕННЫЕ БОЛЕЗНИ ЧЕЛОВЕКА НАСЛЕДСТВЕННЫЕ БОЛЕЗНИ ЧЕЛОВЕКА  Микробиология и биотехнология. Микроорганизмы
Микробиология и биотехнология. Микроорганизмы Упоротые животные кембрийского периода
Упоротые животные кембрийского периода Государственный природный заказник Балтасинский
Государственный природный заказник Балтасинский Внутреннее строение рыб
Внутреннее строение рыб Аквариумные рыбки
Аквариумные рыбки Видообразованиe. Пути видообразования
Видообразованиe. Пути видообразования Articular system
Articular system Factors of evolution
Factors of evolution Биологические науки
Биологические науки Aorta. The walls of the arteries
Aorta. The walls of the arteries Тема урока: Высшие споровые. Плауны, хвощи, папоротники Цель урока: - изучить представителей современных папоротникообразных; -
Тема урока: Высшие споровые. Плауны, хвощи, папоротники Цель урока: - изучить представителей современных папоротникообразных; -  Характеристика древесных растений, используемых в ландшафтной архитектуре. Лекция 1
Характеристика древесных растений, используемых в ландшафтной архитектуре. Лекция 1 Презентация по экологии ВОДНЫЕ РЕСУРСЫ: РАЦИОНАЛЬНОЕ ИСПОЛЬЗОВАНИЕ И ОХРАНА
Презентация по экологии ВОДНЫЕ РЕСУРСЫ: РАЦИОНАЛЬНОЕ ИСПОЛЬЗОВАНИЕ И ОХРАНА  Презентация на тему "Дмитрий Иванович Мендилеев" - скачать презентации по Биологии
Презентация на тему "Дмитрий Иванович Мендилеев" - скачать презентации по Биологии Половое размножение живых организмов
Половое размножение живых организмов Подготовила учитель биологии Захарина Наталья Михайловна
Подготовила учитель биологии Захарина Наталья Михайловна