- Происхождение жизни. Источники информации и логика рассуждений

Содержание
- 2. Откуда мы получаем информацию о том, как возникла жизнь?
- 4. Представим, что мы строим пазл – пытаемся собрать очень большую, сложную, но невероятно красивую картину из
- 5. При сопоставлении фактов возникает смысл и логическая связь. Множество фактов, которые необходимо добыть и связать в
- 6. Источники информации I. Наши знания о том, как реально устроены современные живые системы II. Лабораторные эксперименты
- 7. I. Чтобы узнать КАК возникла жизнь, нужно знать как она устроена. Эволюционный процесс первоначально рассматривался в
- 8. I. Чтобы узнать КАК возникла жизнь, нужно знать как она устроена. Когда стали известны строение и
- 9. Происхождение жизни – это поэтапная молекулярная эволюция Молекулярная эволюция – это наука, изучающая изменения генетических макромолекул
- 11. II. Искусственное моделирование различных этапов происхождения жизни. «Мне думается, что воспроизведение жизни в лаборатории не является
- 12. II. Лабораторные эксперименты 1 этап. (20-80-е годы ХХ века). Основные усилия были направлены на экспериментальное исследование
- 13. II. Лабораторные эксперименты В 1861 г. А. М. Бутлеров, нагревая формальдегид в щелочном растворе Ca(OH)2, получил
- 14. II. Лабораторные эксперименты Опыты С. Миллера по моделированию абиогенного синтеза аминокислот и других органических компонентов жизни.
- 15. Из опытов С.Миллера Особенно богатый набор аминокислот получался при моделировании синтеза вблизи вулканов
- 16. II. Лабораторные эксперименты Синтез аденина из синильной кислоты. Аденин – компонент ДНК и РНК.
- 17. Абиогенный синтез пуринов
- 18. Лабораторное моделирование абиогенного синтеза пиримидиновых рибонуклеотидов (цитидина и уридина) (2010 г)
- 19. Синтез аденина в метеоритах
- 20. II. Лабораторные эксперименты 2 этап. В 90-е годы существенную роль стали играть эксперименты с информационными биологическими
- 21. Позволяет отбирать мутантные молекулы РНК, которые специфически связываются с тем или иным субстратом. РНК- ключевой катализатор
- 22. I. Эволюция РНК в пробирке Технология полимеразной цепной реакции (ПЦР) позволяет копировать (с мутациями) молекулы РНК,
- 23. II. Создание минимально работоспособной клетки Создание небольшого синтетического генома, интегрированного в бактериальную клетку (проект завершён) Создание
- 24. Минимальный метаболизм современной клетки 31 вещество из внешней среды
- 25. Минимальная модель современной клетки Теоретически минимальный набор генов - 241 (102 одиночных генов, 19 генных кластеров)
- 26. II. Заключение. Фактически искусcтвенное моделирование, подобно следственному эксперименту, должно служить важнейшим аргументом того, как возникла жизнь.
- 27. II. Анализ молекулярных реликтов (молекулярных ископаемых) – самый информативный источник. «Молекулярное ископаемое» - абстракция обозначающая любые
- 28. III. Молекулярные реликты «Жизнь в своем развитии никогда не пренебрегала тем, что уже построено, а строила
- 29. III. Молекулярные реликты Современные биохимические механизмы несут на себе отпечаток эволюционного прошлого – «родимые пятна» своей
- 30. III. Что относится к молекулярным реликтам Блочное разделение клеточного метаболизма по характеру синтезируемых веществ. Третий блок
- 31. Синтез аминокислот обеспечивается функционированием промежуточного метаболизма, который должен был возникнуть до стадии биосинтеза аминокислот, а, следовательно,
- 32. III. Молекулярные реликты " Если эволюция метаболических путей шла путем последовательного добавления новых ферментативных реакций к
- 33. Почему РНК возникла до ДНК? Молекула РНК в отличие от ДНК способна образовывать большое разнообразие вторичных,
- 35. ПРИНЦИПЫ МАКРОМОЛЕКУЛЯРНОЙ СТРУКТУРЫ РНК РНК, в отличие от ДНК, одноцепочечна. Высокомолекулярные РНК представляют собой ковалентно непрерывные
- 38. Почему РНК возникла до ДНК? РНК способна удлинять короткие фрагменты РНК РНК способна к частичной неточной
- 39. Может ли РНК воспроизводить РНК? Рибозимы могут воспроизводить РНК из мономеров очень неточно. РНК может воспроизводиться
- 40. Схема репликации рибозимов в опыте Линкольн и Джойса. Исходными субстратами служат 4 олигонуклеотида (два розовых в
- 41. Почему РНК возникла до ДНК? Синтез рибонуклеотидов предшествует синтезу дезкоксирибонуклеотидов Сначала синтезируются компоненты РНК Затем из
- 42. Почему РНК возникла до ДНК? Репликация РНК, которую можно наблюдать в современных вирусах много проще репликации
- 43. Почему РНК возникла до ДНК? В опытах по моделированию абиогенного синтеза рибоза образуется существенно легче, чем
- 44. РНК-кофакторы – реликты мира РНК, существовавшего до появления ДНК. Кофактор (синонимы - кофермент, коэнзим) –небелковое вещество,
- 45. III. Молекулярные реликты Хотя отсутствуют прямые данные о существовании РНКовых организмов, «лицо» РНКового мира обнаруживается в
- 46. III. Молекулярные реликты Кодирующие макромолекулы (ДНК и РНК) эволюционируют с разными скоростями. Наиболее медленно изменяются обычно
- 47. РНК-кофакторы – реликты мира РНК
- 48. РНК-кофакторы – реликты мира РНК
- 49. РНК-кофакторы – универсальные переносчики электронов и атомов водорода.
- 50. РНК-кофакторы – реликты мира РНК Кофермент А, как и любой кофактор, содержит компонент РНК. Этот компонент
- 51. РНК-кофакторы – реликты мира РНК Некоторые РНК-кофакторы влючают не только компоненты РНК, но и белков –
- 52. В чём заключается универсальность РНК-кофактов? В использовании универсальной энергетической валюты – АТФ и других нуклеотидтрифосфатов. АТФ
- 53. Современный метаболизм как палимпсест мира РНК Палимпсест означает то, что написано на месте прежнего текста. Древние
- 54. РНК-кофакторы. Резюме. РНК-кофакторы являлись ключевыми компонентами в биохимических процессах ещё до возникновения ДНК-организмов. Нуклеотидные кофакторы следует
- 55. Транспортные РНК – молекулярные реликты Молекула тРНК относительно небольшая, её длина варьирует от 74 до 95
- 56. Древние тРНК с комплементарными антикодонами кодировались комплементарными РНК-парами. В тРНК некоторые последовательности изменяются с ходом эволюции
- 57. III. Другие молекулярные реликты. Рибосома – одна из самых древних структур, возникшая на ранних этапах возникновения
- 58. Ключевое открытие сравнительной геномики Типичное время исчезновения сходства последовательностей у гомологичных генов сравнимо со временем существования
- 59. Распределение генов по эволюционному возрасту
- 60. Ядро, оболочка и облако ЯДРО - (Почти) универсальные гены, те, что представлены в (почти) всех геномах
- 61. Ядро, оболочка и облако ОБОЛОЧКА - Умеренно консервативная генная оболочка состоит из ортологичных генов, представленных в
- 62. Ядро, оболочка и облако ОБЛАКО. Гены «облака» составляют переменную долю в каждом геноме, обычно в интервале
- 63. Ядро, оболочка и облако у прокариот
- 64. Прогенот – коллективная форма жизни. До появления ДНК живые системы не дивергировали на отдельные виды. Этому
- 65. Традиционные представления о филогении – ветвящееся дерево
- 66. Горизонтальный обмен генами превратил дерево в мицелий Не менее 80% генов в каждом прокариотическом геноме участвовали
- 67. «Горячая» и «холодная» жизнь
- 69. IV. Анализ геологических данных. Они дают два вида информации. Первый вид - геохронологию, которая в частности
- 70. Период массовых метеоритных бомбардировок
- 71. IV. Анализ геологических данных. Геологические исследования позволяют также находить так называемые особые условия, в которых возможен
- 72. Геологические данные позволяют воссоздать условия на Земле в момент возникновения жизни. Ничтожные концентрации свободного кислорода в
- 73. Условия на первобытной Земле
- 74. Геологические данные позволяют воссоздать хронологию и откалибровать молекулярные часы эволюции.
- 75. Строматолиты — ископаемые остатки цианобактериальных матов.
- 76. Древняя глобальная биосфера Была представлена бактериальными матами
- 78. Окаменелости цианобактерий (3,5 млрд. лет)
- 80. Скачать презентацию

Откуда мы получаем информацию о том, как возникла жизнь?
Откуда мы получаем информацию о том, как возникла жизнь?


Представим, что мы строим пазл – пытаемся собрать очень большую,
Представим, что мы строим пазл – пытаемся собрать очень большую,

При сопоставлении фактов возникает смысл и логическая связь.
Множество фактов,
которые необходимо
добыть и
При сопоставлении фактов возникает смысл и логическая связь.
Множество фактов,
которые необходимо
добыть и

Источники информации
I. Наши знания о том, как реально устроены современные живые
Источники информации
I. Наши знания о том, как реально устроены современные живые

I. Чтобы узнать КАК возникла жизнь, нужно знать как она устроена.
Эволюционный
I. Чтобы узнать КАК возникла жизнь, нужно знать как она устроена.
Эволюционный

I. Чтобы узнать КАК возникла жизнь, нужно знать как она устроена.
Когда
I. Чтобы узнать КАК возникла жизнь, нужно знать как она устроена.
Когда

Происхождение жизни – это поэтапная молекулярная эволюция
Молекулярная эволюция – это наука,
Происхождение жизни – это поэтапная молекулярная эволюция
Молекулярная эволюция – это наука,


II. Искусственное моделирование различных этапов происхождения жизни.
«Мне думается, что
II. Искусственное моделирование различных этапов происхождения жизни.
«Мне думается, что

II. Лабораторные эксперименты
1 этап. (20-80-е годы ХХ века). Основные усилия были
II. Лабораторные эксперименты
1 этап. (20-80-е годы ХХ века). Основные усилия были
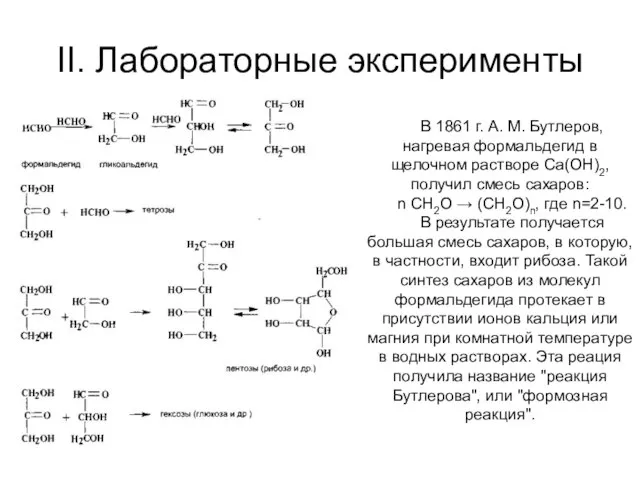
II. Лабораторные эксперименты
В 1861 г. А. М. Бутлеров, нагревая формальдегид в
II. Лабораторные эксперименты
В 1861 г. А. М. Бутлеров, нагревая формальдегид в
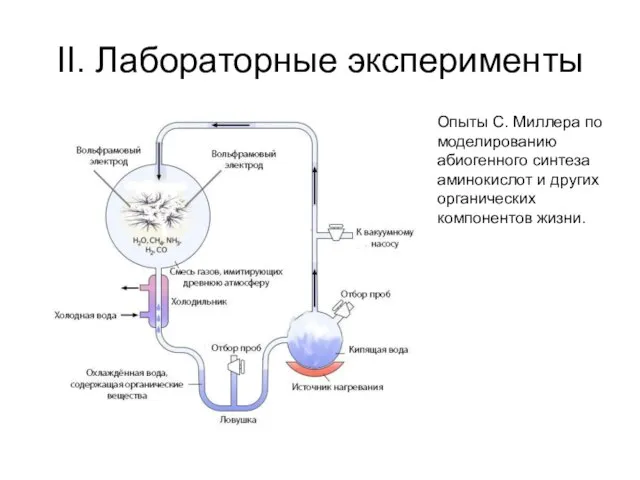
II. Лабораторные эксперименты
Опыты С. Миллера по моделированию абиогенного синтеза аминокислот и
II. Лабораторные эксперименты
Опыты С. Миллера по моделированию абиогенного синтеза аминокислот и

Из опытов С.Миллера
Особенно богатый набор аминокислот получался при моделировании синтеза вблизи
Из опытов С.Миллера
Особенно богатый набор аминокислот получался при моделировании синтеза вблизи
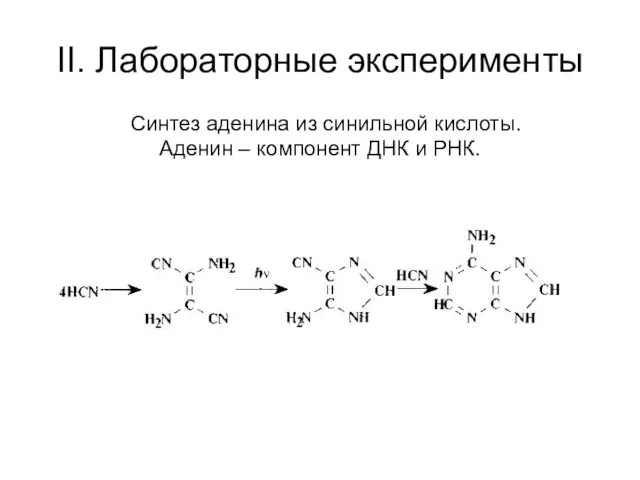
II. Лабораторные эксперименты
Синтез аденина из синильной кислоты.
Аденин – компонент ДНК
II. Лабораторные эксперименты
Синтез аденина из синильной кислоты.
Аденин – компонент ДНК
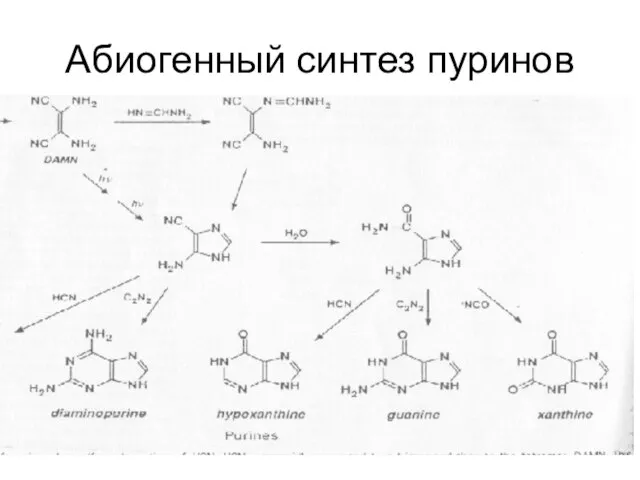
Абиогенный синтез пуринов
Абиогенный синтез пуринов
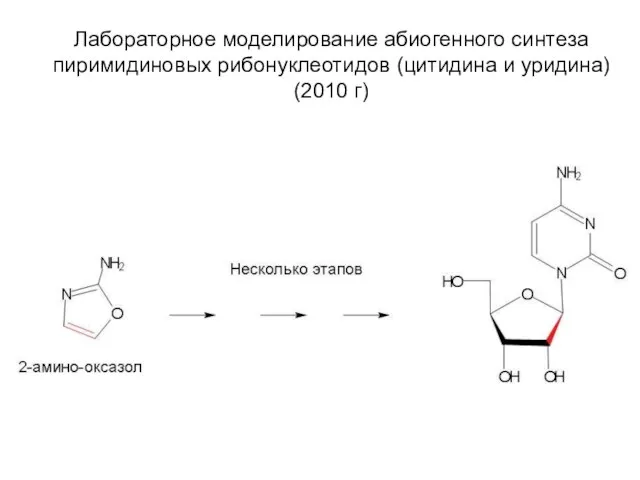
Лабораторное моделирование абиогенного синтеза пиримидиновых рибонуклеотидов (цитидина и уридина) (2010 г)
Лабораторное моделирование абиогенного синтеза пиримидиновых рибонуклеотидов (цитидина и уридина) (2010 г)
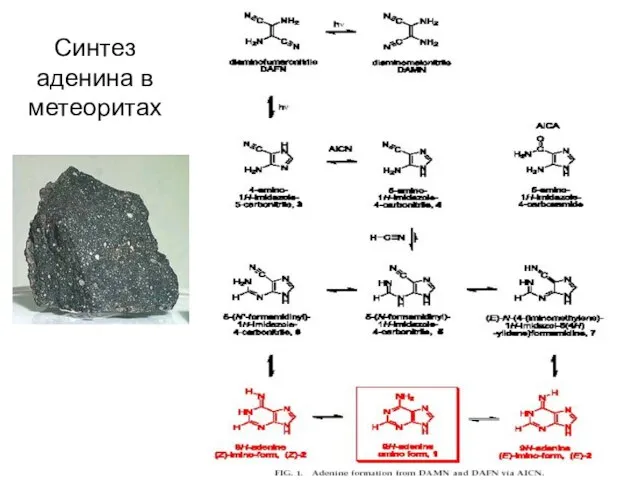
Синтез аденина в метеоритах
Синтез аденина в метеоритах

II. Лабораторные эксперименты
2 этап. В 90-е годы существенную роль стали играть
II. Лабораторные эксперименты
2 этап. В 90-е годы существенную роль стали играть

Позволяет отбирать мутантные молекулы РНК, которые специфически связываются с тем или
Позволяет отбирать мутантные молекулы РНК, которые специфически связываются с тем или
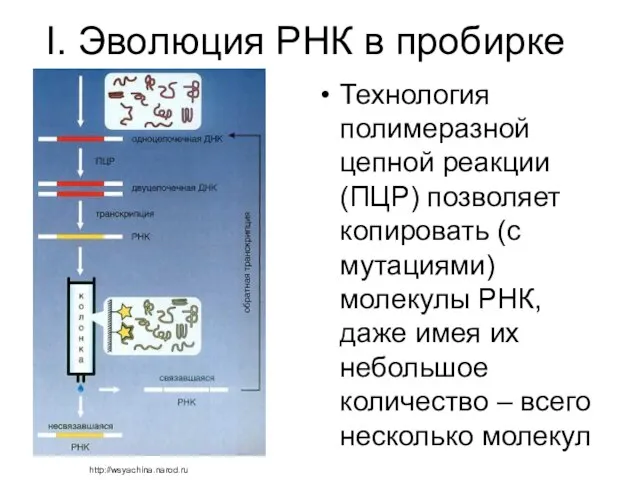
I. Эволюция РНК в пробирке
Технология полимеразной цепной реакции (ПЦР) позволяет копировать
I. Эволюция РНК в пробирке
Технология полимеразной цепной реакции (ПЦР) позволяет копировать

II. Создание минимально работоспособной клетки
Создание небольшого синтетического генома, интегрированного в бактериальную
II. Создание минимально работоспособной клетки
Создание небольшого синтетического генома, интегрированного в бактериальную
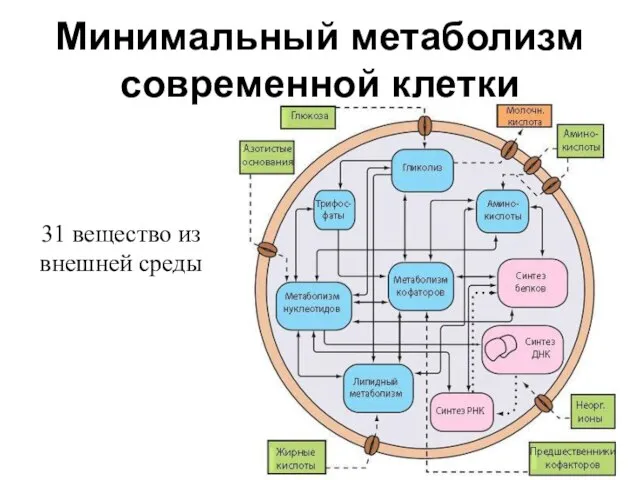
Минимальный метаболизм
современной клетки
31 вещество из внешней среды
Минимальный метаболизм
современной клетки
31 вещество из внешней среды

Минимальная модель современной клетки
Теоретически минимальный набор генов - 241
(102
Минимальная модель современной клетки
Теоретически минимальный набор генов - 241
(102

II. Заключение.
Фактически искусcтвенное моделирование, подобно следственному эксперименту, должно служить важнейшим
II. Заключение.
Фактически искусcтвенное моделирование, подобно следственному эксперименту, должно служить важнейшим

II. Анализ молекулярных реликтов (молекулярных ископаемых) – самый информативный источник.
«Молекулярное ископаемое»
II. Анализ молекулярных реликтов (молекулярных ископаемых) – самый информативный источник.
«Молекулярное ископаемое»

III. Молекулярные реликты
«Жизнь в своем развитии никогда не пренебрегала тем,
III. Молекулярные реликты
«Жизнь в своем развитии никогда не пренебрегала тем,

III. Молекулярные реликты
Современные биохимические механизмы несут на себе отпечаток эволюционного прошлого
III. Молекулярные реликты
Современные биохимические механизмы несут на себе отпечаток эволюционного прошлого
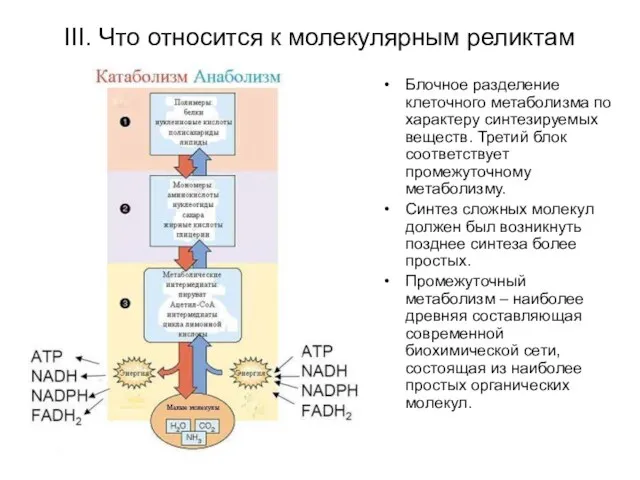
III. Что относится к молекулярным реликтам
Блочное разделение клеточного метаболизма по характеру
III. Что относится к молекулярным реликтам
Блочное разделение клеточного метаболизма по характеру
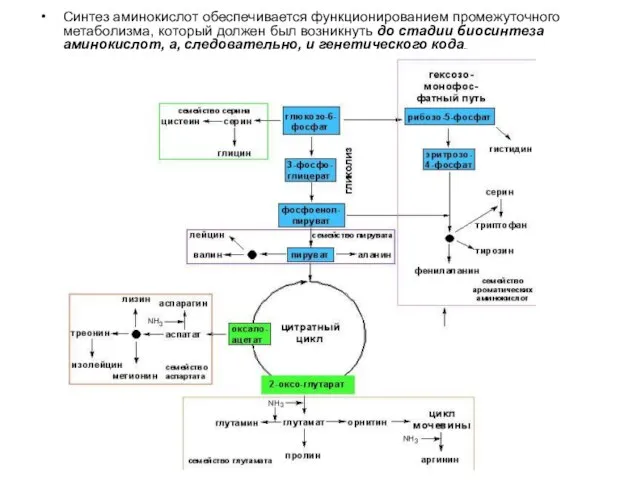
Синтез аминокислот обеспечивается функционированием промежуточного метаболизма, который должен был возникнуть до
Синтез аминокислот обеспечивается функционированием промежуточного метаболизма, который должен был возникнуть до

III. Молекулярные реликты
" Если эволюция метаболических путей шла путем последовательного
III. Молекулярные реликты
" Если эволюция метаболических путей шла путем последовательного
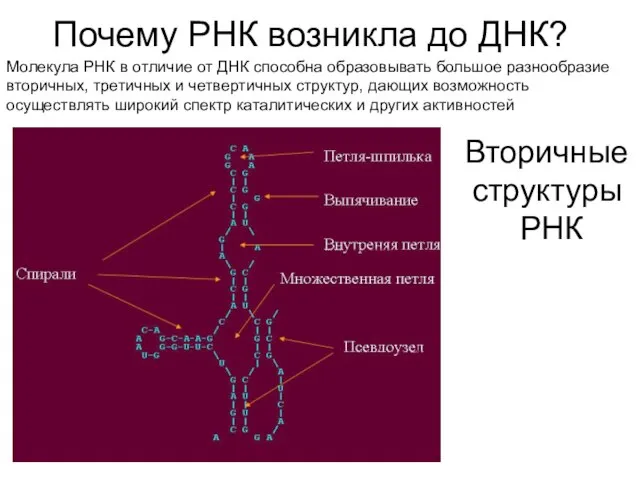
Почему РНК возникла до ДНК?
Молекула РНК в отличие от ДНК способна
Почему РНК возникла до ДНК?
Молекула РНК в отличие от ДНК способна

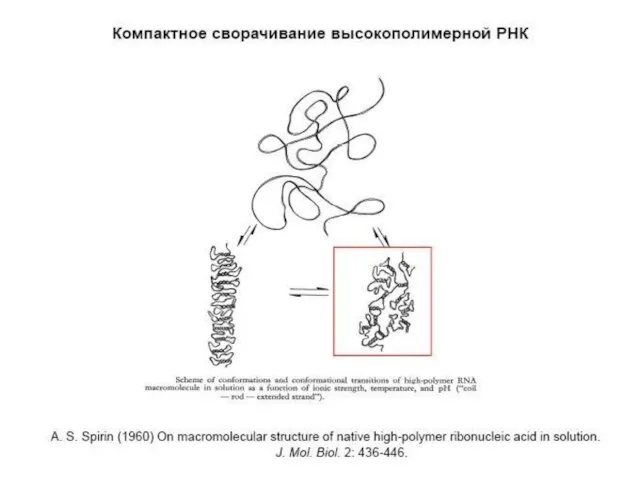
ПРИНЦИПЫ МАКРОМОЛЕКУЛЯРНОЙ СТРУКТУРЫ РНК
РНК, в отличие от ДНК, одноцепочечна.
Высокомолекулярные РНК представляют
ПРИНЦИПЫ МАКРОМОЛЕКУЛЯРНОЙ СТРУКТУРЫ РНК
РНК, в отличие от ДНК, одноцепочечна.
Высокомолекулярные РНК представляют

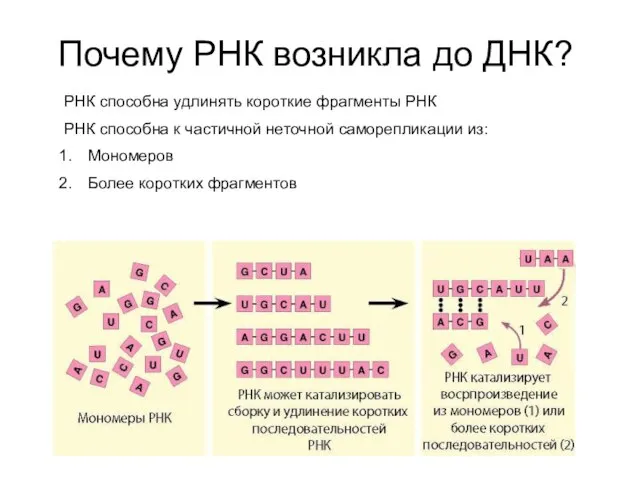
Почему РНК возникла до ДНК?
РНК способна удлинять короткие фрагменты РНК
РНК способна
Почему РНК возникла до ДНК?
РНК способна удлинять короткие фрагменты РНК
РНК способна

Может ли РНК воспроизводить РНК?
Рибозимы могут воспроизводить РНК из мономеров очень
Может ли РНК воспроизводить РНК?
Рибозимы могут воспроизводить РНК из мономеров очень
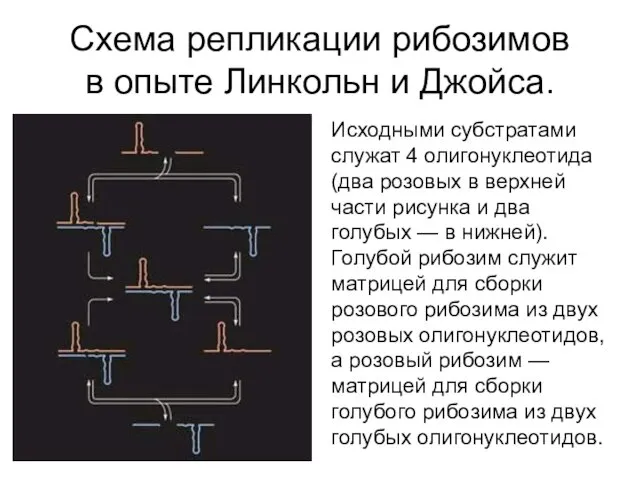
Схема репликации рибозимов в опыте Линкольн и Джойса.
Исходными субстратами служат 4 олигонуклеотида
Схема репликации рибозимов в опыте Линкольн и Джойса.
Исходными субстратами служат 4 олигонуклеотида

Почему РНК возникла до ДНК?
Синтез рибонуклеотидов предшествует синтезу дезкоксирибонуклеотидов
Сначала синтезируются
Почему РНК возникла до ДНК?
Синтез рибонуклеотидов предшествует синтезу дезкоксирибонуклеотидов
Сначала синтезируются

Почему РНК возникла до ДНК?
Репликация РНК, которую можно наблюдать в
Почему РНК возникла до ДНК?
Репликация РНК, которую можно наблюдать в

Почему РНК возникла до ДНК?
В опытах по моделированию абиогенного синтеза рибоза
Почему РНК возникла до ДНК?
В опытах по моделированию абиогенного синтеза рибоза

РНК-кофакторы – реликты мира РНК, существовавшего до появления ДНК.
Кофактор (синонимы -
РНК-кофакторы – реликты мира РНК, существовавшего до появления ДНК.
Кофактор (синонимы -

III. Молекулярные реликты
Хотя отсутствуют прямые данные о существовании РНКовых организмов, «лицо»
III. Молекулярные реликты
Хотя отсутствуют прямые данные о существовании РНКовых организмов, «лицо»

III. Молекулярные реликты
Кодирующие макромолекулы (ДНК и РНК) эволюционируют с разными скоростями.
III. Молекулярные реликты
Кодирующие макромолекулы (ДНК и РНК) эволюционируют с разными скоростями.
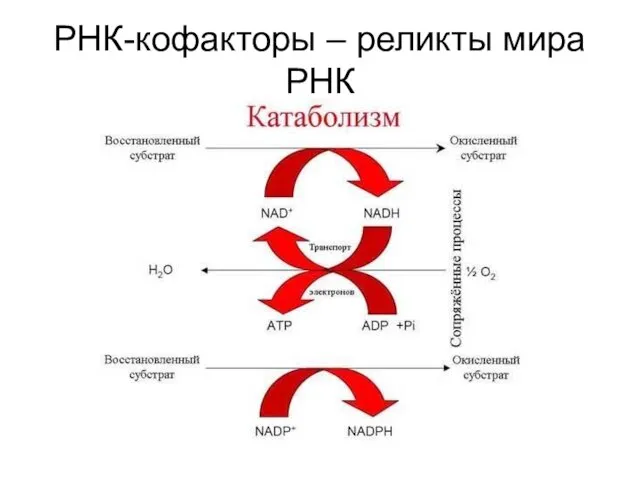
РНК-кофакторы – реликты мира РНК
РНК-кофакторы – реликты мира РНК
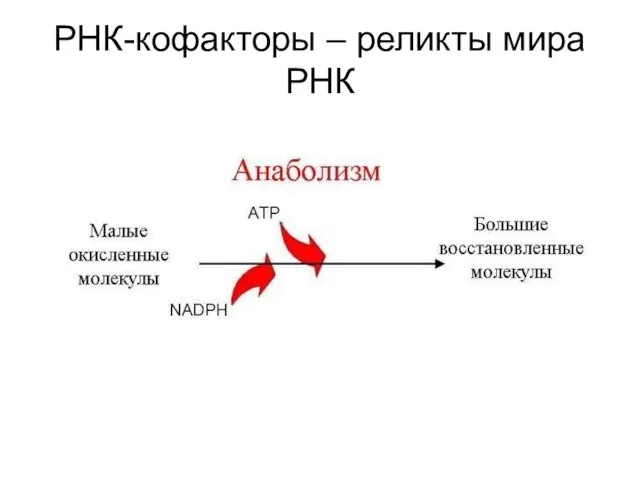
РНК-кофакторы – реликты мира РНК
РНК-кофакторы – реликты мира РНК
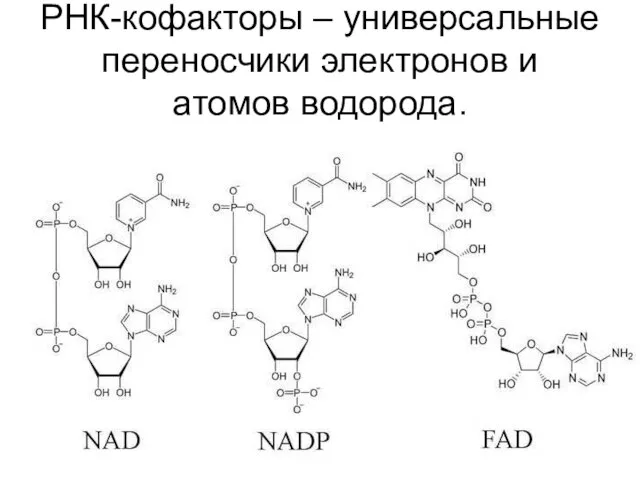
РНК-кофакторы – универсальные переносчики электронов и атомов водорода.
РНК-кофакторы – универсальные переносчики электронов и атомов водорода.
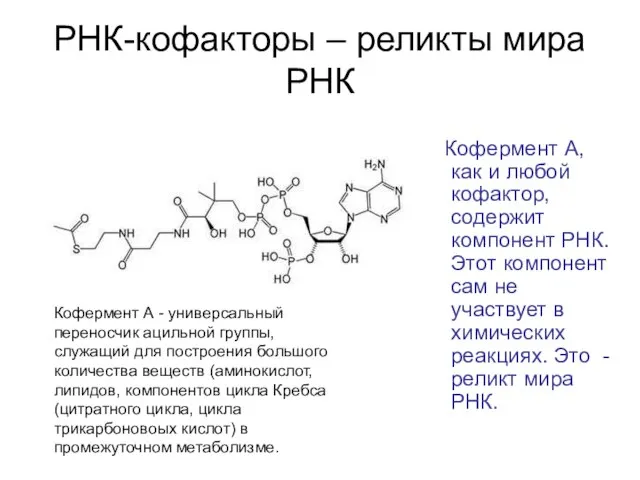
РНК-кофакторы – реликты мира РНК
Кофермент А, как и любой кофактор,
РНК-кофакторы – реликты мира РНК
Кофермент А, как и любой кофактор,
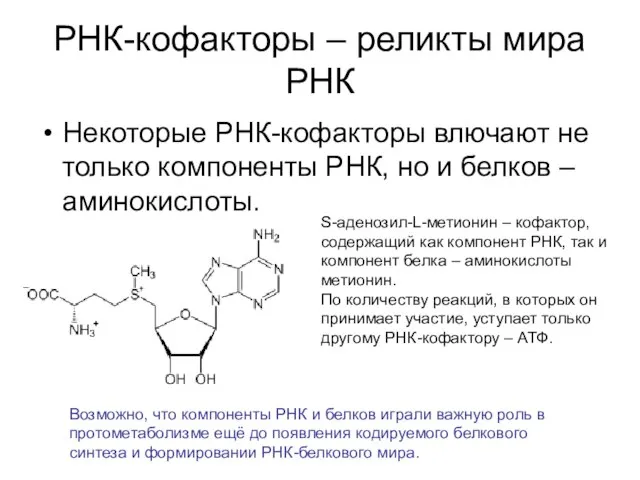
РНК-кофакторы – реликты мира РНК
Некоторые РНК-кофакторы влючают не только компоненты РНК,
РНК-кофакторы – реликты мира РНК
Некоторые РНК-кофакторы влючают не только компоненты РНК,

В чём заключается универсальность РНК-кофактов?
В использовании универсальной энергетической валюты – АТФ
В чём заключается универсальность РНК-кофактов?
В использовании универсальной энергетической валюты – АТФ

Современный метаболизм как палимпсест мира РНК
Палимпсест означает то, что
Современный метаболизм как палимпсест мира РНК
Палимпсест означает то, что

РНК-кофакторы. Резюме.
РНК-кофакторы являлись ключевыми компонентами в биохимических процессах ещё до возникновения
РНК-кофакторы. Резюме.
РНК-кофакторы являлись ключевыми компонентами в биохимических процессах ещё до возникновения
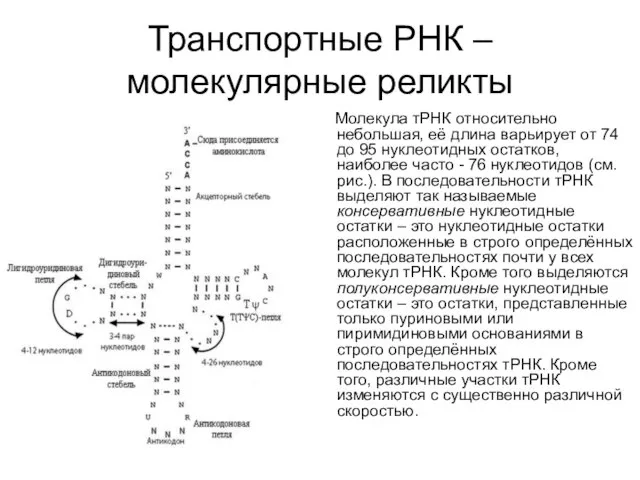
Транспортные РНК – молекулярные реликты
Молекула тРНК относительно небольшая, её длина
Транспортные РНК – молекулярные реликты
Молекула тРНК относительно небольшая, её длина
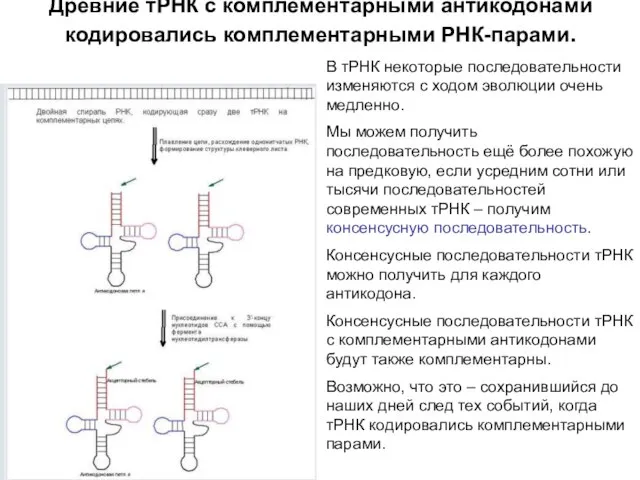
Древние тРНК с комплементарными антикодонами кодировались комплементарными РНК-парами.
В тРНК некоторые последовательности
Древние тРНК с комплементарными антикодонами кодировались комплементарными РНК-парами.
В тРНК некоторые последовательности

III. Другие молекулярные реликты.
Рибосома – одна из самых древних структур, возникшая
III. Другие молекулярные реликты.
Рибосома – одна из самых древних структур, возникшая

Ключевое открытие сравнительной геномики
Типичное время исчезновения сходства последовательностей у гомологичных генов
Ключевое открытие сравнительной геномики
Типичное время исчезновения сходства последовательностей у гомологичных генов
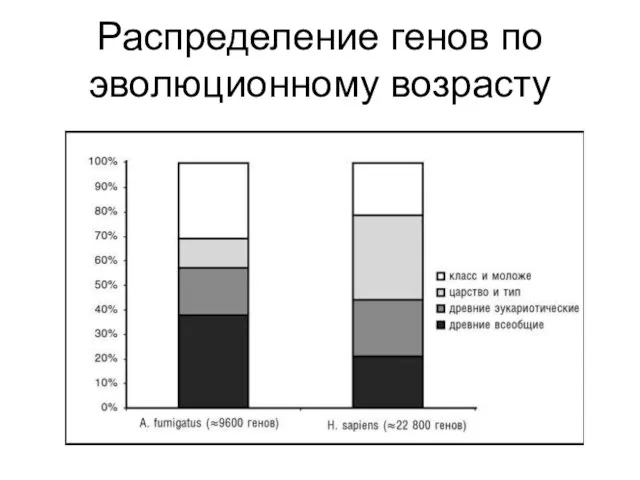
Распределение генов по эволюционному возрасту
Распределение генов по эволюционному возрасту

Ядро, оболочка и облако
ЯДРО - (Почти) универсальные гены, те, что представлены
Ядро, оболочка и облако
ЯДРО - (Почти) универсальные гены, те, что представлены

Ядро, оболочка и облако
ОБОЛОЧКА - Умеренно консервативная генная оболочка состоит из
Ядро, оболочка и облако
ОБОЛОЧКА - Умеренно консервативная генная оболочка состоит из

Ядро, оболочка и облако
ОБЛАКО. Гены «облака» составляют переменную долю в каждом
Ядро, оболочка и облако
ОБЛАКО. Гены «облака» составляют переменную долю в каждом
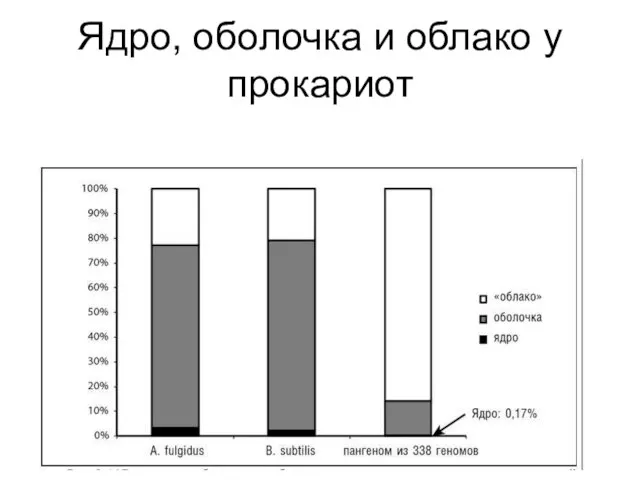
Ядро, оболочка и облако у прокариот
Ядро, оболочка и облако у прокариот

Прогенот – коллективная форма жизни.
До появления ДНК живые системы не дивергировали
Прогенот – коллективная форма жизни.
До появления ДНК живые системы не дивергировали
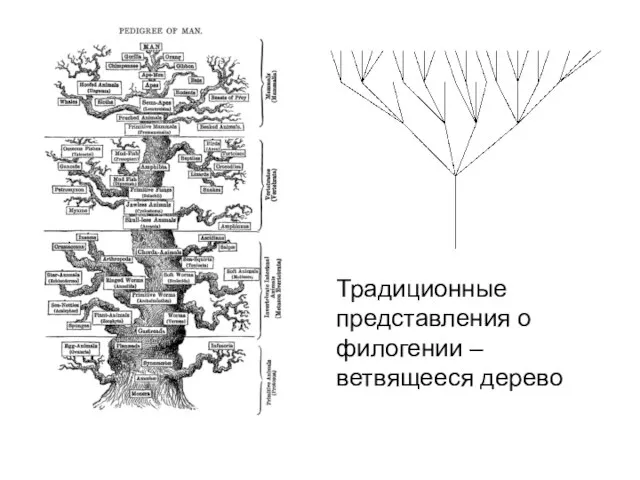
Традиционные представления о филогении – ветвящееся дерево
Традиционные представления о филогении – ветвящееся дерево
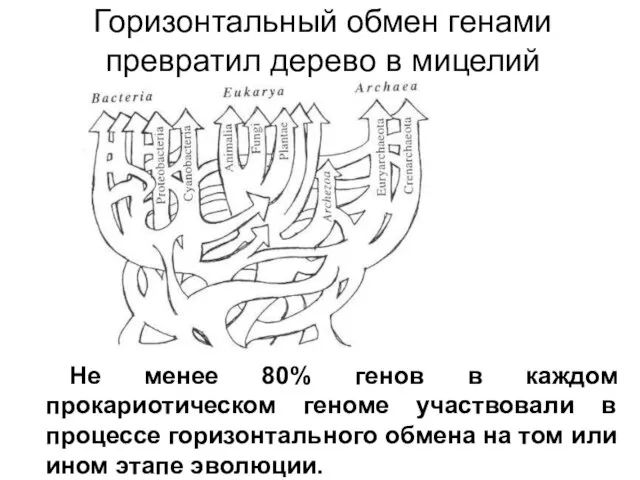
Горизонтальный обмен генами превратил дерево в мицелий
Не менее 80% генов в
Горизонтальный обмен генами превратил дерево в мицелий
Не менее 80% генов в
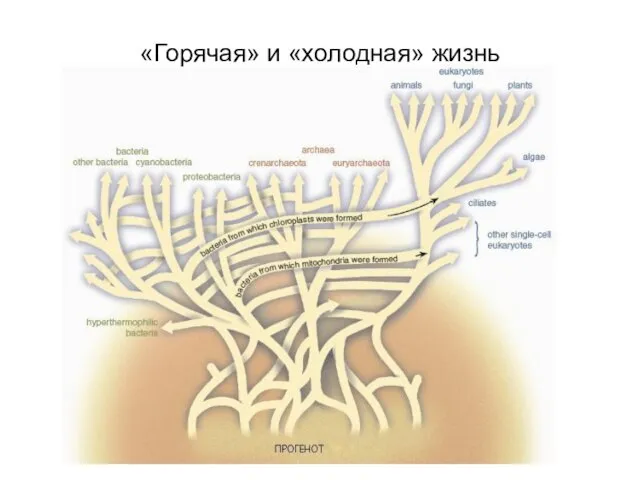
«Горячая» и «холодная» жизнь
«Горячая» и «холодная» жизнь


IV. Анализ геологических данных.
Они дают два вида информации.
IV. Анализ геологических данных.
Они дают два вида информации.

Период массовых метеоритных бомбардировок
Период массовых метеоритных бомбардировок

IV. Анализ геологических данных.
Геологические исследования позволяют также находить так называемые
IV. Анализ геологических данных.
Геологические исследования позволяют также находить так называемые

Геологические данные позволяют воссоздать условия на Земле в момент возникновения жизни.
Ничтожные
Геологические данные позволяют воссоздать условия на Земле в момент возникновения жизни.
Ничтожные

Условия на первобытной Земле
Условия на первобытной Земле
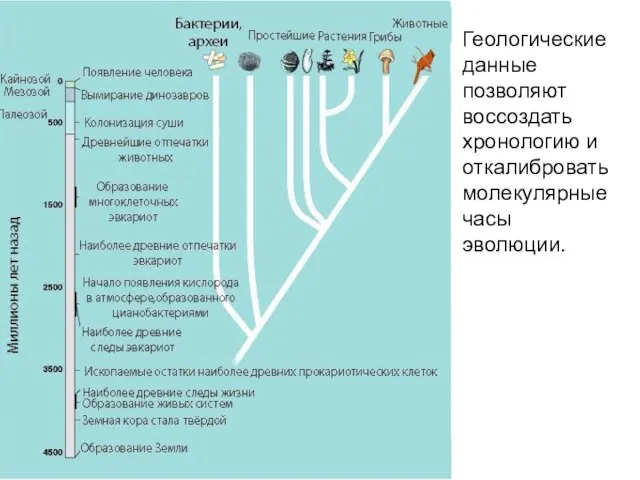
Геологические данные позволяют воссоздать хронологию и откалибровать молекулярные часы эволюции.
Геологические данные позволяют воссоздать хронологию и откалибровать молекулярные часы эволюции.

Строматолиты — ископаемые остатки цианобактериальных матов.
Строматолиты — ископаемые остатки цианобактериальных матов.

Древняя глобальная биосфера
Была представлена бактериальными матами
Древняя глобальная биосфера
Была представлена бактериальными матами


Окаменелости цианобактерий
(3,5 млрд. лет)
Окаменелости цианобактерий
(3,5 млрд. лет)
 Насекомые. Отряд таракановые
Насекомые. Отряд таракановые Пути эволюции силуэта породы немецкая овчарка. Тупиковые пути
Пути эволюции силуэта породы немецкая овчарка. Тупиковые пути Биохимический мониторинг миокардитов у цыплят-бройлеров и алгоритмы разработки схем фармакопрофилактики
Биохимический мониторинг миокардитов у цыплят-бройлеров и алгоритмы разработки схем фармакопрофилактики Презентация на тему "Зрительный анализатор" - скачать презентации по Биологии
Презентация на тему "Зрительный анализатор" - скачать презентации по Биологии Поэтапное рисование насекомых
Поэтапное рисование насекомых Жизнедеятельность клетки
Жизнедеятельность клетки Исчезнувшие животные
Исчезнувшие животные Мышцы туловища
Мышцы туловища Какая у нас память Исследовательская работа
Какая у нас память Исследовательская работа  Хемосинтез
Хемосинтез 20160718_vsyo_proydyot_8_klass
20160718_vsyo_proydyot_8_klass «Диатомовые водоросли (Diatomeae) Выполнила: Студентка 3 курса ф-та естествознания Рвачева Е.И.
«Диатомовые водоросли (Diatomeae) Выполнила: Студентка 3 курса ф-та естествознания Рвачева Е.И. Презентация на тему "Завдання та методи селекції" - скачать бесплатно презентации по Биологии
Презентация на тему "Завдання та методи селекції" - скачать бесплатно презентации по Биологии Особенности высшей нервной деятельности
Особенности высшей нервной деятельности Сердечно-сосудистая, дыхательная и нервная системы
Сердечно-сосудистая, дыхательная и нервная системы Рецепторы гормонов и передача сигналов: рецепторы клеточной поверхности, внутриклеточные рецепторы
Рецепторы гормонов и передача сигналов: рецепторы клеточной поверхности, внутриклеточные рецепторы Презентация на тему Развитие жизни на Земле. Эволюция человека
Презентация на тему Развитие жизни на Земле. Эволюция человека Царство Грибы. Общая характеристика. Многообразие и значение
Царство Грибы. Общая характеристика. Многообразие и значение Животные пустыни
Животные пустыни Домашние птицы
Домашние птицы Ю. Герман. К 110-летию со дня рождения писателя
Ю. Герман. К 110-летию со дня рождения писателя Конкурс презентаций «Интерактивная мозаика» Конкурс презентаций «Интерактивная мозаика» Номинация «Интерактивный кроссворд»
Конкурс презентаций «Интерактивная мозаика» Конкурс презентаций «Интерактивная мозаика» Номинация «Интерактивный кроссворд»  Микрофлора, дисбиозы
Микрофлора, дисбиозы Процессы жизнедеятельности одноклеточных животных 11класс 3 урок Автор: Кудяшева Вера Григорьевна, учитель биологии МОУ СОШ №12 С
Процессы жизнедеятельности одноклеточных животных 11класс 3 урок Автор: Кудяшева Вера Григорьевна, учитель биологии МОУ СОШ №12 С Мое хобби. Комнатные цветы
Мое хобби. Комнатные цветы “Куріння”
“Куріння”  Ткани. Типы тканей и их свойства.
Ткани. Типы тканей и их свойства. Паразитизм
Паразитизм