- Фізіологія збудливих тканин

Содержание
- 2. Література: Покровский В. М. Физиология человека. / В.М. Покровский, Г. Ф. Коротько. - М.: Медицина, 1997;
- 3. Фізіологія (от греч. physis — природа и logos - учение) – наука, що вивчає життєдіяльність (функціонування)
- 4. Напрямки фізіології
- 5. Методи фізіологічних досліджень 1. Експеримент Гострий (вівісекція); В умовах ізольованого органу; Хронічний (І.П. Павлов); 2.Графічна реєстрація
- 6. Об'єктивна графічна реєстрація біоелектричних потенціалів стала основою важливого розділу фізіології - електрофізіології.
- 9. УКРАЇНСЬКА ШКОЛА ФІЗІОЛОГІВ Чаговець В.Ю (1873-1941) Воронцов Д.С. (1886-1965) Костюк П. Г. (1924-2010) Щелков І. П.
- 10. Клетка– основная структурная и функциональная единица организма. Она способна к самообновлению, саморегуляции и самовоспроизведению. Клетки объединяются
- 11. Основні властивості живих клітин Подразливість – здатність клітини активно реагувати на дію факторів зовнішнього чи внутрішнього
- 12. Подразнення – вплив подразника на організм. Збудливість - це здатність збудливих тканин реагувати на подразнення збудженням.
- 13. Протилежний стан збудженню збудливих тканин називається гальмуванням. Гальмування - це активний процес, що супроводжується специфічними змінами
- 14. Протилежний стан збудженню збудливих тканин називається гальмуванням. Гальмування - це активний процес, що супроводжується специфічними змінами
- 15. Будова клітинної мембрани Жидкостно-мозаичная модель (Сингера и Николсона) – белки погружены в фосфолипидный бислой. Липидный бислой
- 16. Трехмерная жидкостно-мозаичная модель клеточной мембраны (по Сингеру—Николсону). А — фосфолипидный бислой, в который погружены белки; Б
- 17. Основні функції клітинної мембрани Бар'єрна; Транспортна; Регуляторна; Контактна; Рецепторна; Секреторна.
- 18. Мембранний потенціал спокою (МПС) – це різниця потенціалів між зовнішньою і внутрішньою сторонами мембрани. Суть теорії
- 19. Умови виникнення МПС • 1. Наличие градиента концентраций для ионов K+, Na+, Cl-; • 2. Избирательно
- 20. 1. Электрохимический градиент для ионов градиент концентрации ионов по обе стороны мембраны (концентрация К+ внутри клетки
- 21. 2. Высокая избирательная проницаемость мембраны для ионов К+, Na+, Cl- В состоянии покоя мембрана проницательна для
- 22. 3. Наличие активного транспорта (Nа+,К+ насоса) в мембране Принцип работы. В клеточной мембране имеется система переносчиков,
- 23. Потенціал дії (ПД) ПД - швидке високоамплітудне коливання МПС, що виникає під час збудження (від -90
- 24. Тривалість ПД 0,3-10 мс
- 25. Фази ПД під час висхідної фази (фаза деполяризації) відбувається не просто зникнення МПС, а виникає різниця
- 26. Тривалість піка потенціалу дії у різних нервових і скелетних м'язових волокон варіює від 0,5 до 3
- 27. ПД виникає в той момент, коли деполяризація мембрани досягає критичного рівня деполяризації. Якщо сила подразника буде
- 28. Відмінності між ПД і ЛВ 1. ПД не залежить від сили подразника, його амплітуда завжди постійна,
- 29. Під час розвитку ПД, виникає рефрактерність (незбудливість) - зниження здатності клітини відповідати на подразнення в результаті
- 30. МАКСИМАЛЬНИЙ РИТМ ІМПУЛЬСАЦІЇ М. Є. Введенський перший звернув увагу на різну здатність збудливих тканин відтворювати високі
- 31. Механізм проведення збудження Проведення збуження вздовж нервових волокон здійснюється за допомогою місцевих (локальних) струмів (Герман, 1899),
- 32. Збільшення швидкості проведення збудження відбувається за рахунок мієлінізації волокон. Мієлінова оболонка виконує роль ізолятора, який переривається
- 33. Закони проведення збудження Закон анатомічної і фізіологічної цілісності волокна - збудження може поширюватися по нервовому волокну
- 34. Фізіологія синапсів
- 35. Синапс – (від грец. synapsis - дотик, з'єднання) місце спеціалізованого контакту між нервовими клітинами або між
- 36. 1. Классификация синапсов по контакту (местонахождению) Нейрон-нейрон нейрон-эффектор (руховий нерв закінчується на волокні скелетного м'яза) -аксон-дендрит
- 38. 3. Классификация синапсов по способу передачи химические электрические смешанные Их большинство простой переход передача и Передача
- 39. Будова хімічного (нервово-м’язового) синапсу Пресинаптична частина Синаптична щілина Постсинаптична частина
- 40. Механізм передачі через синапс 1. ПД по аксону надходить до пресинаптичної мембрани і викликає її деполяризацію
- 41. Нейромедиаторы (нейротрансмиттеры) — биологически активные химические вещества, посредством которых осуществляется передача электрического импульса с нервной клетки
- 42. Ацетилхолин – медиатор нервно-мышечного синапса Ацетилхолин выполняет функцию медиатора в синапсах периферической и ЦНС: Ах выделяется
- 43. γ-Аминомасляная кислота (ГАМК, GABA) Аминокислота, важнейший тормозной нейромедиатор центральной нервной системы человека и млекопитающих. γ-Аминомасляная кислота
- 44. Особливості проведення інформації через хімічний синапс 1. Однобічність проведення. 2.Затримка проведення (Час від надходження нервового імпульсу
- 46. Скачать презентацию

Література:
Покровский В. М. Физиология человека. / В.М. Покровский, Г. Ф. Коротько.
Література:
Покровский В. М. Физиология человека. / В.М. Покровский, Г. Ф. Коротько.

Фізіологія (от греч. physis — природа и logos - учение) –
Фізіологія (от греч. physis — природа и logos - учение) –

Напрямки фізіології
Напрямки фізіології
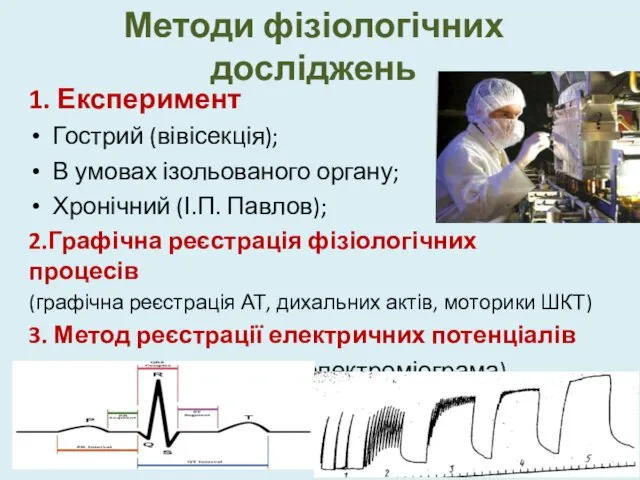
Методи фізіологічних досліджень
1. Експеримент
Гострий (вівісекція);
В умовах ізольованого органу;
Хронічний (І.П. Павлов);
2.Графічна
Методи фізіологічних досліджень
1. Експеримент
Гострий (вівісекція);
В умовах ізольованого органу;
Хронічний (І.П. Павлов);
2.Графічна

Об'єктивна графічна реєстрація біоелектричних потенціалів стала основою важливого розділу фізіології -
Об'єктивна графічна реєстрація біоелектричних потенціалів стала основою важливого розділу фізіології -



УКРАЇНСЬКА ШКОЛА ФІЗІОЛОГІВ
Чаговець В.Ю (1873-1941)
Воронцов Д.С.
(1886-1965)
Костюк П. Г.
(1924-2010)
Щелков І. П.
(1833-1909)
Мечников
УКРАЇНСЬКА ШКОЛА ФІЗІОЛОГІВ
Чаговець В.Ю (1873-1941)
Воронцов Д.С.
(1886-1965)
Костюк П. Г.
(1924-2010)
Щелков І. П.
(1833-1909)
Мечников
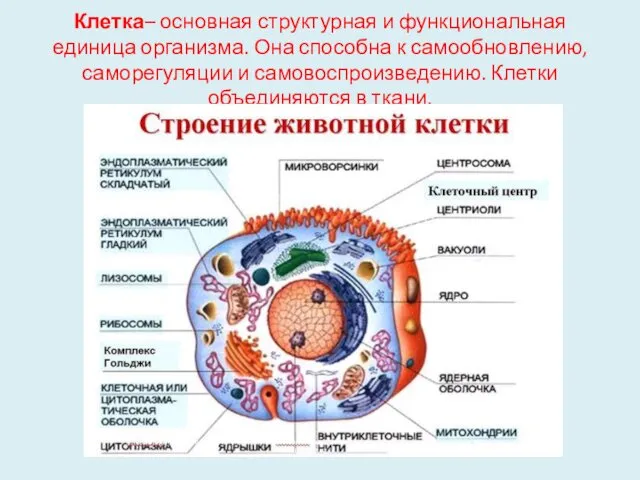
Клетка– основная структурная и функциональная единица организма. Она способна к самообновлению,
Клетка– основная структурная и функциональная единица организма. Она способна к самообновлению,

Основні властивості живих клітин
Подразливість – здатність клітини активно реагувати на дію
Основні властивості живих клітин
Подразливість – здатність клітини активно реагувати на дію

Подразнення – вплив подразника на організм.
Збудливість - це здатність збудливих тканин
Подразнення – вплив подразника на організм.
Збудливість - це здатність збудливих тканин

Протилежний стан збудженню збудливих тканин називається гальмуванням.
Гальмування - це активний
Протилежний стан збудженню збудливих тканин називається гальмуванням.
Гальмування - це активний

Протилежний стан збудженню збудливих тканин називається гальмуванням.
Гальмування - це активний
Протилежний стан збудженню збудливих тканин називається гальмуванням.
Гальмування - це активний
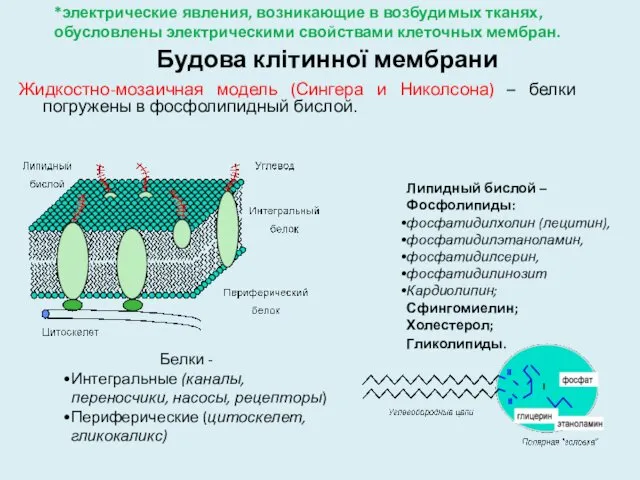
Будова клітинної мембрани
Жидкостно-мозаичная модель (Сингера и Николсона) – белки погружены в
Будова клітинної мембрани
Жидкостно-мозаичная модель (Сингера и Николсона) – белки погружены в
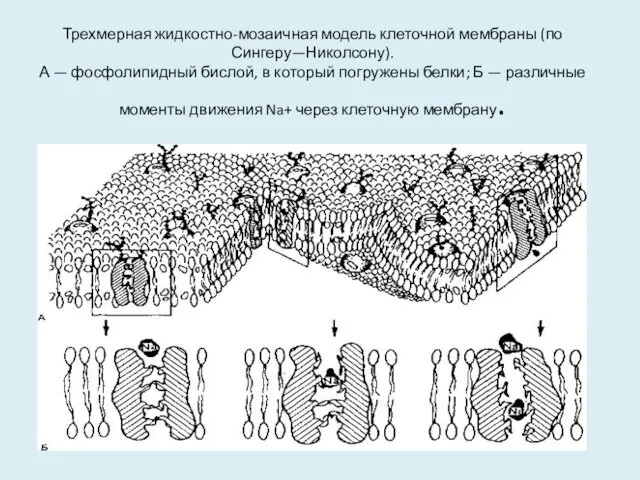
Трехмерная жидкостно-мозаичная модель клеточной мембраны (по Сингеру—Николсону).
А — фосфолипидный бислой, в
Трехмерная жидкостно-мозаичная модель клеточной мембраны (по Сингеру—Николсону). А — фосфолипидный бислой, в
Основні функції клітинної мембрани
Бар'єрна;
Транспортна;
Регуляторна;
Контактна;
Рецепторна;
Секреторна.
Основні функції клітинної мембрани
Бар'єрна;
Транспортна;
Регуляторна;
Контактна;
Рецепторна;
Секреторна.

Мембранний потенціал спокою (МПС) – це різниця потенціалів між зовнішньою і
Мембранний потенціал спокою (МПС) – це різниця потенціалів між зовнішньою і

Умови виникнення МПС
• 1. Наличие градиента концентраций для ионов K+, Na+,
Умови виникнення МПС
• 1. Наличие градиента концентраций для ионов K+, Na+,

1. Электрохимический градиент для ионов
градиент концентрации ионов по обе стороны мембраны
1. Электрохимический градиент для ионов
градиент концентрации ионов по обе стороны мембраны

2. Высокая избирательная проницаемость
мембраны для ионов К+, Na+, Cl-
В состоянии покоя
2. Высокая избирательная проницаемость
мембраны для ионов К+, Na+, Cl-
В состоянии покоя
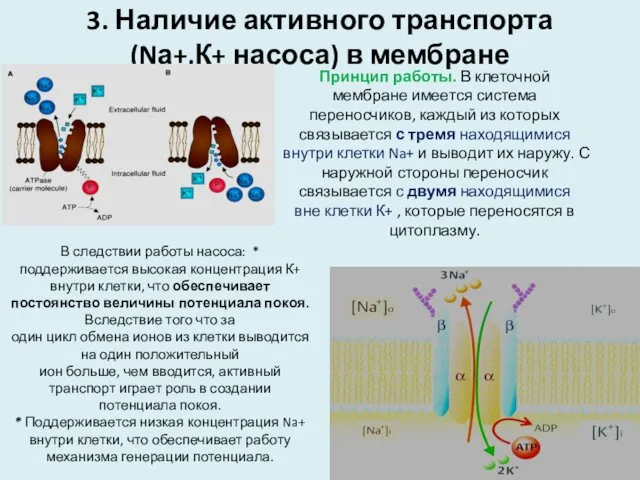
3. Наличие активного транспорта
(Nа+,К+ насоса) в мембране
Принцип работы. В клеточной мембране
3. Наличие активного транспорта
(Nа+,К+ насоса) в мембране
Принцип работы. В клеточной мембране

Потенціал дії (ПД)
ПД - швидке високоамплітудне коливання МПС, що виникає під
Потенціал дії (ПД)
ПД - швидке високоамплітудне коливання МПС, що виникає під
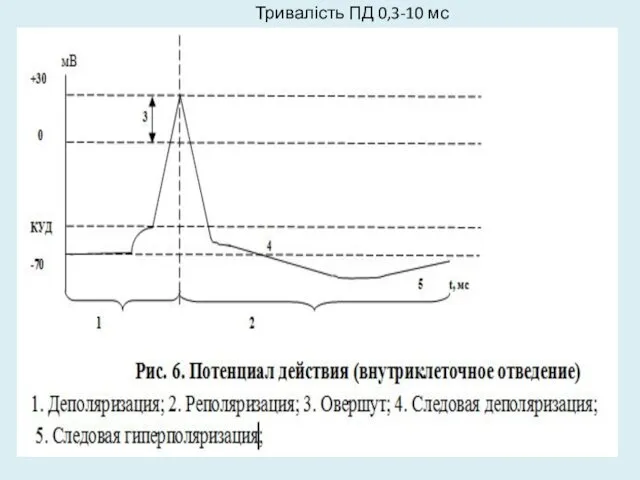
Тривалість ПД 0,3-10 мс
Тривалість ПД 0,3-10 мс

Фази ПД
під час висхідної фази (фаза деполяризації) відбувається не просто зникнення
Фази ПД
під час висхідної фази (фаза деполяризації) відбувається не просто зникнення

Тривалість піка потенціалу дії у різних нервових і скелетних м'язових волокон
Тривалість піка потенціалу дії у різних нервових і скелетних м'язових волокон
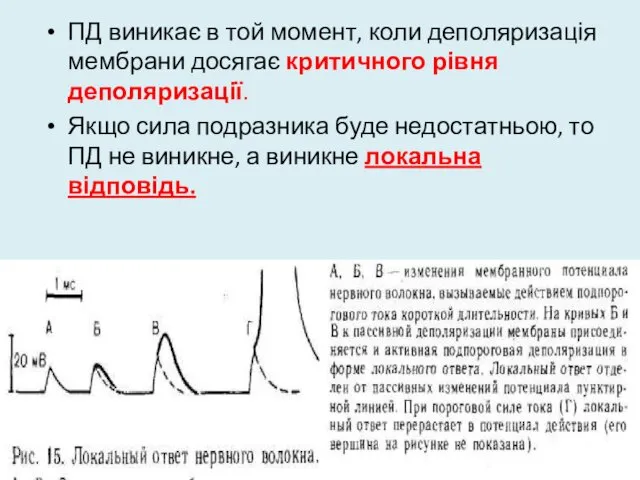
ПД виникає в той момент, коли деполяризація мембрани досягає критичного рівня
ПД виникає в той момент, коли деполяризація мембрани досягає критичного рівня

Відмінності між ПД і ЛВ
1. ПД не залежить від сили
Відмінності між ПД і ЛВ
1. ПД не залежить від сили

Під час розвитку ПД, виникає рефрактерність (незбудливість) - зниження здатності клітини
Під час розвитку ПД, виникає рефрактерність (незбудливість) - зниження здатності клітини

МАКСИМАЛЬНИЙ РИТМ ІМПУЛЬСАЦІЇ
М. Є. Введенський перший звернув увагу на різну здатність
МАКСИМАЛЬНИЙ РИТМ ІМПУЛЬСАЦІЇ
М. Є. Введенський перший звернув увагу на різну здатність

Механізм проведення збудження
Проведення збуження вздовж нервових волокон здійснюється за допомогою місцевих
Механізм проведення збудження
Проведення збуження вздовж нервових волокон здійснюється за допомогою місцевих
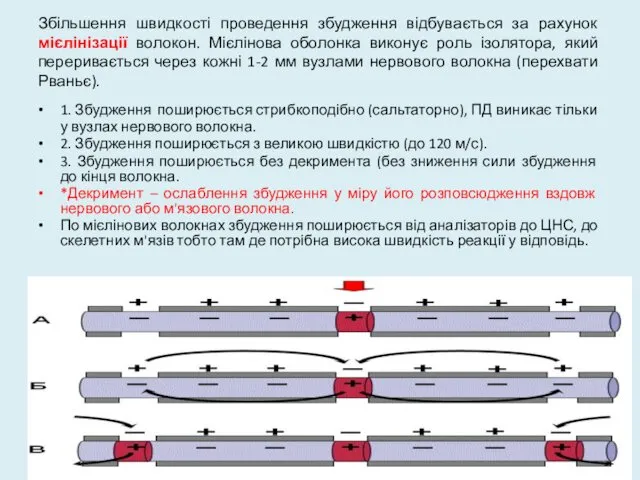
Збільшення швидкості проведення збудження відбувається за рахунок мієлінізації волокон. Мієлінова оболонка
Збільшення швидкості проведення збудження відбувається за рахунок мієлінізації волокон. Мієлінова оболонка

Закони проведення збудження
Закон анатомічної і фізіологічної цілісності волокна - збудження може
Закони проведення збудження
Закон анатомічної і фізіологічної цілісності волокна - збудження може
Фізіологія синапсів
Фізіологія синапсів
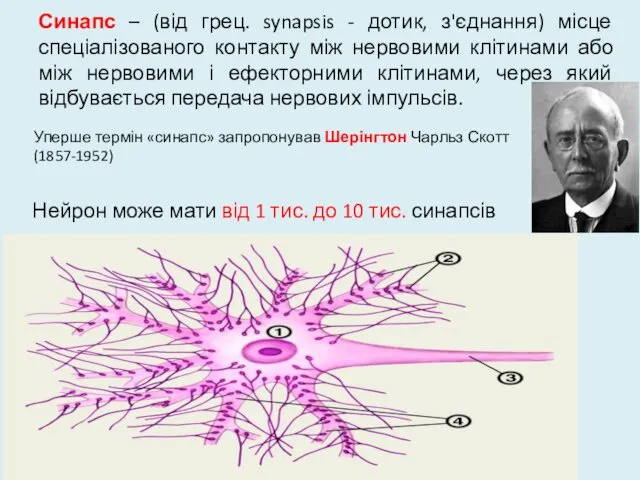
Синапс – (від грец. synapsis - дотик, з'єднання) місце спеціалізованого контакту
Синапс – (від грец. synapsis - дотик, з'єднання) місце спеціалізованого контакту
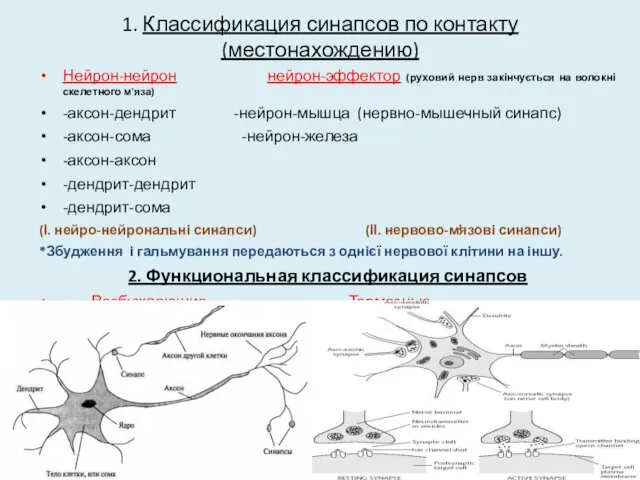
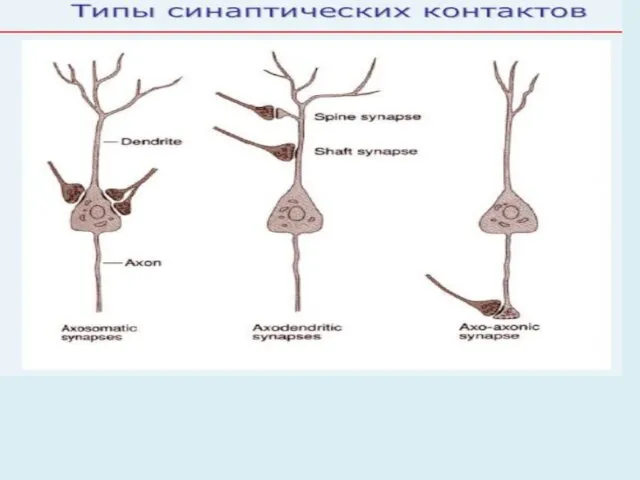
1. Классификация синапсов по контакту (местонахождению)
Нейрон-нейрон нейрон-эффектор (руховий нерв закінчується на
1. Классификация синапсов по контакту (местонахождению)
Нейрон-нейрон нейрон-эффектор (руховий нерв закінчується на
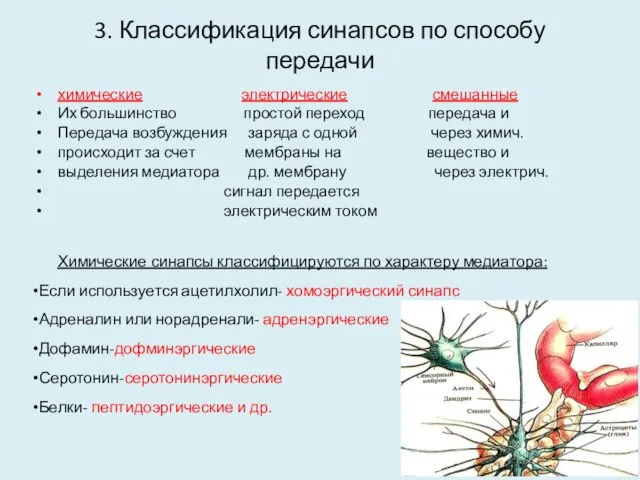
3. Классификация синапсов по способу передачи
химические электрические смешанные
Их большинство простой переход
3. Классификация синапсов по способу передачи
химические электрические смешанные
Их большинство простой переход
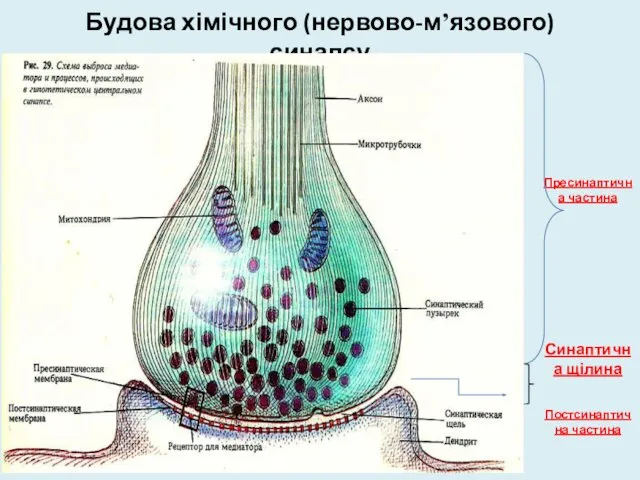
Будова хімічного (нервово-м’язового) синапсу
Пресинаптична частина
Синаптична щілина
Постсинаптична частина
Будова хімічного (нервово-м’язового) синапсу
Пресинаптична частина
Синаптична щілина
Постсинаптична частина
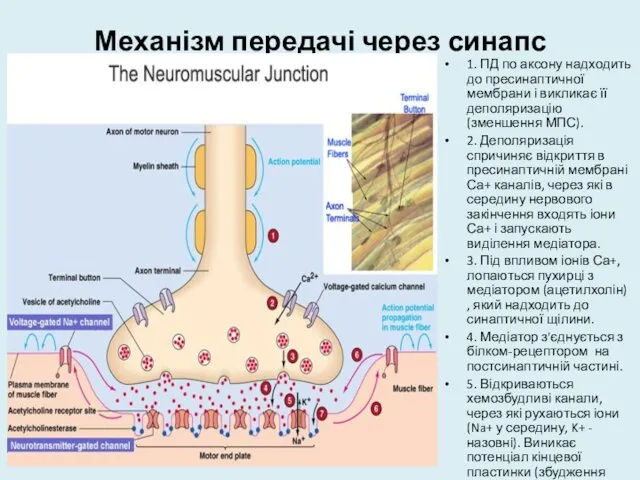
Механізм передачі через синапс
1. ПД по аксону надходить до пресинаптичної мембрани
Механізм передачі через синапс
1. ПД по аксону надходить до пресинаптичної мембрани

Нейромедиаторы (нейротрансмиттеры)
— биологически активные химические вещества, посредством которых осуществляется передача
Нейромедиаторы (нейротрансмиттеры)
— биологически активные химические вещества, посредством которых осуществляется передача

Ацетилхолин – медиатор нервно-мышечного синапса
Ацетилхолин выполняет функцию медиатора в синапсах периферической
Ацетилхолин – медиатор нервно-мышечного синапса
Ацетилхолин выполняет функцию медиатора в синапсах периферической
γ-Аминомасляная кислота (ГАМК, GABA)
Аминокислота, важнейший тормозной нейромедиатор центральной нервной системы
γ-Аминомасляная кислота (ГАМК, GABA)
Аминокислота, важнейший тормозной нейромедиатор центральной нервной системы

Особливості проведення інформації через хімічний синапс
1. Однобічність проведення.
2.Затримка проведення (Час від
Особливості проведення інформації через хімічний синапс
1. Однобічність проведення.
2.Затримка проведення (Час від
 Круговорот углекислого газа в природе
Круговорот углекислого газа в природе Домашнее животное – коза. Выполнила учитель начальных классов МОУ «Лазурненская средняя общеобразовательная школа» Лобанчу
Домашнее животное – коза. Выполнила учитель начальных классов МОУ «Лазурненская средняя общеобразовательная школа» Лобанчу Презентация на тему "Разнообразие культурных растений" - скачать презентации по Биологии
Презентация на тему "Разнообразие культурных растений" - скачать презентации по Биологии Амурский тигр
Амурский тигр Тип Саркодовые
Тип Саркодовые Биохимия крови
Биохимия крови Жизненные процессы клетки
Жизненные процессы клетки Единство химической организации живых организмов. Углеводы
Единство химической организации живых организмов. Углеводы Лекарственные растения содержащие эфирные масла
Лекарственные растения содержащие эфирные масла Презентация на тему Взаимоотношения организмов разных видов
Презентация на тему Взаимоотношения организмов разных видов Биосинтез инвертазы при ферментации гидролизата помола зерна ржи штаммом Aspergillus niger Л-4
Биосинтез инвертазы при ферментации гидролизата помола зерна ржи штаммом Aspergillus niger Л-4 Тестовые задания по теме Наше питание
Тестовые задания по теме Наше питание X Всероссийский турнир юных биологов. Матрёшка. (Задача 4)
X Всероссийский турнир юных биологов. Матрёшка. (Задача 4) Ткани. Типы тканей и их свойства
Ткани. Типы тканей и их свойства Голларктика 2. Неарктическое подцарство
Голларктика 2. Неарктическое подцарство Сердце и сосуды
Сердце и сосуды Система профилактических мероприятий при периодической тимпании телят
Система профилактических мероприятий при периодической тимпании телят Правильное питание школьников. Исследовательский проект по биологии.
Правильное питание школьников. Исследовательский проект по биологии. Технология выращивания посадочного материала
Технология выращивания посадочного материала Травлення в шлунку
Травлення в шлунку Быковская Наталья Владимировна, учитель биологии МОУ «Средняя общеобразовательная школа № 1 с углубленным изучением отдельных
Быковская Наталья Владимировна, учитель биологии МОУ «Средняя общеобразовательная школа № 1 с углубленным изучением отдельных  Бег и сердце
Бег и сердце Биология. Многообразие живых организмов. 7 класс Раздел №3 Царство Растения.
Биология. Многообразие живых организмов. 7 класс Раздел №3 Царство Растения.  Презентация на тему Внутреннее строение земноводных
Презентация на тему Внутреннее строение земноводных Жизненные формы в растительном мире МОУ- СОШ р. п. Пушкино Советский район Саратовская область Учителя биологии, экологии Т. А.
Жизненные формы в растительном мире МОУ- СОШ р. п. Пушкино Советский район Саратовская область Учителя биологии, экологии Т. А. Гаджимустафаева Гаджимустафаева Тамила Шихмагомедовна учитель биологии МБОУ «СОШ №7», г.Когалыма
Гаджимустафаева Гаджимустафаева Тамила Шихмагомедовна учитель биологии МБОУ «СОШ №7», г.Когалыма  Республиканский заочный конкурс детских фотографий В объективе натуралиста. Мир насекомых
Республиканский заочный конкурс детских фотографий В объективе натуралиста. Мир насекомых Презентация по биологии Мамонты
Презентация по биологии Мамонты