- Генетика микроорганизмов

Содержание
- 2. Генетика микроорганизмов
- 3. План занятия 1.Характеристика генетического аппарата бактерий 2.Виды внехромосомного генетического материала бактерий 3.Виды изменчивости бактерий 4.Механизмы передачи
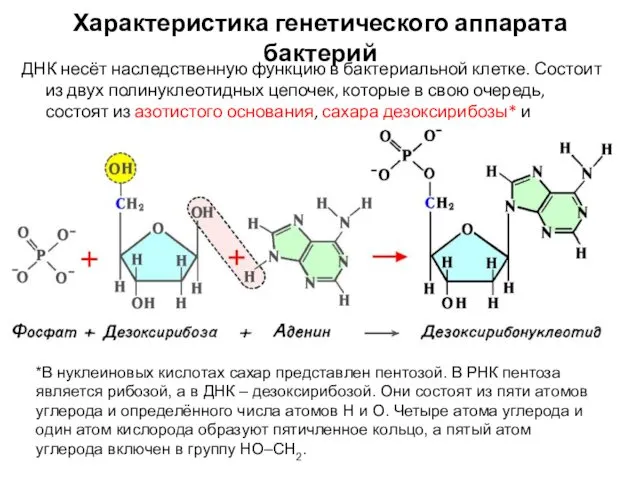
- 4. Характеристика генетического аппарата бактерий ДНК несёт наследственную функцию в бактериальной клетке. Состоит из двух полинуклеотидных цепочек,
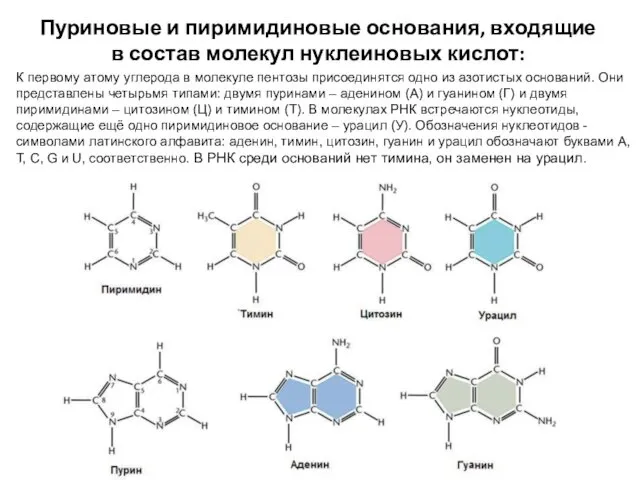
- 5. Пуриновые и пиримидиновые основания, входящие в состав молекул нуклеиновых кислот: К первому атому углерода в молекуле
- 6. Пиримидины (цитозин, тимин и урацил) содержат шестичленные кольца из двух атомов азота и четырех атомов углерода.
- 7. Соединение нуклеотидов в макромолекулу нуклеиновой кислоты происходит путем взаимодействия фосфата одного нуклеотида с гидроксилом пентозы другого
- 8. Последовательность нуклеотидов в цепи молекулы является первичной структурой молекулы ДНК. Соединение цепей друг с другом осуществляется
- 9. Причиной спаривания именно пуринов с пиримидинами является то, что пара из двух пуринов была бы слишком
- 10. Ковалентные связи между атомами в углеводно-фосфатной цепи полинуклеотида имеют определённую пространственную ориентацию, обусловленную так называемыми торсионными
- 11. NB! Таким образом, в структурной организации молекулы ДНК выделяют три уровня: – первичную структуру – последовательность
- 12. https://youtu.be/LdKdjhhMAWY - видео, рекомендованное к просмотру
- 13. Наследственная информация у бактерий хранится в форме проследовательности нуклеотидов в ДНК, которые определяют последовательность аминокислот в
- 14. Гены обозначают строчными начальными буквами названия синтезируемого под их контролем соединения (например, his – гистидиновый ген,
- 15. Кодирование и расшифровка генетической информации у прокариот и эукариот обнаруживают принципиальное сходство. Это доказывает, что механизм
- 16. Внехромосомные факторы наследственности бактерий - плазмиды, транспозоны, IS-последовательности и умеренные бактериофаги, в которых закодированы дополнительные (не
- 17. Плазмиды бактерий Функциональная классификация плазмид основана на свойствах, которыми они наделяют бактерии: 1) F-плазмиды. Кодируют пол
- 18. R-плазмиды R-плазмиды – это плазмиды, детерминирующие множественную лекарственную устойчивость (или резистентность, откуда и название) бактериальной клетки
- 20. Виды изменчивости бактерий К фенотипической изменчивости относятся: Адаптация – приспособление микроорганизмов к новым условиям среды. В
- 21. Генотипическая изменчивость Мутации (от лат. mutatio - изменять) - это передаваемые по наследству структурные изменения генов.
- 22. Мутации могут индуцировать следующие события: модификации оснований (изменения отдельных нуклеотидов), вставки (включение дополнительных оснований), делении (потеря
- 23. Различают несколько видов мутаций: «Молчащие» мутации (мутации «без изменения смысла», то есть не вызывающие изменения аминокислотной
- 24. Генетическая рекомбинация Генетические рекомбинации заключаются в объединении и обычно немедленной перетасовке генов, принадлежащих близкородственным, но генотипически
- 25. Механизмы передачи генетической информации у бактерий У прокариот комбинативные изменения проявляются в результате трансформации, трансдукции, конъюгации.
- 26. Механизмы передачи генетической информации
- 27. А. Конъюгация - требует наличия двух типов клеток: доноров (F+), обладающих F-фактором, и реципиентов (F-), не
- 28. Механизмы передачи генетической информации у бактерий Трансдукция – перенос генов (фрагментов ДНК) от донорской клетки бактерии
- 30. Скачать презентацию

Генетика микроорганизмов
Генетика микроорганизмов

План занятия
1.Характеристика генетического аппарата бактерий
2.Виды внехромосомного генетического материала бактерий
3.Виды изменчивости бактерий
4.Механизмы
План занятия
1.Характеристика генетического аппарата бактерий
2.Виды внехромосомного генетического материала бактерий
3.Виды изменчивости бактерий
4.Механизмы
Характеристика генетического аппарата бактерий
ДНК несёт наследственную функцию в бактериальной клетке. Состоит
Характеристика генетического аппарата бактерий
ДНК несёт наследственную функцию в бактериальной клетке. Состоит
Пуриновые и пиримидиновые основания, входящие в состав молекул нуклеиновых кислот:
К первому
Пуриновые и пиримидиновые основания, входящие в состав молекул нуклеиновых кислот:
К первому

Пиримидины (цитозин, тимин и урацил) содержат шестичленные кольца из двух атомов
Пиримидины (цитозин, тимин и урацил) содержат шестичленные кольца из двух атомов
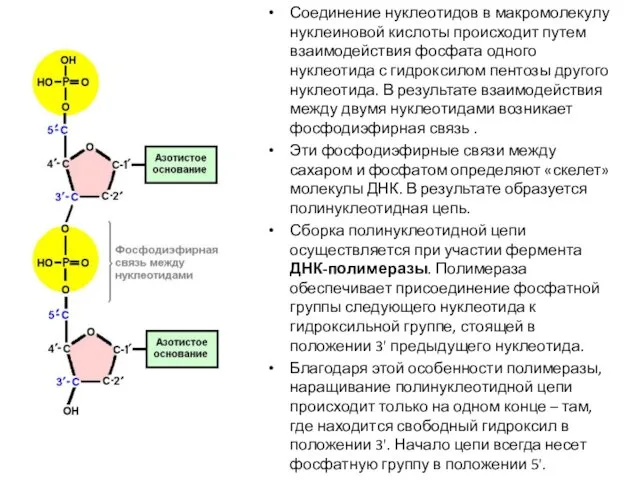
Соединение нуклеотидов в макромолекулу нуклеиновой кислоты происходит путем взаимодействия фосфата одного
Соединение нуклеотидов в макромолекулу нуклеиновой кислоты происходит путем взаимодействия фосфата одного
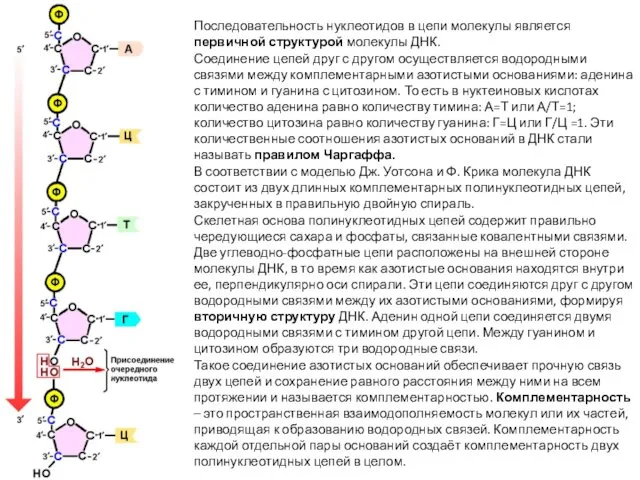
Последовательность нуклеотидов в цепи молекулы является первичной структурой молекулы ДНК.
Соединение цепей
Последовательность нуклеотидов в цепи молекулы является первичной структурой молекулы ДНК.
Соединение цепей
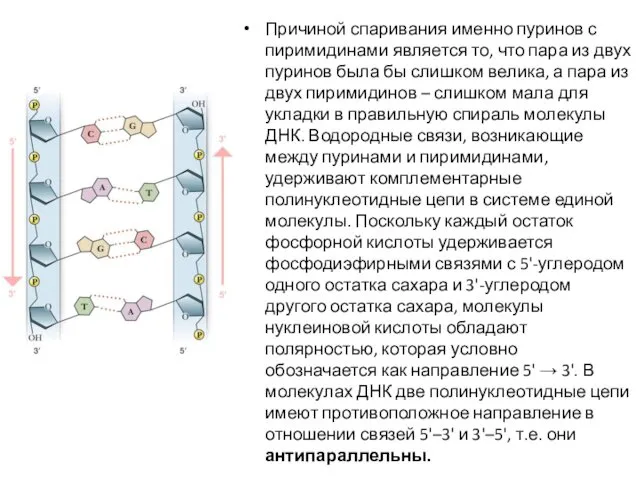
Причиной спаривания именно пуринов с пиримидинами является то, что пара из
Причиной спаривания именно пуринов с пиримидинами является то, что пара из
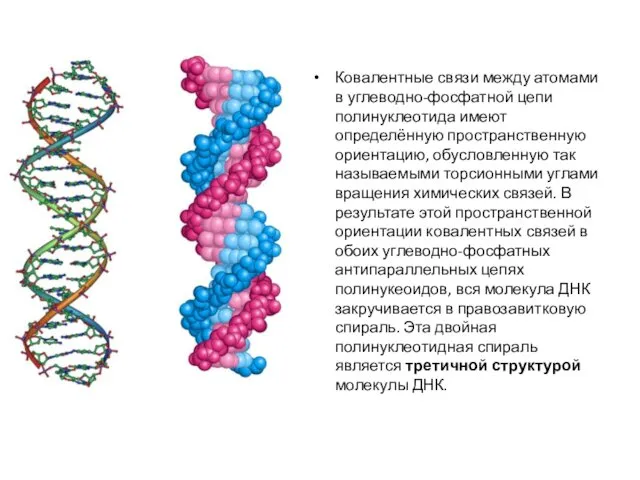
Ковалентные связи между атомами в углеводно-фосфатной цепи полинуклеотида имеют определённую пространственную
Ковалентные связи между атомами в углеводно-фосфатной цепи полинуклеотида имеют определённую пространственную

NB!
Таким образом, в структурной организации молекулы ДНК выделяют три уровня:
–
NB!
Таким образом, в структурной организации молекулы ДНК выделяют три уровня:
–
https://youtu.be/LdKdjhhMAWY - видео, рекомендованное к просмотру
https://youtu.be/LdKdjhhMAWY - видео, рекомендованное к просмотру

Наследственная информация у бактерий хранится в форме проследовательности нуклеотидов в ДНК,
Наследственная информация у бактерий хранится в форме проследовательности нуклеотидов в ДНК,

Гены обозначают строчными начальными буквами названия синтезируемого под их контролем соединения
Гены обозначают строчными начальными буквами названия синтезируемого под их контролем соединения

Кодирование и расшифровка генетической информации у прокариот и эукариот обнаруживают принципиальное
Кодирование и расшифровка генетической информации у прокариот и эукариот обнаруживают принципиальное

Внехромосомные факторы наследственности бактерий - плазмиды, транспозоны, IS-последовательности и умеренные бактериофаги,
Внехромосомные факторы наследственности бактерий - плазмиды, транспозоны, IS-последовательности и умеренные бактериофаги,

Плазмиды бактерий
Функциональная классификация плазмид основана на свойствах, которыми они наделяют бактерии:
Плазмиды бактерий
Функциональная классификация плазмид основана на свойствах, которыми они наделяют бактерии:

R-плазмиды
R-плазмиды – это плазмиды, детерминирующие множественную лекарственную устойчивость (или резистентность,
R-плазмиды
R-плазмиды – это плазмиды, детерминирующие множественную лекарственную устойчивость (или резистентность,


Виды изменчивости бактерий
К фенотипической изменчивости относятся:
Адаптация – приспособление микроорганизмов к новым
Виды изменчивости бактерий
К фенотипической изменчивости относятся:
Адаптация – приспособление микроорганизмов к новым

Генотипическая изменчивость
Мутации (от лат. mutatio - изменять) - это передаваемые по
Генотипическая изменчивость
Мутации (от лат. mutatio - изменять) - это передаваемые по

Мутации могут индуцировать следующие события: модификации оснований (изменения отдельных нуклеотидов), вставки
Мутации могут индуцировать следующие события: модификации оснований (изменения отдельных нуклеотидов), вставки

Различают несколько видов мутаций:
«Молчащие» мутации (мутации «без изменения смысла», то
Различают несколько видов мутаций:
«Молчащие» мутации (мутации «без изменения смысла», то

Генетическая рекомбинация
Генетические рекомбинации заключаются в объединении и обычно немедленной перетасовке генов,
Генетическая рекомбинация
Генетические рекомбинации заключаются в объединении и обычно немедленной перетасовке генов,

Механизмы передачи генетической информации у бактерий
У прокариот комбинативные изменения проявляются в
Механизмы передачи генетической информации у бактерий
У прокариот комбинативные изменения проявляются в
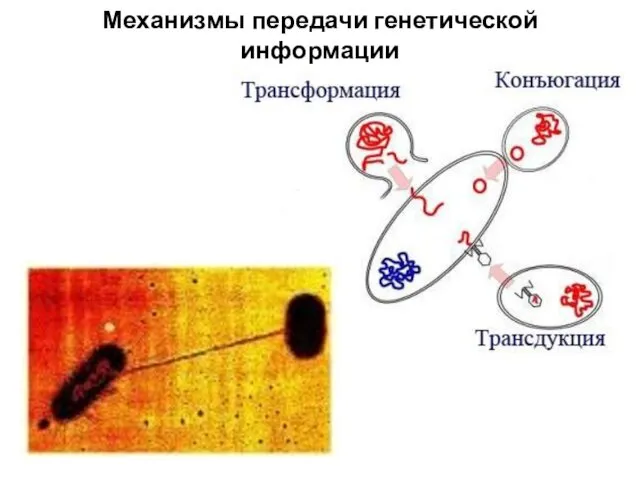
Механизмы передачи генетической информации
Механизмы передачи генетической информации
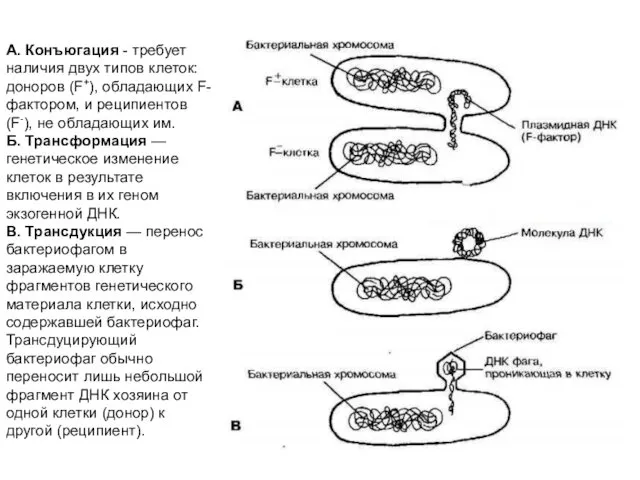
А. Конъюгация - требует наличия двух типов клеток: доноров (F+), обладающих
А. Конъюгация - требует наличия двух типов клеток: доноров (F+), обладающих

Механизмы передачи генетической информации у бактерий
Трансдукция – перенос генов (фрагментов ДНК)
Механизмы передачи генетической информации у бактерий
Трансдукция – перенос генов (фрагментов ДНК)
 Рослинні жири Підготувала учениця 11-А класу Бойко Наталія
Рослинні жири Підготувала учениця 11-А класу Бойко Наталія  Насекомые
Насекомые История и методология космической биологии
История и методология космической биологии Презентация на тему Круговорот воды в природе
Презентация на тему Круговорот воды в природе Экология лесных насекомых. (Лекция 3)
Экология лесных насекомых. (Лекция 3) Отряд жесткокрылые
Отряд жесткокрылые Цветок – генеративный орган, его строение и значение
Цветок – генеративный орган, его строение и значение Презентация на тему "Природа Евразии" - скачать презентации по Биологии
Презентация на тему "Природа Евразии" - скачать презентации по Биологии Триггерные точки верхней части руки
Триггерные точки верхней части руки Огурец Герман F1
Огурец Герман F1 Матричные биосинтезы-2
Матричные биосинтезы-2 Физиология анализаторов. Часть 1
Физиология анализаторов. Часть 1 Стресс, стресс!
Стресс, стресс!  Органойды клетки
Органойды клетки Грибы. Съедобные грибы
Грибы. Съедобные грибы Примитивные организмы
Примитивные организмы Строение и функции белков
Строение и функции белков Деление клетки. Митоз
Деление клетки. Митоз Загадочный витамин В17
Загадочный витамин В17 Кроманьонец
Кроманьонец Презентация на тему "Размножение и оплодотворение у растений" - презентации по Биологии
Презентация на тему "Размножение и оплодотворение у растений" - презентации по Биологии Тритоны. Среда обитания
Тритоны. Среда обитания Наука биология
Наука биология Мы – такие разные. Расы
Мы – такие разные. Расы Инфузория - туфелька
Инфузория - туфелька Каталитическая активность ферментов
Каталитическая активность ферментов Почему листья зеленые?
Почему листья зеленые? В мире животных
В мире животных