- Генетика микроорганизмов. (Лекция 4)

Содержание
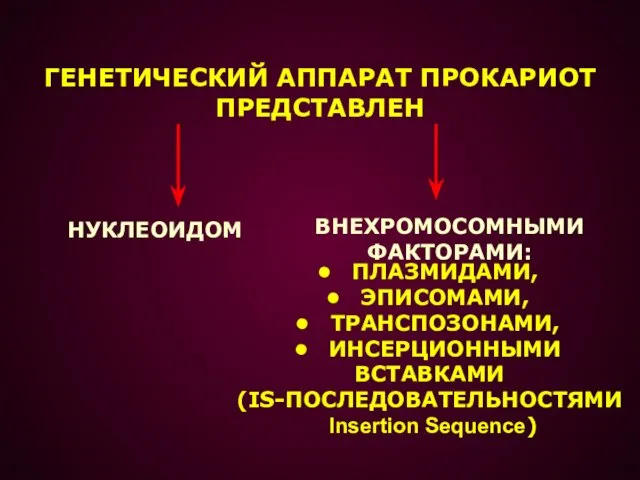
- 2. ГЕНЕТИЧЕСКИЙ АППАРАТ ПРОКАРИОТ ПРЕДСТАВЛЕН НУКЛЕОИДОМ ПЛАЗМИДАМИ, ЭПИСОМАМИ, ТРАНСПОЗОНАМИ, ИНСЕРЦИОННЫМИ ВСТАВКАМИ (IS-ПОСЛЕДОВАТЕЛЬНОСТЯМИ Insertion Sequence) ВНЕХРОМОСОМНЫМИ ФАКТОРАМИ:
- 3. ПЛАЗМИДЫ - ВНЕХРОМОСОМНЫЕ ГЕНЕТИЧЕСКИЕ СТРУКТУРЫ БАКТЕРИЙ. НЕБОЛЬШИЕ МОЛЕКУЛЫ ДНК, СПОСОБНЫЕ К АВТОНОМНОЙ РЕПЛИКАЦИИ. ПЛАЗМИДЫ ЛОКАЛИЗУЮТСЯ В
- 4. СВОБОДНЫЕ ПЛАЗМИДЫ СПОСОБНЫ К АВТОНОМНОЙ ОТ ХРОМОСОМЫ РЕПЛИКАЦИИ ТРАНСМИССИВНЫЕ ПЛАЗМИДЫ НЕТРАНСМИССИВНЫЕ ПЛАЗМИДЫ САМОСТОЯТЕЛЬНО ПЕРЕДАЮТСЯ ДРУГИМ ОСОБЯМ
- 5. ПРИОБРЕТЕНИЕ ИЛИ УТРАТА ПЛАЗМИДЫ ПРИВОДИТ К ПРИОБРЕТЕНИЮ ИЛИ УТРАТЕ ОДНОГО ИЛИ НЕСКОЛЬКИХ ПРИЗНАКОВ, В НЕКОТОРЫХ КЛЕТКАХ
- 6. R-ПЛАЗМИДА (ФАКТОР РЕЗИСТЕНТНОСТИ) - ДЕТЕРМИНИРОВАНИЕ СИНТЕЗА ФЕРМЕНТОВ, РАСЩЕПЛЯЮЩИХ АНТИБИОТИКИ, ТОРМОЖЕНИЕ ПЕРЕНОСА АНТИБИОТИКА ЧЕРЕЗ КЛЕТОЧНЫЕ МЕМБРАНЫ. СОСТОИТ
- 7. COL –ПЛАЗМИДЫ - КОНТРОЛИРУЮТ СИНТЕЗ БАКТЕРИОЦИНОВ, КОТОРЫЕ АКТИВНЫ В ОТНОШЕНИИ БЛИЗКОРОДСТВЕННЫХ ВИДОВ БАКТЕРИЙ. ХАРАКТЕРНО АВТОНОМНОЕ СОСТОЯНИЕ,
- 8. F-ПЛАЗМИДА (ФАКТОР ФЕРТИЛЬНОСТИ) – КОНТРОЛИРУЕТ СИНТЕЗ СЕКС-ПИЛИ, КОНЪЮГАЦИЮ И ПЕРЕНОС ГЕНОВ ХРОМОСОМЫ И НЕТРАНСМИССИВНЫХ ПЛАЗМИД ОТ
- 9. ИНСЕРЦИОННЫЕ ПОСЛЕДОВАТЕЛЬНОСТИ (IS) – ЛИНЕЙНЫЕ ФРАГМЕНТЫ ДВУХЦЕПОЧЕЧНОЙ ДНК (ОТ 200 ДО 2000 П. Н.), СОДЕРЖАТ ТОЛЬКО
- 10. ТРАНСПОЗОНЫ – ПОСЛЕДОВАТЕЛЬНОСТИ ДНК (более 2000 п.н.), СОДЕРЖАТ КРОМЕ ГЕНОВ, ОТВЕТСТВЕННЫХ ЗА ТРАНСПОЗИЦИЮ, СТРУКТУРНЫЕ ГЕНЫ, ОПРЕДЕЛЯЮЩИЕ
- 11. ТРАНСПОЗОНЫ - КОНТРОЛИРУЮТ РЕЗИСТЕНТНОСТЬ К АНТИБИОТИКАМ, ИОНАМ ТЯЖЕЛЫХ МЕТАЛЛОВ, СПОСОБНОСТЬ К КАТАБОЛИЗМУ ЛАКТОЗЫ, РАФФИНОЗЫ, ДЕГРАДАЦИИ ТОЛУОЛА,
- 12. ОСНОВНАЯ ФУНКЦИЯ ГЕНЕТИЧЕСКОГО АППАРАТА – КОНТРОЛЬ НАСЛЕДСТВЕННОСТИ И ИЗМЕНЧИВОСТИ
- 13. ИЗМЕНЧИВОСТЬ - СВОЙСТВО ОРГАНИЗМОВ ПРИОБРЕТАТЬ НОВЫЕ ИЛИ УТРАЧИВАТЬ ИСХОДНЫЕ ПРИЗНАКИ. ИЗМЕНЧИВОСТЬ У БОЛЬШИНСТВА М/О ВЫРАЖЕНА В
- 14. В ТО ЖЕ ВРЕМЯ НЕКОТОРЫЕ ВИДЫ БАКТЕРИЙ (НАПРИМЕР, АРХЕБАКТЕРИИ) И ОТДЕЛЬНЫЕ ИХ ПРИЗНАКИ (ФОРМА, РАЗМЕРЫ, СТРУКТУРА
- 15. ГЕНОТИПИЧЕСКАЯ ИЗМЕНЧИВОСТЬ ОБЕСПЕЧИВАЕТСЯ МУТАЦИЯМИ ГЕНЕТИЧЕСКИМИ РЕКОМБИНАЦИЯМИ
- 16. МУТАЦИИ У БАКТЕРИЙ ГЕНОМНЫЕ ХРОМОСОМНЫЕ ГЕННЫЕ ЗАКЛЮЧАЮТСЯ В ИНТЕГРАЦИИ В ХРОМОСОМУ И ПЕРЕМЕЩЕНИИ ПО НЕЙ ТРАНСПОЗОНОВ,
- 17. СПОНТАННЫЕ МУТАЦИИ ОБУСЛОВЛЕНЫ ОШИБКАМИ РЕПЛИКАЦИИ ГЕНОМА В ПРОЦЕССЕ ДЕЛЕНИЯ ОСОБЕЙ И ОШИБКАМИ РЕПАРАЦИИ ПОВРЕЖДЕННОГО ГЕНОМА, А
- 18. ИНДУЦИРОВАННЫЕ МУТАЦИИ ПОЯВЛЯЮТСЯ В РЕЗУЛЬТАТЕ ДЕЙСТВИЯ МУТАГЕНОВ, К КОТОРЫМ ОТНОСЯТСЯ УФ-ИЗЛУЧЕНИЕ, ИОНИЗИРУЮЩЕЕ ИЗЛУЧЕНИЕ, ХИМИЧЕСКИЕ ВЕЩЕСТВА, ВЕЩЕСТВА
- 19. СУДЬБА МУТАНТОВ ОПРЕДЕЛЯЕТСЯ ИХ ЖИЗНЕСПОСОБНОСТЬЮ И ОТБОРОМ. В СЕЛЕКТИВНОЙ СРЕДЕ МУТАНТЫ МОГУТ ПРИОБРЕСТИ ДОМИНИРУЮЩЕЕ ПОЛОЖЕНИЕ В
- 20. ГЕНЕТИЧЕСКИЕ РЕКОМБИНАЦИИИ – ПРОЦЕСС ОБРАЗОВАНИЯ ГЕНОМОВ, СОДЕРЖАЩИХ ГЕНЕТИЧЕСКИЙ МАТЕРИАЛ ОТ ДВУХ РОДИТЕЛЬСКИХ ФОРМ – БАКТЕРИИ-ДОНОРА (D)
- 21. ТРАНСФОРМАЦИЯ – ПРОЦЕСС ПЕРЕНОСА ГЕНЕТИЧЕСКОГО МАТЕРИАЛА, ПРИ КОТОРОМ КЛЕТКА РЕЦИПИЕНТ ПОГЛОЩАЕТ ГЕНЕТИЧЕСКИЙ МАТЕРИАЛ В ФОРМЕ СВОБОДНОЙ
- 22. СПОСОБНОСТЬ ДНК ПРОНИКАТЬ В КЛЕТКУ РЕЦИПИЕНТА ЗАВИСИТ ОТ «СОСТОЯНИЯ»ДНК (ФРАГМЕНТИРОВАННАЯ МОЛЕКУЛА ДНК) И ОТ ФИЗИОЛОГИЧЕСКОГО СОСТОЯНИЯ
- 23. Процесс трансформации включает несколько фаз: адсорбция ДНК-донора на клетке-реципиенте проникновение ДНК внутрь клетки-реципиента соединение ДНК с
- 24. ТРАНСФОРМАЦИЯ D R
- 25. ТРАНСДУКЦИЕЙ НАЗЫВАЕТСЯ ПЕРЕНОС ГЕНЕТИЧЕСКОГО МАТЕРИАЛА ИЗ ОДНОЙ КЛЕТКИ В ДРУГУЮ С ПОМОЩЬЮ БАКТЕРИОФАГОВ Этот способ генетического
- 26. Трансдукция оказывается возможной, если в процессе размножения фага одна из частиц случайно захватывает фрагмент бактериальной хромосомы.
- 27. ТРАНСДУКЦИЯ D R
- 28. КОНЪЮГАЦИЯ – ПРОЦЕСС ПЕРЕДАЧИ ГЕНЕТИЧЕСКОГО МАТЕРИАЛА ОТ ОДНОЙ КЛЕТКИ К ДРУГОЙ ПРИ ИХ НЕПОСРЕДСТВЕННОМ КОНТАКТЕ, ПРИ
- 29. ПРИ КОНЪЮГАЦИИ F+ КЛЕТКА ПРИСОЕДИНЯЕТСЯ К F- КЛЕТКЕ ПРИ ПОМОЩИ F ПИЛИ F ПЛАЗМИДА РЕПЛИЦИРУЕТСЯ ПО
- 30. КОНЪЮГАЦИЯ D R
- 31. КОНЪЮГАЦИЯ У БАКТЕРИЙ
- 32. МЕТОДЫ ИЗУЧЕНИЯ ГЕНОМА БАКТЕРИЙ ОСНОВАНЫ НА ПРИМЕНЕНИИ КОМПЛЕКСА ГЕНЕТИЧЕСКИХ, БИОХИМИЧЕСКИХ МЕТОДОВ, А ТАКЖЕ КУЛЬТУРАЛЬНОГО МЕТОДА
- 33. ПЦР – метод амплификации, т.е. получения большого числа копий нужного гена или его фрагмента в условиях
- 34. ПЦР ШИРОКО ИСПОЛЬЗУЕТСЯ ДЛЯ РАННЕЙ ДИАГНОСТИКИ ВИЧ-ИНФЕКЦИИ, ВИРУСНЫХ ГЕПАТИТОВ, КЛЕЩЕВОГО ЭНЦЕФАЛИТА, ТУБЕРКУЛЕЗА, ВЕНЕРИЧЕСКИХ ЗАБОЛЕВАНИЙ И Т.Д.
- 35. ВЫСОКИЙ ПОКАЗАТЕЛЬ ЧУВСТВИТЕЛЬНОСТИ И СПЕЦИФИЧНОСТИ (ДО 1000 М/О В 1 МЛ); ВОЗМОЖНОСТЬ ОДНОВРЕМЕННОГО ВЫЯВЛЕНИЯ НЕСКОЛЬКИХ МИКРООРГАНИЗМОВ
- 36. РЕАКЦИОННАЯ СМЕСЬ ДЛЯ ПОЛУЧЕНИЯ НУЖНОЙ ДНК СОДЕРЖИТ: ИССЛЕДУЕМАЯ ДНК-МАТРИЦА, СУБСТРАТЫ РЕАКЦИИ-ДЕЗОКСИНУКЛЕОЗИДТРИФОСФАТЫ (DATP, DCTP, DGTP И TTP)
- 37. Цикл ПЦР включает 3 этапа: Денатурация – исходная смесь нагревается до 94°С, при этом нити ДНК
- 40. ЭТИ ЭТАПЫ ПОВТОРЯЮТСЯ МНОГОКРАТНО В ПРИБОРЕ – АМПЛИФИКАТОРЕ (ТЕРМОЦИКЛЕРЕ), ЧТО ПОЗВОЛЯЕТ ПОЛУЧИТЬ ОГРОМНОЕ КОЛИЧЕСТВО КОПИЙ НУЖНОГО
- 41. THERMOCYCLER ДЛЯ ПОСТАНОВКИ ПЦР
- 42. Современный амплификатор Corbett
- 43. Амплифицированный фрагмент выявляют в процессе электрофореза в агарозном геле
- 45. Скачать презентацию
ГЕНЕТИЧЕСКИЙ АППАРАТ ПРОКАРИОТ
ПРЕДСТАВЛЕН
НУКЛЕОИДОМ
ПЛАЗМИДАМИ,
ЭПИСОМАМИ,
ТРАНСПОЗОНАМИ,
ГЕНЕТИЧЕСКИЙ АППАРАТ ПРОКАРИОТ
ПРЕДСТАВЛЕН
НУКЛЕОИДОМ
ПЛАЗМИДАМИ,
ЭПИСОМАМИ,
ТРАНСПОЗОНАМИ,

ПЛАЗМИДЫ - ВНЕХРОМОСОМНЫЕ ГЕНЕТИЧЕСКИЕ
СТРУКТУРЫ БАКТЕРИЙ.
НЕБОЛЬШИЕ МОЛЕКУЛЫ ДНК,
СПОСОБНЫЕ К АВТОНОМНОЙ РЕПЛИКАЦИИ.
ПЛАЗМИДЫ
ПЛАЗМИДЫ - ВНЕХРОМОСОМНЫЕ ГЕНЕТИЧЕСКИЕ
СТРУКТУРЫ БАКТЕРИЙ.
НЕБОЛЬШИЕ МОЛЕКУЛЫ ДНК,
СПОСОБНЫЕ К АВТОНОМНОЙ РЕПЛИКАЦИИ.
ПЛАЗМИДЫ
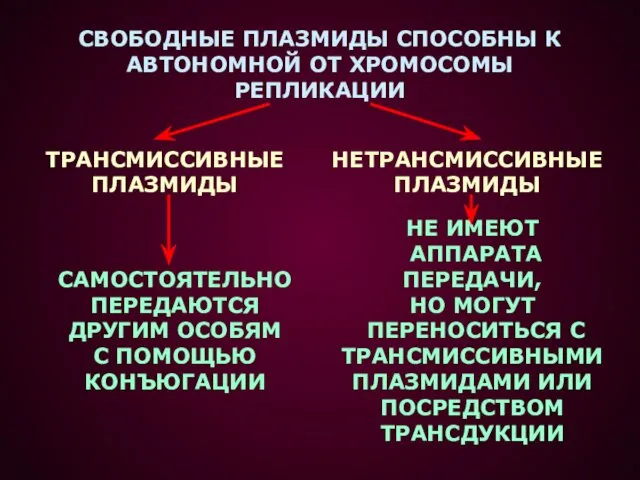
СВОБОДНЫЕ ПЛАЗМИДЫ СПОСОБНЫ К
АВТОНОМНОЙ ОТ ХРОМОСОМЫ
РЕПЛИКАЦИИ
ТРАНСМИССИВНЫЕ
ПЛАЗМИДЫ
НЕТРАНСМИССИВНЫЕ
ПЛАЗМИДЫ
САМОСТОЯТЕЛЬНО
ПЕРЕДАЮТСЯ
ДРУГИМ ОСОБЯМ
СВОБОДНЫЕ ПЛАЗМИДЫ СПОСОБНЫ К
АВТОНОМНОЙ ОТ ХРОМОСОМЫ
РЕПЛИКАЦИИ
ТРАНСМИССИВНЫЕ
ПЛАЗМИДЫ
НЕТРАНСМИССИВНЫЕ
ПЛАЗМИДЫ
САМОСТОЯТЕЛЬНО
ПЕРЕДАЮТСЯ
ДРУГИМ ОСОБЯМ

ПРИОБРЕТЕНИЕ ИЛИ УТРАТА ПЛАЗМИДЫ
ПРИВОДИТ К ПРИОБРЕТЕНИЮ ИЛИ УТРАТЕ
ОДНОГО ИЛИ
ПРИОБРЕТЕНИЕ ИЛИ УТРАТА ПЛАЗМИДЫ
ПРИВОДИТ К ПРИОБРЕТЕНИЮ ИЛИ УТРАТЕ
ОДНОГО ИЛИ

R-ПЛАЗМИДА (ФАКТОР РЕЗИСТЕНТНОСТИ) -
ДЕТЕРМИНИРОВАНИЕ СИНТЕЗА ФЕРМЕНТОВ,
РАСЩЕПЛЯЮЩИХ АНТИБИОТИКИ,
ТОРМОЖЕНИЕ ПЕРЕНОСА
R-ПЛАЗМИДА (ФАКТОР РЕЗИСТЕНТНОСТИ) -
ДЕТЕРМИНИРОВАНИЕ СИНТЕЗА ФЕРМЕНТОВ,
РАСЩЕПЛЯЮЩИХ АНТИБИОТИКИ,
ТОРМОЖЕНИЕ ПЕРЕНОСА

COL –ПЛАЗМИДЫ - КОНТРОЛИРУЮТ СИНТЕЗ
БАКТЕРИОЦИНОВ, КОТОРЫЕ АКТИВНЫ
В ОТНОШЕНИИ БЛИЗКОРОДСТВЕННЫХ
COL –ПЛАЗМИДЫ - КОНТРОЛИРУЮТ СИНТЕЗ
БАКТЕРИОЦИНОВ, КОТОРЫЕ АКТИВНЫ
В ОТНОШЕНИИ БЛИЗКОРОДСТВЕННЫХ

F-ПЛАЗМИДА
(ФАКТОР ФЕРТИЛЬНОСТИ) –
КОНТРОЛИРУЕТ СИНТЕЗ СЕКС-ПИЛИ,
КОНЪЮГАЦИЮ И ПЕРЕНОС ГЕНОВ
F-ПЛАЗМИДА
(ФАКТОР ФЕРТИЛЬНОСТИ) –
КОНТРОЛИРУЕТ СИНТЕЗ СЕКС-ПИЛИ,
КОНЪЮГАЦИЮ И ПЕРЕНОС ГЕНОВ

ИНСЕРЦИОННЫЕ ПОСЛЕДОВАТЕЛЬНОСТИ (IS) –
ЛИНЕЙНЫЕ ФРАГМЕНТЫ
ДВУХЦЕПОЧЕЧНОЙ ДНК (ОТ 200 ДО
ИНСЕРЦИОННЫЕ ПОСЛЕДОВАТЕЛЬНОСТИ (IS) –
ЛИНЕЙНЫЕ ФРАГМЕНТЫ
ДВУХЦЕПОЧЕЧНОЙ ДНК (ОТ 200 ДО

ТРАНСПОЗОНЫ –
ПОСЛЕДОВАТЕЛЬНОСТИ ДНК (более 2000 п.н.),
СОДЕРЖАТ КРОМЕ ГЕНОВ, ОТВЕТСТВЕННЫХ
ТРАНСПОЗОНЫ –
ПОСЛЕДОВАТЕЛЬНОСТИ ДНК (более 2000 п.н.),
СОДЕРЖАТ КРОМЕ ГЕНОВ, ОТВЕТСТВЕННЫХ

ТРАНСПОЗОНЫ -
КОНТРОЛИРУЮТ РЕЗИСТЕНТНОСТЬ
К АНТИБИОТИКАМ, ИОНАМ ТЯЖЕЛЫХ МЕТАЛЛОВ,
СПОСОБНОСТЬ К КАТАБОЛИЗМУ
ЛАКТОЗЫ, РАФФИНОЗЫ,
ТРАНСПОЗОНЫ -
КОНТРОЛИРУЮТ РЕЗИСТЕНТНОСТЬ
К АНТИБИОТИКАМ, ИОНАМ ТЯЖЕЛЫХ МЕТАЛЛОВ,
СПОСОБНОСТЬ К КАТАБОЛИЗМУ
ЛАКТОЗЫ, РАФФИНОЗЫ,

ОСНОВНАЯ ФУНКЦИЯ ГЕНЕТИЧЕСКОГО
АППАРАТА –
КОНТРОЛЬ
НАСЛЕДСТВЕННОСТИ И
ИЗМЕНЧИВОСТИ
ОСНОВНАЯ ФУНКЦИЯ ГЕНЕТИЧЕСКОГО
АППАРАТА –
КОНТРОЛЬ
НАСЛЕДСТВЕННОСТИ И
ИЗМЕНЧИВОСТИ

ИЗМЕНЧИВОСТЬ - СВОЙСТВО ОРГАНИЗМОВ
ПРИОБРЕТАТЬ НОВЫЕ ИЛИ УТРАЧИВАТЬ
ИСХОДНЫЕ ПРИЗНАКИ.
ИЗМЕНЧИВОСТЬ У
ПРИОБРЕТАТЬ НОВЫЕ ИЛИ УТРАЧИВАТЬ
ИСХОДНЫЕ ПРИЗНАКИ.
ИЗМЕНЧИВОСТЬ У

В ТО ЖЕ ВРЕМЯ НЕКОТОРЫЕ ВИДЫ
БАКТЕРИЙ
(НАПРИМЕР, АРХЕБАКТЕРИИ)
И ОТДЕЛЬНЫЕ ИХ
БАКТЕРИЙ
(НАПРИМЕР, АРХЕБАКТЕРИИ)
И ОТДЕЛЬНЫЕ ИХ

ГЕНОТИПИЧЕСКАЯ ИЗМЕНЧИВОСТЬ
ОБЕСПЕЧИВАЕТСЯ
МУТАЦИЯМИ
ГЕНЕТИЧЕСКИМИ
РЕКОМБИНАЦИЯМИ
ГЕНОТИПИЧЕСКАЯ ИЗМЕНЧИВОСТЬ
ОБЕСПЕЧИВАЕТСЯ
МУТАЦИЯМИ
ГЕНЕТИЧЕСКИМИ
РЕКОМБИНАЦИЯМИ
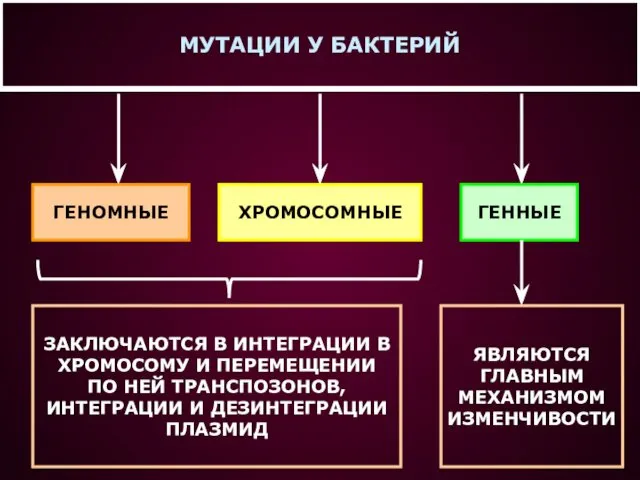
МУТАЦИИ У БАКТЕРИЙ
ГЕНОМНЫЕ
ХРОМОСОМНЫЕ
ГЕННЫЕ
ЗАКЛЮЧАЮТСЯ В ИНТЕГРАЦИИ В
ХРОМОСОМУ И ПЕРЕМЕЩЕНИИ
ПО НЕЙ
МУТАЦИИ У БАКТЕРИЙ
ГЕНОМНЫЕ
ХРОМОСОМНЫЕ
ГЕННЫЕ
ЗАКЛЮЧАЮТСЯ В ИНТЕГРАЦИИ В
ХРОМОСОМУ И ПЕРЕМЕЩЕНИИ
ПО НЕЙ

СПОНТАННЫЕ МУТАЦИИ ОБУСЛОВЛЕНЫ
ОШИБКАМИ РЕПЛИКАЦИИ ГЕНОМА В ПРОЦЕССЕ
ДЕЛЕНИЯ ОСОБЕЙ И
СПОНТАННЫЕ МУТАЦИИ ОБУСЛОВЛЕНЫ
ОШИБКАМИ РЕПЛИКАЦИИ ГЕНОМА В ПРОЦЕССЕ
ДЕЛЕНИЯ ОСОБЕЙ И

ИНДУЦИРОВАННЫЕ МУТАЦИИ ПОЯВЛЯЮТСЯ
В РЕЗУЛЬТАТЕ ДЕЙСТВИЯ МУТАГЕНОВ,
К КОТОРЫМ ОТНОСЯТСЯ
УФ-ИЗЛУЧЕНИЕ,
ИОНИЗИРУЮЩЕЕ
ИНДУЦИРОВАННЫЕ МУТАЦИИ ПОЯВЛЯЮТСЯ
В РЕЗУЛЬТАТЕ ДЕЙСТВИЯ МУТАГЕНОВ,
К КОТОРЫМ ОТНОСЯТСЯ
УФ-ИЗЛУЧЕНИЕ,
ИОНИЗИРУЮЩЕЕ

СУДЬБА МУТАНТОВ ОПРЕДЕЛЯЕТСЯ ИХ
ЖИЗНЕСПОСОБНОСТЬЮ И ОТБОРОМ.
В СЕЛЕКТИВНОЙ СРЕДЕ МУТАНТЫ
СУДЬБА МУТАНТОВ ОПРЕДЕЛЯЕТСЯ ИХ
ЖИЗНЕСПОСОБНОСТЬЮ И ОТБОРОМ.
В СЕЛЕКТИВНОЙ СРЕДЕ МУТАНТЫ

ГЕНЕТИЧЕСКИЕ РЕКОМБИНАЦИИИ –
ПРОЦЕСС ОБРАЗОВАНИЯ ГЕНОМОВ,
СОДЕРЖАЩИХ ГЕНЕТИЧЕСКИЙ
МАТЕРИАЛ ОТ ДВУХ
ГЕНЕТИЧЕСКИЕ РЕКОМБИНАЦИИИ –
ПРОЦЕСС ОБРАЗОВАНИЯ ГЕНОМОВ,
СОДЕРЖАЩИХ ГЕНЕТИЧЕСКИЙ
МАТЕРИАЛ ОТ ДВУХ

ТРАНСФОРМАЦИЯ – ПРОЦЕСС ПЕРЕНОСА ГЕНЕТИЧЕСКОГО МАТЕРИАЛА, ПРИ КОТОРОМ КЛЕТКА РЕЦИПИЕНТ ПОГЛОЩАЕТ
ТРАНСФОРМАЦИЯ – ПРОЦЕСС ПЕРЕНОСА ГЕНЕТИЧЕСКОГО МАТЕРИАЛА, ПРИ КОТОРОМ КЛЕТКА РЕЦИПИЕНТ ПОГЛОЩАЕТ

СПОСОБНОСТЬ ДНК ПРОНИКАТЬ В КЛЕТКУ РЕЦИПИЕНТА ЗАВИСИТ
ОТ «СОСТОЯНИЯ»ДНК (ФРАГМЕНТИРОВАННАЯ МОЛЕКУЛА
СПОСОБНОСТЬ ДНК ПРОНИКАТЬ В КЛЕТКУ РЕЦИПИЕНТА ЗАВИСИТ
ОТ «СОСТОЯНИЯ»ДНК (ФРАГМЕНТИРОВАННАЯ МОЛЕКУЛА

Процесс трансформации включает несколько фаз:
адсорбция ДНК-донора на клетке-реципиенте
проникновение ДНК внутрь клетки-реципиента
соединение
Процесс трансформации включает несколько фаз:
адсорбция ДНК-донора на клетке-реципиенте
проникновение ДНК внутрь клетки-реципиента
соединение
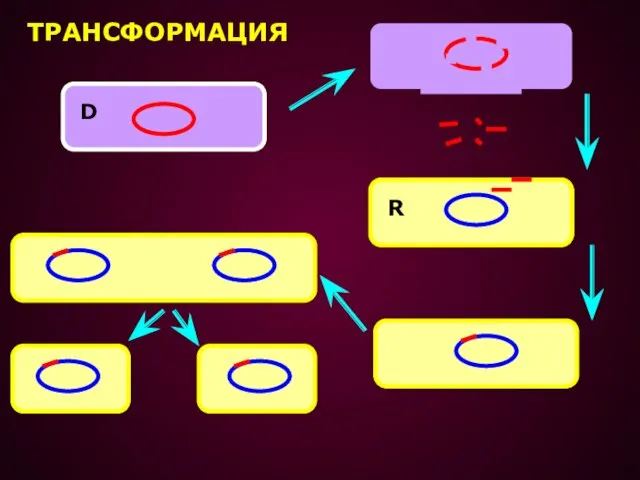
ТРАНСФОРМАЦИЯ
D
R
ТРАНСФОРМАЦИЯ
D
R

ТРАНСДУКЦИЕЙ НАЗЫВАЕТСЯ ПЕРЕНОС ГЕНЕТИЧЕСКОГО МАТЕРИАЛА ИЗ ОДНОЙ КЛЕТКИ В ДРУГУЮ С
ТРАНСДУКЦИЕЙ НАЗЫВАЕТСЯ ПЕРЕНОС ГЕНЕТИЧЕСКОГО МАТЕРИАЛА ИЗ ОДНОЙ КЛЕТКИ В ДРУГУЮ С

Трансдукция оказывается возможной, если в процессе размножения фага одна из частиц
Трансдукция оказывается возможной, если в процессе размножения фага одна из частиц
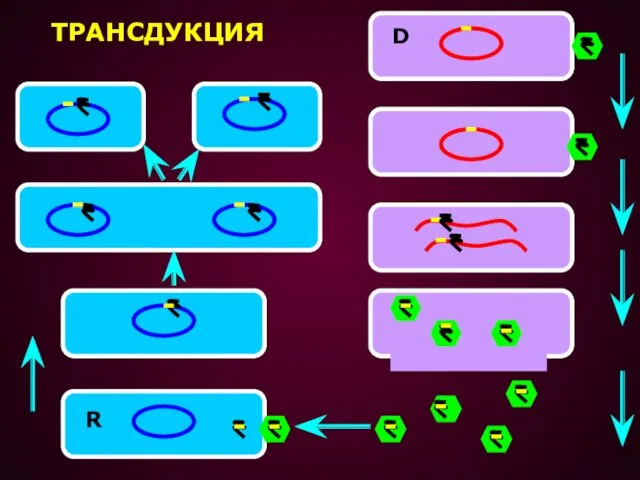
ТРАНСДУКЦИЯ
D
R
ТРАНСДУКЦИЯ
D
R

КОНЪЮГАЦИЯ – ПРОЦЕСС ПЕРЕДАЧИ ГЕНЕТИЧЕСКОГО МАТЕРИАЛА ОТ ОДНОЙ КЛЕТКИ К ДРУГОЙ
КОНЪЮГАЦИЯ – ПРОЦЕСС ПЕРЕДАЧИ ГЕНЕТИЧЕСКОГО МАТЕРИАЛА ОТ ОДНОЙ КЛЕТКИ К ДРУГОЙ

ПРИ КОНЪЮГАЦИИ F+ КЛЕТКА ПРИСОЕДИНЯЕТСЯ К F- КЛЕТКЕ ПРИ ПОМОЩИ
ПРИ КОНЪЮГАЦИИ F+ КЛЕТКА ПРИСОЕДИНЯЕТСЯ К F- КЛЕТКЕ ПРИ ПОМОЩИ
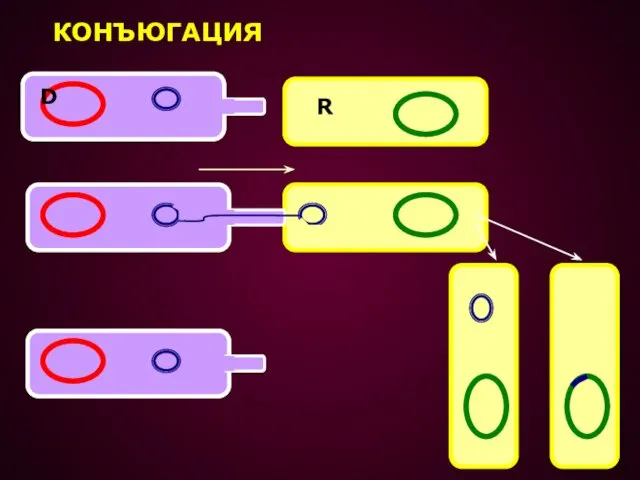
КОНЪЮГАЦИЯ
D
R
КОНЪЮГАЦИЯ
D
R
КОНЪЮГАЦИЯ У БАКТЕРИЙ
КОНЪЮГАЦИЯ У БАКТЕРИЙ

МЕТОДЫ ИЗУЧЕНИЯ ГЕНОМА БАКТЕРИЙ
ОСНОВАНЫ НА ПРИМЕНЕНИИ КОМПЛЕКСА
ГЕНЕТИЧЕСКИХ, БИОХИМИЧЕСКИХ МЕТОДОВ,
А ТАКЖЕ
МЕТОДЫ ИЗУЧЕНИЯ ГЕНОМА БАКТЕРИЙ
ОСНОВАНЫ НА ПРИМЕНЕНИИ КОМПЛЕКСА
ГЕНЕТИЧЕСКИХ, БИОХИМИЧЕСКИХ МЕТОДОВ,
А ТАКЖЕ

ПЦР – метод амплификации,
т.е. получения большого числа копий
нужного гена
ПЦР – метод амплификации,
т.е. получения большого числа копий
нужного гена

ПЦР ШИРОКО ИСПОЛЬЗУЕТСЯ ДЛЯ РАННЕЙ ДИАГНОСТИКИ ВИЧ-ИНФЕКЦИИ, ВИРУСНЫХ ГЕПАТИТОВ, КЛЕЩЕВОГО ЭНЦЕФАЛИТА,
ПЦР ШИРОКО ИСПОЛЬЗУЕТСЯ ДЛЯ РАННЕЙ ДИАГНОСТИКИ ВИЧ-ИНФЕКЦИИ, ВИРУСНЫХ ГЕПАТИТОВ, КЛЕЩЕВОГО ЭНЦЕФАЛИТА,

ВЫСОКИЙ ПОКАЗАТЕЛЬ ЧУВСТВИТЕЛЬНОСТИ И СПЕЦИФИЧНОСТИ (ДО 1000 М/О В 1 МЛ);
ВОЗМОЖНОСТЬ
ВОЗМОЖНОСТЬ

РЕАКЦИОННАЯ СМЕСЬ ДЛЯ ПОЛУЧЕНИЯ НУЖНОЙ ДНК СОДЕРЖИТ:
ИССЛЕДУЕМАЯ ДНК-МАТРИЦА,
СУБСТРАТЫ РЕАКЦИИ-ДЕЗОКСИНУКЛЕОЗИДТРИФОСФАТЫ
РЕАКЦИОННАЯ СМЕСЬ ДЛЯ ПОЛУЧЕНИЯ НУЖНОЙ ДНК СОДЕРЖИТ:
ИССЛЕДУЕМАЯ ДНК-МАТРИЦА,
СУБСТРАТЫ РЕАКЦИИ-ДЕЗОКСИНУКЛЕОЗИДТРИФОСФАТЫ

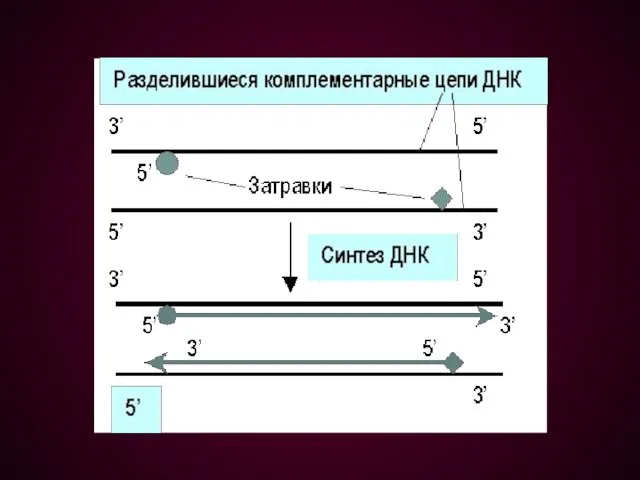
Цикл ПЦР включает 3 этапа:
Денатурация – исходная смесь нагревается до
Цикл ПЦР включает 3 этапа:
Денатурация – исходная смесь нагревается до


ЭТИ ЭТАПЫ ПОВТОРЯЮТСЯ МНОГОКРАТНО В ПРИБОРЕ – АМПЛИФИКАТОРЕ (ТЕРМОЦИКЛЕРЕ), ЧТО ПОЗВОЛЯЕТ
ЭТИ ЭТАПЫ ПОВТОРЯЮТСЯ МНОГОКРАТНО В ПРИБОРЕ – АМПЛИФИКАТОРЕ (ТЕРМОЦИКЛЕРЕ), ЧТО ПОЗВОЛЯЕТ

THERMOCYCLER ДЛЯ ПОСТАНОВКИ ПЦР
THERMOCYCLER ДЛЯ ПОСТАНОВКИ ПЦР

Современный амплификатор Corbett
Современный амплификатор Corbett

Амплифицированный фрагмент выявляют в процессе электрофореза в агарозном геле
Амплифицированный фрагмент выявляют в процессе электрофореза в агарозном геле
 Способы размножения животных. Оплодотворение
Способы размножения животных. Оплодотворение Торможение условных рефлексов Торможение условных рефлексов
Торможение условных рефлексов Торможение условных рефлексов  1 закон Менделя
1 закон Менделя Презентация на тему Корень Строение и функции
Презентация на тему Корень Строение и функции  Что есть что в мире библейских растений. Кедр - символ величия, славы и богатства
Что есть что в мире библейских растений. Кедр - символ величия, славы и богатства Технологічні карти
Технологічні карти Презентация на тему "Хвостатые" - скачать презентации по Биологии
Презентация на тему "Хвостатые" - скачать презентации по Биологии Деление покрытосеменных растений на классы и семейства
Деление покрытосеменных растений на классы и семейства Реализация генетической информации у про- и эукариот
Реализация генетической информации у про- и эукариот Предмет и задачи микробиологии и иммунологии
Предмет и задачи микробиологии и иммунологии Вода - источник жизни
Вода - источник жизни Репродукция вирусов
Репродукция вирусов Эмбриональное развитие человека
Эмбриональное развитие человека Презентация на тему "Охрана природы" - скачать презентации по Биологии
Презентация на тему "Охрана природы" - скачать презентации по Биологии Чарльз Дарвин и его теория возникновения жизни
Чарльз Дарвин и его теория возникновения жизни Побег и почки. Внешнее строение листа Урок биологии в 6 классе Учитель: Мироненко Л.Ю.
Побег и почки. Внешнее строение листа Урок биологии в 6 классе Учитель: Мироненко Л.Ю. Углеводы (сахариды)
Углеводы (сахариды) Цветок, его строение и значение
Цветок, его строение и значение Методы выявления и учета вредителей капусты и лука
Методы выявления и учета вредителей капусты и лука Терминация репликации. Теломеры. Их репликация и роль в функционировании эукариотической клетки
Терминация репликации. Теломеры. Их репликация и роль в функционировании эукариотической клетки Общие вопросы функциональной анатомии сенсорных систем
Общие вопросы функциональной анатомии сенсорных систем Дыхательная система
Дыхательная система Томаты
Томаты Красная книга
Красная книга «Болезни глаз. Гигиена зрения». Учебный проект по биологии ученицы 8 класса Железногорской санаторно-лесной школы Гербаль Люд
«Болезни глаз. Гигиена зрения». Учебный проект по биологии ученицы 8 класса Железногорской санаторно-лесной школы Гербаль Люд Эра динозавров в цифрах
Эра динозавров в цифрах Клеточный цикл и его регуляция
Клеточный цикл и его регуляция Анатомо-морфологические особенности и основные физиологические функции организма
Анатомо-морфологические особенности и основные физиологические функции организма