- Реализация генетической информации у про- и эукариот

Содержание
- 2. План лекции. Ген, определение. Строение генов про- и эукариот. Генетический код и его свойства. Реализация генетической
- 3. Термин ген был предложен датчанином Йогансеном в 1909 году. Природа гена была не ясна, но описывались
- 4. Итак, свойства гена: (не путаем со свойствами генетического кода!) Дискретность - имеет определенный размер и позицию
- 5. Н.К.Кольцов и идея матричного синтеза Бывшее здание Института экспериментальной биологии Н.К. Кольцова в Москве на ул.
- 6. Путь идеи: Кольцов Тимофеев-Ресовский Дельбрюк Уотсон Никола́й Влади́мирович Тимофе́ев-Ресо́вский (1900 – 1981) — биолог, генетик. Основные
- 7. Но к концу 40-х годов XX века стало ясно, что гены – это ДНК Дж. Уотсон
- 8. Центральная догма молекулярной биологии:
- 9. В генах закодированы не только белки ДНК Ген иРНК Белок Функция нкРНК Функция
- 10. Что же такое ген сегодня? Определение. Ген – участок молекулы ДНК, несущий информацию о первичной структуре
- 11. генетика эпигенетика транскриптомика геномика протеомика А сама генетика за ХХ век разрослась и дала ветви
- 12. Рассмотрим этапы реализации наследственной информации на примере синтеза белка.
- 13. Живые организмы делятся на два больших надцарства: Прокариоты (доядерные) Эукариоты (ядерные) растения грибы животные археи синезеленые
- 14. Размеры геномов разных видов (в гаплоидном наборе). 4.6 Mb – размер генома кишечной палочки (бактерия) 670
- 15. Основные отличия организации генов про- и эукариот: Прокариоты До 90% ДНК это гены, кодирующие белки Гены
- 16. Чем сложнее организм, тем больше у него в геноме не кодирующей белки ДНК Величина белок-кодирующей части
- 17. Строение лактозного оперона бактерии кишечной палочки (E.coli). Ген I промотор оператор Ген Z Ген Y Ген
- 18. У прокариот транскрипция (1) и трансляция(2) не разделены ни в пространстве, ни во времени прокариотическая клетка
- 19. Особенности экспрессии генов у прокариот дают нам ряд преимуществ Многие антибиотики (аминогликозиды, тетрациклины, хлорамфеникол) связываются с
- 20. Рассмотрим этапы синтеза белка у эукариот
- 21. Типичный ген эукариот имеет свой промотор и несколько регуляторов регуляторы промотор лидер трейлер кодирующая область -
- 22. Этапы реализации наследственной информации (синтеза белка) Транскрипция - синтез РНК (всех видов) по матрице ДНК. (Происходит
- 23. 1. Транскрипция – синтез РНК по матрице ДНК Транскрипция включает: Инициацию Элонгацию Терминацию Факторов, участвующих в
- 24. Факторы транскрипции Специфические факторы транскрипции (например, гормоны) инициируют или подавляют транскрипцию определенных генов, связываясь с участками
- 25. ДНК-связывающий домен типа «лейциновая молния» в комплексе с ДНК. ДНК-связывающий домен типа «спираль-петля-спираль» в комплексе с
- 26. Специфические факторы влияют на уровень транскрипции
- 27. Транскрипция: инициация ДНК кодогенная(смысловая) цепь матричная цепь 3’ 5’ 5’ 3’ Нуклеотиды РНК Фермент РНК-полимераза Есть
- 28. Транскрипция: элонгация ДНК кодогенная(смысловая) цепь матричная цепь 3’ 5’ 5’ 3’ РНК-полимераза движется вдоль матричной цепи
- 29. Транскрипция: терминация ДНК 3’ 5’ 5’ 3’ РНК отделяется от ДНК 5’ 3’ РНК Сигналом терминации
- 30. Сейчас сезон грибов… (Медицинские аспекты регуляции транскрипции) Гриб бледная поганка (Amanita), содержит яд α-аманитин. LD50 (доза
- 31. 2. Процессинг мРНК 5’ 3’ Кэпирование Присоединение полиаденилового «хвоста» Вырезание интронов Сплайсинг экзонов Модификация нуклеотидов Незрелая
- 32. Сплайсинг – вырезание интронов и сшивание экзонов – осуществляет сплайсосома Сплайсосома в собранном виде Сплайсосому образуют
- 33. Сплайсосома распознает интроны по определенным последовательностям Молекула пре-мРНК обязательно содержит специфические последовательности, распознаваемые во время сборки
- 34. Сплайсинг может регулироваться. 94% генов у человека подвержены альтернативному сплайсингу, а у остальных 6% просто нет
- 35. 3.Трансляция Трансляция – синтез полипептида из аминокислот в рибосоме по матрице иРНК. Трансляция происходит в соответствии
- 36. Свойства генетического кода (не путать со свойствами гена!): Триплетность – одна аминокислота шифруется тремя нуклеотидами. Специфичность
- 37. Есть разные представления таблицы генетического кода
- 38. Амиокислоты записывают по трем первым буквам названия или одной буквой
- 39. В трансляции участвуют: иРНК тРНК Рибосомы ( в состав рибосомы входит рРНК и белки) Факторы трансляции
- 40. Процессинг тРНК
- 41. Нетипичные азотистые основания в тРНК Инозин способен образовывать водородные связи с разными азотистыми основаниями
- 42. тРНК (транспортная РНК) переносит аминокислоты к рибосоме Cодержит от 76 до 90 нуклеотидов. Число видов тРНК
- 43. Wobble hypothesis – теория неоднозначного соответствия, гипотеза качания, предложена Ф.Криком. Нестрогое соответствие оснований в 3-ей позиции
- 44. Почему третье основание не столь важно? Специфичность кодон-антикодонового взаимодействия обеспечивается главным образом двумя первыми основаниями кодонов;
- 45. Аминокислота присоединяется к 3’ концу «своей» тРНК аминокислота + тРНК + АТФ → аминоацил-тРНК + АМФ
- 46. Рибосома - немембранный органоид клетки, состоящий из белков и рРНК. иРНК Две молекулы тРНК Малая субъединица
- 47. 1.Инициация. Трансляция начинается с того, что иРНК соединяется с малой субъединицей рибосомы АУГ УАЦ Большая субъединица
- 48. 2. Элонгация. У рибосомы формируется функциональный центр А У Г Ц Г Г Г Ц Ц
- 49. 2. Элонгация. Между аминокислотами возникает пептидная связь СО-NH А У Г Ц Г Г Г Ц
- 50. 2. Элонгация. После образования пептидной связи тРНК уходит, а рибосома сдвигается на 1 триплет, что называется
- 51. 2. Элонгация. Подходят новые тРНК, образуются пептидные связи, рибосома движется вдоль иРНК… А У Г Ц
- 52. 3. Терминация. Когда в А-участке оказывается один из трех возможных стоп-триплетов трансляция заканчивается А У Г
- 53. Рибосома вновь распадается на субъединицы В синтезе длинного полипептида может участвовать несколько рибосом. Они образуют полисому
- 54. 4. Процессинг белка. В ходе трансляции образуется полипептид (цепь аминокислотных остатков) - это первичная структура белка.
- 55. Примеры малых пептидов: окситоцин соединения, обладающие гормональной активностью (глюкагон, окситоцин, вазопрессин и др.); вещества, регулирующие пищеварительные
- 56. Примеры белков:самый большой известный белок – титин, также известный как тайтин или коннектин — самый большой
- 57. Процессинг индивидуален у каждого белка Фолдинг – преобретение белком его трехмерной структуры.
- 58. К основным реакциям процессинга белков относятся: 1. Удаление с N-конца метионина или даже нескольких аминокислот. 2.
- 59. Примеры белков: цинковые пальцы Цинковый палец (англ. zinc finger) — тип белковой структуры, небольшой белковый мотив,
- 60. Примеры белков: лейциновая молния Лейциновая застёжка-молния (также лейциновая застёжка, лейциновая молния, англ. leucine zipper) — тип
- 61. Фолдинг белков Фолдинг – это процесс укладки вытянутой полипептидной цепи в правильную трехмерную структуру. Для обеспечения
- 62. Путь секретируемого белка лежит через каналы ЭПС к аппарату Гольджи
- 63. Пример: процессинг инсулина 1) Препроинсулин, 110 аминокислотных остатков (L — лидерный пептид, B — участок, C
- 65. Неправильный фолдинг и прионные болезни
- 66. Регуляция экспрессии генов у эукариот
- 67. История вопроса
- 68. XIX век. Август Вейсман и теория детерминант половая клетка дифференцировка и смерть клетки тела непрерывный ряд
- 69. Это справедливо в части случаях: у некоторых червей и членистоногих Вейсман опирался на данные о том,
- 70. точно известно общее число клеток тела(959) и что из чего разовьется. Например, у небольшого круглого червя
- 71. Иначе обстоят дела у большинства других животных, в том числе человека. Зигота бластомеры личинки Морской еж
- 72. Развитие стали представлять как процесс включения и выключения генов (а не их разрушения, как полагал Вейсман)
- 73. В ХХ века была сформулирована гипотеза дифференциальной активности генов Ядро каждой клетки содержит полный набор генов
- 74. Опыт Джона Гёрдона по клонирование африканской шпорцевой лягушки. Гердон проводил опыты в 1960-х годах В 1990-х
- 75. Политенные хромосомы личинок двукрылых Dolly the sheep А, Б – личинка в возрасте 110 час. В
- 76. Гены можно разделить на конститутивные, т.е. всегда активные. Их иногда называют «генами домашнего хозяйства» регулируемые, т.е.
- 77. Регуляция активности гена Позитивная (индукция), включение Негативная (репрессия), выключение
- 78. Регуляция экспрессии генов у прокариот См. Предыдущую лекцию
- 79. Регуляция экспрессии генов у эукариот
- 80. Регуляция экспрессии генов у эукариот происходит на всех этапах синтеза белка. Изменение число копий генов Регуляция
- 81. 1. Число копий гена
- 82. 1. Изменение числа копий гена Х Увеличение – амплификация. Например, у амфибий синтезируются дополнительные копии генов
- 83. В выключении Х-хромосомы у самок млекопитающих участвует нкРНК Канадский ученый Барр (1908 – 1995) (и его
- 84. Инактивация Х-хромосомы у самок млекопитающих В части клеток активна Х от отца, в части от матери
- 85. Xist РНК окружает ту Х хромосому, с которой экспрессируется, и превращает ее в гетерохроматин – тельце
- 86. 2. Транскрипция Самый частый уровень регуляции экспрессии гена
- 87. 2. Регуляция транскрипции Много белков принимают в этом участие Энхансеры – усилители – включают и усиливают
- 88. Многие белки благодаря своей конфигурации могут связываться с ДНК (являются регуляторами транскрипции) Спираль-петля-спираль (helix-loop-helix) Лейциновая молния
- 89. На активность транскрипции влияют гистоны – белки хромосом Ацетилирование гистонов ослабляет связь ДНК с ними и
- 90. Метилирование цитозина в ДНК, наоборот, подавляет транскрипцию Нет метилирования – активный ген Есть метилирование – ген
- 91. У многоклеточных эукариот в роли регуляторов транскрипции выступают гормоны клетка ядро ДНК гормон рецептор транскрипция Каскад
- 92. Стероидные гормоны в комплексе с рецептором связываются прямо с ДНК
- 93. 3. Процессинг РНК
- 94. Регуляция процессинга РНК Альтернативный сплайсинг Редактирование РНК Изменение РНК путем вставок, делеций или изменения азотистых оснований.
- 95. Альтернативный сплайсинг приводит к появлению изоформ белков
- 96. Показано, что у человека 94 % генов подвержено альтернативному сплайсингу (у остальных 6 % генов нет
- 97. Пример редактирования РНК (эдитинг) Ген аполипопротеина ДНК Пре-мРНК кишечник печень без редактирования редатирование: ЦАА ? УАА
- 98. 4. Трансляция
- 99. Регуляция трансляции Удлинение или укорочение времени жизни иРНК: Пример: гормон пролактин удлиняет время жизни иРНК для
- 100. 5. Процессинг белка
- 101. 5. Регуляция процессинга белка Коллаген – основной белок соединительной ткани, около 30% всего белка тела. Есть
- 102. Синтез коллагена сложен и включает много посттрансляционных преобразований Нарушение сборки коллагеновых фибрилл наблюдается при болезнях соединительной
- 104. Несса Кери пишет:
- 106. Скачать презентацию

План лекции.
Ген, определение. Строение генов про- и эукариот.
Генетический код и его
План лекции.
Ген, определение. Строение генов про- и эукариот.
Генетический код и его

Термин ген был предложен датчанином Йогансеном в 1909 году.
Природа гена была
Термин ген был предложен датчанином Йогансеном в 1909 году. Природа гена была

Итак, свойства гена:
(не путаем со свойствами генетического кода!)
Дискретность - имеет определенный
Итак, свойства гена:
(не путаем со свойствами генетического кода!)
Дискретность - имеет определенный

Н.К.Кольцов и идея матричного синтеза
Бывшее здание Института экспериментальной биологии Н.К. Кольцова
Н.К.Кольцов и идея матричного синтеза
Бывшее здание Института экспериментальной биологии Н.К. Кольцова

Путь идеи: Кольцов Тимофеев-Ресовский Дельбрюк Уотсон
Никола́й Влади́мирович Тимофе́ев-Ресо́вский
(1900 – 1981)
Путь идеи: Кольцов Тимофеев-Ресовский Дельбрюк Уотсон
Никола́й Влади́мирович Тимофе́ев-Ресо́вский
(1900 – 1981)

Но к концу 40-х годов XX века стало ясно, что гены
Но к концу 40-х годов XX века стало ясно, что гены
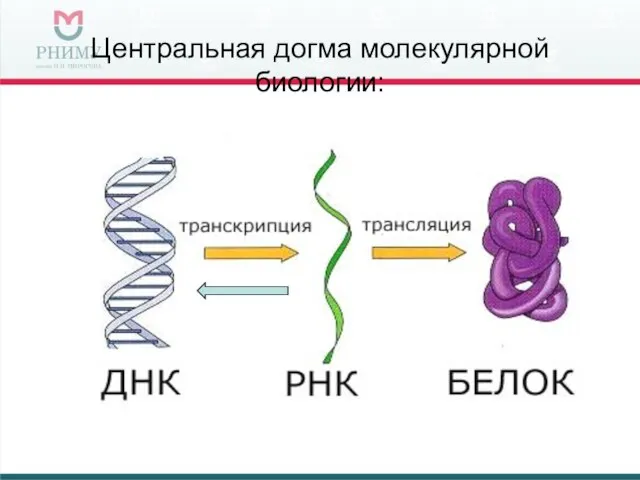
Центральная догма молекулярной биологии:
Центральная догма молекулярной биологии:
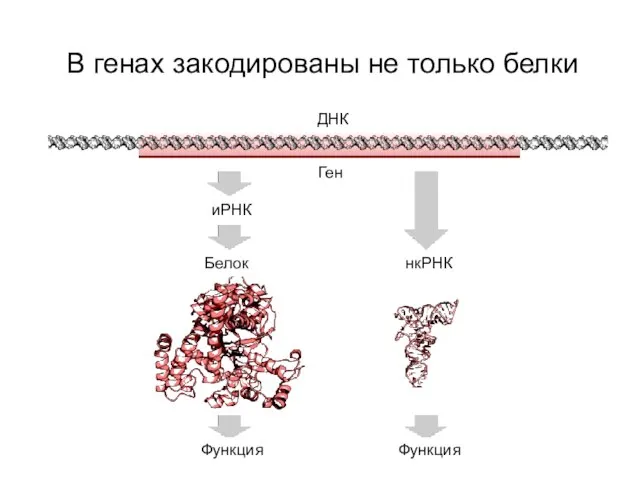
В генах закодированы не только белки
ДНК
Ген
иРНК
Белок
Функция
нкРНК
Функция
В генах закодированы не только белки
ДНК
Ген
иРНК
Белок
Функция
нкРНК
Функция
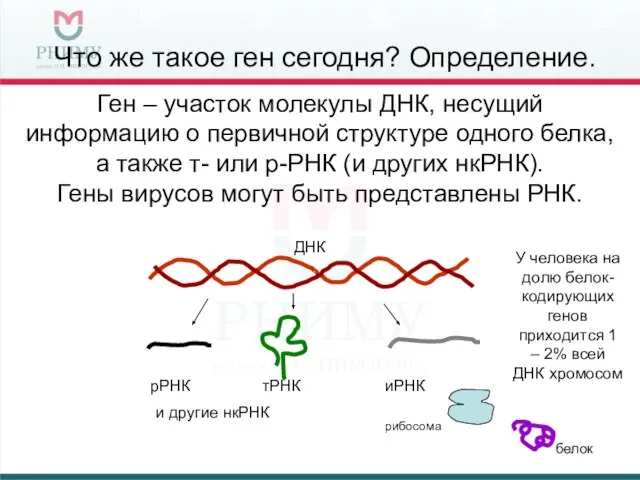
Что же такое ген сегодня? Определение.
Ген – участок молекулы ДНК, несущий
Что же такое ген сегодня? Определение.
Ген – участок молекулы ДНК, несущий
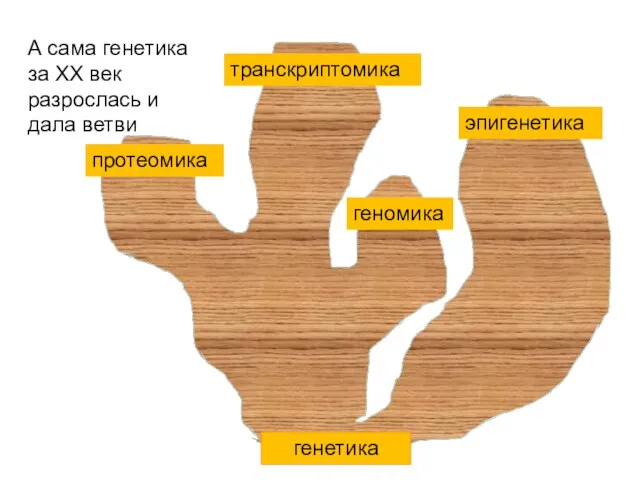
генетика
эпигенетика
транскриптомика
геномика
протеомика
А сама генетика за ХХ век разрослась и дала ветви
генетика
эпигенетика
транскриптомика
геномика
протеомика
А сама генетика за ХХ век разрослась и дала ветви

Рассмотрим этапы реализации наследственной информации на примере синтеза белка.
Рассмотрим этапы реализации наследственной информации на примере синтеза белка.
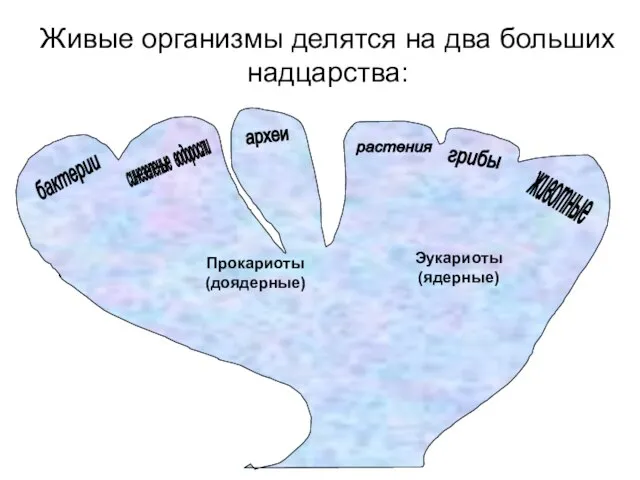
Живые организмы делятся на два больших надцарства:
Прокариоты (доядерные)
Эукариоты (ядерные)
растения
грибы
животные
Живые организмы делятся на два больших надцарства:
Прокариоты (доядерные)
Эукариоты (ядерные)
растения
грибы
животные

Размеры геномов разных видов
(в гаплоидном наборе).
4.6 Mb – размер генома
Размеры геномов разных видов
(в гаплоидном наборе).
4.6 Mb – размер генома

Основные отличия организации генов про- и эукариот:
Прокариоты
До 90% ДНК это
Основные отличия организации генов про- и эукариот:
Прокариоты
До 90% ДНК это
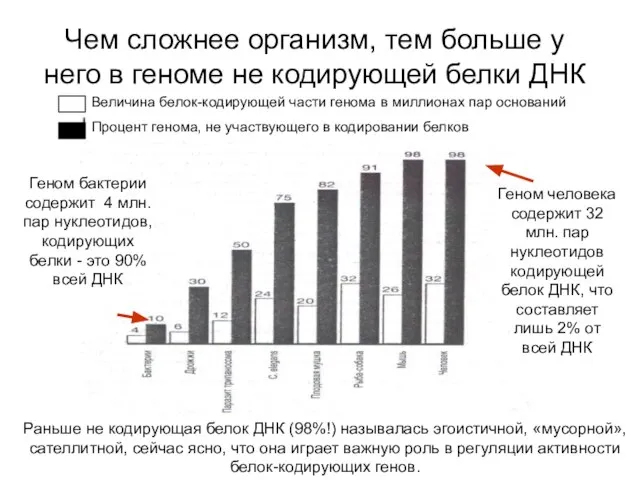
Чем сложнее организм, тем больше у него в геноме не кодирующей
Чем сложнее организм, тем больше у него в геноме не кодирующей
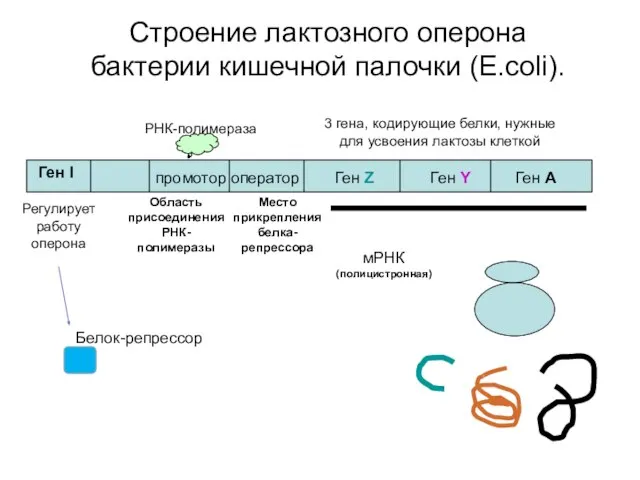
Строение лактозного оперона бактерии кишечной палочки (E.coli).
Ген I
промотор оператор
Ген Z
Ген
Строение лактозного оперона бактерии кишечной палочки (E.coli).
Ген I
промотор оператор
Ген Z
Ген
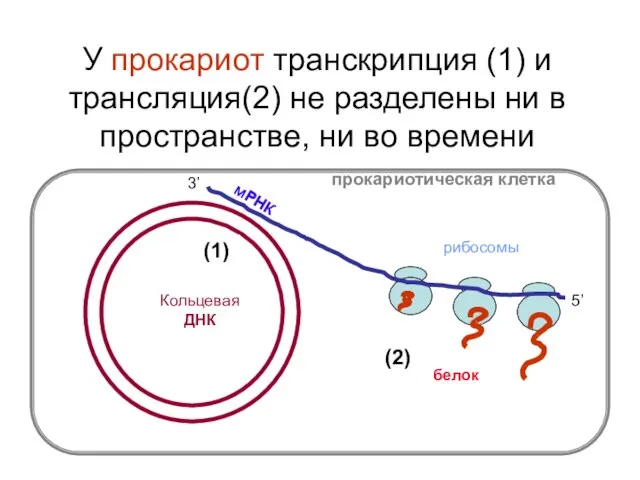
У прокариот транскрипция (1) и трансляция(2) не разделены ни в пространстве,
У прокариот транскрипция (1) и трансляция(2) не разделены ни в пространстве,

Особенности экспрессии генов у прокариот дают нам ряд преимуществ
Многие антибиотики (аминогликозиды,
Особенности экспрессии генов у прокариот дают нам ряд преимуществ
Многие антибиотики (аминогликозиды,

Рассмотрим этапы синтеза белка у эукариот
Рассмотрим этапы синтеза белка у эукариот
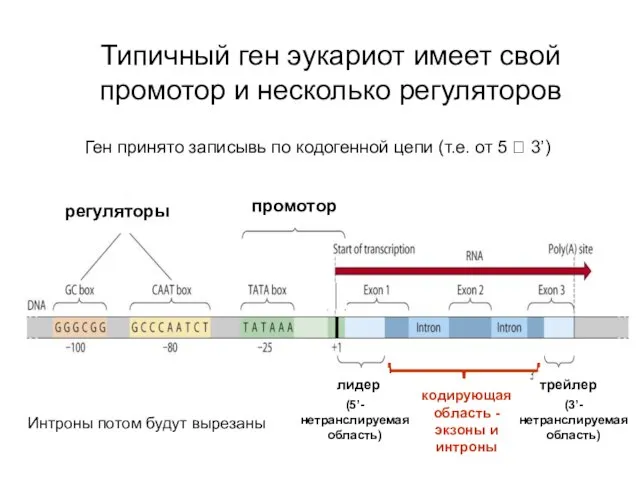
Типичный ген эукариот имеет свой промотор и несколько регуляторов
регуляторы
промотор
лидер
трейлер
кодирующая область -
Типичный ген эукариот имеет свой промотор и несколько регуляторов
регуляторы
промотор
лидер
трейлер
кодирующая область -

Этапы реализации наследственной информации (синтеза белка)
Транскрипция - синтез РНК (всех видов)
Этапы реализации наследственной информации (синтеза белка)
Транскрипция - синтез РНК (всех видов)
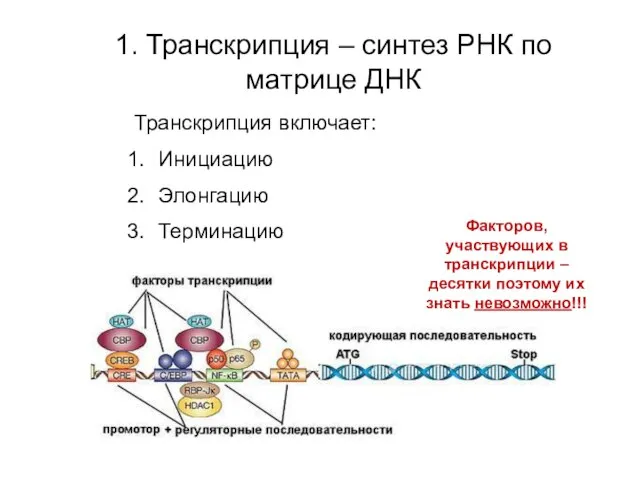
1. Транскрипция – синтез РНК по матрице ДНК
Транскрипция включает:
Инициацию
Элонгацию
Терминацию
Факторов, участвующих
1. Транскрипция – синтез РНК по матрице ДНК
Транскрипция включает:
Инициацию
Элонгацию
Терминацию
Факторов, участвующих
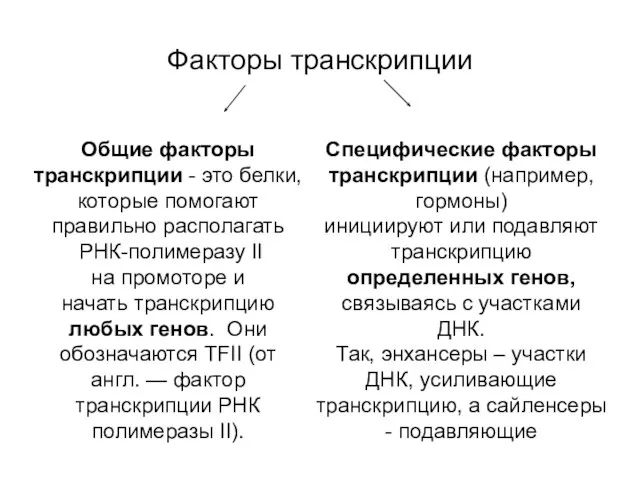
Факторы транскрипции
Специфические факторы транскрипции (например, гормоны)
инициируют или подавляют транскрипцию определенных
Факторы транскрипции
Специфические факторы транскрипции (например, гормоны)
инициируют или подавляют транскрипцию определенных
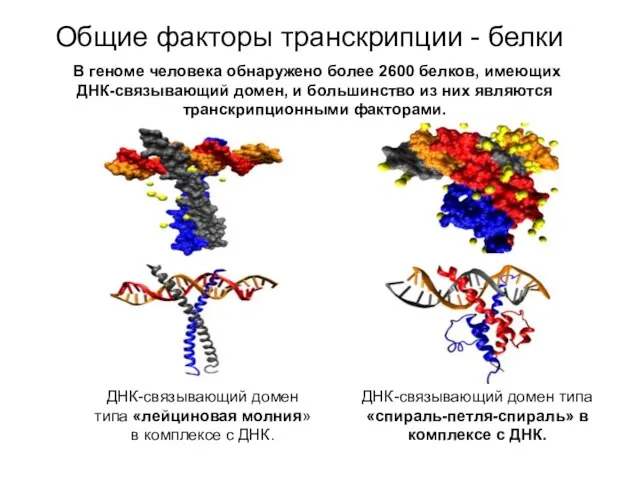
ДНК-связывающий домен типа «лейциновая молния» в комплексе с ДНК.
ДНК-связывающий домен
ДНК-связывающий домен типа «лейциновая молния» в комплексе с ДНК.
ДНК-связывающий домен
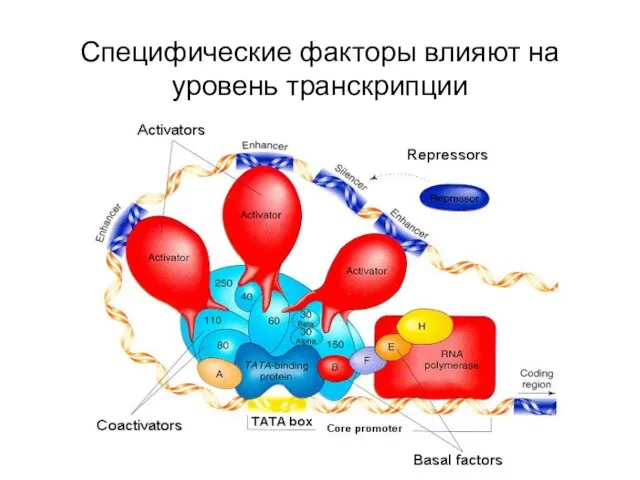
Специфические факторы влияют на уровень транскрипции
Специфические факторы влияют на уровень транскрипции
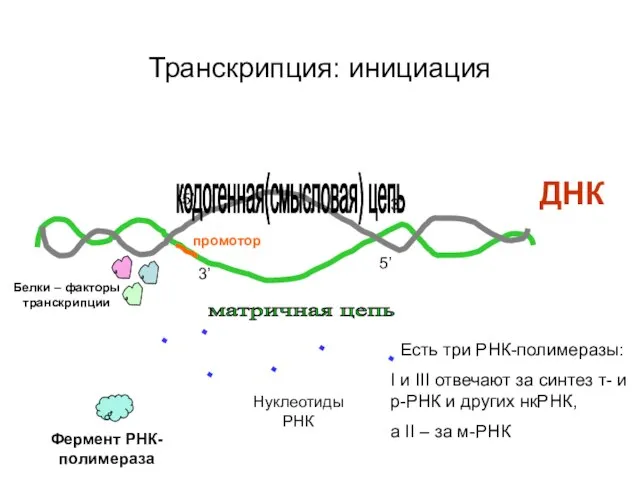
Транскрипция: инициация
ДНК
кодогенная(смысловая) цепь
матричная цепь
3’
5’
5’
3’
Нуклеотиды РНК
Фермент РНК-полимераза
Есть три РНК-полимеразы:
I и
Транскрипция: инициация
ДНК
кодогенная(смысловая) цепь
матричная цепь
3’
5’
5’
3’
Нуклеотиды РНК
Фермент РНК-полимераза
Есть три РНК-полимеразы:
I и

Транскрипция: элонгация
ДНК
кодогенная(смысловая) цепь
матричная цепь
3’
5’
5’
3’
РНК-полимераза движется вдоль матричной цепи ДНК
Транскрипция: элонгация
ДНК
кодогенная(смысловая) цепь
матричная цепь
3’
5’
5’
3’
РНК-полимераза движется вдоль матричной цепи ДНК
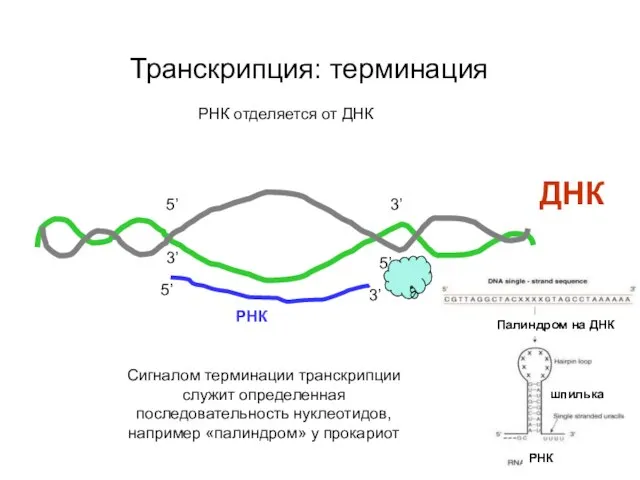
Транскрипция: терминация
ДНК
3’
5’
5’
3’
РНК отделяется от ДНК
5’
3’
РНК
Сигналом терминации транскрипции служит определенная последовательность нуклеотидов,
Транскрипция: терминация
ДНК
3’
5’
5’
3’
РНК отделяется от ДНК
5’
3’
РНК
Сигналом терминации транскрипции служит определенная последовательность нуклеотидов,

Сейчас сезон грибов… (Медицинские аспекты регуляции транскрипции)
Гриб бледная поганка (Amanita), содержит
Сейчас сезон грибов… (Медицинские аспекты регуляции транскрипции)
Гриб бледная поганка (Amanita), содержит

2. Процессинг мРНК
5’
3’
Кэпирование
Присоединение полиаденилового «хвоста»
Вырезание интронов
Сплайсинг экзонов
Модификация нуклеотидов
Незрелая мРНК
Кэп (7метил-гуанозинтирфосфат)
Хвост из
2. Процессинг мРНК
5’
3’
Кэпирование
Присоединение полиаденилового «хвоста»
Вырезание интронов
Сплайсинг экзонов
Модификация нуклеотидов
Незрелая мРНК
Кэп (7метил-гуанозинтирфосфат)
Хвост из
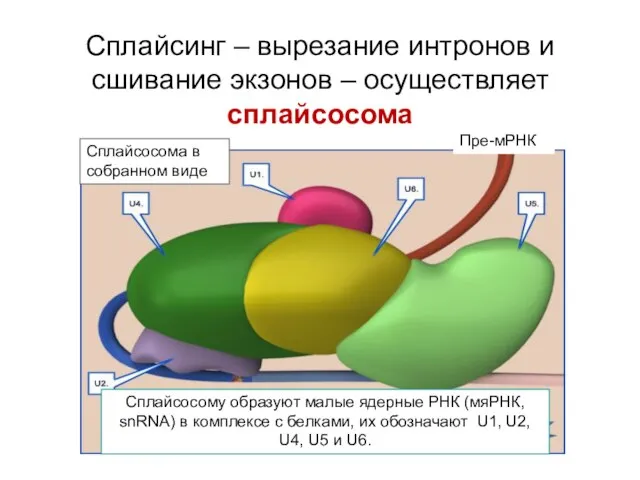
Сплайсинг – вырезание интронов и сшивание экзонов – осуществляет сплайсосома
Сплайсосома в
Сплайсинг – вырезание интронов и сшивание экзонов – осуществляет сплайсосома
Сплайсосома в
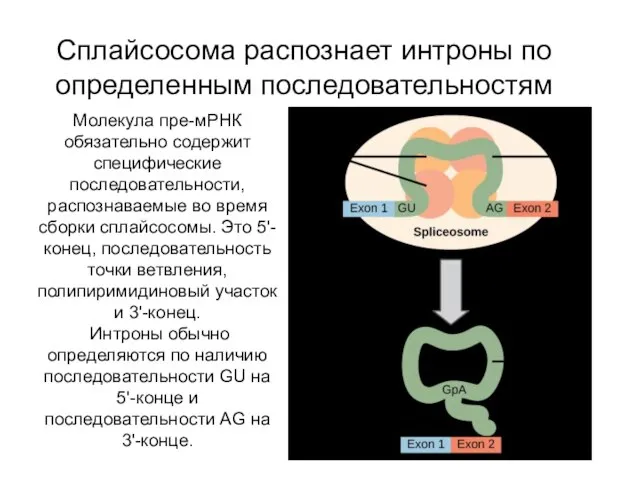
Сплайсосома распознает интроны по определенным последовательностям
Молекула пре-мРНК обязательно содержит специфические последовательности,
Сплайсосома распознает интроны по определенным последовательностям
Молекула пре-мРНК обязательно содержит специфические последовательности,
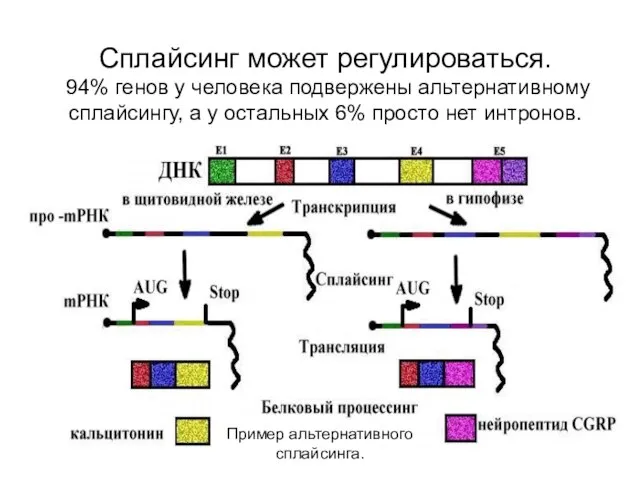
Сплайсинг может регулироваться.
94% генов у человека подвержены альтернативному сплайсингу, а
Сплайсинг может регулироваться. 94% генов у человека подвержены альтернативному сплайсингу, а
3.Трансляция
Трансляция – синтез полипептида из аминокислот в рибосоме по матрице
3.Трансляция
Трансляция – синтез полипептида из аминокислот в рибосоме по матрице

Свойства генетического кода (не путать со свойствами гена!):
Триплетность – одна аминокислота
Свойства генетического кода (не путать со свойствами гена!):
Триплетность – одна аминокислота
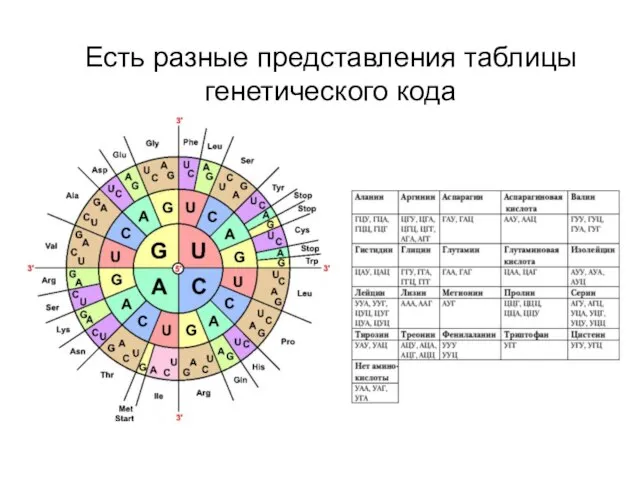
Есть разные представления таблицы генетического кода
Есть разные представления таблицы генетического кода

Амиокислоты записывают по трем первым буквам названия или одной буквой
Амиокислоты записывают по трем первым буквам названия или одной буквой
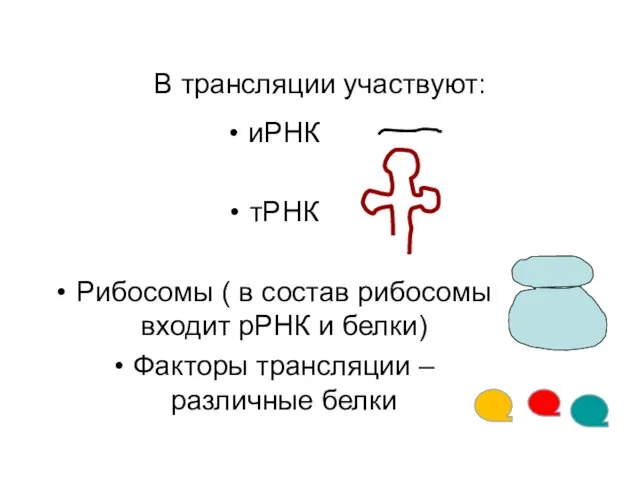
В трансляции участвуют:
иРНК
тРНК
Рибосомы ( в состав рибосомы входит рРНК и белки)
Факторы
В трансляции участвуют:
иРНК
тРНК
Рибосомы ( в состав рибосомы входит рРНК и белки)
Факторы
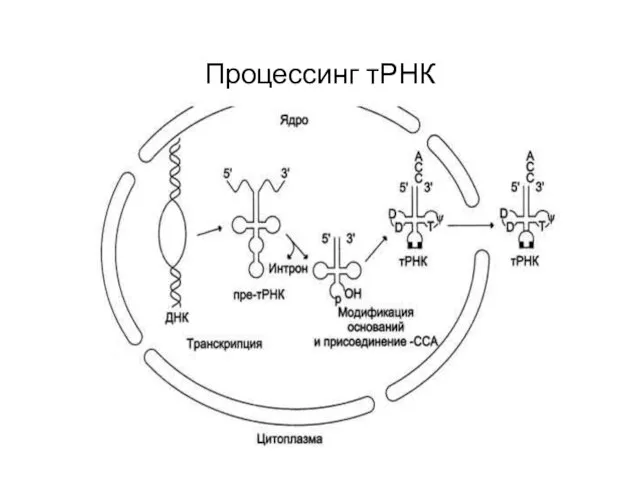
Процессинг тРНК
Процессинг тРНК
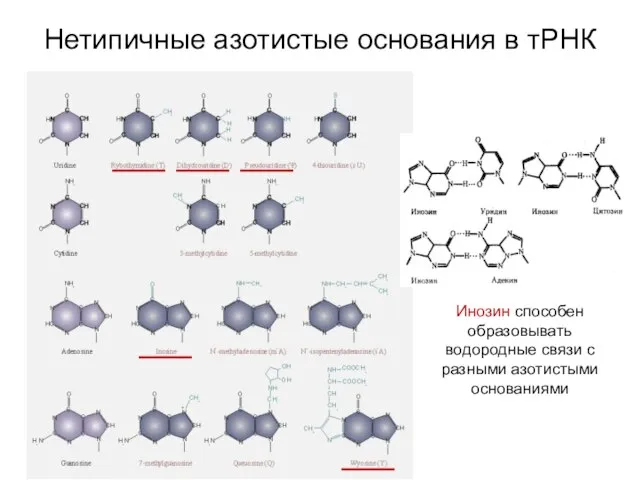
Нетипичные азотистые основания в тРНК
Инозин способен образовывать водородные связи с разными
Нетипичные азотистые основания в тРНК
Инозин способен образовывать водородные связи с разными
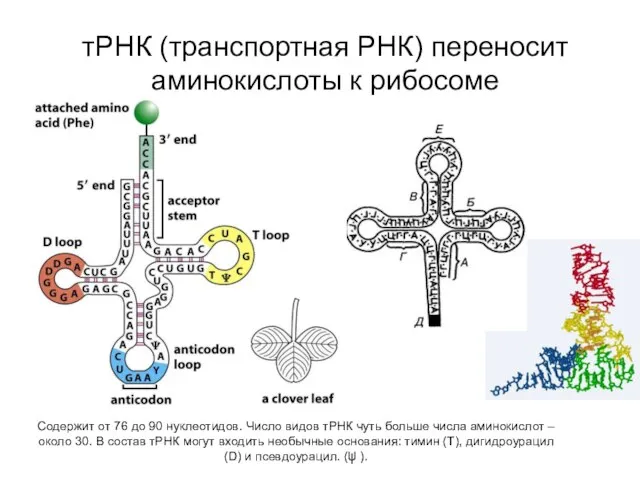
тРНК (транспортная РНК) переносит аминокислоты к рибосоме
Cодержит от 76 до 90
тРНК (транспортная РНК) переносит аминокислоты к рибосоме
Cодержит от 76 до 90
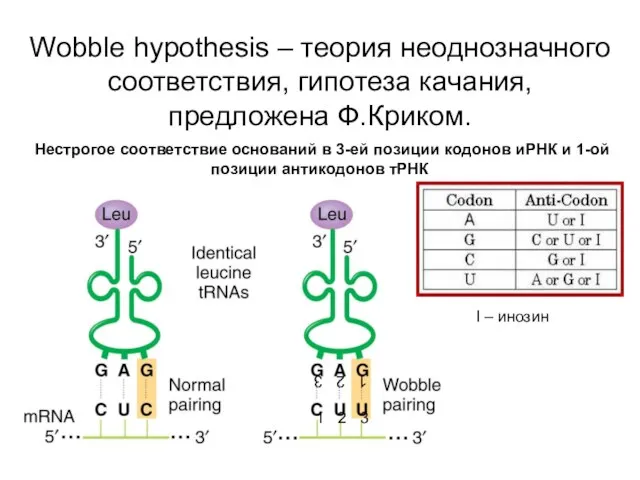
Wobble hypothesis – теория неоднозначного соответствия, гипотеза качания, предложена Ф.Криком.
Нестрогое
Wobble hypothesis – теория неоднозначного соответствия, гипотеза качания, предложена Ф.Криком.
Нестрогое

Почему третье основание не столь важно?
Специфичность кодон-антикодонового взаимодействия обеспечивается главным образом
Почему третье основание не столь важно?
Специфичность кодон-антикодонового взаимодействия обеспечивается главным образом
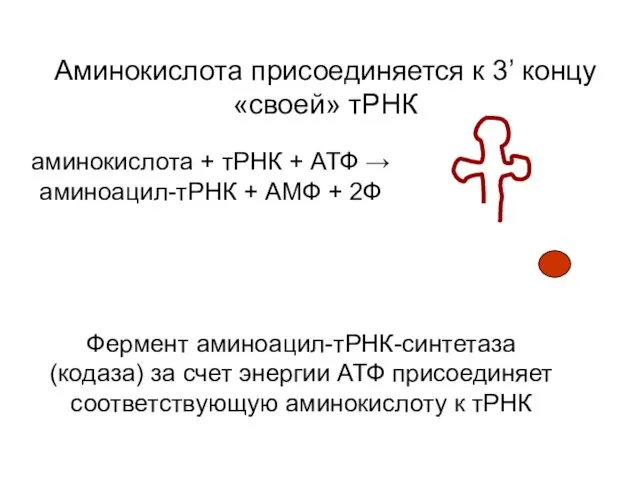
Аминокислота присоединяется к 3’ концу «своей» тРНК
аминокислота + тРНК +
Аминокислота присоединяется к 3’ концу «своей» тРНК
аминокислота + тРНК +
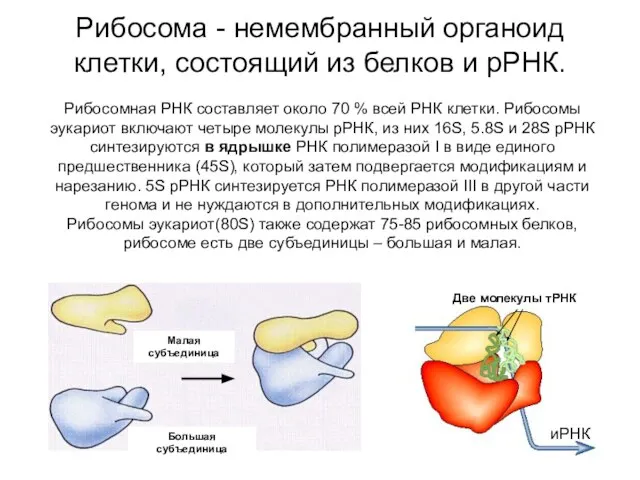
Рибосома - немембранный органоид клетки, состоящий из белков и рРНК.
иРНК
Две молекулы
Рибосома - немембранный органоид клетки, состоящий из белков и рРНК.
иРНК
Две молекулы
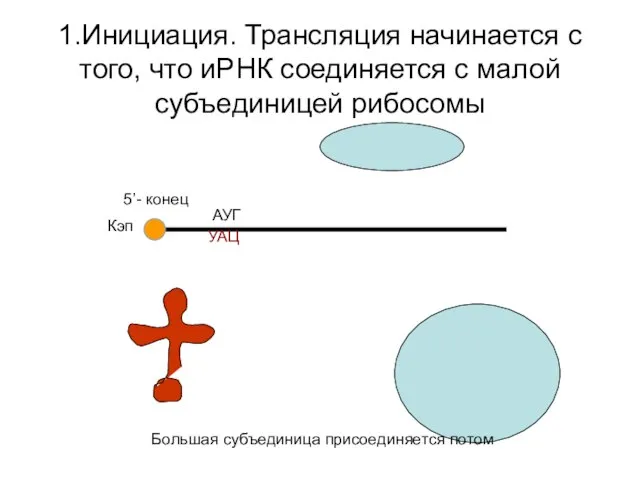
1.Инициация. Трансляция начинается с того, что иРНК соединяется с малой субъединицей
1.Инициация. Трансляция начинается с того, что иРНК соединяется с малой субъединицей
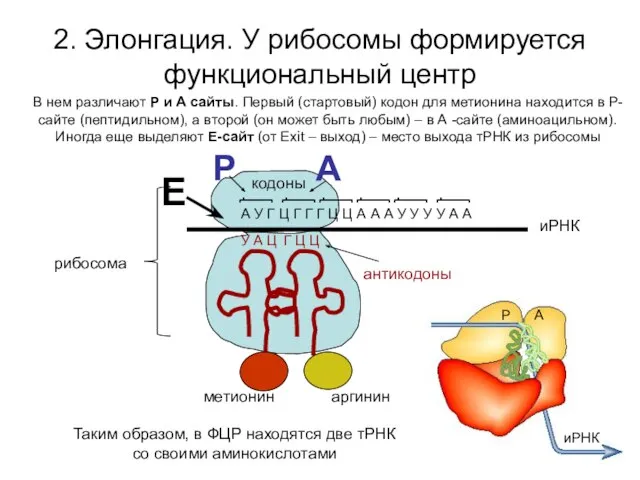
2. Элонгация. У рибосомы формируется функциональный центр
А У Г Ц Г
2. Элонгация. У рибосомы формируется функциональный центр
А У Г Ц Г
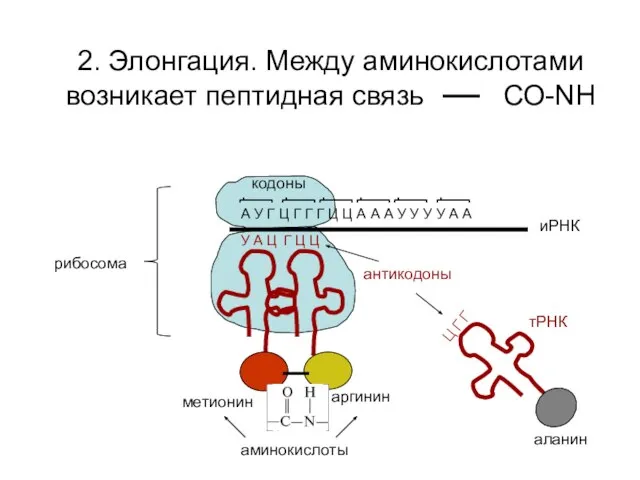
2. Элонгация. Между аминокислотами возникает пептидная связь СО-NH
А У Г Ц
2. Элонгация. Между аминокислотами возникает пептидная связь СО-NH
А У Г Ц
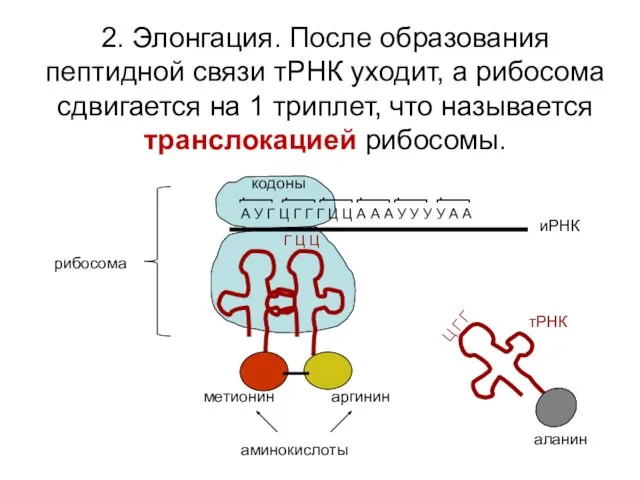
2. Элонгация. После образования пептидной связи тРНК уходит, а рибосома сдвигается
2. Элонгация. После образования пептидной связи тРНК уходит, а рибосома сдвигается
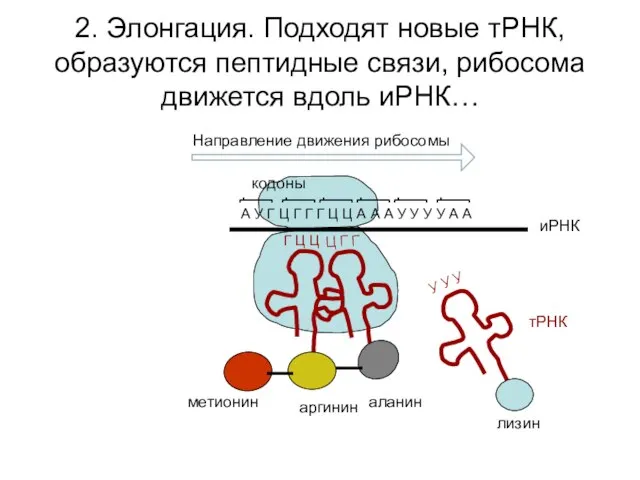
2. Элонгация. Подходят новые тРНК, образуются пептидные связи, рибосома движется вдоль
2. Элонгация. Подходят новые тРНК, образуются пептидные связи, рибосома движется вдоль
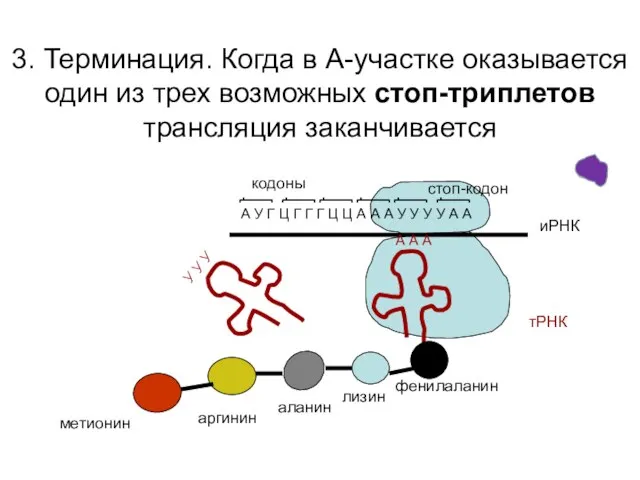
3. Терминация. Когда в А-участке оказывается один из трех возможных стоп-триплетов
3. Терминация. Когда в А-участке оказывается один из трех возможных стоп-триплетов
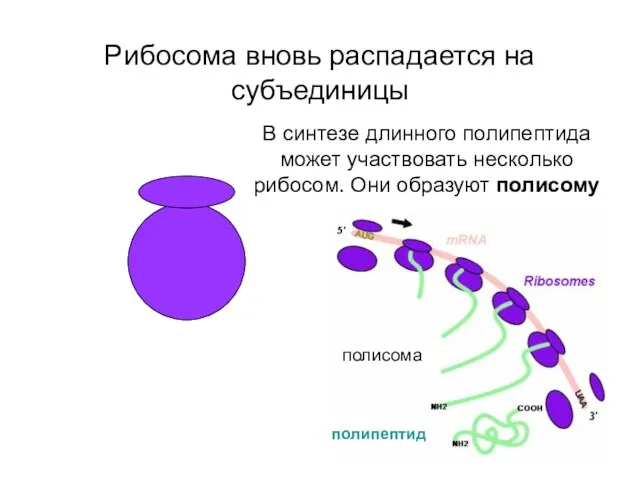
Рибосома вновь распадается на субъединицы
В синтезе длинного полипептида может участвовать несколько
Рибосома вновь распадается на субъединицы
В синтезе длинного полипептида может участвовать несколько
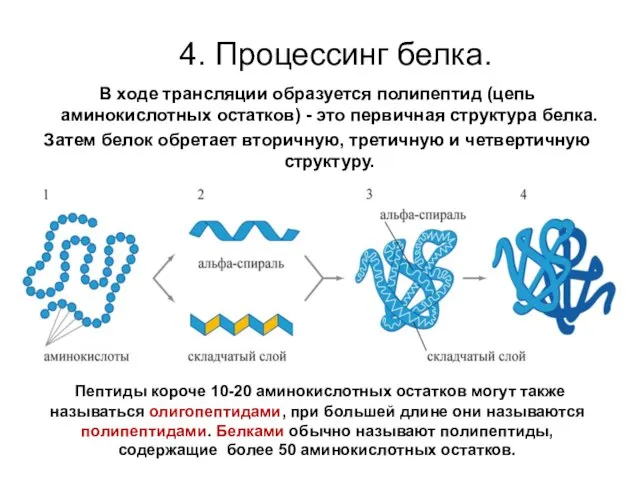
4. Процессинг белка.
В ходе трансляции образуется полипептид (цепь аминокислотных остатков)
4. Процессинг белка.
В ходе трансляции образуется полипептид (цепь аминокислотных остатков)
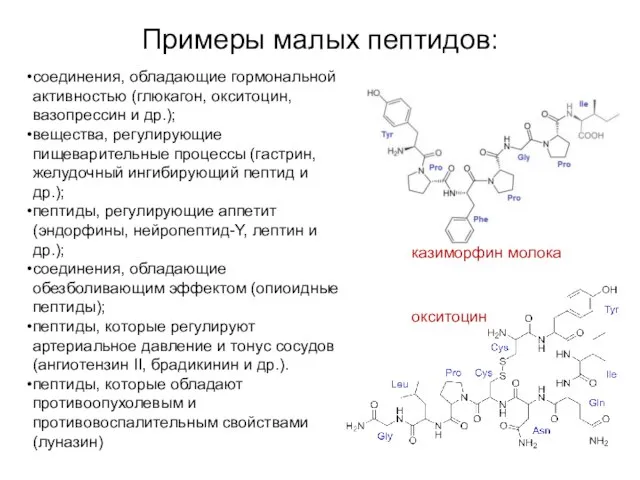
Примеры малых пептидов:
окситоцин
соединения, обладающие гормональной активностью (глюкагон, окситоцин, вазопрессин и др.);
вещества,
Примеры малых пептидов:
окситоцин
соединения, обладающие гормональной активностью (глюкагон, окситоцин, вазопрессин и др.);
вещества,

Примеры белков:самый большой известный белок – титин,
также известный как тайтин или
Примеры белков:самый большой известный белок – титин,
также известный как тайтин или
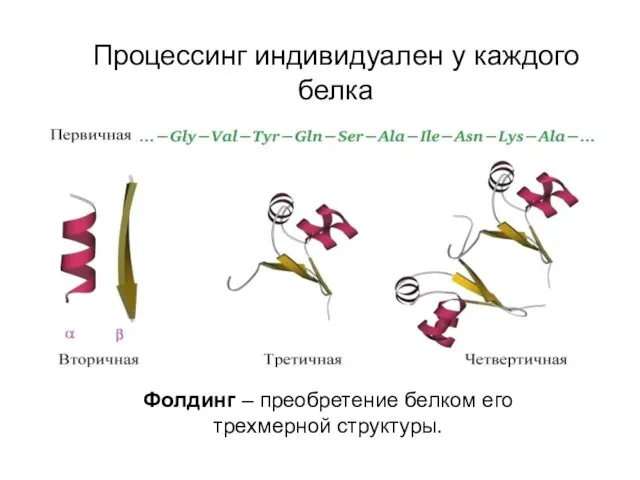
Процессинг индивидуален у каждого белка
Фолдинг – преобретение белком его трехмерной структуры.
Процессинг индивидуален у каждого белка
Фолдинг – преобретение белком его трехмерной структуры.

К основным реакциям процессинга белков относятся:
1. Удаление с N-конца метионина или
К основным реакциям процессинга белков относятся:
1. Удаление с N-конца метионина или

Примеры белков: цинковые пальцы
Цинковый палец (англ. zinc finger) — тип белковой структуры,
Примеры белков: цинковые пальцы
Цинковый палец (англ. zinc finger) — тип белковой структуры,
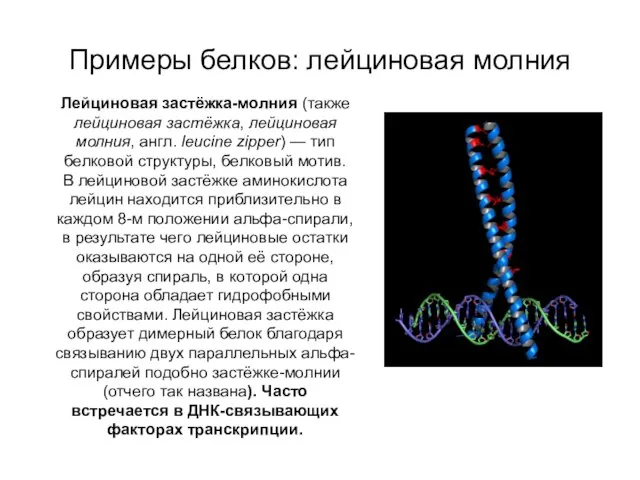
Примеры белков: лейциновая молния
Лейциновая застёжка-молния (также лейциновая застёжка, лейциновая молния, англ. leucine
Примеры белков: лейциновая молния
Лейциновая застёжка-молния (также лейциновая застёжка, лейциновая молния, англ. leucine

Фолдинг белков
Фолдинг – это процесс укладки вытянутой полипептидной цепи в правильную
Фолдинг белков
Фолдинг – это процесс укладки вытянутой полипептидной цепи в правильную
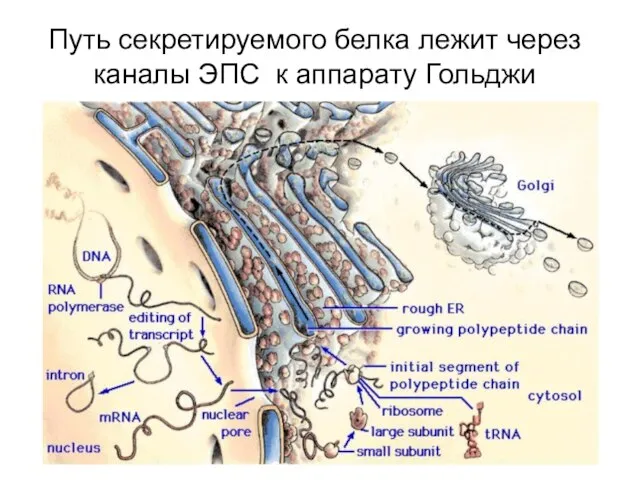
Путь секретируемого белка лежит через каналы ЭПС к аппарату Гольджи
Путь секретируемого белка лежит через каналы ЭПС к аппарату Гольджи
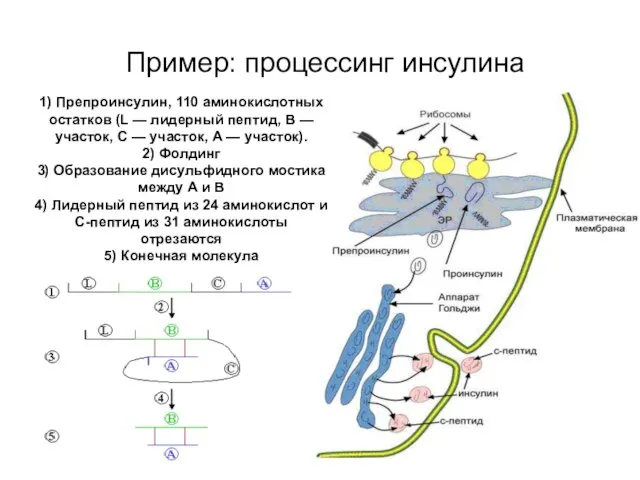
Пример: процессинг инсулина
1) Препроинсулин, 110 аминокислотных остатков (L — лидерный пептид, B —
Пример: процессинг инсулина
1) Препроинсулин, 110 аминокислотных остатков (L — лидерный пептид, B —
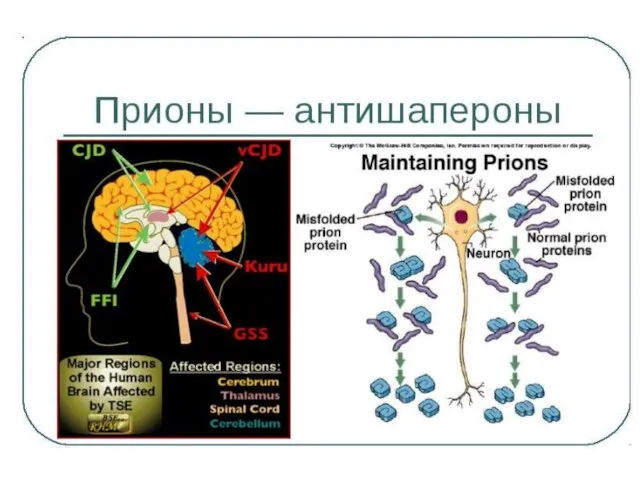

Неправильный фолдинг и прионные болезни
Неправильный фолдинг и прионные болезни

Регуляция экспрессии генов у эукариот
Регуляция экспрессии генов у эукариот

История вопроса
История вопроса

XIX век. Август Вейсман и теория детерминант
половая клетка
дифференцировка и смерть
клетки тела
непрерывный
XIX век. Август Вейсман и теория детерминант
половая клетка
дифференцировка и смерть
клетки тела
непрерывный

Это справедливо в части случаях: у некоторых червей и членистоногих
Вейсман опирался
Это справедливо в части случаях: у некоторых червей и членистоногих
Вейсман опирался
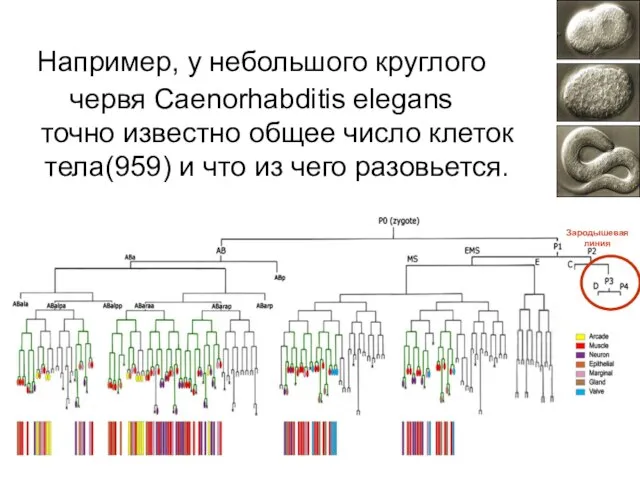
точно известно общее число клеток тела(959) и что из чего разовьется.
точно известно общее число клеток тела(959) и что из чего разовьется.
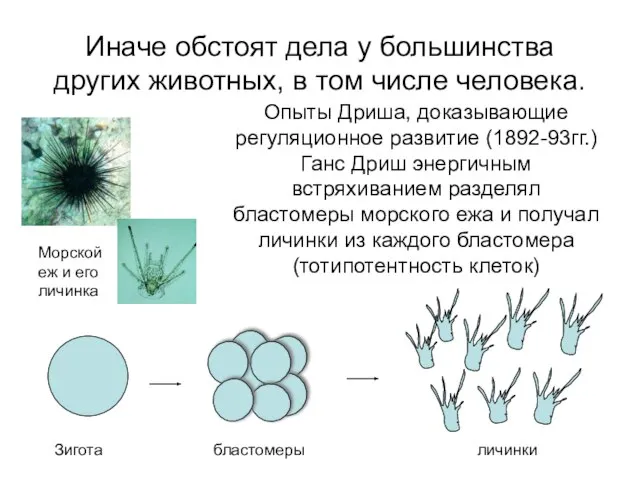
Иначе обстоят дела у большинства других животных, в том числе человека.
Иначе обстоят дела у большинства других животных, в том числе человека.

Развитие стали представлять как процесс включения и выключения генов
(а не
Развитие стали представлять как процесс включения и выключения генов (а не

В ХХ века была сформулирована гипотеза дифференциальной активности генов
Ядро каждой клетки
В ХХ века была сформулирована гипотеза дифференциальной активности генов
Ядро каждой клетки
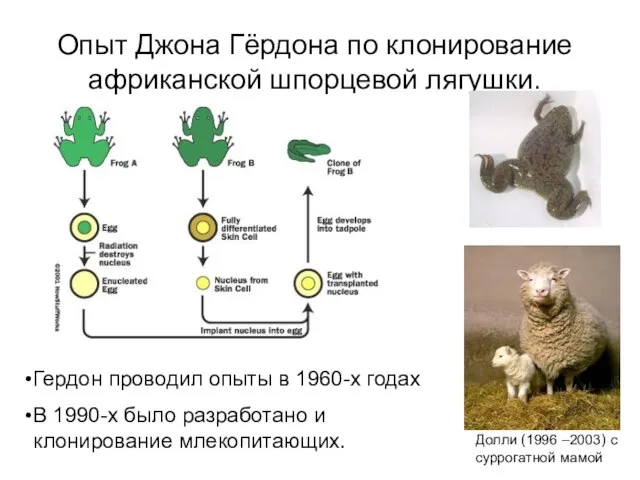
Опыт Джона Гёрдона по клонирование африканской шпорцевой лягушки.
Гердон проводил опыты в
Опыт Джона Гёрдона по клонирование африканской шпорцевой лягушки.
Гердон проводил опыты в
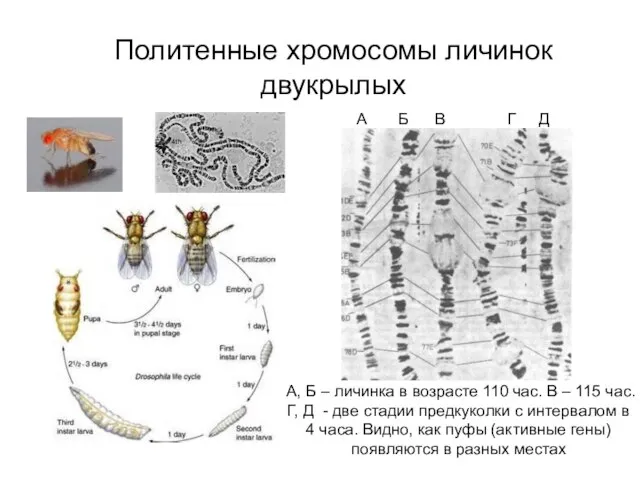
Политенные хромосомы личинок двукрылых
Dolly the sheep
А, Б – личинка в возрасте
Политенные хромосомы личинок двукрылых
Dolly the sheep
А, Б – личинка в возрасте

Гены можно разделить на
конститутивные, т.е. всегда активные.
Их иногда называют «генами
Гены можно разделить на
конститутивные, т.е. всегда активные.
Их иногда называют «генами
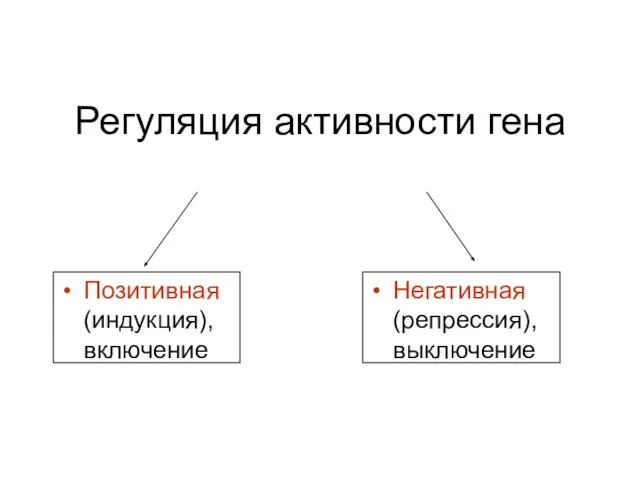
Регуляция активности гена
Позитивная (индукция), включение
Негативная (репрессия), выключение
Регуляция активности гена
Позитивная (индукция), включение
Негативная (репрессия), выключение

Регуляция экспрессии генов у прокариот
См. Предыдущую лекцию
Регуляция экспрессии генов у прокариот
См. Предыдущую лекцию

Регуляция экспрессии генов у эукариот
Регуляция экспрессии генов у эукариот

Регуляция экспрессии генов у эукариот происходит на всех этапах синтеза белка.
Изменение
Регуляция экспрессии генов у эукариот происходит на всех этапах синтеза белка.
Изменение

1. Число копий гена
1. Число копий гена
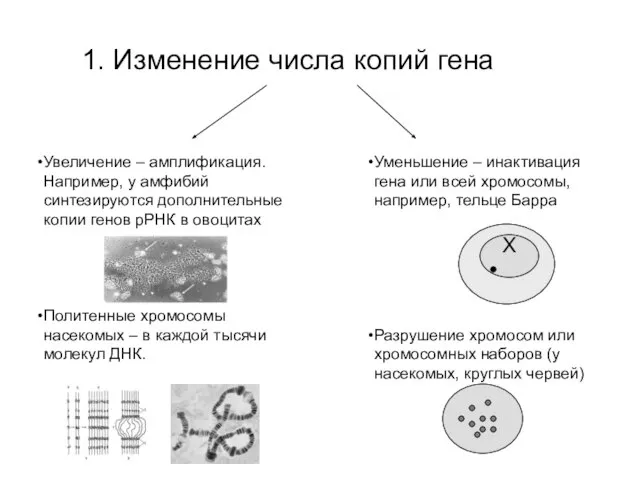
1. Изменение числа копий гена
Х
Увеличение – амплификация. Например, у амфибий синтезируются
1. Изменение числа копий гена
Х
Увеличение – амплификация. Например, у амфибий синтезируются
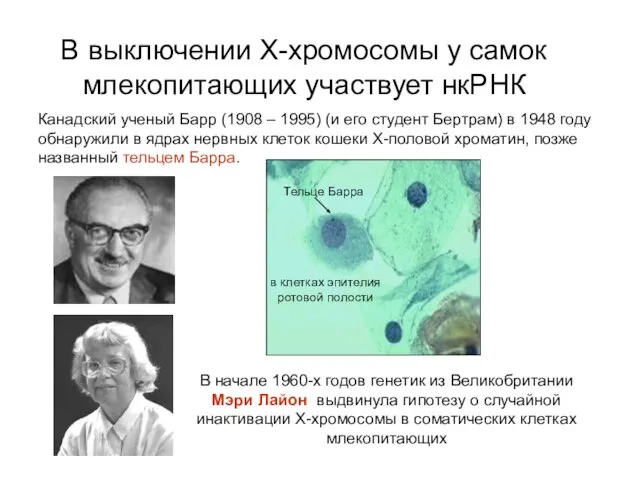
В выключении Х-хромосомы у самок млекопитающих участвует нкРНК
Канадский ученый Барр (1908
В выключении Х-хромосомы у самок млекопитающих участвует нкРНК
Канадский ученый Барр (1908
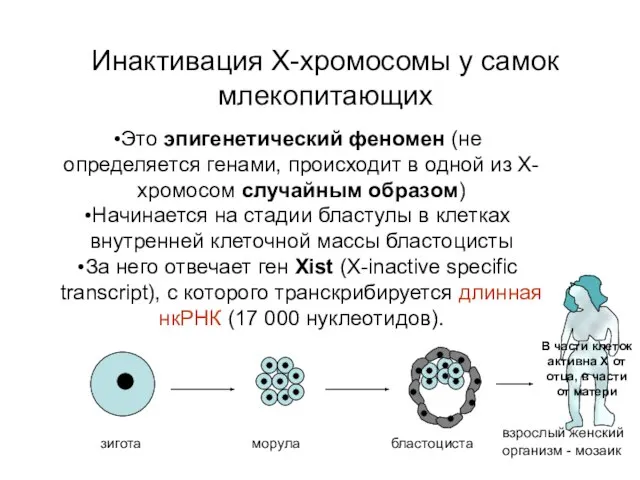
Инактивация Х-хромосомы у самок млекопитающих
В части клеток активна Х от отца,
Инактивация Х-хромосомы у самок млекопитающих
В части клеток активна Х от отца,
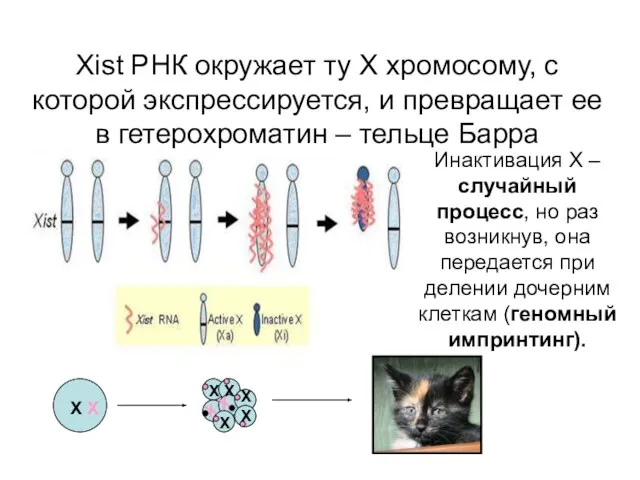
Xist РНК окружает ту Х хромосому, с которой экспрессируется, и превращает
Xist РНК окружает ту Х хромосому, с которой экспрессируется, и превращает

2. Транскрипция
Самый частый уровень регуляции экспрессии гена
2. Транскрипция
Самый частый уровень регуляции экспрессии гена
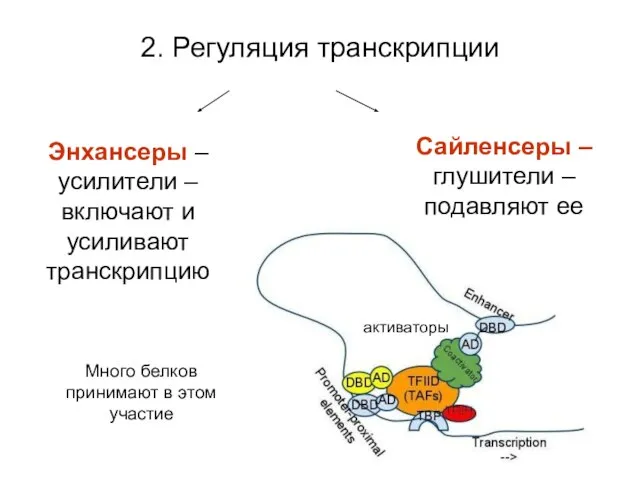
2. Регуляция транскрипции
Много белков принимают в этом участие
Энхансеры – усилители
2. Регуляция транскрипции
Много белков принимают в этом участие
Энхансеры – усилители
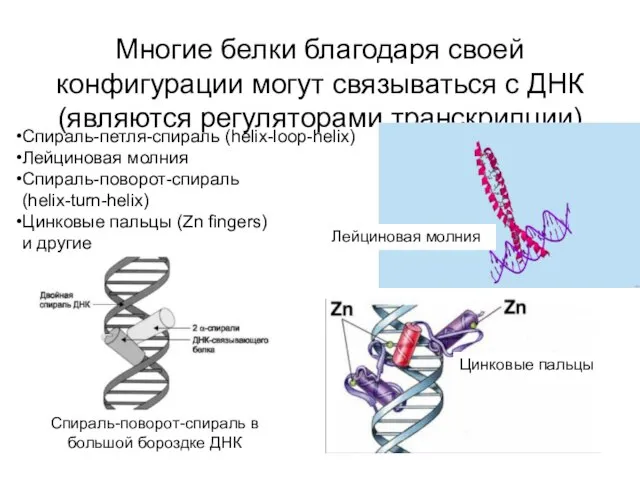
Многие белки благодаря своей конфигурации могут связываться с ДНК (являются регуляторами
Многие белки благодаря своей конфигурации могут связываться с ДНК (являются регуляторами
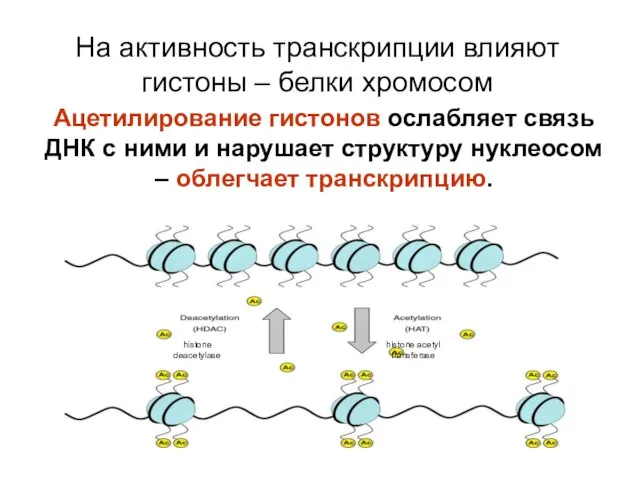
На активность транскрипции влияют гистоны – белки хромосом
Ацетилирование гистонов ослабляет
На активность транскрипции влияют гистоны – белки хромосом
Ацетилирование гистонов ослабляет
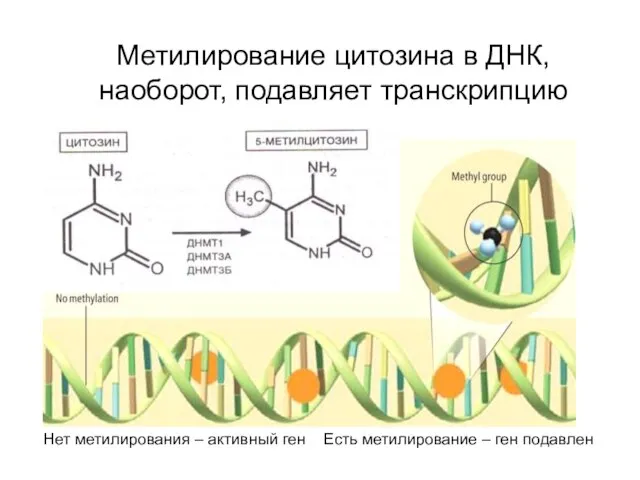
Метилирование цитозина в ДНК, наоборот, подавляет транскрипцию
Нет метилирования – активный ген
Метилирование цитозина в ДНК, наоборот, подавляет транскрипцию
Нет метилирования – активный ген
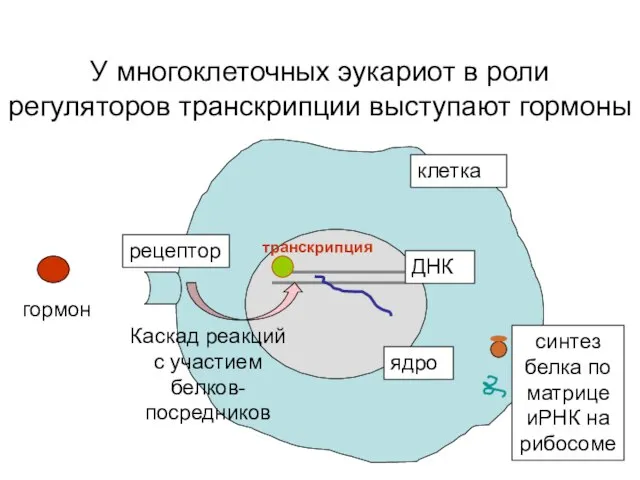
У многоклеточных эукариот в роли регуляторов транскрипции выступают гормоны
клетка
ядро
ДНК
гормон
рецептор
транскрипция
Каскад реакций с
У многоклеточных эукариот в роли регуляторов транскрипции выступают гормоны
клетка
ядро
ДНК
гормон
рецептор
транскрипция
Каскад реакций с
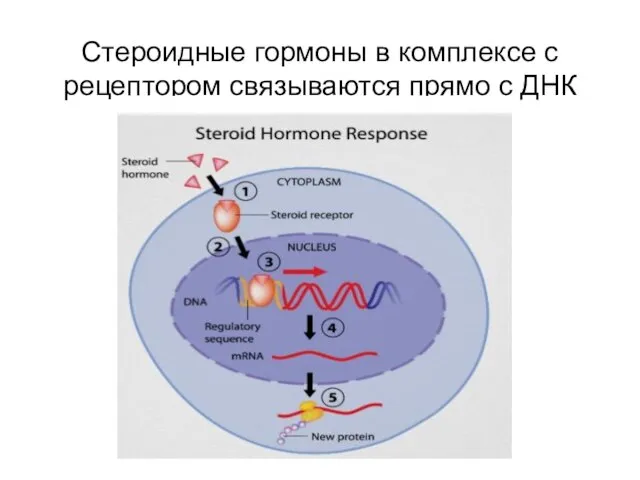
Стероидные гормоны в комплексе с рецептором связываются прямо с ДНК
Стероидные гормоны в комплексе с рецептором связываются прямо с ДНК

3. Процессинг РНК
3. Процессинг РНК
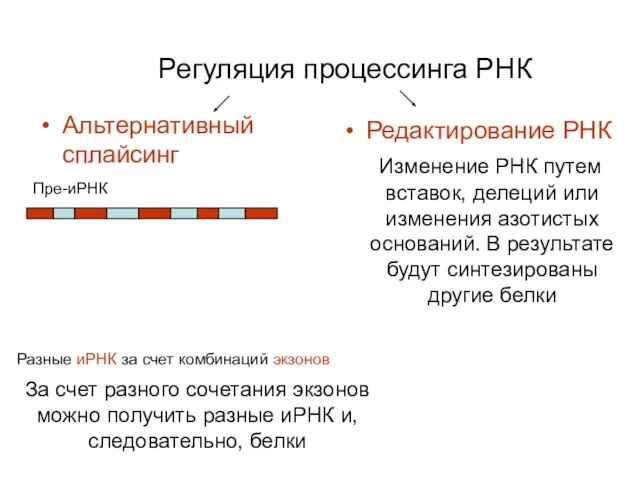
Регуляция процессинга РНК
Альтернативный сплайсинг
Редактирование РНК
Изменение РНК путем вставок,
Регуляция процессинга РНК
Альтернативный сплайсинг
Редактирование РНК
Изменение РНК путем вставок,
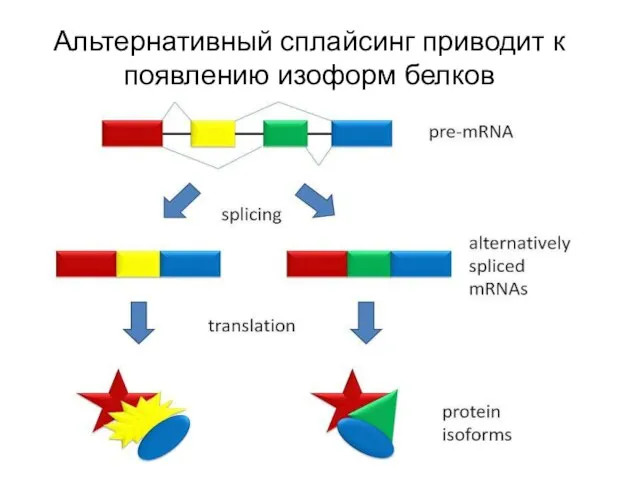
Альтернативный сплайсинг приводит к появлению изоформ белков
Альтернативный сплайсинг приводит к появлению изоформ белков

Показано, что у человека 94 % генов подвержено альтернативному сплайсингу (у остальных
Показано, что у человека 94 % генов подвержено альтернативному сплайсингу (у остальных
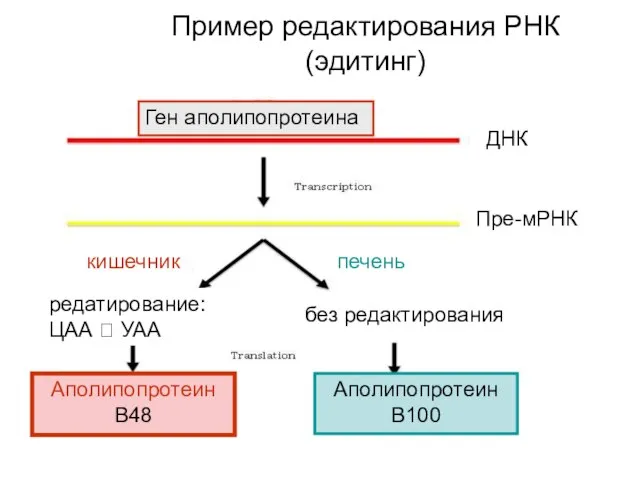
Пример редактирования РНК (эдитинг)
Ген аполипопротеина
ДНК
Пре-мРНК
кишечник
печень
без редактирования
редатирование: ЦАА ? УАА
Аполипопротеин В48
Аполипопротеин В100
Пример редактирования РНК (эдитинг)
Ген аполипопротеина
ДНК
Пре-мРНК
кишечник
печень
без редактирования
редатирование: ЦАА ? УАА
Аполипопротеин В48
Аполипопротеин В100

4. Трансляция
4. Трансляция

Регуляция трансляции
Удлинение или укорочение времени жизни иРНК:
Пример: гормон пролактин удлиняет
Регуляция трансляции
Удлинение или укорочение времени жизни иРНК:
Пример: гормон пролактин удлиняет

5. Процессинг белка
5. Процессинг белка

5. Регуляция процессинга белка
Коллаген – основной белок соединительной ткани, около 30%
5. Регуляция процессинга белка
Коллаген – основной белок соединительной ткани, около 30%
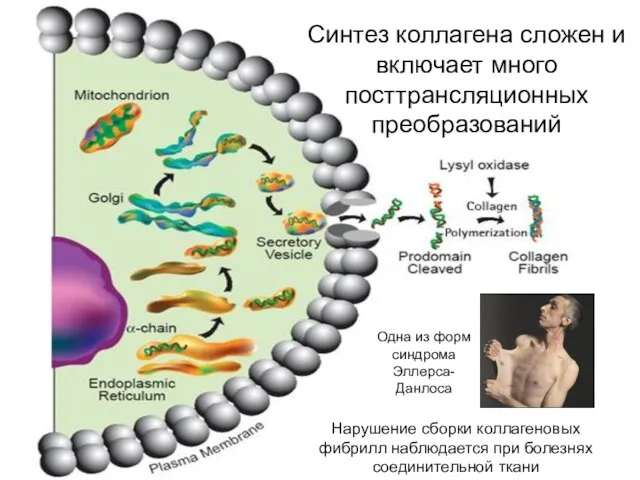
Синтез коллагена сложен и включает много посттрансляционных преобразований
Нарушение сборки коллагеновых фибрилл
Синтез коллагена сложен и включает много посттрансляционных преобразований
Нарушение сборки коллагеновых фибрилл

Несса Кери пишет:
Несса Кери пишет:
 Систематика грибов
Систематика грибов Эпителиально-стромальные соотношения слизистой слепой кишки при питании диспергированной пищей
Эпителиально-стромальные соотношения слизистой слепой кишки при питании диспергированной пищей Деревья России
Деревья России Строение органов дыхания у беспозвоночных и позвоночных животных
Строение органов дыхания у беспозвоночных и позвоночных животных Популяция как форма существования вида
Популяция как форма существования вида Технология критического мышления в процессе преподавания биологии Бабушкина Ж.В. учитель биологии и географии МБОУ «Еланская С
Технология критического мышления в процессе преподавания биологии Бабушкина Ж.В. учитель биологии и географии МБОУ «Еланская С Цитоплазма (2 фракции)
Цитоплазма (2 фракции) Типы питания животных
Типы питания животных Подвесной сад Кокедама
Подвесной сад Кокедама Бактериологическое исследование мясных консервов
Бактериологическое исследование мясных консервов Методологический редукционизм в генетике
Методологический редукционизм в генетике Работу выполнила учитель начальных классов МКОУ Советской СОШ
Работу выполнила учитель начальных классов МКОУ Советской СОШ  Развитие психики в онтогенезе. Постнатальный период. Ранний постнатальный период
Развитие психики в онтогенезе. Постнатальный период. Ранний постнатальный период Влияние качества воды реки Еруслан на здоровье школьников Выполнил: ученик 7 класса Довыденко Алексей МОУ - СОШ с.Карпенк
Влияние качества воды реки Еруслан на здоровье школьников Выполнил: ученик 7 класса Довыденко Алексей МОУ - СОШ с.Карпенк Презентация на тему "Грибы. Разнообразие грибов. Значение грибов в природе и жизни человека" - скачать презентации по Биологи
Презентация на тему "Грибы. Разнообразие грибов. Значение грибов в природе и жизни человека" - скачать презентации по Биологи Презентация на тему Размножение и индивидуальное развитие организмов Биология 10 класс
Презентация на тему Размножение и индивидуальное развитие организмов Биология 10 класс  Витамины
Витамины Птицы. Первые вестники весны
Птицы. Первые вестники весны Виды и породы животных
Виды и породы животных Сохранить планету для людей
Сохранить планету для людей Вклад Д.И. Менделеева и Г. Менделя в развитие наук
Вклад Д.И. Менделеева и Г. Менделя в развитие наук Генетика. Взаимодействие генов
Генетика. Взаимодействие генов Кактус
Кактус Зоология позвоночных
Зоология позвоночных Презентация на тему "Антропоген" - скачать презентации по Биологии
Презентация на тему "Антропоген" - скачать презентации по Биологии ЭНДОКРИННАЯ СИСТЕМА. Тесты. Шильцова Наталья Петровна - учитель биологии МОУ СОШ№9, пос. Восточно-Горняцкий(Горняцкий) г Белая Кали
ЭНДОКРИННАЯ СИСТЕМА. Тесты. Шильцова Наталья Петровна - учитель биологии МОУ СОШ№9, пос. Восточно-Горняцкий(Горняцкий) г Белая Кали Презентация на тему "Генетика пола" - скачать презентации по Биологии
Презентация на тему "Генетика пола" - скачать презентации по Биологии Презентация на тему "Лекарственные растения" - скачать презентации по Биологии
Презентация на тему "Лекарственные растения" - скачать презентации по Биологии