- Генетика мікроорганізмів

Содержание
- 2. План лекції Будова клітинного генетичного апарату. Позахромосомні елементи спадковості. Мутації. Рекомбінації. Основи генної інженерії. Молекулярні біотехнології.
- 3. Історія розвитку молекулярної біотехнології
- 4. Історія розвитку молекулярної біотехнології
- 5. F. Crick i J. Watson – відкривачі структури ДНК
- 7. Генетичний матеріал у бактерій представлений: хромосомою позахромосомними елементами спадковості: плазмідами транспозонами IS-елементами
- 8. Плазміди можуть мати лінійну або кільцеву структуру, є необов‘язковими компонентами мікробної клітини; здатні до самостійної реплікації.
- 9. Мігруючі генетичні елементи TGATC ACTAG GCGTATCG CGCATAGC TGATC ACTAG GCGTATCG CGCATAGC Гени для транспозиції IS IS
- 10. 1. Координуюча: взаємодія транспозонів, плазмід, помірних фагів між собою та хромосомою бактерії, забезпечуючи їх реплікацію. 2.
- 11. 1. Регуляторна. 2. Кодуюча. 3. Індукують генні мутації за типом делеції або інверсії. 4. Включення їх
- 12. Класифікація плазмід За розміщенням в клітині: позахромосомні інтегровані За типом передачі: кон’югативні (трансмісивні, мають tra-ген) некон’югативні
- 13. Сol – продукція коліцинів HLy – продукція гемолізинів Tol – розщеплення толуолу, ксилолу Ent – продукція
- 14. Функціональні властивості плазмід Антибіотико резистентність пеніцилін Загибель клітини R-плазміда пеніцилін Розмноження антибітикоcтійких бактерій фертильність реципієнт F-плазміда
- 15. 1. Регуляторна (якщо ген не здатний реплікуватись за рахунок втрати його частини, плазміди вносять власний реплікон).
- 16. високий темп втрати ознаки збільшення темпів втрати ознаки під впливом температури або хімічних сполук спільна втрата
- 17. За походженням: спонтанні; індуковані. За локалізацією: нуклеоїдні; цитоплазматичні. За кількістю генів, що мутували: генні; хромосомні. За
- 18. інверсія дуплікація делеція дислокація Хромосомні мутації:
- 19. делеція інсерція (вставка) заміна: транзиція (пуринова основа на пуринову, піримідинова – на піримідинову) трансверзія (пуринова основа
- 20. 1. Пряма мутація 2. Зворотна (обернена) мутація або реверсія 3. Супресорна мутація (внутрішньогенна, позагенна). Веде до
- 21. Мутагенні фактори Фізичні: 1. УФО (λ-260 НМ) – найсильніша мутагенна дія – утворюються димери тиміну, заміна
- 22. Мутагенні фактори Хімічні: 1. Азотиста кислота - дезамінує нуклеотиди 2. N-нітрозометилсечовина – супермутаген, канцероген 3. Етилметансульфонат
- 23. Дія різних мутагенів на бактерії Деякі фізичні й хімічні фактори підвищують частоту мутацій. Ультрафіолетове випромінювання та
- 24. R і S форми колоній
- 25. Властивості мікробів S-колоній: Клітини нормальної морфології Дифузне помутніння бульйону У рухомих видів є джгутики У капсульних
- 26. Методи виявлення мутантів За різницею в швидкості росту (посів на мінімальне середовище; мутанти виростають) Різна здатність
- 27. Метод реплік для виявлення ауксотрофних мутантів Повноцінне середовище Мінімальне середовище (ауксотрофи не ростуть)
- 28. Світлова репарація – Репарація тимінових димерів Гуанін Цитозин Тимін Аденін Ковалентний звґязок (тиміновий димер) розщеплення тимінових
- 29. Темнова репарація 1. Деградація прилеглих до пошкодженої ділянок ДНК 2. Вирізання за допомогою рестриктаз пошкоджених ділянок
- 30. SOS-реактивація При множинних ушкодженнях ділянки з мутаціями переводяться в неактивний стан, а їх роль виконує неушкоджена
- 31. ТРАНСФОРМАЦІЯ Бактеріальна хромосома Фрагменти ДНК Інтеграція ДНК в хромосому Деградація А В ДНК плазміди Бактеріальна хромосома
- 32. Трансформація (досліди Грифітса, 1928; Евері Мк Леода і Макарті, 1944)
- 33. ТРАНСДУКЦІЯ Викликають помірні, дефектні фаги. Види: загальна (генералізована) специфічна абортивна
- 34. ТРАНСДУКЦІЯ (Ціндер і Ледерберг, 1952) ДНК бактеріофагу Хромосома клітини хазяїна Клітина А (донор) Частинки фагу Лізис
- 35. Спеціалізована трансдукція
- 36. ВІДМІННОСТІ ТРАНСДУКЦІЇ ТА ФАГОВОЇ КОНВЕРСІЇ Трансдукція – перенос генетичної інформації з клітини в клітину за допомогою
- 37. КОН’ЮГАЦІЯ та її механізм (Ледерберг і Тейтум, 1946)
- 38. Кон’югація
- 39. рекомбінації Трансдукція Кон’югація Трансформація
- 40. Молеку- лярна біологія Мікро- біологія Біохімія Хімічна інженерія Генетика Клітинна біологія Молекулярна біотехнологія Високо- врожайні культури
- 41. ПРОДУЦЕНТИ, що найчастіше використовуються в біотехнології ЕУКАРІОТИ – дріжджі, плісняви, культури клітин тварин, людини та рослин
- 42. Хромосомна карта E. coli
- 43. Переваги одноклітинних продуцентів: велика швидкість розмноження використання простих, дешевих субстратів можливість одержання "суперпродуцентів" шляхом генетичної селекції
- 44. Біотехнологічні продукти мікроорганізмів - продуцентів самі клітини як джерело цільового продукту крупні молекули (ферменти, токсини, антигени,
- 45. СФЕРИ ВИКОРИСТАННЯ БІОТЕХНОЛОГІЇ
- 46. Основні продукти, які отримують за допомогою біотехнології
- 47. Деякі гормони людини, які продукуються рекомбінантними мікроорганізмами
- 48. ГЕНЕТИЧНА ІНЖЕНЕРІЯ – спрямована зміна геному продуцента в потрібному для людини напрямку: пересадка в геном продуцента
- 49. "ІНСТРУМЕНТИ" ДЛЯ ГЕНЕТИЧНОЇ ІНЖЕНЕРІЇ ФЕРМЕНТИ (рестриктази, лігази, зворотня транскріптаза) ВЕКТОРИ (плазміди, помірні бактеріофаги, косміди, транспозони, віруси)
- 50. СХЕМА ГЕНЕТИЧНО - ІНЖЕНЕРНОГО ЕКСПЕРИМЕНТУ визначення локалізації потрібного гену (сиквенс, генетичне мапування) - клонування (виділення) потрібного
- 51. БІОТЕХНОЛОГІЯ Ферменти, ліки Рекомбінантні рослини Білки Ядро Ізольований ген Вставка у вектор
- 52. ГЕНОТЕРАПІЯ
- 53. ГЕНЕТИКА ВІРУСІВ Способи збільшення інформації: дворазове зчитування одієї іРНК з інших ініціюючих кодонів зсув рамки трансляції
- 54. У вірусів можуть бути: Модифікації (зміна складу білків капсиду, суперкапсиду під впливом клітин). Мутації (розмір бляшок
- 55. ВИДИ ГЕНЕТИЧНИХ РЕКОМБІНАЦІЙ У ВІРУСІВ 1. Рекомбінація: Обмін генами (міжгенна) та їх частинами (внутрішньогенна) Схрещування близьких
- 56. ВИДИ ГЕНЕТИЧНИХ РЕКОМБІНАЦІЙ У ВІРУСІВ 2. Множинна реактивація: вірусна інфекція викликається при зараженні віріонами з пошкодженим
- 57. ВИДИ РЕКОМБІНАЦІЙ У ВІРУСІВ 4. Гетерозиготність: одночасній репродукції декількох віріонів, різних за спадковими властивостями, утворюються віріони,
- 58. ВИДИ РЕКОМБІНАЦІЙ У ВІРУСІВ 5. Транскапсидація: частина чужерідного генетичного матерілау, заключеного всередині капсиду іншого вірусу, здатна
- 59. ВИДИ РЕКОМБІНАЦІЙ У ВІРУСІВ 6. Крос-реактивація (рятування маркера): реактивація інактивованого геному неінактивованим (подібно до множинної реактивації)
- 61. Скачать презентацию

План лекції
Будова клітинного генетичного апарату.
Позахромосомні елементи спадковості.
Мутації.
Рекомбінації.
Основи генної інженерії. Молекулярні
План лекції
Будова клітинного генетичного апарату.
Позахромосомні елементи спадковості.
Мутації.
Рекомбінації.
Основи генної інженерії. Молекулярні

Історія розвитку молекулярної біотехнології
Історія розвитку молекулярної біотехнології

Історія розвитку молекулярної біотехнології
Історія розвитку молекулярної біотехнології

F. Crick i J. Watson –
відкривачі структури ДНК
F. Crick i J. Watson –
відкривачі структури ДНК


Генетичний матеріал
у бактерій представлений:
хромосомою
позахромосомними елементами спадковості:
плазмідами
транспозонами
IS-елементами
Генетичний матеріал
у бактерій представлений:
хромосомою
позахромосомними елементами спадковості:
плазмідами
транспозонами
IS-елементами

Плазміди можуть мати лінійну або кільцеву структуру, є необов‘язковими компонентами мікробної
Плазміди можуть мати лінійну або кільцеву структуру, є необов‘язковими компонентами мікробної
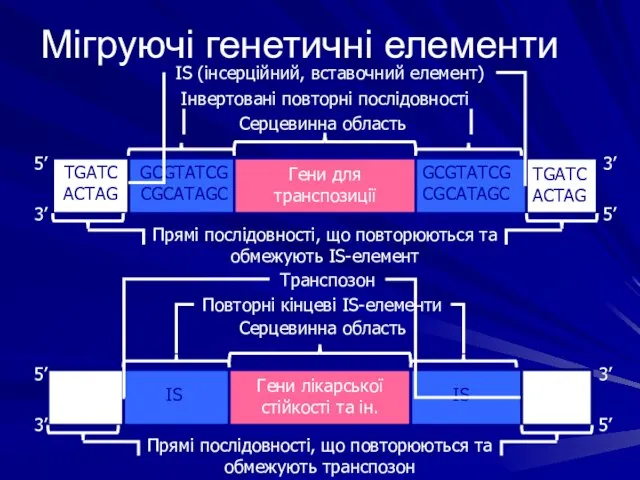
Мігруючі генетичні елементи
TGATC
ACTAG
GCGTATCG
CGCATAGC
TGATC
ACTAG
GCGTATCG
CGCATAGC
Гени для транспозиції
IS
IS
Гени лікарської стійкості та ін.
5’
3’
3’
5’
3’
5’
3’
5’
Прямі послідовності, що
Мігруючі генетичні елементи
TGATC
ACTAG
GCGTATCG
CGCATAGC
TGATC
ACTAG
GCGTATCG
CGCATAGC
Гени для транспозиції
IS
IS
Гени лікарської стійкості та ін.
5’
3’
3’
5’
3’
5’
3’
5’
Прямі послідовності, що

1. Координуюча: взаємодія транспозонів, плазмід, помірних фагів між собою та хромосомою
1. Координуюча: взаємодія транспозонів, плазмід, помірних фагів між собою та хромосомою

1. Регуляторна.
2. Кодуюча.
3. Індукують генні мутації за типом делеції або інверсії.
4.
1. Регуляторна.
2. Кодуюча.
3. Індукують генні мутації за типом делеції або інверсії.
4.

Класифікація плазмід
За розміщенням в клітині:
позахромосомні
інтегровані
За типом передачі:
Класифікація плазмід
За розміщенням в клітині:
позахромосомні
інтегровані
За типом передачі:

Сol – продукція коліцинів
HLy – продукція гемолізинів
Tol – розщеплення толуолу, ксилолу
Сol – продукція коліцинів
HLy – продукція гемолізинів
Tol – розщеплення толуолу, ксилолу

Функціональні властивості плазмід
Антибіотико
резистентність
пеніцилін
Загибель клітини
R-плазміда
пеніцилін
Розмноження антибітикоcтійких бактерій
фертильність
реципієнт
F-плазміда
F-пілі
вірулентність
Не продукує токсин
плазміда
вірулентності
токсин
донор
метаболізм
плазміда
метаболізму
Функціональні властивості плазмід
Антибіотико
резистентність
пеніцилін
Загибель клітини
R-плазміда
пеніцилін
Розмноження антибітикоcтійких бактерій
фертильність
реципієнт
F-плазміда
F-пілі
вірулентність
Не продукує токсин
плазміда
вірулентності
токсин
донор
метаболізм
плазміда
метаболізму

1. Регуляторна (якщо ген не здатний реплікуватись за рахунок втрати його
1. Регуляторна (якщо ген не здатний реплікуватись за рахунок втрати його

високий темп втрати ознаки
збільшення темпів втрати ознаки під впливом температури або
високий темп втрати ознаки
збільшення темпів втрати ознаки під впливом температури або

За походженням: спонтанні;
індуковані.
За локалізацією: нуклеоїдні;
цитоплазматичні.
За
За походженням: спонтанні;
індуковані.
За локалізацією: нуклеоїдні;
цитоплазматичні.
За

інверсія
дуплікація
делеція
дислокація
Хромосомні мутації:
інверсія
дуплікація
делеція
дислокація
Хромосомні мутації:

делеція
інсерція (вставка)
заміна:
транзиція (пуринова основа на пуринову, піримідинова – на
делеція
інсерція (вставка)
заміна:
транзиція (пуринова основа на пуринову, піримідинова – на

1. Пряма мутація
2. Зворотна (обернена) мутація або реверсія
3. Супресорна мутація (внутрішньогенна,
1. Пряма мутація
2. Зворотна (обернена) мутація або реверсія
3. Супресорна мутація (внутрішньогенна,

Мутагенні фактори
Фізичні:
1. УФО (λ-260 НМ) – найсильніша мутагенна дія – утворюються
Мутагенні фактори
Фізичні:
1. УФО (λ-260 НМ) – найсильніша мутагенна дія – утворюються

Мутагенні фактори
Хімічні:
1. Азотиста кислота - дезамінує нуклеотиди
2. N-нітрозометилсечовина – супермутаген, канцероген
3.
Мутагенні фактори
Хімічні:
1. Азотиста кислота - дезамінує нуклеотиди
2. N-нітрозометилсечовина – супермутаген, канцероген
3.
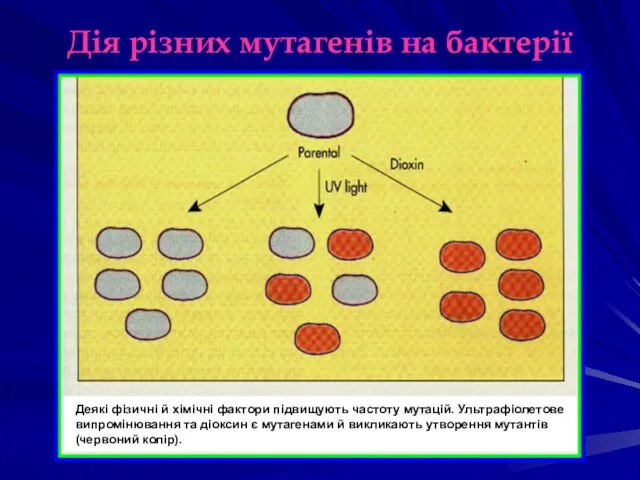
Дія різних мутагенів на бактерії
Деякі фізичні й хімічні фактори підвищують частоту
Дія різних мутагенів на бактерії
Деякі фізичні й хімічні фактори підвищують частоту

R і S форми колоній
R і S форми колоній

Властивості мікробів S-колоній:
Клітини нормальної морфології
Дифузне помутніння бульйону
У рухомих видів є джгутики
У
Властивості мікробів S-колоній:
Клітини нормальної морфології
Дифузне помутніння бульйону
У рухомих видів є джгутики
У

Методи виявлення мутантів
За різницею в швидкості росту (посів на мінімальне середовище;
Методи виявлення мутантів
За різницею в швидкості росту (посів на мінімальне середовище;
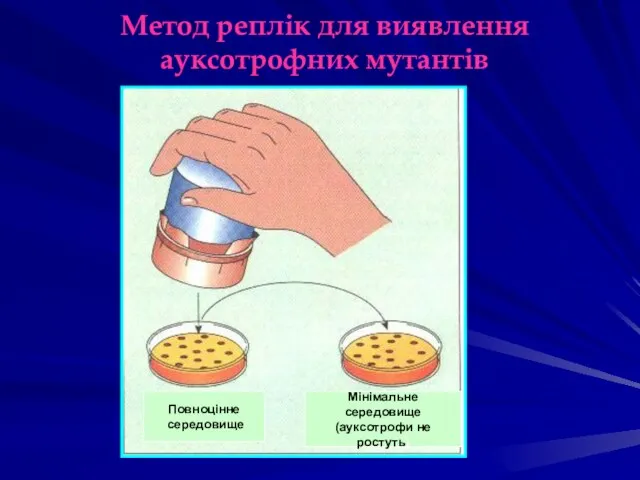
Метод реплік для виявлення ауксотрофних мутантів
Повноцінне
середовище
Мінімальне середовище
(ауксотрофи не ростуть)
Метод реплік для виявлення ауксотрофних мутантів
Повноцінне
середовище
Мінімальне середовище
(ауксотрофи не ростуть)
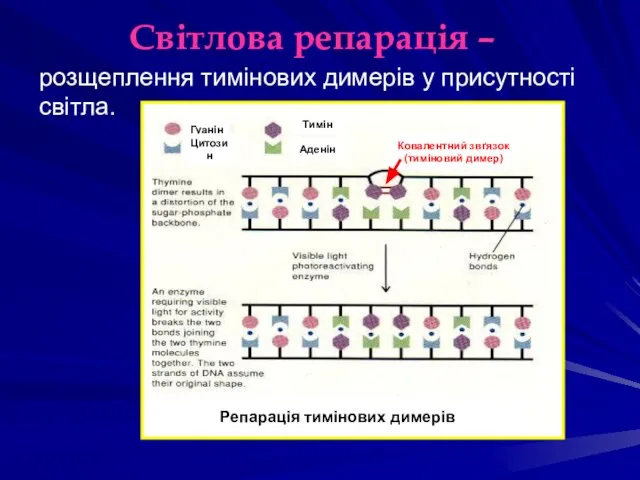
Світлова репарація –
Репарація тимінових димерів
Гуанін
Цитозин
Тимін
Аденін
Ковалентний звґязок
(тиміновий димер)
розщеплення тимінових
Світлова репарація –
Репарація тимінових димерів
Гуанін
Цитозин
Тимін
Аденін
Ковалентний звґязок
(тиміновий димер)
розщеплення тимінових
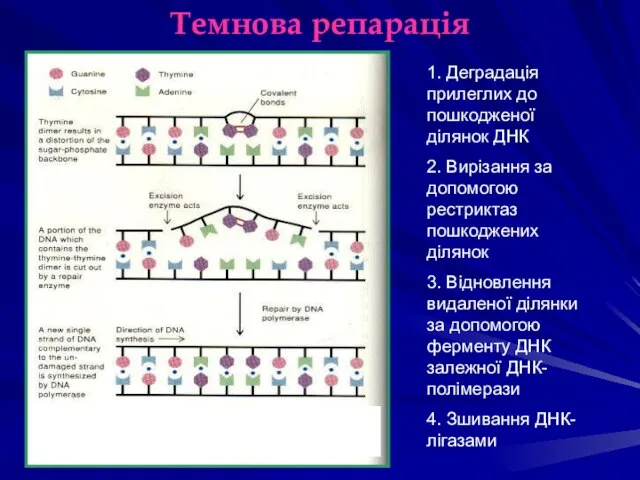
Темнова репарація
1. Деградація прилеглих до пошкодженої ділянок ДНК
2. Вирізання за допомогою
Темнова репарація
1. Деградація прилеглих до пошкодженої ділянок ДНК
2. Вирізання за допомогою

SOS-реактивація
При множинних ушкодженнях ділянки з мутаціями переводяться в неактивний стан, а
SOS-реактивація
При множинних ушкодженнях ділянки з мутаціями переводяться в неактивний стан, а
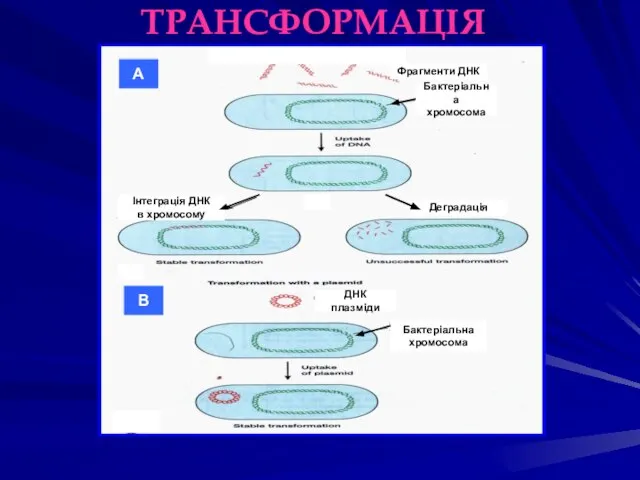
ТРАНСФОРМАЦІЯ
Бактеріальна
хромосома
Фрагменти ДНК
Інтеграція ДНК
в хромосому
Деградація
А
В
ДНК плазміди
Бактеріальна
хромосома
ТРАНСФОРМАЦІЯ
Бактеріальна
хромосома
Фрагменти ДНК
Інтеграція ДНК
в хромосому
Деградація
А
В
ДНК плазміди
Бактеріальна
хромосома
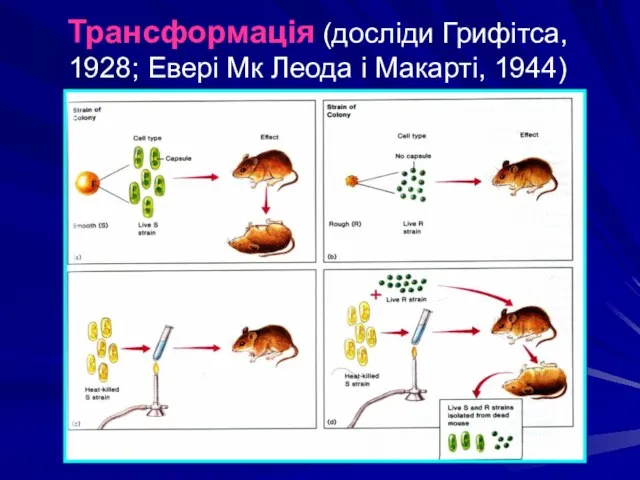
Трансформація (досліди Грифітса, 1928; Евері Мк Леода і Макарті, 1944)
Трансформація (досліди Грифітса, 1928; Евері Мк Леода і Макарті, 1944)

ТРАНСДУКЦІЯ
Викликають помірні, дефектні фаги.
Види:
загальна (генералізована)
специфічна
абортивна
ТРАНСДУКЦІЯ
Викликають помірні, дефектні фаги.
Види:
загальна (генералізована)
специфічна
абортивна
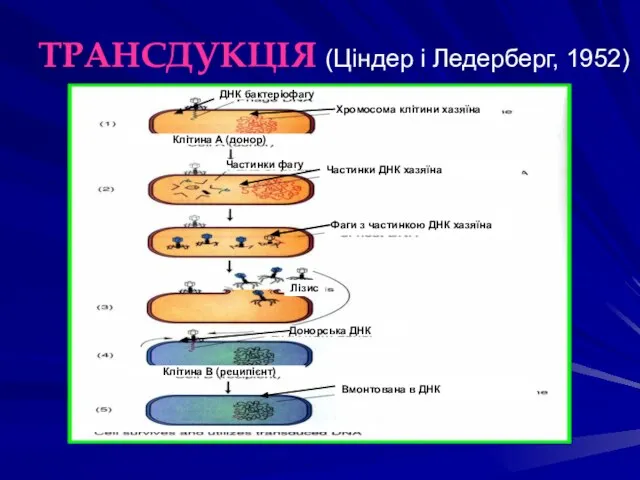
ТРАНСДУКЦІЯ (Ціндер і Ледерберг, 1952)
ДНК бактеріофагу
Хромосома клітини хазяїна
Клітина А (донор)
Частинки фагу
Лізис
Фаги
ТРАНСДУКЦІЯ (Ціндер і Ледерберг, 1952)
ДНК бактеріофагу
Хромосома клітини хазяїна
Клітина А (донор)
Частинки фагу
Лізис
Фаги
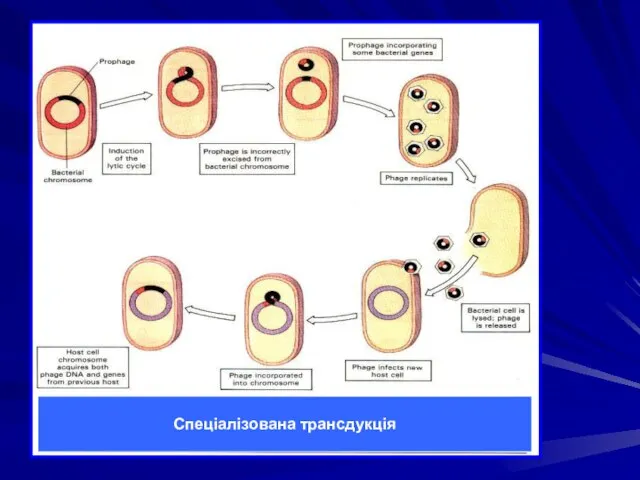
Спеціалізована трансдукція
Спеціалізована трансдукція

ВІДМІННОСТІ ТРАНСДУКЦІЇ ТА ФАГОВОЇ КОНВЕРСІЇ
Трансдукція – перенос генетичної інформації з клітини
ВІДМІННОСТІ ТРАНСДУКЦІЇ ТА ФАГОВОЇ КОНВЕРСІЇ
Трансдукція – перенос генетичної інформації з клітини
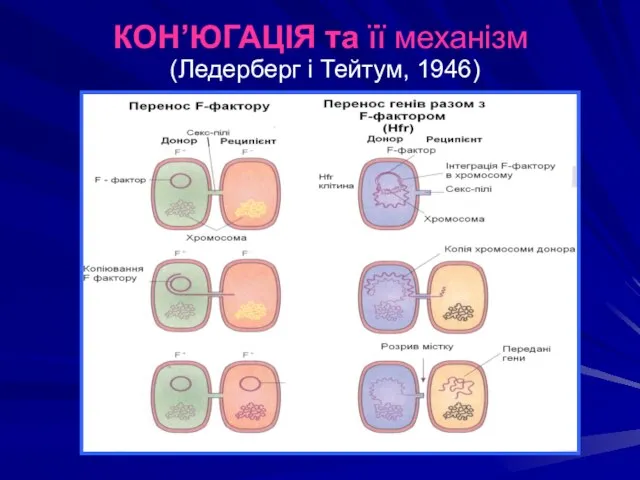
КОН’ЮГАЦІЯ та її механізм
(Ледерберг і Тейтум, 1946)
КОН’ЮГАЦІЯ та її механізм
(Ледерберг і Тейтум, 1946)
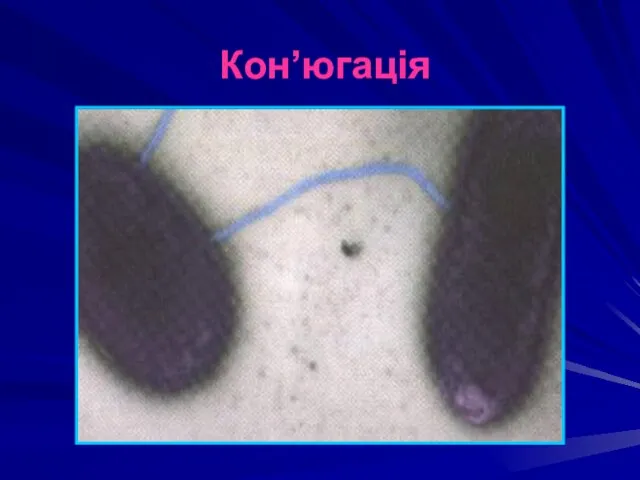
Кон’югація
Кон’югація
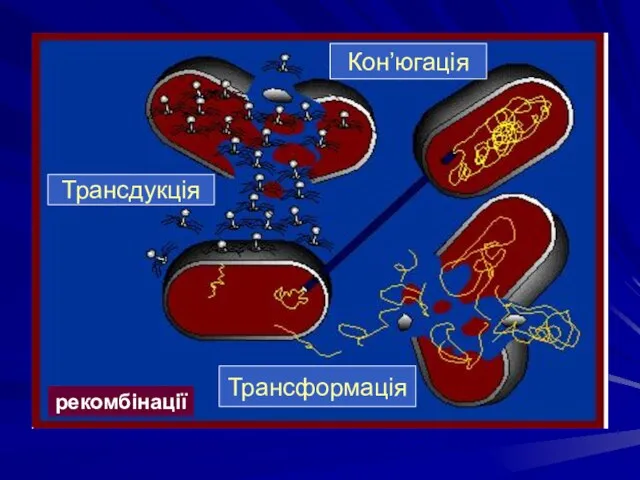
рекомбінації
Трансдукція
Кон’югація
Трансформація
рекомбінації
Трансдукція
Кон’югація
Трансформація
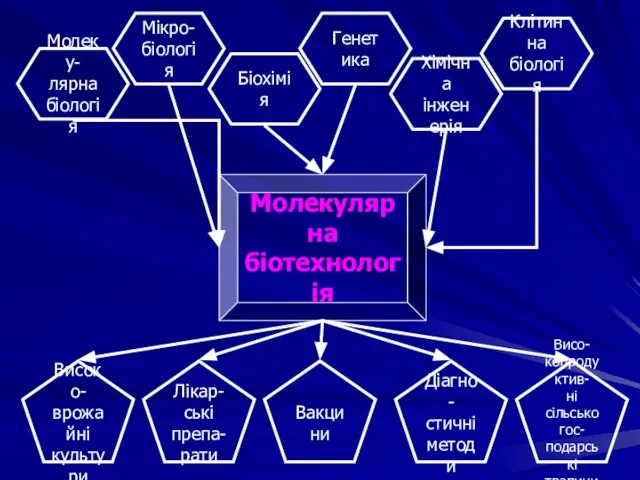
Молеку-
лярна
біологія
Мікро-
біологія
Біохімія
Хімічна
інженерія
Генетика
Клітинна
біологія
Молекулярна
біотехнологія
Високо-
врожайні
культури
Лікар-
ські препа-
рати
Вакцини
Діагно-
стичні
методи
Висо-
копродуктив-
ні сільськогос-
подарські
тварини
Молеку-
лярна
біологія
Мікро-
біологія
Біохімія
Хімічна
інженерія
Генетика
Клітинна
біологія
Молекулярна
біотехнологія
Високо-
врожайні
культури
Лікар-
ські препа-
рати
Вакцини
Діагно-
стичні
методи
Висо-
копродуктив-
ні сільськогос-
подарські
тварини

ПРОДУЦЕНТИ, що найчастіше використовуються в біотехнології
ЕУКАРІОТИ – дріжджі, плісняви, культури
ПРОДУЦЕНТИ, що найчастіше використовуються в біотехнології
ЕУКАРІОТИ – дріжджі, плісняви, культури
Хромосомна карта E. coli
Хромосомна карта E. coli

Переваги одноклітинних продуцентів:
велика швидкість розмноження
використання простих, дешевих субстратів
можливість одержання "суперпродуцентів" шляхом
Переваги одноклітинних продуцентів:
велика швидкість розмноження
використання простих, дешевих субстратів
можливість одержання "суперпродуцентів" шляхом

Біотехнологічні продукти мікроорганізмів - продуцентів
самі клітини як джерело цільового продукту
крупні молекули
Біотехнологічні продукти мікроорганізмів - продуцентів
самі клітини як джерело цільового продукту
крупні молекули
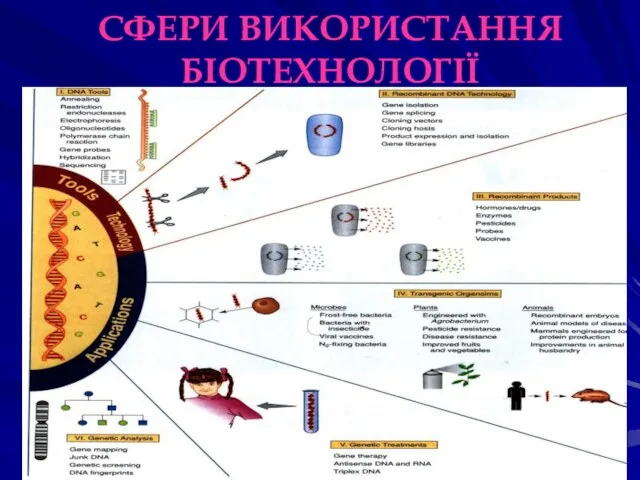
СФЕРИ ВИКОРИСТАННЯ БІОТЕХНОЛОГІЇ
СФЕРИ ВИКОРИСТАННЯ БІОТЕХНОЛОГІЇ

Основні продукти, які отримують за допомогою біотехнології
Основні продукти, які отримують за допомогою біотехнології

Деякі гормони людини, які продукуються рекомбінантними мікроорганізмами
Деякі гормони людини, які продукуються рекомбінантними мікроорганізмами

ГЕНЕТИЧНА ІНЖЕНЕРІЯ –
спрямована зміна геному продуцента в потрібному для людини напрямку:
ГЕНЕТИЧНА ІНЖЕНЕРІЯ –
спрямована зміна геному продуцента в потрібному для людини напрямку:

"ІНСТРУМЕНТИ"
ДЛЯ ГЕНЕТИЧНОЇ ІНЖЕНЕРІЇ
ФЕРМЕНТИ (рестриктази, лігази, зворотня транскріптаза)
ВЕКТОРИ (плазміди, помірні бактеріофаги,
"ІНСТРУМЕНТИ"
ДЛЯ ГЕНЕТИЧНОЇ ІНЖЕНЕРІЇ
ФЕРМЕНТИ (рестриктази, лігази, зворотня транскріптаза)
ВЕКТОРИ (плазміди, помірні бактеріофаги,

СХЕМА ГЕНЕТИЧНО - ІНЖЕНЕРНОГО ЕКСПЕРИМЕНТУ
визначення локалізації потрібного гену (сиквенс, генетичне мапування)
СХЕМА ГЕНЕТИЧНО - ІНЖЕНЕРНОГО ЕКСПЕРИМЕНТУ
визначення локалізації потрібного гену (сиквенс, генетичне мапування)
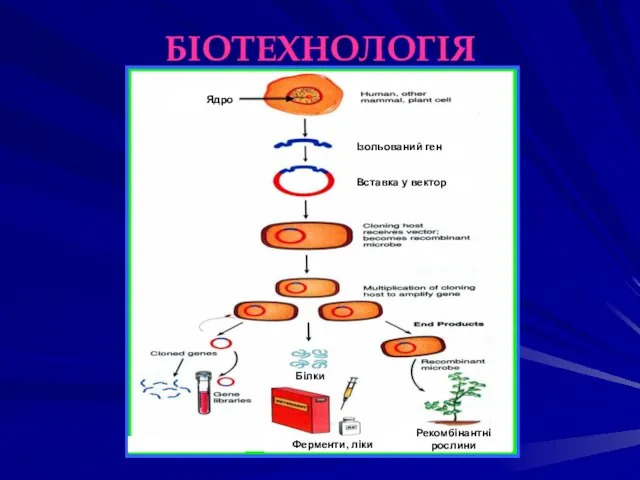
БІОТЕХНОЛОГІЯ
Ферменти, ліки
Рекомбінантні
рослини
Білки
Ядро
Ізольований ген
Вставка у вектор
БІОТЕХНОЛОГІЯ
Ферменти, ліки
Рекомбінантні
рослини
Білки
Ядро
Ізольований ген
Вставка у вектор
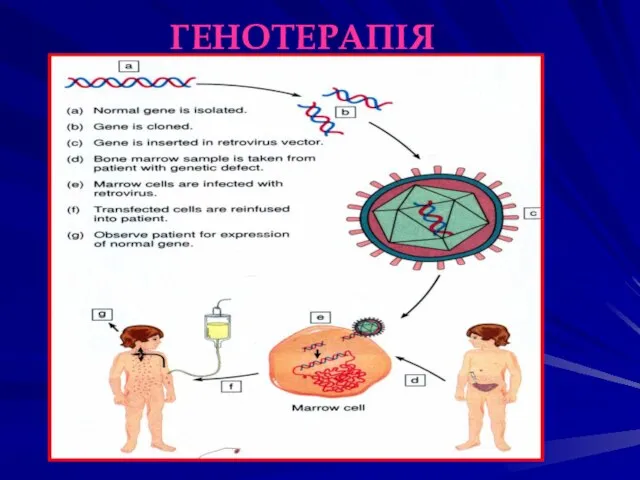
ГЕНОТЕРАПІЯ
ГЕНОТЕРАПІЯ

ГЕНЕТИКА ВІРУСІВ
Способи збільшення інформації:
дворазове зчитування одієї іРНК з інших ініціюючих кодонів
зсув
ГЕНЕТИКА ВІРУСІВ
Способи збільшення інформації:
дворазове зчитування одієї іРНК з інших ініціюючих кодонів
зсув

У вірусів можуть бути:
Модифікації (зміна складу білків капсиду, суперкапсиду під впливом
У вірусів можуть бути:
Модифікації (зміна складу білків капсиду, суперкапсиду під впливом

ВИДИ ГЕНЕТИЧНИХ РЕКОМБІНАЦІЙ У ВІРУСІВ
1. Рекомбінація: Обмін генами (міжгенна) та їх
ВИДИ ГЕНЕТИЧНИХ РЕКОМБІНАЦІЙ У ВІРУСІВ
1. Рекомбінація: Обмін генами (міжгенна) та їх

ВИДИ ГЕНЕТИЧНИХ РЕКОМБІНАЦІЙ У ВІРУСІВ
2. Множинна реактивація: вірусна інфекція викликається при
ВИДИ ГЕНЕТИЧНИХ РЕКОМБІНАЦІЙ У ВІРУСІВ
2. Множинна реактивація: вірусна інфекція викликається при

ВИДИ РЕКОМБІНАЦІЙ У ВІРУСІВ
4. Гетерозиготність: одночасній репродукції декількох віріонів, різних за
ВИДИ РЕКОМБІНАЦІЙ У ВІРУСІВ
4. Гетерозиготність: одночасній репродукції декількох віріонів, різних за

ВИДИ РЕКОМБІНАЦІЙ У ВІРУСІВ
5. Транскапсидація: частина чужерідного генетичного матерілау, заключеного всередині
ВИДИ РЕКОМБІНАЦІЙ У ВІРУСІВ
5. Транскапсидація: частина чужерідного генетичного матерілау, заключеного всередині

ВИДИ РЕКОМБІНАЦІЙ У ВІРУСІВ
6. Крос-реактивація (рятування маркера): реактивація інактивованого геному неінактивованим
ВИДИ РЕКОМБІНАЦІЙ У ВІРУСІВ
6. Крос-реактивація (рятування маркера): реактивація інактивованого геному неінактивованим
 Дикие животные
Дикие животные КИМ для подготовки учащихся 5 класса к ВПР по биологии
КИМ для подготовки учащихся 5 класса к ВПР по биологии Значение рыб в природе и жизни человека
Значение рыб в природе и жизни человека Пищеварительная система 3 (печень, поджелудочная железы)
Пищеварительная система 3 (печень, поджелудочная железы) Сцепленное наследование
Сцепленное наследование Характерные особенности растений семейств крестоцветные и пасленовые
Характерные особенности растений семейств крестоцветные и пасленовые Хвойные растения
Хвойные растения  Паразитизм Составитель Большаков С. В.
Паразитизм Составитель Большаков С. В.  Основы микробиологии и иммунологии
Основы микробиологии и иммунологии Микробиология плодов и овощей
Микробиология плодов и овощей Презентация на тему "Пищеварение и всасывание в кишечнике" - скачать презентации по Биологии
Презентация на тему "Пищеварение и всасывание в кишечнике" - скачать презентации по Биологии Статистические показатели популяции
Статистические показатели популяции Клеточная теория
Клеточная теория Типы экологических взаимодействий
Типы экологических взаимодействий  Тема « Обмен веществ. Обменные процессы в организме»
Тема « Обмен веществ. Обменные процессы в организме» Что делает человека человеком
Что делает человека человеком Однодольные растения. Лилейные
Однодольные растения. Лилейные Презентация на тему: «Генетически модифицированные объекты» Выполнил 8-а Руководитель: Аржакова Юлия Юрьевна
Презентация на тему: «Генетически модифицированные объекты» Выполнил 8-а Руководитель: Аржакова Юлия Юрьевна  Кости туловища
Кости туловища ПРОКАРИОТЫ
ПРОКАРИОТЫ Orientări în anatomie:puncte,axe,planuri anatomice. Introducere in sistemul osos Relieful osos
Orientări în anatomie:puncte,axe,planuri anatomice. Introducere in sistemul osos Relieful osos Роль человека в биосфере. Тема 58
Роль человека в биосфере. Тема 58 Тип Инфузории или Ресничные. Подцарство Простейшие. Мультимедийная презентация урока биологии в 7 классе
Тип Инфузории или Ресничные. Подцарство Простейшие. Мультимедийная презентация урока биологии в 7 классе Valgud. Valkude ehitus
Valgud. Valkude ehitus Тварини дбають про своє потомство
Тварини дбають про своє потомство Презентация на тему Дикорастущие и культурные растения
Презентация на тему Дикорастущие и культурные растения Медицинская биохимия Раздел биохимии, изучающий нарушения метаболизма, лежащие в основе различных заболеваний
Медицинская биохимия Раздел биохимии, изучающий нарушения метаболизма, лежащие в основе различных заболеваний  Защита растений в ландшафте
Защита растений в ландшафте