- Клетки преимплантационного эмбриона: потенции и пластичность

Содержание
- 2. Выбор судьбы клетками раннего эмбриона: решение в 2 этапа 1 этап: внутри или снаружи (вопрос: за
- 3. Дробление клеток раннего эмбриона: насколько клетки идентичны? PLoS One. 2016 Mar 31;11(3). The Principal Forces of
- 4. Дробление клеток раннего эмбриона: насколько клетки идентичны? Эксперименты по исследованию асимметрии ооцита и раннего эмбриона овцы
- 5. 32 бластомера и более Компактизация и поляризация клеток эмбриона: вот она, классическая схема с формированием адгезионных
- 6. Период компактизации Переход от 8 к 16 Переход от 16 к 32 Клеток мало для формирования
- 7. Асимметричная локализация в клетках: aPKC, Par3 и ezrin ezrin aPKC aPKC Par3 Louvet et al., 1996
- 8. Компактизация и поляризация клеток эмбриона: как круглые клетки становятся плоскими? Fierro-González et al., Cadherin-dependent filopodia control
- 9. Протеин-киназа Hippo (у млекопитающих называется Mst1/2); Этот каскад контролирует пролиферацию и апоптоз; Мутации в этом гене
- 10. Hippo сигналинг: преобразование позиционной информации в молекулярный каскад Gata3 – ранний маркер клеток ТЭ Oct4, Sox2
- 11. Сигнальный путь Hippo в ранних эмбрионах Выключение Hippo во внешних клетках – вторичное событие по отношению
- 12. Hippo сигналинг недостаточен, необходима «внутренняя» среда Щелевые контакты Адгезионные контакты → сигналиг через цитоплазматические тирозин-киназы Фокальные
- 13. Классические представления: В ранней бластоцисте все клетки ВКМ с самого начала различаются по положению, в результате
- 14. FGFR1 Gata6, FGFR1-4 Преимплантационный эпибласт Первичная эндодерма 2008: PrE - предшественники первичной энтодермы (гипобласта) мигрируют к
- 15. С развитием преимплантационного эмбриона происходит потеря потенции клеток
- 16. Является ли у бластоцисты человека разделение клеток на ТЭ и ВКМ окончательным ? Paepe C.D. et
- 17. Результаты микрохирургических операций: Наружные клетки, помещенные внутрь эмбриона оставались в этой позиции и начинали экспрессировать NANOG:
- 18. Гипометилирование ДНК Метилирование гистонов Декомпактизация хроматина Время, которое клетка занимает внутреннюю позицию; подверженность действию внеклеточного матрикса:
- 19. Bischoff et al., 2008 События в жизни преимплантационного эмбриона: что влияет на время их наступления? Эмбрион
- 20. Переход от тоти- к прюрипотенции у клеток ВКМ (эмбрионы мыши) Период первичной плюрипотентности 4-е 5-е клеточные
- 21. Возвращаемся к первому слайду: Выбор судьбы клетками раннего эмбриона - решение в 2 этапа: 1 этап:
- 22. Вопрос для обсуждения: за счет каких клеточных процессов клетки эмбриона становятся трофэктодермы или внутренней клеточной массы?
- 25. Скачать презентацию

Выбор судьбы клетками раннего эмбриона: решение в 2 этапа
1 этап: внутри
Выбор судьбы клетками раннего эмбриона: решение в 2 этапа
1 этап: внутри

Дробление клеток раннего эмбриона: насколько клетки идентичны?
PLoS One. 2016 Mar 31;11(3).
Дробление клеток раннего эмбриона: насколько клетки идентичны?
PLoS One. 2016 Mar 31;11(3).
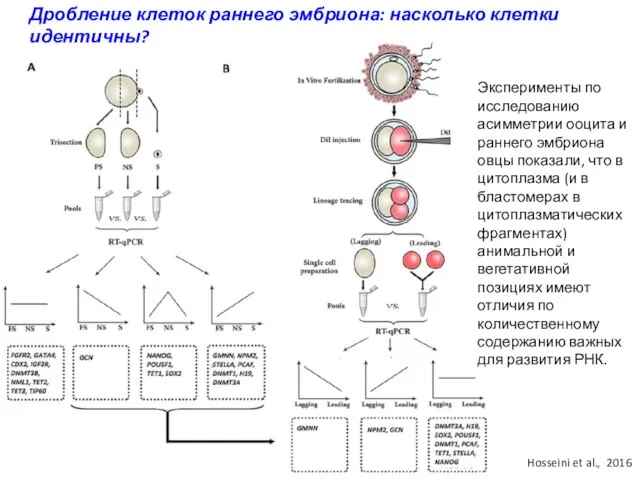
Дробление клеток раннего эмбриона: насколько клетки идентичны?
Эксперименты по исследованию асимметрии ооцита
Дробление клеток раннего эмбриона: насколько клетки идентичны?
Эксперименты по исследованию асимметрии ооцита
32 бластомера
и более
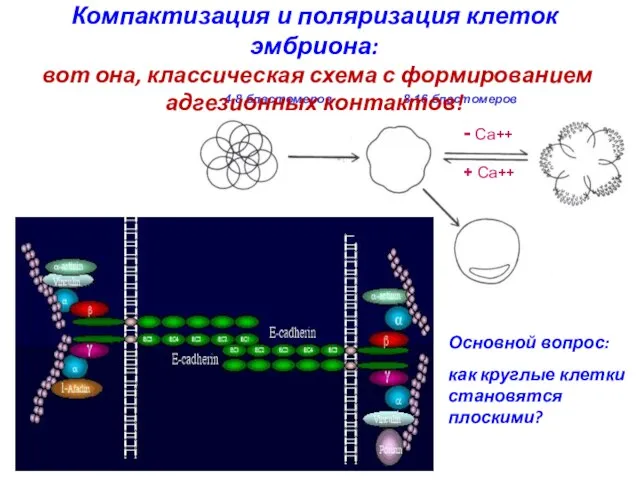
Компактизация и поляризация клеток эмбриона:
вот она,
32 бластомера
и более
Компактизация и поляризация клеток эмбриона:
вот она,
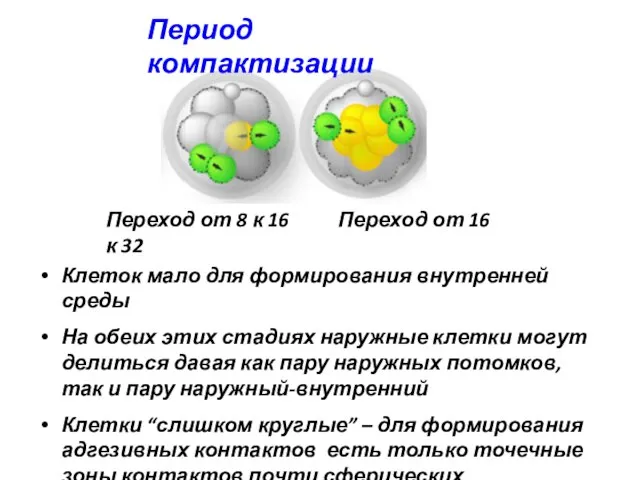
Период компактизации
Переход от 8 к 16 Переход от 16 к 32
Клеток
Период компактизации
Переход от 8 к 16 Переход от 16 к 32
Клеток
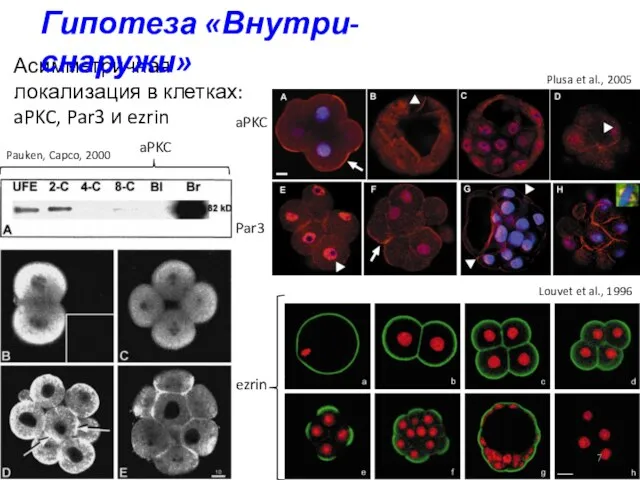
Асимметричная локализация в клетках: aPKC, Par3 и ezrin
ezrin
aPKC
aPKC
Par3
Louvet et al., 1996
Pauken,
Асимметричная локализация в клетках: aPKC, Par3 и ezrin
ezrin
aPKC
aPKC
Par3
Louvet et al., 1996
Pauken,
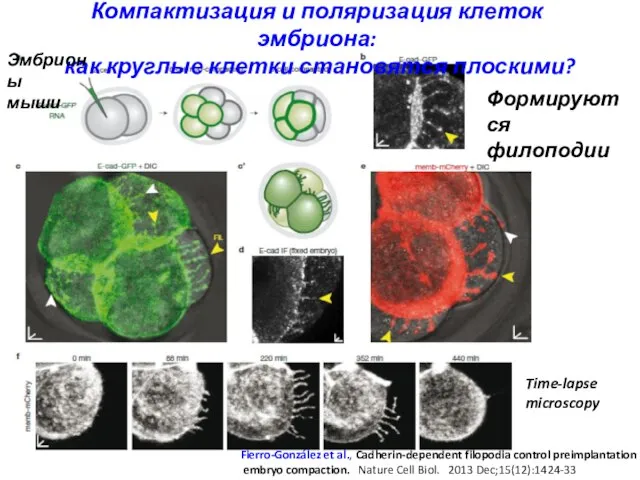
Компактизация и поляризация клеток эмбриона:
как круглые клетки становятся плоскими?
Fierro-González et
Компактизация и поляризация клеток эмбриона:
как круглые клетки становятся плоскими?
Fierro-González et
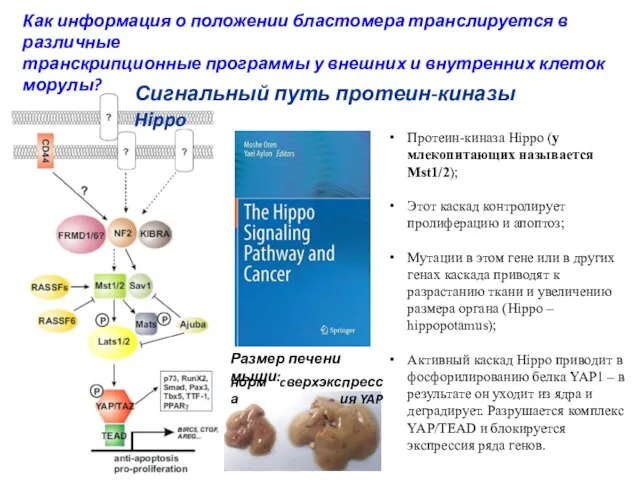
Протеин-киназа Hippo (у млекопитающих называется Mst1/2);
Этот каскад контролирует пролиферацию и апоптоз;
Протеин-киназа Hippo (у млекопитающих называется Mst1/2);
Этот каскад контролирует пролиферацию и апоптоз;
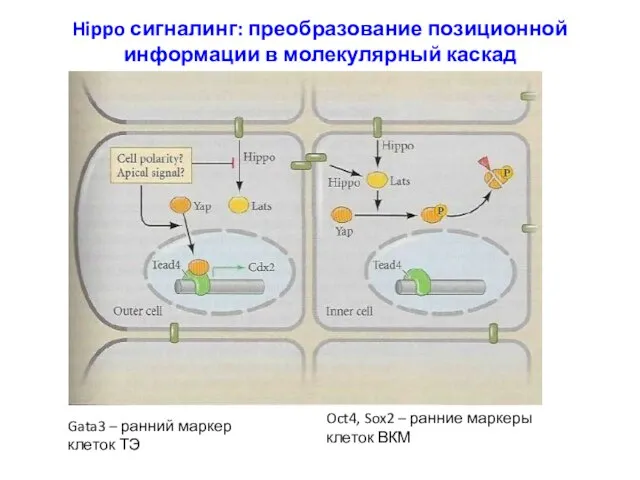
Hippo сигналинг: преобразование позиционной информации в молекулярный каскад
Gata3 – ранний маркер
Hippo сигналинг: преобразование позиционной информации в молекулярный каскад
Gata3 – ранний маркер
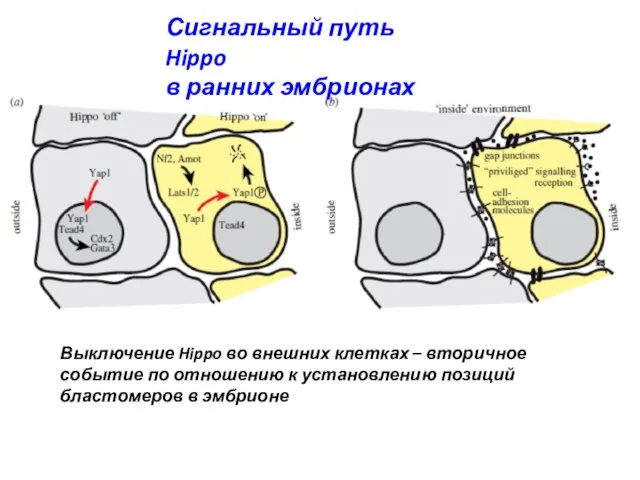
Сигнальный путь Hippo
в ранних эмбрионах
Выключение Hippo во внешних клетках
Сигнальный путь Hippo
в ранних эмбрионах
Выключение Hippo во внешних клетках
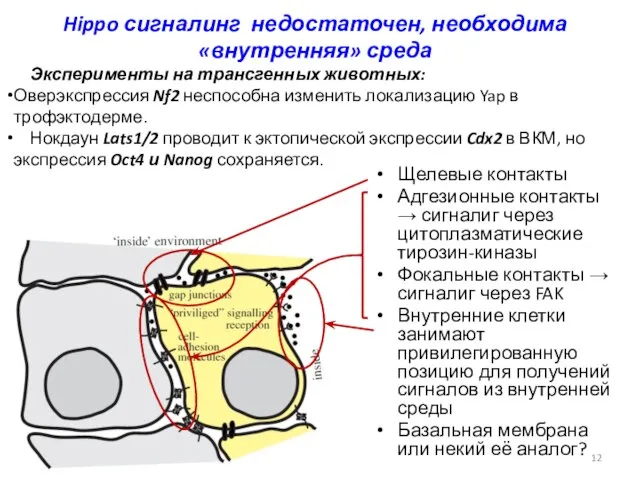
Hippo сигналинг недостаточен, необходима «внутренняя» среда
Щелевые контакты
Адгезионные контакты → сигналиг через
Hippo сигналинг недостаточен, необходима «внутренняя» среда
Щелевые контакты
Адгезионные контакты → сигналиг через

Классические представления:
В ранней бластоцисте все клетки ВКМ с самого начала
Классические представления:
В ранней бластоцисте все клетки ВКМ с самого начала
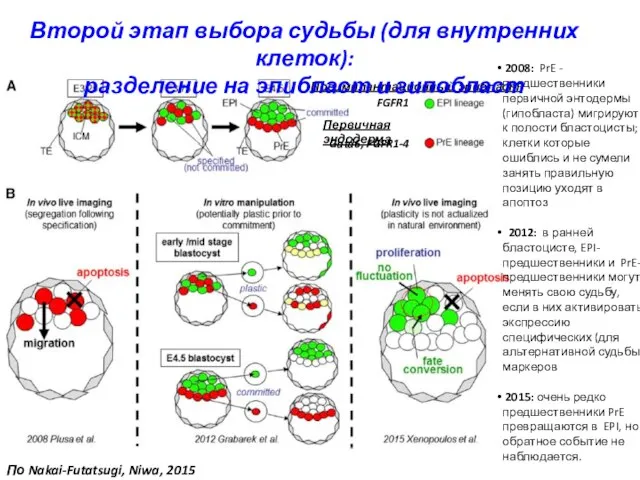
FGFR1
Gata6, FGFR1-4
Преимплантационный эпибласт
Первичная эндодерма
2008: PrE - предшественники первичной энтодермы
FGFR1
Gata6, FGFR1-4
Преимплантационный эпибласт
Первичная эндодерма
2008: PrE - предшественники первичной энтодермы
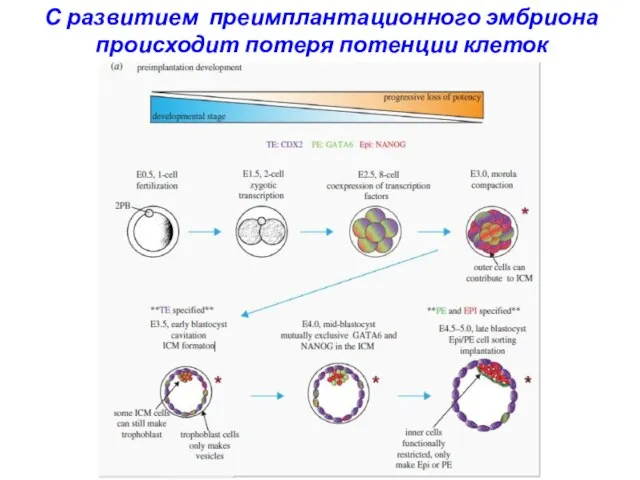
С развитием преимплантационного эмбриона происходит потеря потенции клеток
С развитием преимплантационного эмбриона происходит потеря потенции клеток
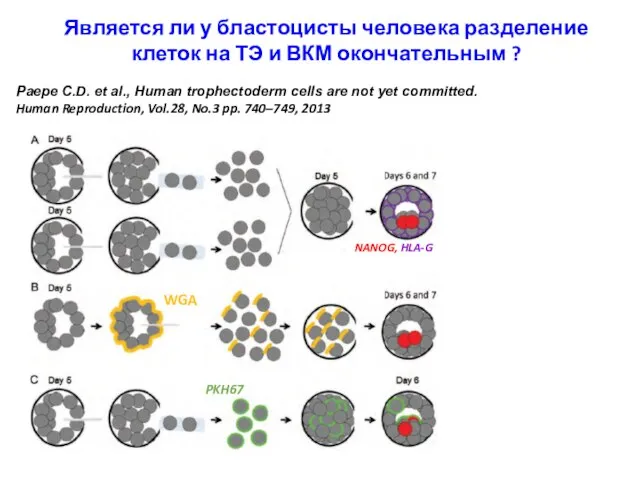
Является ли у бластоцисты человека разделение клеток на ТЭ и ВКМ
Является ли у бластоцисты человека разделение клеток на ТЭ и ВКМ
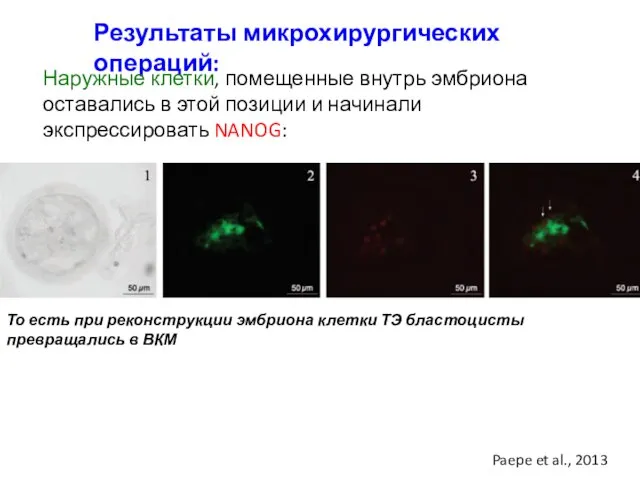
Результаты микрохирургических операций:
Наружные клетки, помещенные внутрь эмбриона оставались в этой позиции
Результаты микрохирургических операций:
Наружные клетки, помещенные внутрь эмбриона оставались в этой позиции

Гипометилирование ДНК
Метилирование гистонов
Декомпактизация хроматина
Время, которое клетка занимает внутреннюю позицию; подверженность действию
Гипометилирование ДНК
Метилирование гистонов
Декомпактизация хроматина
Время, которое клетка занимает внутреннюю позицию; подверженность действию
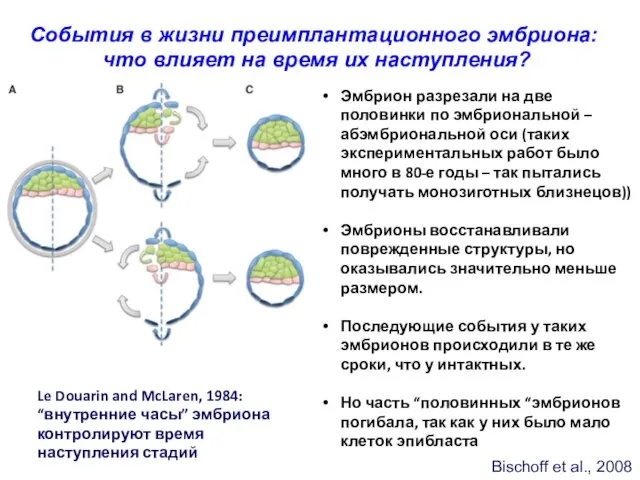
Bischoff et al., 2008
События в жизни преимплантационного эмбриона:
что влияет на
Bischoff et al., 2008
События в жизни преимплантационного эмбриона:
что влияет на
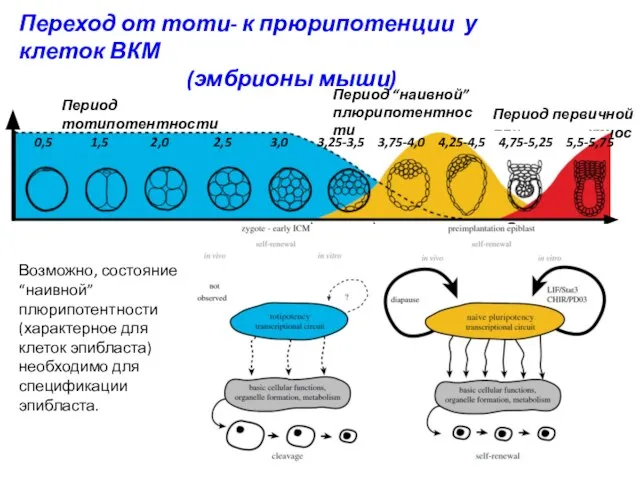
Переход от тоти- к прюрипотенции у клеток ВКМ
(эмбрионы мыши)
Период первичной
Переход от тоти- к прюрипотенции у клеток ВКМ
(эмбрионы мыши)
Период первичной

Возвращаемся к первому слайду:
Выбор судьбы клетками раннего эмбриона - решение в
Возвращаемся к первому слайду:
Выбор судьбы клетками раннего эмбриона - решение в

Вопрос для обсуждения:
за счет каких клеточных процессов клетки эмбриона становятся
Вопрос для обсуждения:
за счет каких клеточных процессов клетки эмбриона становятся

 Ядро клетки. Строение и функции
Ядро клетки. Строение и функции Пищеварительная система
Пищеварительная система Презентация на тему Пищеварительная система
Презентация на тему Пищеварительная система Біологія в ХХІ столітті. Генетика. Штучний геном
Біологія в ХХІ столітті. Генетика. Штучний геном  Кожный анализатор
Кожный анализатор Огляд відділів евкаріотичних водоростей
Огляд відділів евкаріотичних водоростей Пятнистый олень
Пятнистый олень Презентация Проблемы Мирового океана
Презентация Проблемы Мирового океана Способы размножение растений. Размножение споровых растений
Способы размножение растений. Размножение споровых растений Изучение генетической структуры популяции. Внешние различия
Изучение генетической структуры популяции. Внешние различия Декоративные цветы
Декоративные цветы Водоросли
Водоросли Морские обитатели контраст форм
Морские обитатели контраст форм Природные стимуляторы роста и защитные биопрепараты
Природные стимуляторы роста и защитные биопрепараты Витамины сырья и продуктов питания
Витамины сырья и продуктов питания Презентация на тему Возникновение жизни и историческое развитие органического мира
Презентация на тему Возникновение жизни и историческое развитие органического мира  Презентация на тему "В мире инфекций" - скачать презентации по Биологии
Презентация на тему "В мире инфекций" - скачать презентации по Биологии Садовые цветы
Садовые цветы Презентация на тему Класс Ленточные черви
Презентация на тему Класс Ленточные черви СЕМЕЙСТВО ЛИЛЕЙНЫХ: СТРОЕНИЕ ЦВЕТКА ТЮЛЬПАНА
СЕМЕЙСТВО ЛИЛЕЙНЫХ: СТРОЕНИЕ ЦВЕТКА ТЮЛЬПАНА Генетические основы селекции. Вклад Н.И. Вавилова в развитие селекции (11 класс)
Генетические основы селекции. Вклад Н.И. Вавилова в развитие селекции (11 класс) Презентация на тему Понятие о биоценозе
Презентация на тему Понятие о биоценозе  Хоперский заповедник
Хоперский заповедник English riddles. Животные
English riddles. Животные Грибы: антиоксидантное действие
Грибы: антиоксидантное действие Все мышцы тела человека
Все мышцы тела человека Высшая нервная деятельность. Высшие мозговые функции
Высшая нервная деятельность. Высшие мозговые функции Физиология Дыхательной Системы Лаборатория полисистемных исследований
Физиология Дыхательной Системы Лаборатория полисистемных исследований