- Клеточные аспекты дифференцировки в корне

Содержание
- 2. Строение апикальной меристемы корня (в сравнении со стеблем). Терминальное/субтерминальное положение. По: Raven
- 3. Выделение зон связано с их производными. Теория гистогенов. Промеристема (покоящийся центр), каллиптроген, периблема, плерома.
- 4. “Закрытый” и “открытый” типы апикальной меристемы корня. По: Raven
- 5. Модельный процесс - полярный транспорт ауксина и дифференциация тканей корня
- 6. Градиент ауксина с максимумом в каллиптрогене достигается полярным распределением переносчиков. КАНАЛИЗАЦИЯ ПТА: изначально незначительное доминирование базипетального
- 7. (b) визуализация при помощи DR5::GUS - максимум ауксина в калиптрогене (c) и значительному изменению структуры, однако
- 8. Экспрессия индуцируемого ауксином промотера DR5 A-F (А) Максимальная экспрессия DR5rev::PEH A в каллиптрогене и верхнем ярусе
- 9. Роль покоящегося центра в апикальной меристеме корня.
- 10. Root Development. Ben Scheres, Philip Benfey, and Liam Dolan, pages 1–18.The Arabidopsis Book, eds. C.R. Somerville
- 11. Регуляция АМК. WOX5 подавляет в ПЦ CYCD3;3. В инициалях плеромы, CEI, LRC/Epi и ROW1 ограничивают экспрессию
- 12. Так же, как АМП, АМ корня устроена по принципу ниши стволовых клеток. Индуктор - клетки покоящегося
- 13. Заложение боковых корней. По: Raven
- 14. Модельный процесс - образование бокового корня Вход в новый митотический цикл, или выход из клеточного цикла
- 15. Градиент ауксина в ходе развития зачатка бокового корня (.(B) DR5::GUS в калиптрогене сформированного бокового корня. Обработка
- 16. По: Raven Дифференциация клеток эндодермы и определение числа слоев коры
- 17. Дифференциация клеток эндодермы и определение числа слоев коры По: Rost
- 18. Клетки эндодермы и внутреннего слоя коровой паренхимы образуются в результате периклинального деления одной из инициалей периблемы.Т.о.
- 19. Ген SCARECROW - маркер клеток покоящегося центра и эндодермы a) in situ гибридизацияSCARECROW. b) конструкция SCR:
- 20. Ген SHORTROOT (SHR) необходим для дифференциации клеток эндодермы. Транскрипция в стеле, белок - в будущих клетках
- 21. Предполагаемый механизм регуляции дифференциаци клеток эндодермы генами SHORTROOT (SHR) и SCARECROW (SCR): SHR, проходя по плазмодесмам
- 22. Дополнительные слои эндодермы в корнях трансгенных растений, экспрессирующих ген SHR под промотером SCR.
- 23. У мутанта scarecrow в коре не достает одного слоя клеток, а имеющийся слой сочетает в себе
- 24. Модельный процесс - образование корневых волосков (трихобластов)
- 25. (A) Ризодерма состоит из 2 типов клеток: трихобластов и атрихобластов. (B) трихобласты всегда граничат с двумя
- 26. Модель сигнальной регуляции программы развития корневого волоска Информационные молекулы из клеток коровой паренхимы активируют комплексы ингибирующие
- 27. Латеральные меристемы: прокамбий, камбий. Их производные.
- 29. Дифференцировка прокамбия в верхушке побега происходит под влиянием ауксина - гормона, синтезирующегося в растущих листовых зачатках.
- 30. Гипотеза канализации ауксинового потока
- 31. Ген из класса HD-ZIPIII (ATHB8) - самый ранний маркер клеток прокамбия.
- 32. Клетки прокамбия характеризуются экспрессией генов KNOX, практически всех генов “адаксиальности”, маркеров “покоящегося центра” корня, PIN.
- 33. Сходства и различия прокамбия и камбия.
- 34. Срез стебля травянистого двудольного растения первичного строения. Проводящие пучки = листовые следы - тяжи соединяющие стебель
- 35. Камбий. Происхождение: прокамбий, межпучковая паренхима, перицикл в корне. Местоположение: побеговая и корневая системы вторичного строения. Функция:
- 36. Образовние камбия - еще одной “бессмертной” ткани и переход от пучкового строения стебля к кольцевому
- 38. Однородность/неоднородность в пределах камбиальной зоны: камбиальные инициали (“стволовые клетки”, их производные – будущие инициали ксилемы и
- 39. Делиться и дифференцироваться – основное свойство камбия. «Правило Санио» - каждая камбиальная инициаль делится четыре раза.
- 40. Nature Reviews Molecular Cell Biology 2; 33-39 (2001); Laurie G. Smith PLANT CELL DIVISION: BUILDING WALLS
- 41. Phragmoplastin dynamics: multiple forms, microtubule association and their roles in cell plate formation in plants Zonglie
- 42. Особенности цитокинеза клеток камбия. Камбий – сезонно функционирующая меристема. Ультраструктурные характеристики покоящегося и активного камбия.
- 43. Камбий – еще одна ниша стволовых клеток в теле растения?
- 44. Молекулярные методы для уточнения структуры камбиальной зоны.
- 47. Камбиальные инициали со стороны флоэмы: гомолог CLV1, (PttCLV1) ANT, PttKAN1 и KNOX. В камбиальной зоне со
- 48. АМП, АМК, камбий устроены по принципу ниши стволовых клеток. Их деятельность поддерживается благодаря CLE-WOX взаимодействиям. WOX5,
- 49. Пролиферацию камбия контролирует несколько комплиментарных механизмов: (1) Модуль CLE-PXY-WOX. PHLOEM INTERCALATED WITH XYLEM (PXY, a leucine-rich
- 50. Транскрипционные факторы - “мегарегуляторы” “мегапрограмм” развития KNOX CLE (CLAVATA-подобные) WOX (WUS-подобные) HD-ZIP III KANADI и YABBY
- 51. Экспрессия ТФ нередко находится под контролем фитогормонов ФИТОГОРМОНЫ, органические вещества небольшого молекулярного веса, образуемые в малых
- 52. Ауксин синтезируется в верхушке побега и передвигается вдоль стебля сверху вниз по паренхимным клеткам со скоростью
- 53. Цитокинины синтезируются главным образом в кончиках корней (но также и L1 АМП) и перемещаются оттуда во
- 54. Mitogenic signals are required for proper progression of the cell cycle. A lack of these signals
- 55. Формирование корней на побегах Формирование корней в каллусе Формирование каллуса Развитие придаточных почек Развитие пазушных почек
- 56. Molecular mechanisms of cytokinin action in different Arabidopsis plant organs. (a) Cartoon of an adult Arabidopsis
- 57. Корреляции в росте органов и частей побега. Взаимосвязь в росте листа и междоузлия Взаимосвязь в развитии
- 58. Соотношение ауксина и цитокннина определяет архитектуру растений: ауксин - апикальное доминирование, цитокинин - развитие пазушных почек
- 59. Гиббереллины - гормоны роста. биосинтез - молодые листья, кончик корня, части цветка, формирующиеся и прорастающие семена)
- 60. гиббереллины способны стимулировать превращение генеративной меристемы в цветковую. Гиббереллин-зависимый путь инициации цветения становится основным в условиях
- 61. Программированная клеточная смерть. Этот термин описывает любой процесс, запускаемый и контролируемый самим растением, в ходе которого
- 62. Основные морфотипы программированной клеточной смерти гибели у растений 1. Гиперчувствительная реакция (HR) или патогенез. Относительно быстро
- 63. Различия: у растений главная роль в PCD вакуоли и пластид, отсутствие специального механизма поглощения останков, иные
- 64. Вакуоль. Мембрана - тонопласт. До 90% объема клетки. Структурная простота и полифункциональность. Обновление клеточных органелл (кислые
- 65. Программированная клеточная смерть. Автофагия. самоперевариванивание посредством автофагосом с последующей секрецией их содержимого в центральную вакуоль (атофагия)
- 66. Программированная клеточная смерть. Автофагия, остановленная на полпути. Интактная плазмалемма. Ситовидные элементы.
- 67. Программированная клеточная смерть. Трахеальные элементы. Формирование вторичной клеточной стенки и последующий гидролиз протопласта после разрыва тонопласта
- 68. Программированная клеточная смерть. Трахеальные элементы.
- 69. Программированная клеточная смерть. Лизигения. лизигения – гидролиз протопласта и клеточной стенки после разрыва тонопласта и освобождения
- 70. Старение. Хромопласты. Основные пигменты каротиноиды, ксантофиллы. Окраска цветков, плодов, осенних листьев. Многочисленные масляные тельца. упорядоченная разборка
- 71. БРАССИНОСТЕРОИДЫ, группа природных регуляторов роста растений, производные ненасыщенных оксистероидов с лактонной группой в кольце В. Ускоряют
- 72. В прокамбиальных клетках скоординированные сигналы ауксина и цитокинина активируют экспрессию генов, поддерживающих их меристематичность. Ауксиновый путь
- 73. in vitro трансдифференцировка клеток мезофилла (MC) в элементы ксилемы. Стадия 1. Утрата способности к фотосинтезу и
- 74. АБК в листьях, а также в корневом чехлике. Транспорт на короткие расстояния происходит путем диффузии, на
- 75. Стрессовые воздействия (ранения, водный дефицит, низкая температура), а также высокие концентрации ауксинов и, иногда, цитокининов резко
- 76. Механизм действия фитогормонов в основных чертах сходен с механизмом действия гормонов животных. Чувствительные клетки воспринимают гормон
- 78. Скачать презентацию
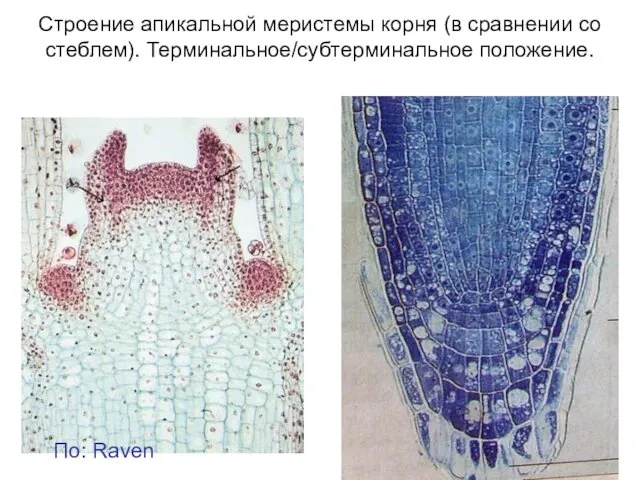
Строение апикальной меристемы корня (в сравнении со стеблем). Терминальное/субтерминальное положение.
По: Raven
Строение апикальной меристемы корня (в сравнении со стеблем). Терминальное/субтерминальное положение.
По: Raven

Выделение зон связано с их производными. Теория гистогенов. Промеристема (покоящийся центр),
Выделение зон связано с их производными. Теория гистогенов. Промеристема (покоящийся центр),

“Закрытый” и “открытый” типы апикальной меристемы корня.
По: Raven
“Закрытый” и “открытый” типы апикальной меристемы корня.
По: Raven
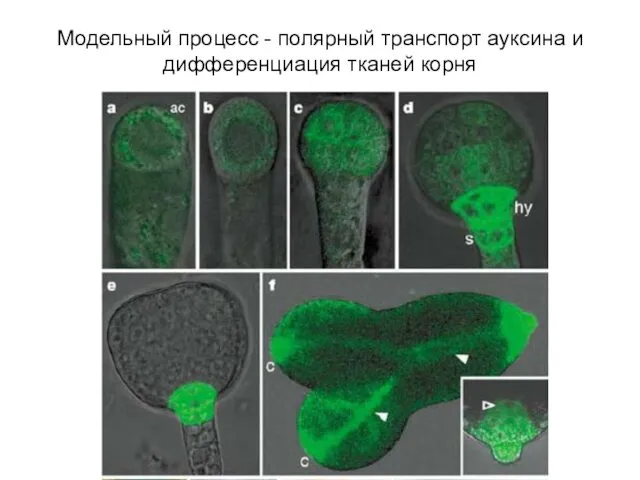
Модельный процесс - полярный транспорт ауксина и дифференциация тканей корня
Модельный процесс - полярный транспорт ауксина и дифференциация тканей корня
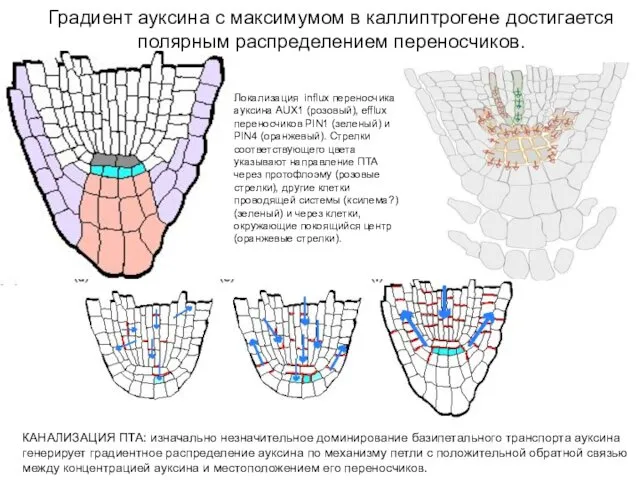
Градиент ауксина с максимумом в каллиптрогене достигается полярным распределением переносчиков.
КАНАЛИЗАЦИЯ
Градиент ауксина с максимумом в каллиптрогене достигается полярным распределением переносчиков.
КАНАЛИЗАЦИЯ
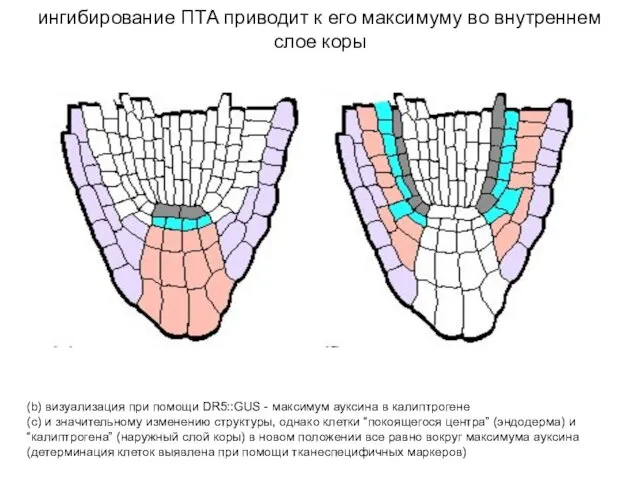
(b) визуализация при помощи DR5::GUS - максимум ауксина в калиптрогене
(c) и
(b) визуализация при помощи DR5::GUS - максимум ауксина в калиптрогене
(c) и

Экспрессия индуцируемого ауксином промотера DR5 A-F
(А) Максимальная экспрессия DR5rev::PEH A
Экспрессия индуцируемого ауксином промотера DR5 A-F
(А) Максимальная экспрессия DR5rev::PEH A
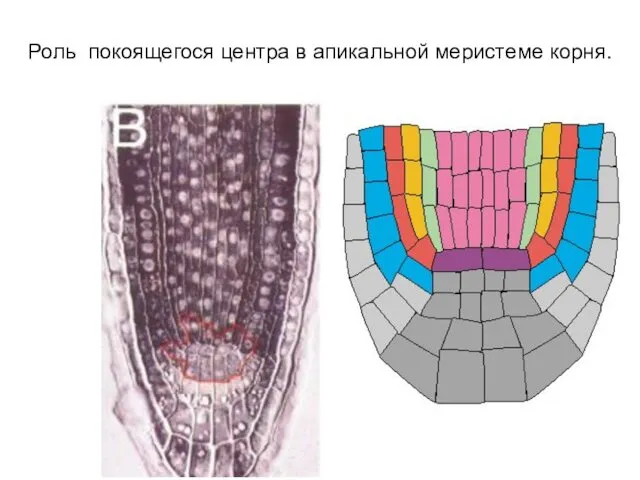
Роль покоящегося центра в апикальной меристеме корня.
Роль покоящегося центра в апикальной меристеме корня.
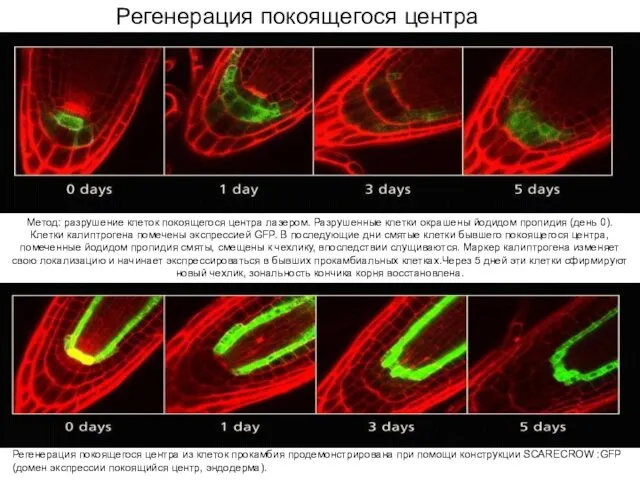
Root Development. Ben Scheres, Philip Benfey, and Liam Dolan, pages 1–18.The Arabidopsis Book, eds.
Root Development. Ben Scheres, Philip Benfey, and Liam Dolan, pages 1–18.The Arabidopsis Book, eds.
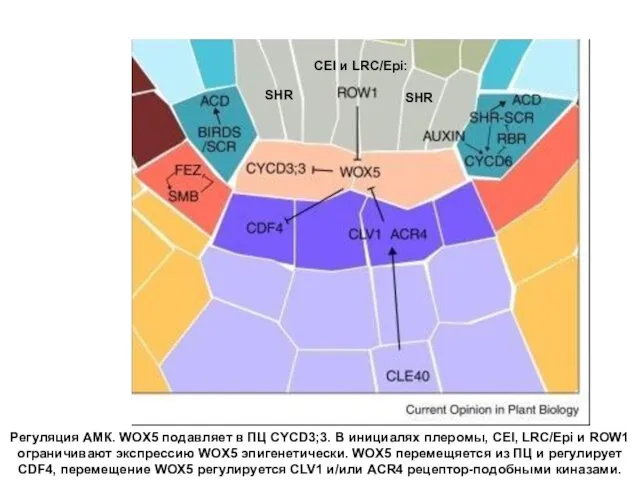
Регуляция АМК. WOX5 подавляет в ПЦ CYCD3;3. В инициалях плеромы, CEI,
Регуляция АМК. WOX5 подавляет в ПЦ CYCD3;3. В инициалях плеромы, CEI,
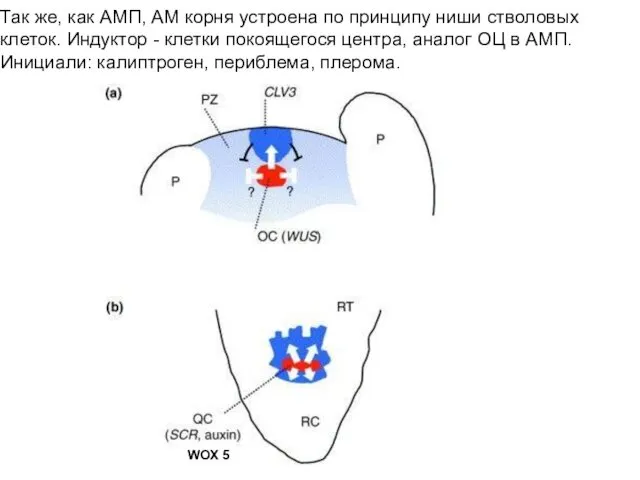
Так же, как АМП, АМ корня устроена по принципу ниши стволовых
Так же, как АМП, АМ корня устроена по принципу ниши стволовых
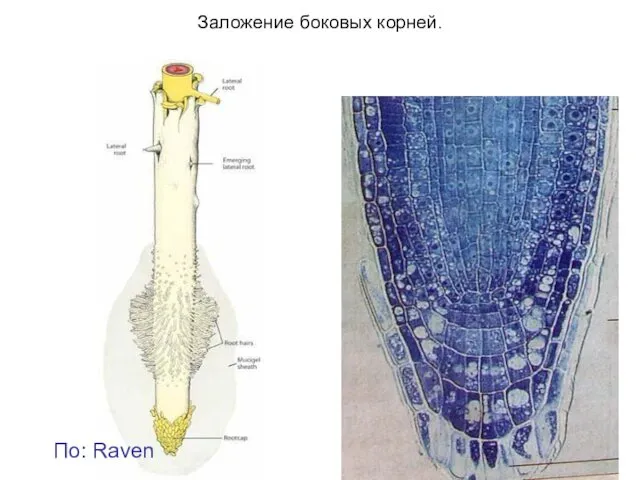
Заложение боковых корней.
По: Raven
Заложение боковых корней.
По: Raven
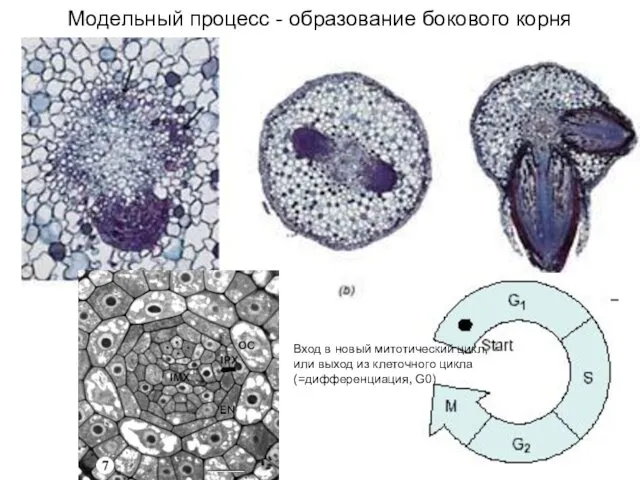
Модельный процесс - образование бокового корня
Вход в новый митотический цикл,
или
Модельный процесс - образование бокового корня
Вход в новый митотический цикл,
или
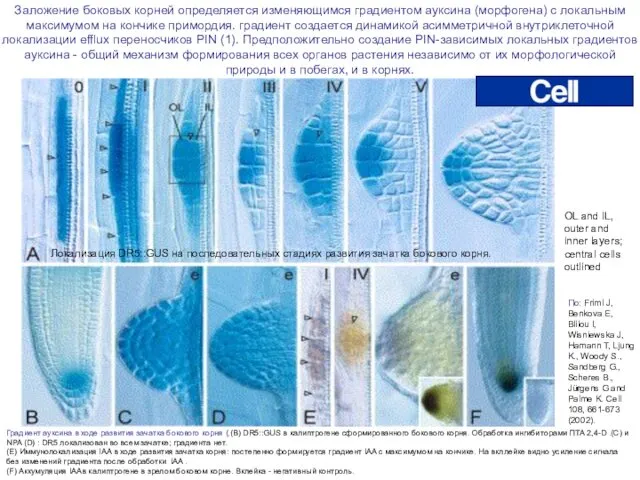
Градиент ауксина в ходе развития зачатка бокового корня (.(B) DR5::GUS в
Градиент ауксина в ходе развития зачатка бокового корня (.(B) DR5::GUS в
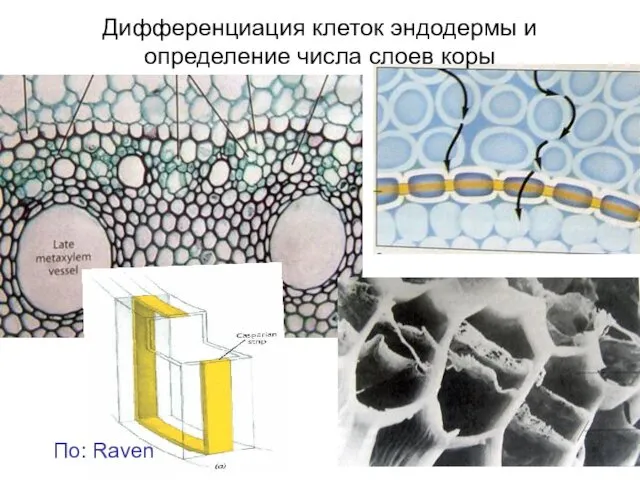
По: Raven
Дифференциация клеток эндодермы и определение числа слоев коры
По: Raven
Дифференциация клеток эндодермы и определение числа слоев коры

Дифференциация клеток эндодермы и определение числа слоев коры
По: Rost
Дифференциация клеток эндодермы и определение числа слоев коры
По: Rost
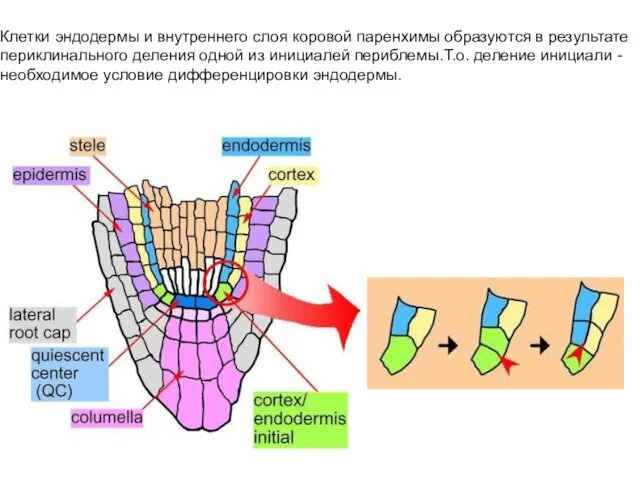
Клетки эндодермы и внутреннего слоя коровой паренхимы образуются в результате периклинального
Клетки эндодермы и внутреннего слоя коровой паренхимы образуются в результате периклинального
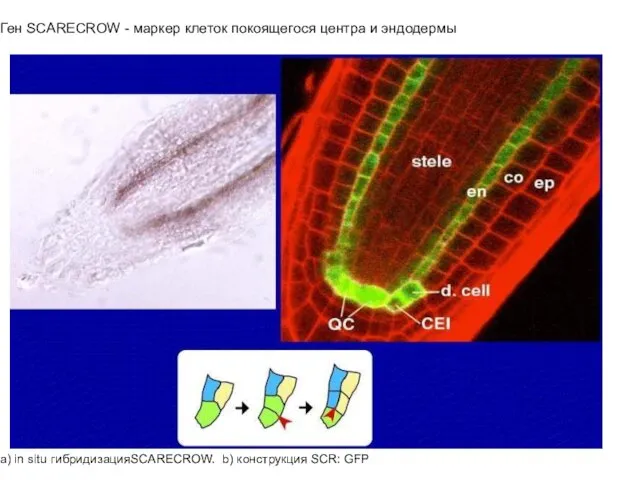
Ген SCARECROW - маркер клеток покоящегося центра и эндодермы
a) in situ
Ген SCARECROW - маркер клеток покоящегося центра и эндодермы
a) in situ
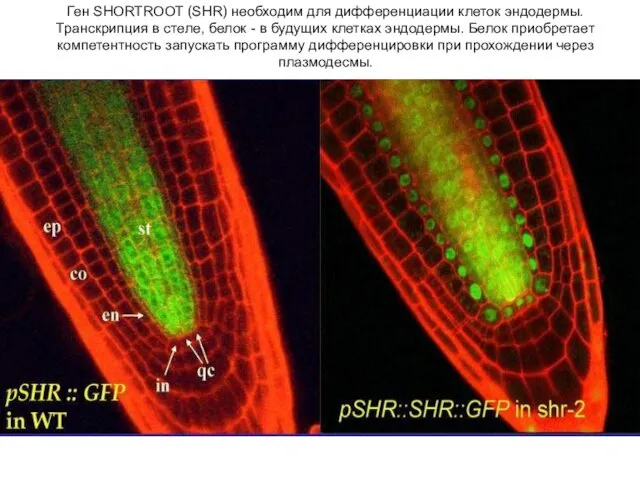
Ген SHORTROOT (SHR) необходим для дифференциации клеток эндодермы.
Транскрипция в стеле,
Ген SHORTROOT (SHR) необходим для дифференциации клеток эндодермы.
Транскрипция в стеле,
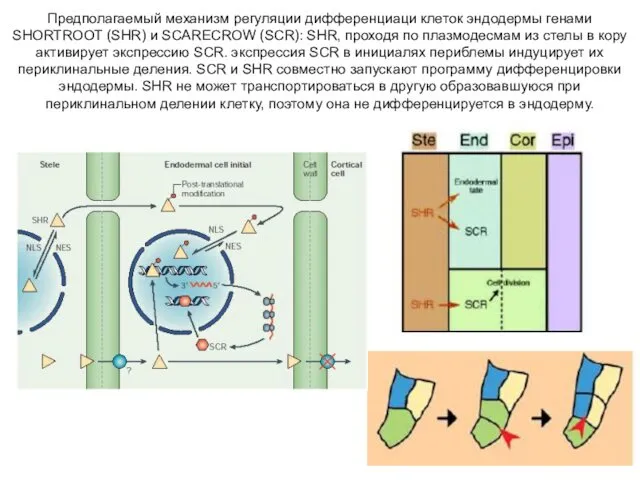
Предполагаемый механизм регуляции дифференциаци клеток эндодермы генами SHORTROOT (SHR) и SCARECROW
Предполагаемый механизм регуляции дифференциаци клеток эндодермы генами SHORTROOT (SHR) и SCARECROW
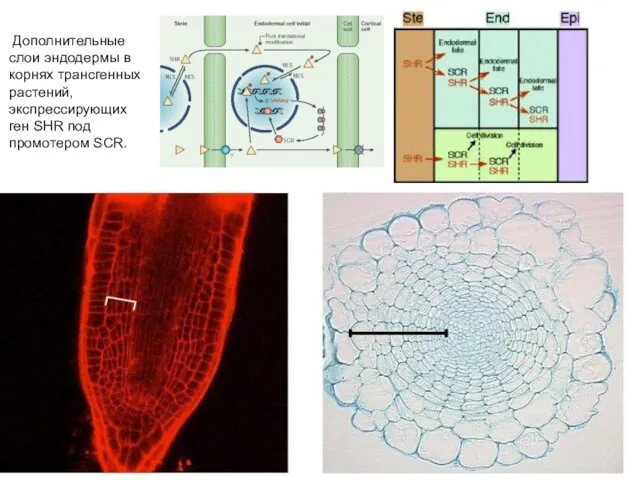
Дополнительные слои эндодермы в корнях трансгенных растений, экспрессирующих ген SHR
Дополнительные слои эндодермы в корнях трансгенных растений, экспрессирующих ген SHR
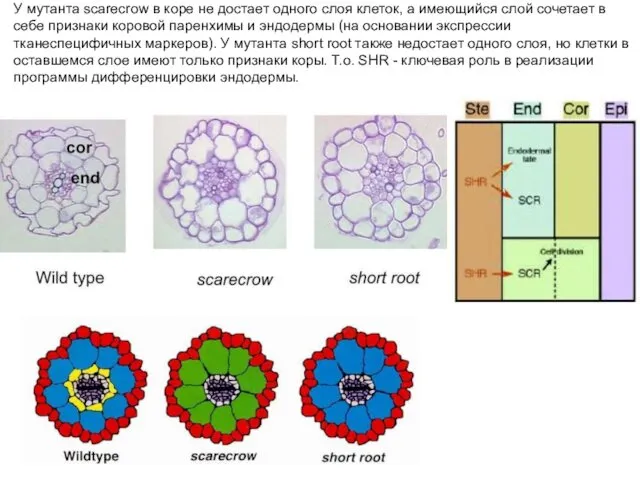
У мутанта scarecrow в коре не достает одного слоя клеток, а
У мутанта scarecrow в коре не достает одного слоя клеток, а
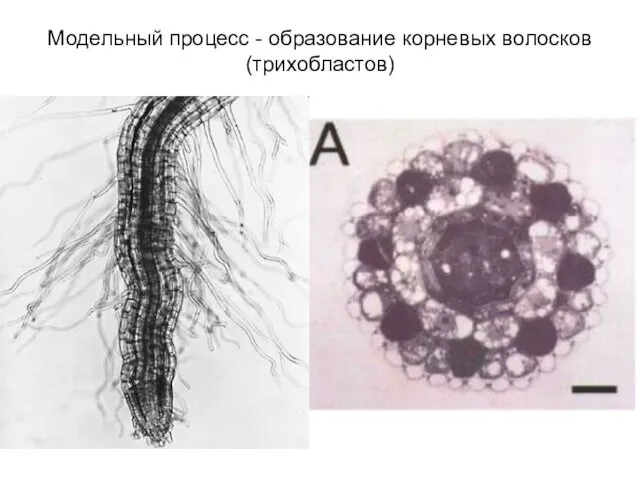
Модельный процесс - образование корневых волосков (трихобластов)
Модельный процесс - образование корневых волосков (трихобластов)
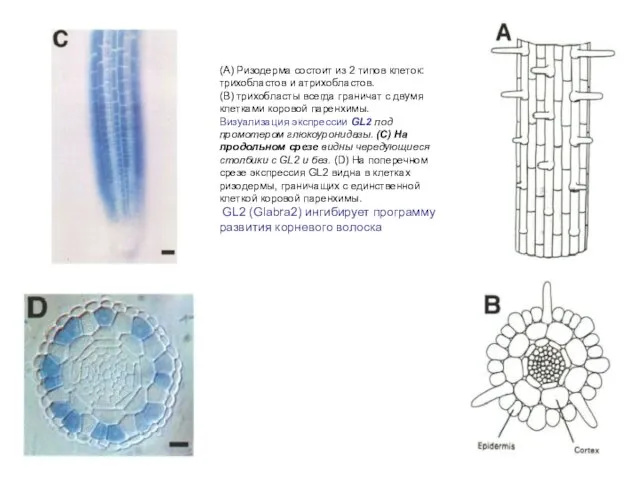
(A) Ризодерма состоит из 2 типов клеток: трихобластов и атрихобластов.
(B) трихобласты
(A) Ризодерма состоит из 2 типов клеток: трихобластов и атрихобластов.
(B) трихобласты
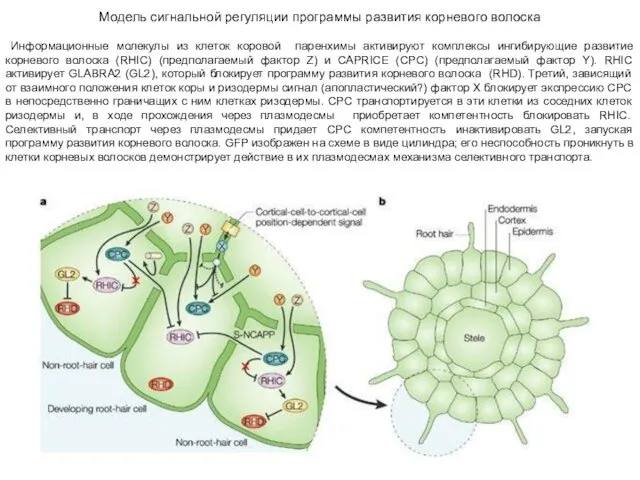
Модель сигнальной регуляции программы развития корневого волоска
Информационные молекулы из
Модель сигнальной регуляции программы развития корневого волоска
Информационные молекулы из

Латеральные меристемы: прокамбий, камбий.
Их производные.
Латеральные меристемы: прокамбий, камбий.
Их производные.

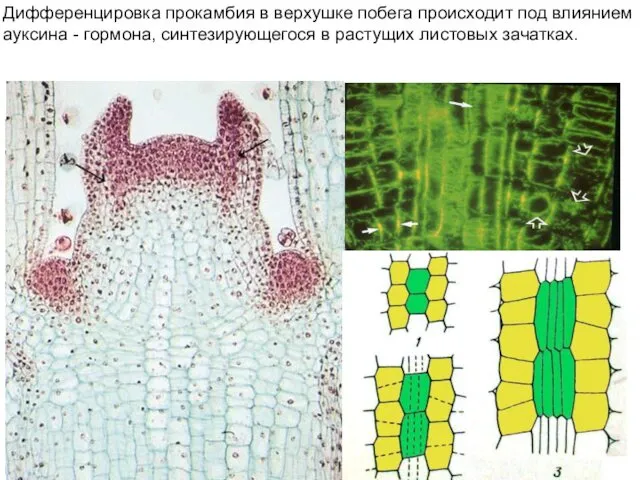
Дифференцировка прокамбия в верхушке побега происходит под влиянием ауксина - гормона,
Дифференцировка прокамбия в верхушке побега происходит под влиянием ауксина - гормона,
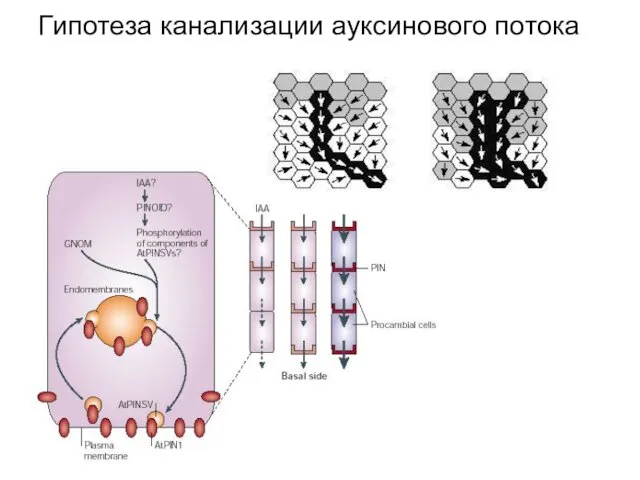
Гипотеза канализации ауксинового потока
Гипотеза канализации ауксинового потока
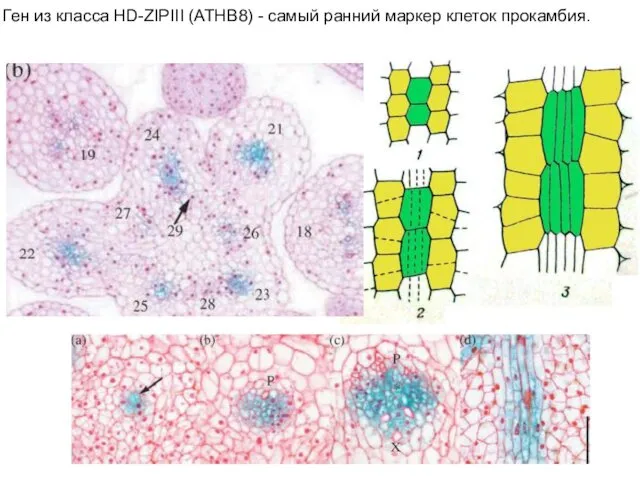
Ген из класса HD-ZIPIII (ATHB8) - самый ранний маркер клеток прокамбия.
Ген из класса HD-ZIPIII (ATHB8) - самый ранний маркер клеток прокамбия.
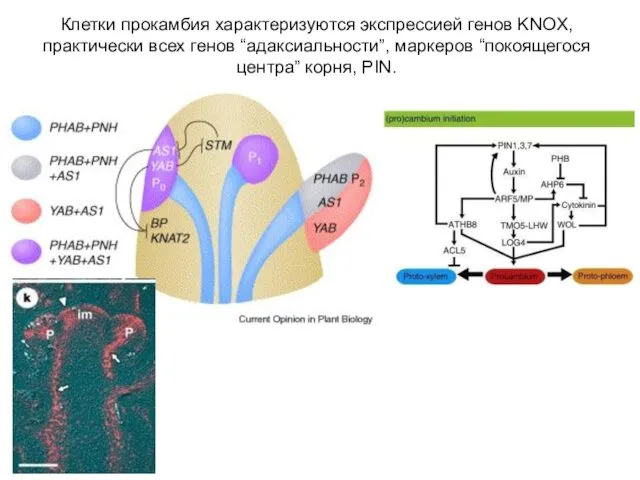
Клетки прокамбия характеризуются экспрессией генов KNOX, практически всех генов “адаксиальности”, маркеров
Клетки прокамбия характеризуются экспрессией генов KNOX, практически всех генов “адаксиальности”, маркеров

Сходства и различия прокамбия и камбия.
Сходства и различия прокамбия и камбия.
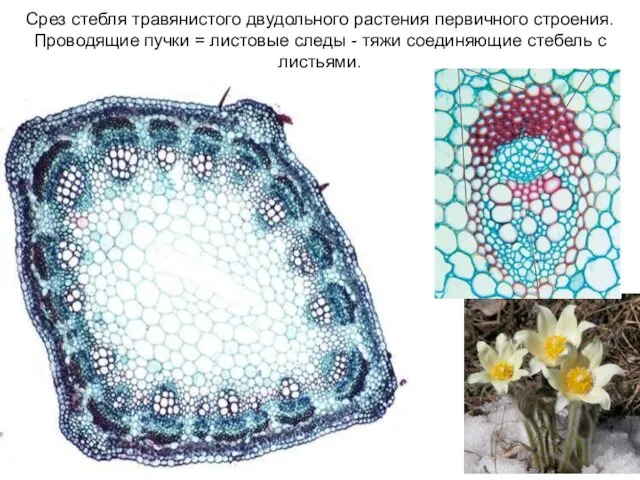
Срез стебля травянистого двудольного растения первичного строения. Проводящие пучки = листовые
Срез стебля травянистого двудольного растения первичного строения. Проводящие пучки = листовые
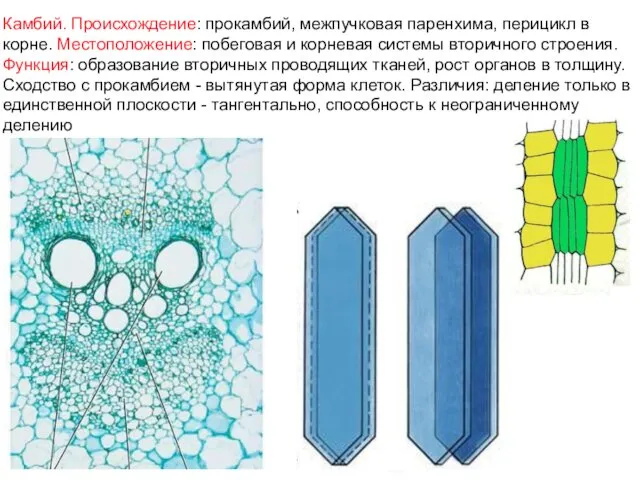
Камбий. Происхождение: прокамбий, межпучковая паренхима, перицикл в корне. Местоположение: побеговая и
Камбий. Происхождение: прокамбий, межпучковая паренхима, перицикл в корне. Местоположение: побеговая и
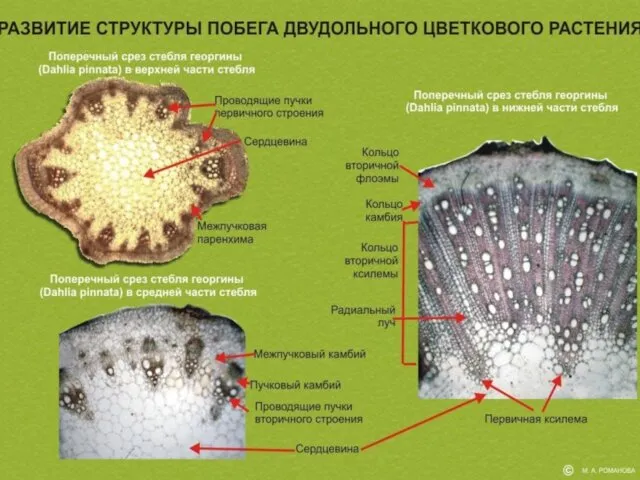
Образовние камбия - еще одной “бессмертной” ткани и переход от пучкового
Образовние камбия - еще одной “бессмертной” ткани и переход от пучкового

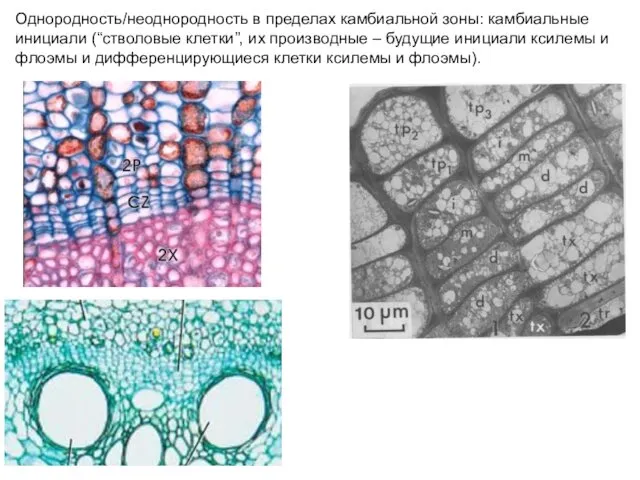
Однородность/неоднородность в пределах камбиальной зоны: камбиальные инициали (“стволовые клетки”, их производные
Однородность/неоднородность в пределах камбиальной зоны: камбиальные инициали (“стволовые клетки”, их производные
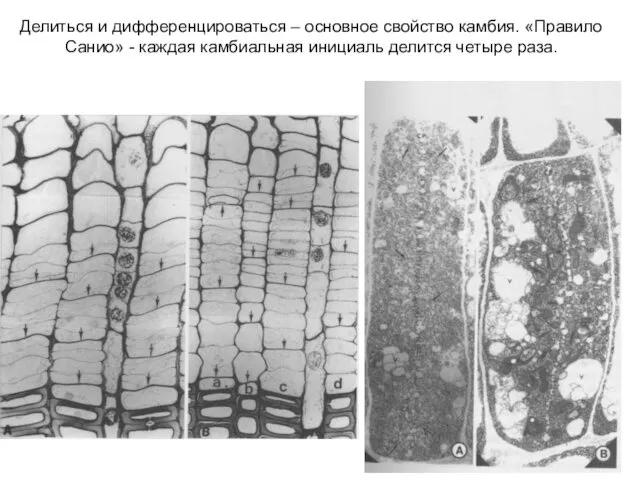
Делиться и дифференцироваться – основное свойство камбия. «Правило Санио» - каждая
Делиться и дифференцироваться – основное свойство камбия. «Правило Санио» - каждая

Nature Reviews Molecular Cell Biology 2; 33-39 (2001);
Laurie G. Smith PLANT CELL
Nature Reviews Molecular Cell Biology 2; 33-39 (2001); Laurie G. Smith PLANT CELL
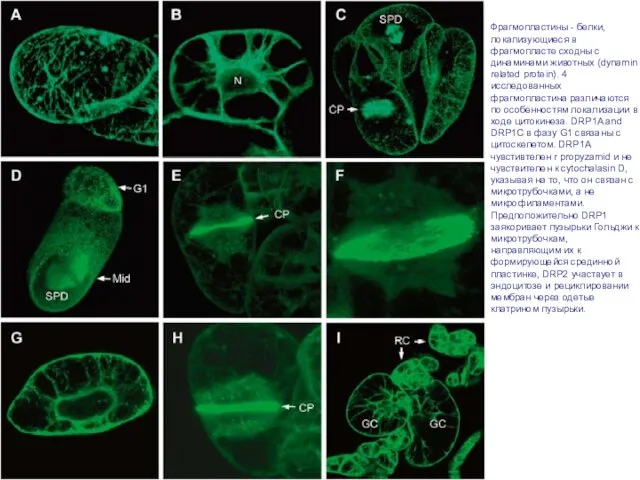
Phragmoplastin dynamics: multiple forms, microtubule association and their roles in cell
Phragmoplastin dynamics: multiple forms, microtubule association and their roles in cell
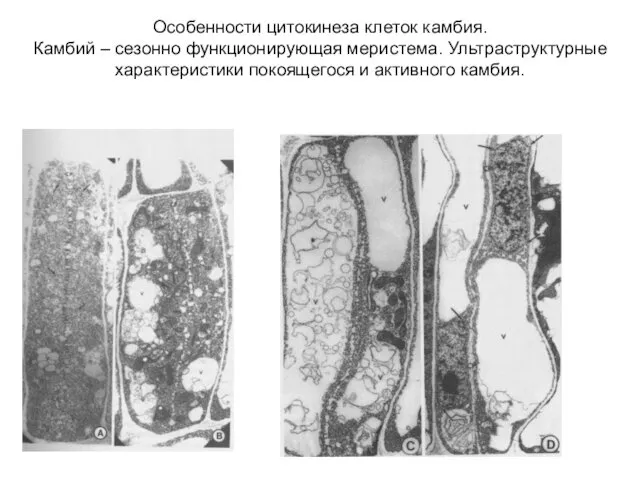
Особенности цитокинеза клеток камбия.
Камбий – сезонно функционирующая меристема. Ультраструктурные характеристики
Особенности цитокинеза клеток камбия.
Камбий – сезонно функционирующая меристема. Ультраструктурные характеристики

Камбий – еще одна ниша стволовых клеток в теле растения?
Камбий – еще одна ниша стволовых клеток в теле растения?
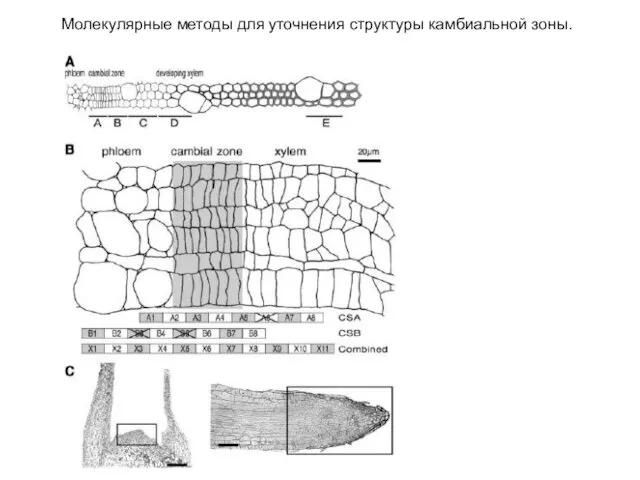
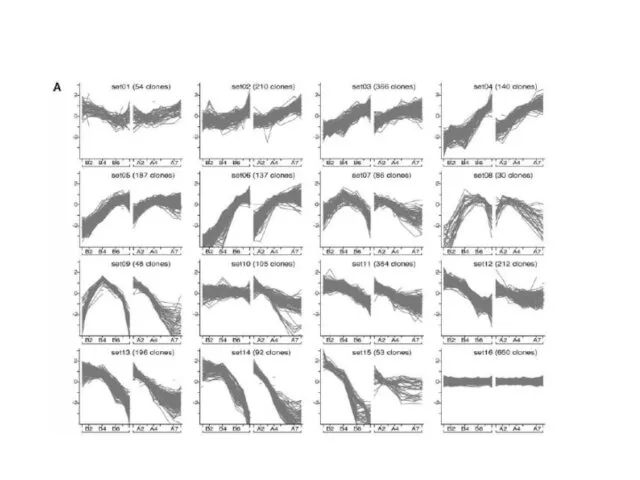
Молекулярные методы для уточнения структуры камбиальной зоны.
Молекулярные методы для уточнения структуры камбиальной зоны.

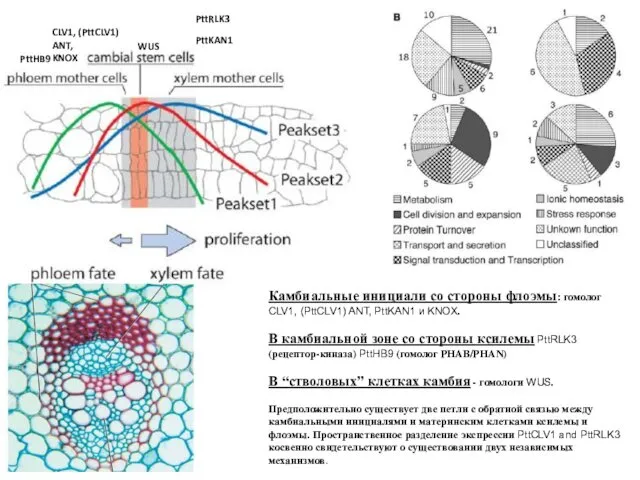
Камбиальные инициали со стороны флоэмы: гомолог CLV1, (PttCLV1) ANT, PttKAN1 и
Камбиальные инициали со стороны флоэмы: гомолог CLV1, (PttCLV1) ANT, PttKAN1 и
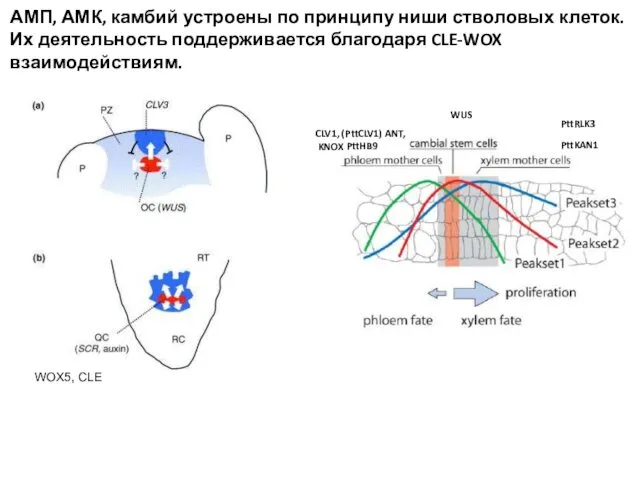
АМП, АМК, камбий устроены по принципу ниши стволовых клеток. Их деятельность
АМП, АМК, камбий устроены по принципу ниши стволовых клеток. Их деятельность
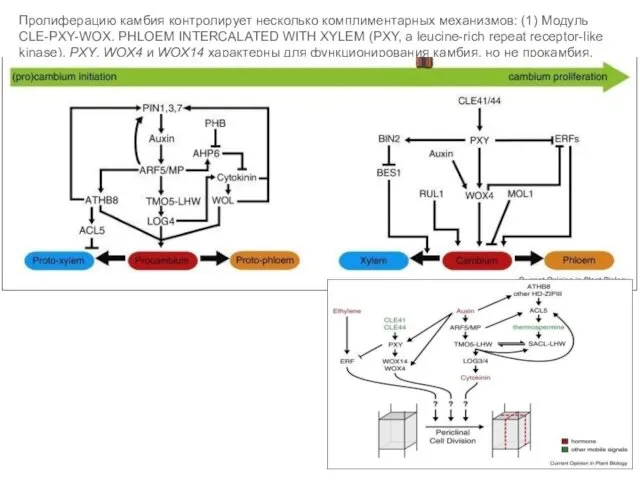
Пролиферацию камбия контролирует несколько комплиментарных механизмов: (1) Модуль CLE-PXY-WOX. PHLOEM INTERCALATED
Пролиферацию камбия контролирует несколько комплиментарных механизмов: (1) Модуль CLE-PXY-WOX. PHLOEM INTERCALATED

Транскрипционные факторы - “мегарегуляторы” “мегапрограмм” развития
KNOX
CLE (CLAVATA-подобные)
WOX (WUS-подобные)
HD-ZIP III
KANADI и YABBY
“Мегарегуляторы”
Транскрипционные факторы - “мегарегуляторы” “мегапрограмм” развития
KNOX
CLE (CLAVATA-подобные)
WOX (WUS-подобные)
HD-ZIP III
KANADI и YABBY
“Мегарегуляторы”

Экспрессия ТФ нередко находится под контролем фитогормонов
ФИТОГОРМОНЫ, органические вещества небольшого молекулярного
Экспрессия ТФ нередко находится под контролем фитогормонов
ФИТОГОРМОНЫ, органические вещества небольшого молекулярного
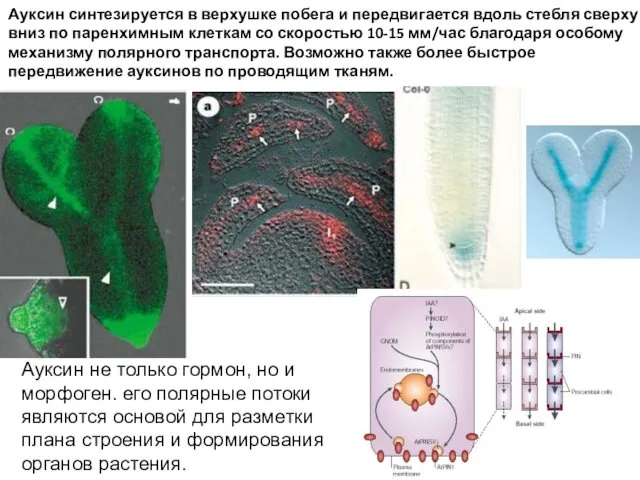
Ауксин синтезируется в верхушке побега и передвигается вдоль стебля сверху вниз
Ауксин синтезируется в верхушке побега и передвигается вдоль стебля сверху вниз
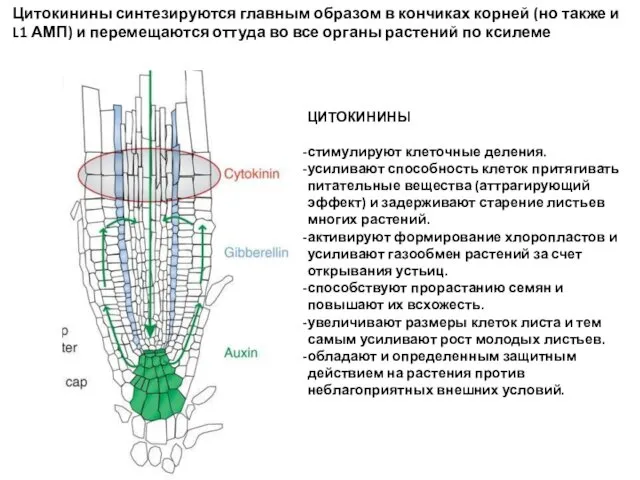
Цитокинины синтезируются главным образом в кончиках корней (но также и L1
Цитокинины синтезируются главным образом в кончиках корней (но также и L1
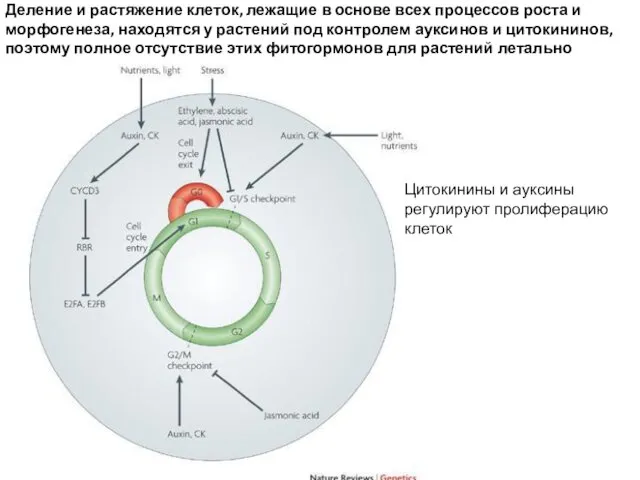
Mitogenic signals are required for proper progression of the cell cycle.
Mitogenic signals are required for proper progression of the cell cycle.

Формирование корней на побегах
Формирование корней в каллусе
Формирование каллуса
Развитие придаточных почек
Развитие пазушных
Формирование корней на побегах
Формирование корней в каллусе
Формирование каллуса
Развитие придаточных почек
Развитие пазушных
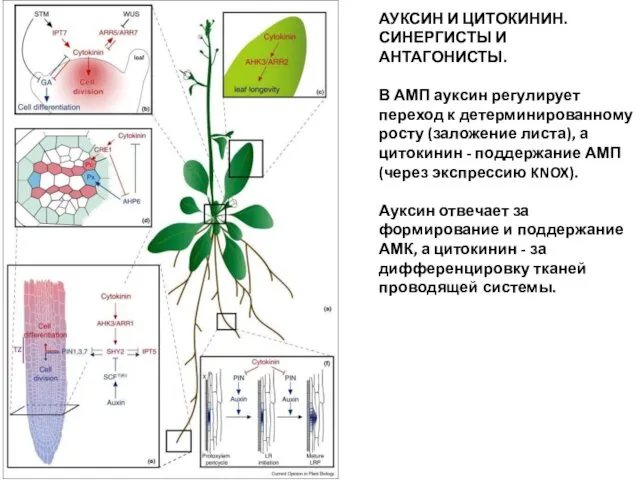
Molecular mechanisms of cytokinin action in different Arabidopsis plant organs.
Molecular mechanisms of cytokinin action in different Arabidopsis plant organs.
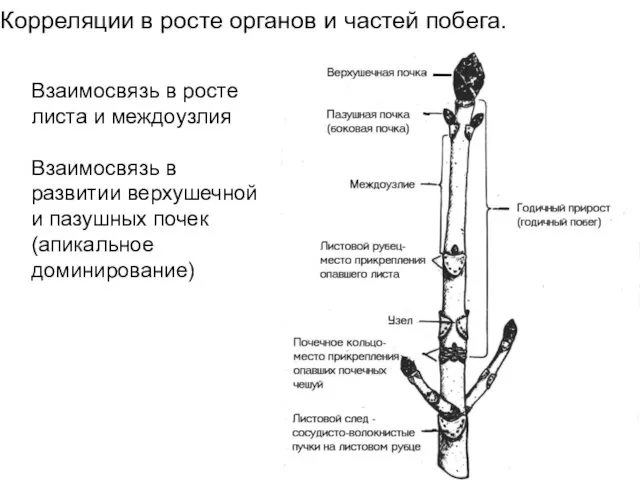
Корреляции в росте органов и частей побега.
Взаимосвязь в росте листа и
Корреляции в росте органов и частей побега.
Взаимосвязь в росте листа и
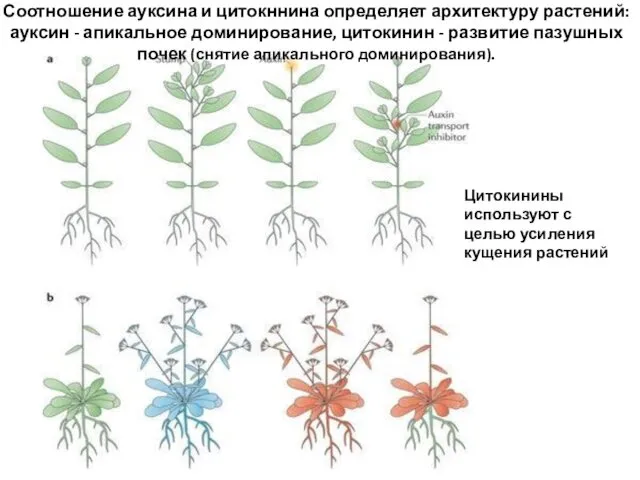
Соотношение ауксина и цитокннина определяет архитектуру растений: ауксин - апикальное доминирование,
Соотношение ауксина и цитокннина определяет архитектуру растений: ауксин - апикальное доминирование,
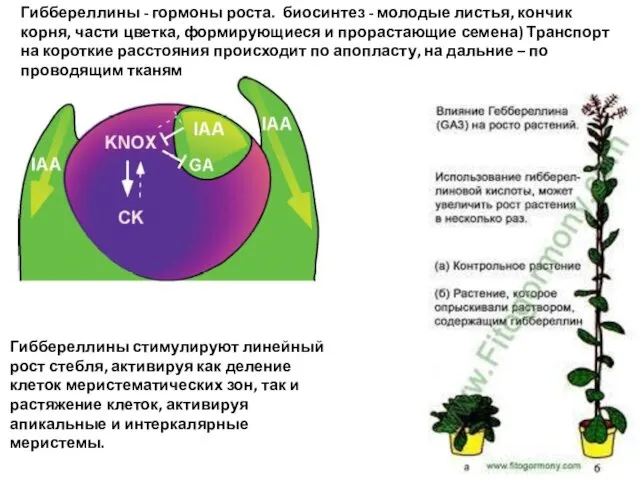
Гиббереллины - гормоны роста. биосинтез - молодые листья, кончик корня, части
Гиббереллины - гормоны роста. биосинтез - молодые листья, кончик корня, части
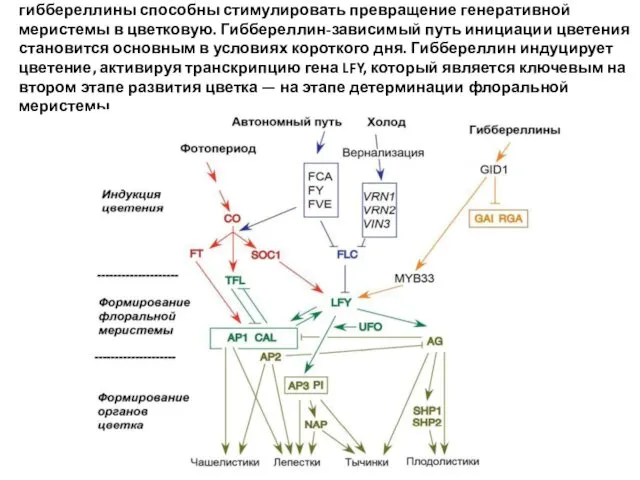
гиббереллины способны стимулировать превращение генеративной меристемы в цветковую. Гиббереллин-зависимый путь инициации
гиббереллины способны стимулировать превращение генеративной меристемы в цветковую. Гиббереллин-зависимый путь инициации
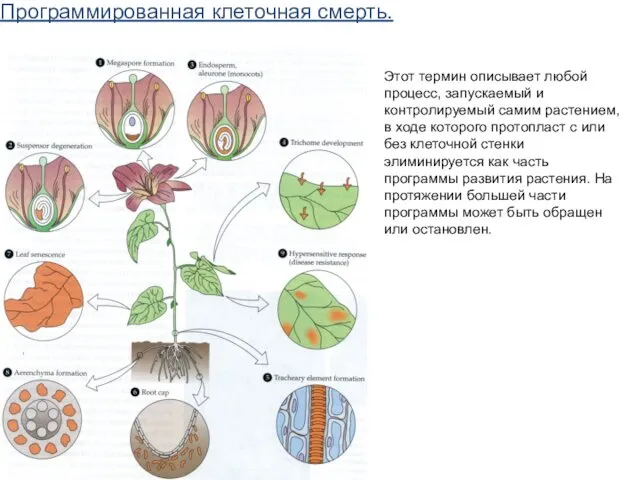
Программированная клеточная смерть.
Этот термин описывает любой процесс, запускаемый и контролируемый
Программированная клеточная смерть.
Этот термин описывает любой процесс, запускаемый и контролируемый

Основные морфотипы программированной клеточной смерти гибели у растений
1. Гиперчувствительная реакция
Основные морфотипы программированной клеточной смерти гибели у растений
1. Гиперчувствительная реакция
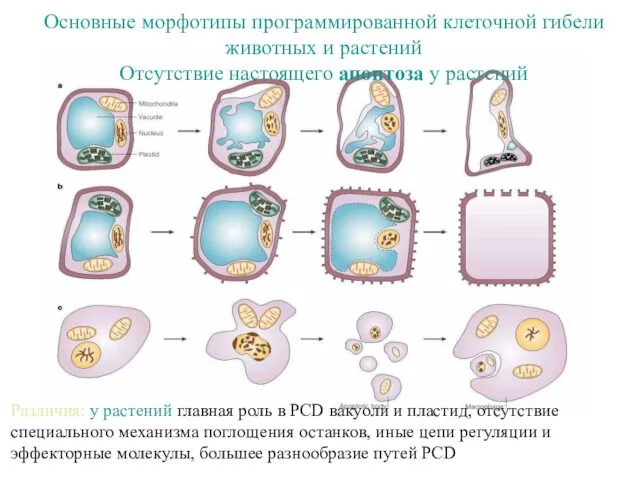
Различия: у растений главная роль в PCD вакуоли и пластид, отсутствие
Различия: у растений главная роль в PCD вакуоли и пластид, отсутствие
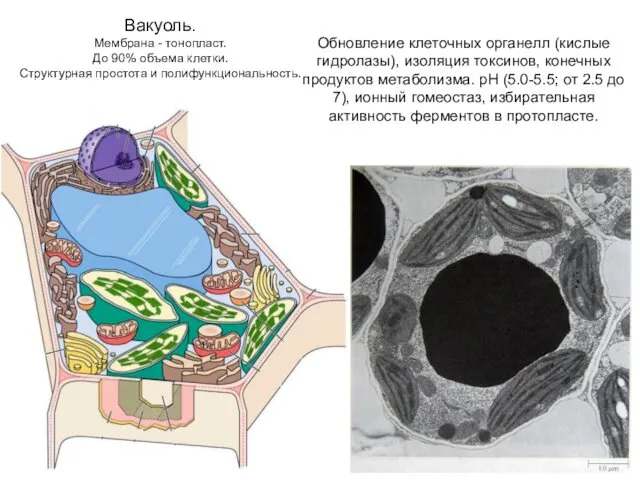
Вакуоль.
Мембрана - тонопласт.
До 90% объема клетки.
Структурная простота и полифункциональность.
Обновление
Вакуоль.
Мембрана - тонопласт.
До 90% объема клетки.
Структурная простота и полифункциональность.
Обновление

Программированная клеточная смерть. Автофагия.
самоперевариванивание посредством автофагосом с последующей секрецией их содержимого
Программированная клеточная смерть. Автофагия.
самоперевариванивание посредством автофагосом с последующей секрецией их содержимого

Программированная клеточная смерть. Автофагия, остановленная на полпути. Интактная плазмалемма. Ситовидные элементы.
Программированная клеточная смерть. Автофагия, остановленная на полпути. Интактная плазмалемма. Ситовидные элементы.
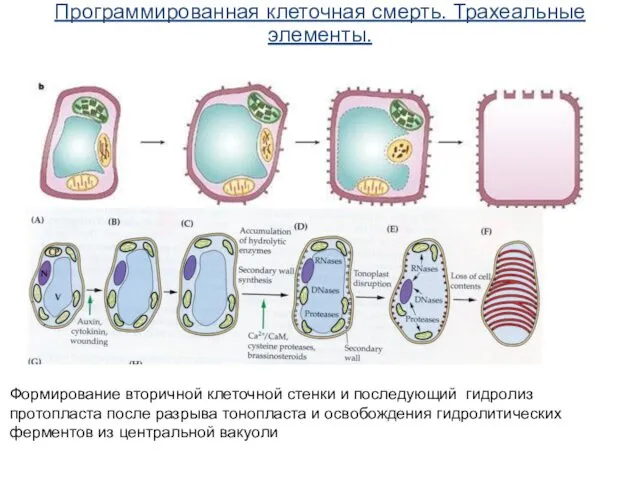
Программированная клеточная смерть. Трахеальные элементы.
Формирование вторичной клеточной стенки и последующий гидролиз
Программированная клеточная смерть. Трахеальные элементы.
Формирование вторичной клеточной стенки и последующий гидролиз
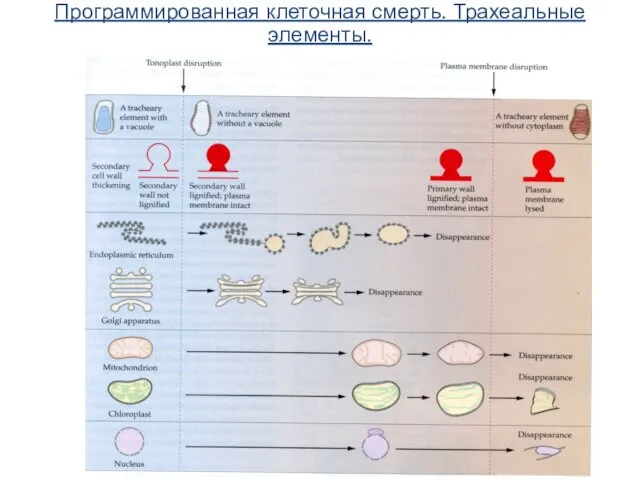
Программированная клеточная смерть. Трахеальные элементы.
Программированная клеточная смерть. Трахеальные элементы.
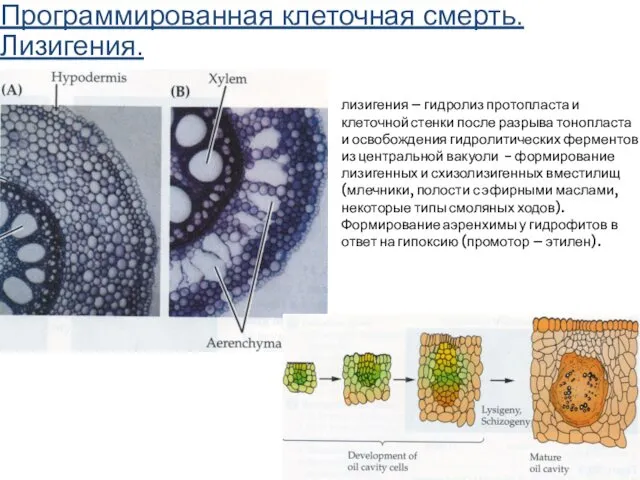
Программированная клеточная смерть. Лизигения.
лизигения – гидролиз протопласта и клеточной стенки
Программированная клеточная смерть. Лизигения.
лизигения – гидролиз протопласта и клеточной стенки

Старение.
Хромопласты. Основные пигменты каротиноиды, ксантофиллы. Окраска цветков, плодов, осенних листьев. Многочисленные
Старение.
Хромопласты. Основные пигменты каротиноиды, ксантофиллы. Окраска цветков, плодов, осенних листьев. Многочисленные

БРАССИНОСТЕРОИДЫ, группа природных регуляторов роста растений, производные ненасыщенных оксистероидов с лактонной
БРАССИНОСТЕРОИДЫ, группа природных регуляторов роста растений, производные ненасыщенных оксистероидов с лактонной
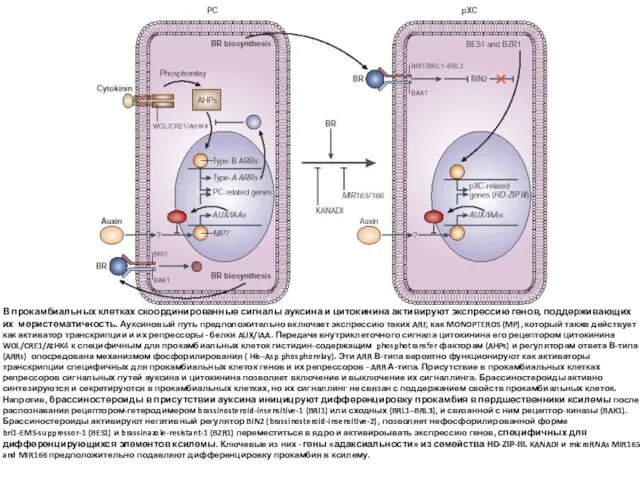
В прокамбиальных клетках скоординированные сигналы ауксина и цитокинина активируют экспрессию генов,
В прокамбиальных клетках скоординированные сигналы ауксина и цитокинина активируют экспрессию генов,

in vitro трансдифференцировка клеток мезофилла (MC) в элементы ксилемы.
Стадия 1. Утрата
in vitro трансдифференцировка клеток мезофилла (MC) в элементы ксилемы.
Стадия 1. Утрата

АБК в листьях, а также в корневом чехлике. Транспорт на короткие
АБК в листьях, а также в корневом чехлике. Транспорт на короткие

Стрессовые воздействия (ранения, водный дефицит, низкая температура), а также высокие концентрации
Стрессовые воздействия (ранения, водный дефицит, низкая температура), а также высокие концентрации

Механизм действия фитогормонов в основных чертах сходен с механизмом действия гормонов
Механизм действия фитогормонов в основных чертах сходен с механизмом действия гормонов
 Грибы съедобные и несъедобные
Грибы съедобные и несъедобные Половое и бесполое размножение Обобщение
Половое и бесполое размножение Обобщение Микроскопические грибы в жизни человека
Микроскопические грибы в жизни человека Как зимуют травы, кустарники, деревья
Как зимуют травы, кустарники, деревья Половая система
Половая система Высшая нервная деятельность. Высшие мозговые функции
Высшая нервная деятельность. Высшие мозговые функции Размножение организмов 6 класс - Презентация
Размножение организмов 6 класс - Презентация Головной мозг
Головной мозг Анализаторы. Органы чувств. Глаз. Ухо
Анализаторы. Органы чувств. Глаз. Ухо Биологические свойства вирусов
Биологические свойства вирусов Особливості Хамелеону
Особливості Хамелеону Состав йогурта с картофельной клетчаткой
Состав йогурта с картофельной клетчаткой Сходство и различия клеток растений грибов и животных
Сходство и различия клеток растений грибов и животных Рептилии. Происхождение рептилий
Рептилии. Происхождение рептилий Систематика эукариотов и прокариотов. Морфология микроорганизмов (лекции №1)
Систематика эукариотов и прокариотов. Морфология микроорганизмов (лекции №1) Ткани животных
Ткани животных Презентация на тему Генетически модифицированные продукты
Презентация на тему Генетически модифицированные продукты Химический состав клетки. Белки (протеины)
Химический состав клетки. Белки (протеины) Особенности строения и жизнедеятельности Ракообразных
Особенности строения и жизнедеятельности Ракообразных Презентация на тему "Привычки и здоровье" - скачать презентации по Биологии
Презентация на тему "Привычки и здоровье" - скачать презентации по Биологии МИНЗДРАВ И МЫ ПРЕДУПРЕЖДАЕМ... Выполнила учащаяся 6 класса Исмоилова Луиза Руководитель проекта учитель ИКТ
МИНЗДРАВ И МЫ ПРЕДУПРЕЖДАЕМ... Выполнила учащаяся 6 класса Исмоилова Луиза Руководитель проекта учитель ИКТ  Селекция – эволюция, направляемая человеком
Селекция – эволюция, направляемая человеком Антропогенные экосистемы С.Е. ДРОМАШКО
Антропогенные экосистемы С.Е. ДРОМАШКО  Растения-паразиты Презентация к уроку экологии в 6 классе
Растения-паразиты Презентация к уроку экологии в 6 классе  Жан Батист Ламарк. 1744 ― 1828
Жан Батист Ламарк. 1744 ― 1828 Мутации
Мутации Физиология ретикулярной формации
Физиология ретикулярной формации Формула цветка
Формула цветка