- Метаболические пути

Содержание
- 2. ОСНОВНЫЕ МЕТАБОЛИЧЕСКИЕ ПУТИ Характер метаболизма в тканях во многом определяется питанием. У человека и ряда других
- 3. Тканевое дыхание Это процесс потребление клетками тканей организма кислорода, который участвует в биологическом окислении. Такой вид
- 4. Дыхательная цепь состоит из: 1. НАД - зависимой дегидрогеназы; 2. ФАД- зависимой дегидрогеназы; 3. Убихинона (КоQ);
- 5. Механизм работы электронтранспортной цепи Наружная мембрана митохондрии (рис. 4.8.1)проницаема для большинства мелких молекул и ионов, внутренняя
- 6. Механизм работы электронтранспортной цепи Процессы окисления и образования АТФ из АДФ и фосфорной кислоты т.е. фосфорилирования
- 7. Метаболизм углеводов У всех млекопитающих глюкоза в клетках превращается в пируват и лактат по метаболическому пути,
- 8. Метаболизм углеводов Таким образом, глюкоза служит главным видом топлива для многих тканей, однако она (а также
- 9. Метаболизм липидов Источником длинноцепочечных жирных кислот служат синтез de novo из ацетил-СоА (в свою очередь образующегося
- 10. Метаболизм аминокислот Аминокислоты необходимы для синтеза белков. Некоторые из них должны обязательно поступать с пищей (незаменимые
- 11. Список литературы Марри Р., Греннер Д., Мейес П., Родуэлл В. Биохимия человека: В 2-х томах. Т.
- 13. Скачать презентацию

ОСНОВНЫЕ МЕТАБОЛИЧЕСКИЕ ПУТИ
Характер метаболизма в тканях во многом определяется питанием. У
ОСНОВНЫЕ МЕТАБОЛИЧЕСКИЕ ПУТИ
Характер метаболизма в тканях во многом определяется питанием. У

Тканевое дыхание
Это процесс потребление клетками тканей организма кислорода, который участвует в
Тканевое дыхание
Это процесс потребление клетками тканей организма кислорода, который участвует в

Дыхательная цепь состоит из:
1. НАД - зависимой дегидрогеназы;
2. ФАД- зависимой дегидрогеназы;
3.
Дыхательная цепь состоит из:
1. НАД - зависимой дегидрогеназы;
2. ФАД- зависимой дегидрогеназы;
3.
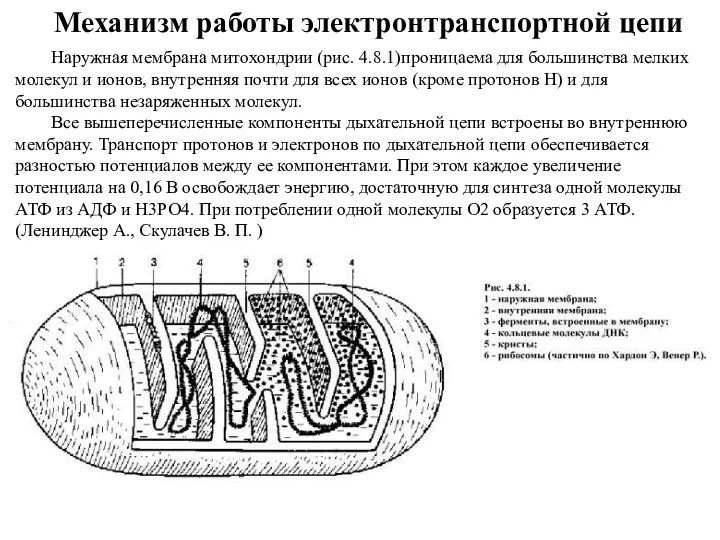
Механизм работы электронтранспортной цепи
Наружная мембрана митохондрии (рис. 4.8.1)проницаема для большинства мелких
Механизм работы электронтранспортной цепи
Наружная мембрана митохондрии (рис. 4.8.1)проницаема для большинства мелких

Механизм работы электронтранспортной цепи
Процессы окисления и образования АТФ из АДФ и
Механизм работы электронтранспортной цепи
Процессы окисления и образования АТФ из АДФ и

Метаболизм углеводов
У всех млекопитающих глюкоза в клетках превращается в пируват и
Метаболизм углеводов
У всех млекопитающих глюкоза в клетках превращается в пируват и

Метаболизм углеводов
Таким образом, глюкоза служит главным видом топлива для многих тканей,
Метаболизм углеводов
Таким образом, глюкоза служит главным видом топлива для многих тканей,
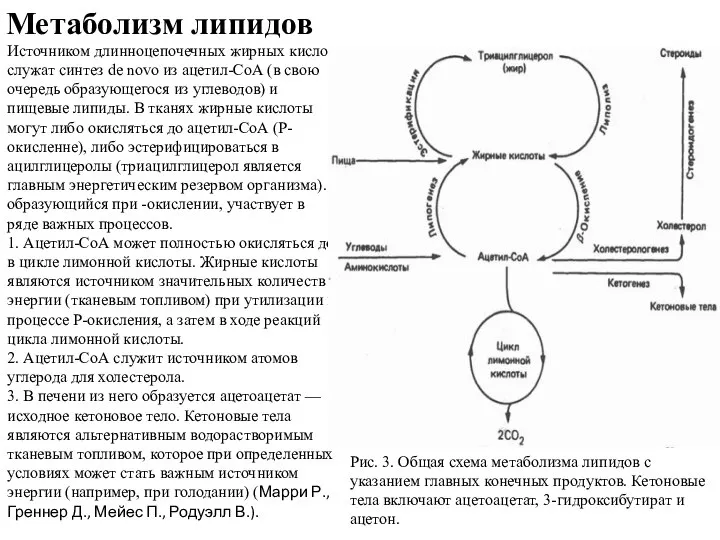
Метаболизм липидов
Источником длинноцепочечных жирных кислот служат синтез de novo из ацетил-СоА
Метаболизм липидов
Источником длинноцепочечных жирных кислот служат синтез de novo из ацетил-СоА
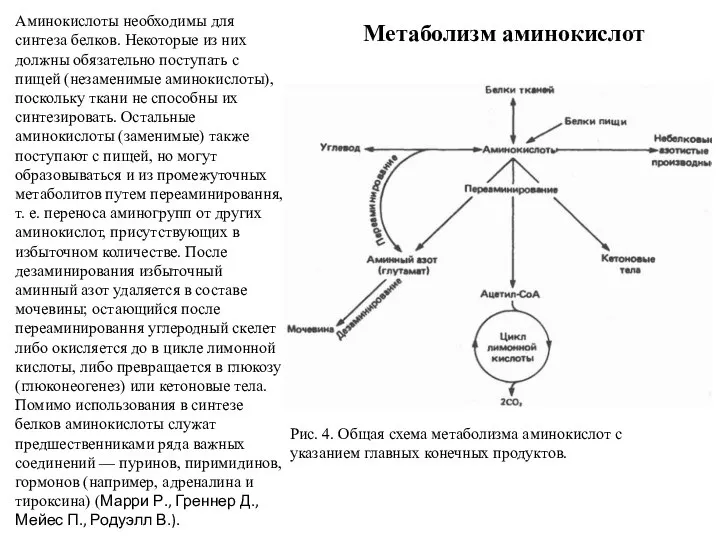
Метаболизм аминокислот
Аминокислоты необходимы для синтеза белков. Некоторые из них должны обязательно
Метаболизм аминокислот
Аминокислоты необходимы для синтеза белков. Некоторые из них должны обязательно

Список литературы
Марри Р., Греннер Д., Мейес П., Родуэлл В. Биохимия
Список литературы
Марри Р., Греннер Д., Мейес П., Родуэлл В. Биохимия
 Критические периоды раннего онтогенеза
Критические периоды раннего онтогенеза 20170927_krasnaya_kniga_buryatii
20170927_krasnaya_kniga_buryatii Птицы под нашим окном
Птицы под нашим окном Презентация по биологии Отряд хищные
Презентация по биологии Отряд хищные  Шенграбен и Аустерлиц «Мне совестно было писать о нашем торжестве, не описав наших неудач и нашего срама» Л.Н. Толстой, черновы
Шенграбен и Аустерлиц «Мне совестно было писать о нашем торжестве, не описав наших неудач и нашего срама» Л.Н. Толстой, черновы Віруси Віруси — неклітинні форми живих організмів , які складаються з нуклеїнової кислоти (ДНК або РНК) і білкової оболонки, зр
Віруси Віруси — неклітинні форми живих організмів , які складаються з нуклеїнової кислоти (ДНК або РНК) і білкової оболонки, зр Биогеоценотический уровень
Биогеоценотический уровень Пресноводная жемчужница Margaritifera margaritifera L. рек северной Европы: особенности экологии и пути сохранения
Пресноводная жемчужница Margaritifera margaritifera L. рек северной Европы: особенности экологии и пути сохранения ЖИВОТНЫЕ ПРОШЛОГО Выполнили Сазанова Д., Алпеева А. 5 « А» класс МБОУ СОШ № 27
ЖИВОТНЫЕ ПРОШЛОГО Выполнили Сазанова Д., Алпеева А. 5 « А» класс МБОУ СОШ № 27  Органы чувств
Органы чувств Bill Gates is one of the most influential people in the world. He is cofounder of the most recognized brands in the computer
Bill Gates is one of the most influential people in the world. He is cofounder of the most recognized brands in the computer Шаблон тренажера
Шаблон тренажера Простейшие Жгутиковые
Простейшие Жгутиковые Биологиялық белсенді қосылыстардың иондалу және липофильділік мәселелері
Биологиялық белсенді қосылыстардың иондалу және липофильділік мәселелері Конярство. Конярство в Україні
Конярство. Конярство в Україні Антропогенез «Создает человека природа, но развивает и образует его общество». &
Антропогенез «Создает человека природа, но развивает и образует его общество». & Трутовик лиственничный
Трутовик лиственничный Проблемы увядающей кожи. Серия омолаживающей продукции TS
Проблемы увядающей кожи. Серия омолаживающей продукции TS Новые представления о строении и функции системы кожных покровов. Морфологические основы действия ретиноидов
Новые представления о строении и функции системы кожных покровов. Морфологические основы действия ретиноидов Птицы Переволоцкого района
Птицы Переволоцкого района Скандинавская ходьба и возрастные изменения
Скандинавская ходьба и возрастные изменения Презентация на тему Особенности растительной клетки
Презентация на тему Особенности растительной клетки  Наші пухнасті друзі
Наші пухнасті друзі Внутреннее строение земноводных Учитель биологии МОУ Неклюдовская СОШ Отряскиной Т.А.
Внутреннее строение земноводных Учитель биологии МОУ Неклюдовская СОШ Отряскиной Т.А. Презентация на тему "Общие свойства живого" - скачать презентации по Биологии
Презентация на тему "Общие свойства живого" - скачать презентации по Биологии Тема урока: «Развитие опорно-двигательной системы» РАЗРАБОТАЛА УЧИТЕЛЬ БИОЛОГИИ МБОУ СОШ №3 Г. БОГОРОДСКА НИЖЕГОРОДСКОЙ ОБЛАСТИ
Тема урока: «Развитие опорно-двигательной системы» РАЗРАБОТАЛА УЧИТЕЛЬ БИОЛОГИИ МБОУ СОШ №3 Г. БОГОРОДСКА НИЖЕГОРОДСКОЙ ОБЛАСТИ  Дикорастущие деревья Крыма
Дикорастущие деревья Крыма Флора и Фауна
Флора и Фауна