- Микология. Место грибов в живом мире, строение, общая характеристика, разнообразие

Содержание
- 2. Место грибов в живом мире, строение, общая характеристика, разнообразие
- 3. «Fungorum ordo chaos est» C. Linnaeus Наука о грибах называется микология (от греческих слов myces –
- 4. Критерии для группировки организмов Структурно-морфологический сравнительное морфологическое описание признаков организмов, обнаруживаемых визуально и различных методов микроскопии
- 5. Морфологические признаки, используемые для описания вида у дрожжевых грибов Рост в жидком сусле ● образование пленки,
- 6. Физиолого-биохимические признаки, используемые для описания вида у дрожжевых грибов ● Брожение сахаров: 5-10 источников. ● Ассимиляция
- 7. Филогенетический анализ позволяет установить родство организмов по генотипам , а не сходство по фенотипам, что может
- 8. Участки рДНК, используемые для определения филогенетического положения грибов Между рибосомальными генами имеются внутригенные и межгенные спейсерные
- 9. Филогенетическое дерево базидиомицетовых дрожжей
- 10. «Нужно всегда помнить, что любое построенное дерево является лишь филогенетической гипотезой, с некоторой долей вероятности описывающей
- 11. Карл Линней (1707-1778) “Система природы”, 1735 “ Minerals exist; plants exist and live; animals exist, live
- 12. Э.Фриз в 1831 г. предложил выделить грибы в самостоятельное царство живого мира, но у большинства биологов
- 13. Пять царств Monera (Procaryota) – отсутствие ядерной мембраны. Эукариоты: Protista – одноклеточные подвижные микроорганизмы с ядром
- 14. автотрофы гетеротрофы (способны к синтезу органических (питаются готовыми органическими веществ из неорганических) веществами) хемотрофы фототрофы осмотрофы
- 15. Грибы – эукариотические гетеротрофные организмы (не имеющие хлорофилла) с абсорционным (осмотрофным) типом питания, размножаются спорами, их
- 16. ГРИБЫ Грибы имеют способ питания, отличающий их от других эукариотных организмов - всасывание питательных веществ из
- 17. 4. Грибы накачивают из окружающей среды в мицелий воду с растворенными питательными веществами благодаря высокому тургорному
- 18. 5. Сахара, проникающие в клетку, быстро превращаются в шестиатомный спирт манит, дисахарид трегалозу, животный крахмал –
- 19. 7. Для распространения спор грибы образуют на поверхности субстрата различные органы спороношения. Сигналы к споруляции. 8.
- 20. Слои клеточной стенки Neurospora crassa: г – микрофибриллы хитина в слое белка, в – белок, б
- 22. Строение мицелия Неограниченный верхушечный рост. Отсутствует сопряженность цитокинеза и митоза (многоядерность). Миграция органелл.
- 23. Ультраструктура септ Аскомицеты: простая септа Базидиомицеты: сложная септа (долипоровая с перфорированной мембранной парентосомой)
- 24. Септа аскомицетов с тельцами Воронина
- 25. МИЦЕЛИАЛЬНЫЙ ОБРАЗ ЖИЗНИ (ОСОБЕННОСТИ) 1. Апикальный рост Скорость удлинения кончика гифы Neurospora crassa составляет 16 мкм
- 26. Покоящиеся и проводящие структуры Склероции, хламидоспоры Синнемы – сросшиеся боковыми сторонами гифы Крупные синнемы – тяжи
- 27. ОСОБЕННОСТИ ДРОЖЖЕВОГО РОСТА 1.Диффузный (а не апикальный) рост 2. Преобладание в клеточной стенке маннана; хитин –
- 28. Мицелиально-дрожевой диморфизм Истинный мицелий Псевдомицелий Дрожжевой рост
- 29. МИЦЕЛИАЛЬНО-ДРОЖЖЕВОЙ ДИМОРФИЗМ ОБУСЛОВЛЕН: Газовым составом среды (аэробные/анаэробные условия) Состоянием ядер в клетке (базидиальные головневые – дикариотический
- 30. Фитопатогены Грибы применяют ферменты для лизиса кутикулы и клеточной стенки H. Strasser
- 31. Fungi use mechanical force to penetrate host cuticle H. Strasser
- 32. апрессорий гаусторий
- 33. Митоз Ядерная оболочка сохраняется во все фазы (закрытый митоз) Центриоли только у жгутиковых грибов. У бол-ва
- 34. Геном у грибов – 25-40 млн. нуклеотидных пар. Низкий процент повторяющихся последовательностей (10-15%). Митохондриальный геном –
- 35. Размножение Вегетативное (мицелий, оидии, хламидоспоры), бесполое (зооспоры, спорангиоспоры, конидии) и половое (смена ядерных фаз, три стадии
- 36. Явление разноядерности – наличие в клетках ядер, гетероаллельных по некоторым генам. Заменяет грибам гетерозиготность. Если у
- 37. Секторность грибной колонии
- 38. Парасексуальный (псевдополой) процесс – слияние гаплоидных ядер гетероаллельных по какому-то локусу может дать начало диплоидному гетерозиготному
- 39. Анастомозы, происходящие между соседними гифами внутри колонии, армируют ее дают более прочной, сохраняют ее единство при
- 41. Совместимость - слияние колоний и отсутствие разграничительных линий, при сращивании изолятов равнозначных морфотипов Сryptonectria parasitica (поражает
- 42. Барраж - линия несовместимости в зоне контакта двух колоний при сокультивировании изолятов различных морфотипов Сryptonectria parasitica.
- 43. Вегетативная несовместимость – механизм защиты грибов от вирусной инфекции, так как вирусы снижают способность особи к
- 44. ОБЩАЯ ХАРАКТЕРИСТИКА ГРИБОВ Углеводы клеточной стенки – хитин и глюканы (маннаны) Синтез лизина – через альфа-аминоадипиновую
- 45. Что же дал филогенетический анализ для определения места грибов в системе живых организмов?
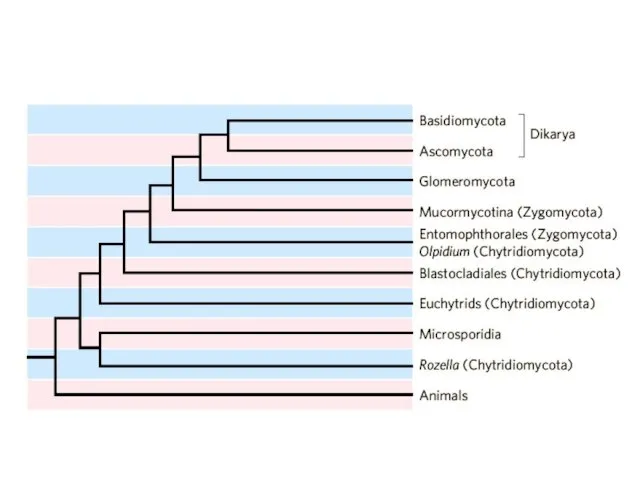
- 47. Организмы, изучаемые микологами, относятся к трем большим независимо эволюционирующим группам, которым разные систематики присваивают ранги выше
- 48. ** – диаминопимелиновую кислоту
- 50. . A: Posteriorly uniflagellate zoospores. Note the refractive lipid sac and slipper shape. B: Aggregation on
- 51. Cryptomycota
- 52. Cryptomycota Cryptomycota
- 53. Tree of Life: Fungi
- 54. Kingdom: Fungi R. T. Moore, 1980 (Hibbett et al., 2007. A higher-level phylogenetic classification of the
- 55. Glomeromycota Glomeromycota Истинные грибы
- 56. Иерархическая классификация Regnum- Kingdom- Царство Fungi Divisio- Phylum- Отдел Basidiomycota Classis- Class- Класс Basidiomycetes Ordo- Order-
- 57. Agaricus bisporus (Lange) Imbach Lange первым описал этот гриб как Coprinus bisporus Imbach позднее перевел этот
- 58. Экологические группы грибов
- 59. Сапротрофные грибы Гифы грибов в почве в люминисцентном микроскопе Посев из почвы на среду Чапека Плодовые
- 60. Ксилотрофные грибы – разрушители древесины Домовой гриб (Serpula lacrimans) Трутовые грибы (пор. Aphyllophorales) Бурая гниль Белая
- 61. Фитопатогенные грибы Фитопатология – одна из важнейших отраслей экологической и прикладной биологии Раневые паразиты (Nectria) Мучнисторосяные
- 62. Микопаразитические грибы Микопаразитические грибы, растущие на плодовых телах агариковых грибов
- 63. Грибы – паразиты животных Энтомопатогенные грибы Entomophthora (зигомицеты) Beauveria bassiana Энтомопатогенные грибы используются для производства биоинсектецидов
- 64. Хищные грибы Хищные грибы (Arthrobotrys, Monacrosporium, Dactylaria, Caenorhabditis) могут жить как сапротрофы, но способны улавливать мелких
- 65. Царство Fungi (Eumycota) Отделы: Chytridiomycota Образуют подвижные споры называемые зооспорами Зигота, мейоз происходит в покоящемся спорангии,
- 66. Какое количество видов грибов существует? По разным источникам грибов от 100 000 до 250 000 видов;
- 67. Оценка максимально возможного разнообразия грибов Цветковых растений на Британских островах описано = 2,000 видов Грибов на
- 68. (a) Estimated totals of global diversity are given as ranges as the estimates depend on the
- 69. Потенциал исследований видового разнообразия грибов Грибы в экстремальных местообитаниях Обширные регионы не изучены (тропики, Антартида, отдаленные
- 71. Когда возникли грибы? Science vol. 289
- 72. Грибы древние организмы Многие виды грибов возникли 450 млн.лет назад, а, возможно, и более 1 млрд.
- 73. Скелетные Биотические события в истории Земли Симбиогенез Прокариоты цианобактерии Растения Водо-росли Млрд. лет назад * -
- 74. Berbee and Taylor, 2001 Расчеты на основе уточненной скорости замены нуклеотидных последовательностей – ( 1.26% за
- 75. Berbee and Taylor, 1993 Ascomycetes отошли от базидиомицетов Дрожжи, обычные плесени эволюционировали после покрытосеменных Glomales отошли
- 76. Heckman et al. (2001). Molecular Evidence for the Early Colonization of Land by Fungi and Plants
- 77. Грибы, из водоемов, стали переходить к наземной жизни около 1,3 млрд. лет назад (Blair Hedges, Science,
- 78. Aскокaрп в Asteroxylon http://www.xs4all.nl/~steurh/engrhyn/erhynie.html Растения и грибы в ископаемых отложениях (Ринийских сланцах, Ранний Девон, ~ 408-360
- 79. Ландшафт и растительный покров на Земле в Меловой период (56-146 млн. лет назад) широколиственные деревья (1),
- 80. Archaeomarasmius (Hibbett et al. 1997) Protomycena (Hibbett et al.,1997) Aureofungus (3,4) в Доминиканском янтаре (23,8 до
- 81. Fungi
- 82. Продолжительность жизни грибов От нескольких суток до тысячилетий.
- 83. Л-2
- 85. ОСОБЕННОСТИ ГРИБОВ И ГРИБОПОДОБНЫХ ОРГАНИЗМОВ
- 86. CНROMISTA Hyphochytriomycota Labyrinthulomycota Oomycota FUNGI Ascomycota Ascomycetes Neolectomycetes Pneumocystidomycetes Saccharomycetes Schizosaccharomycetes Taphrinomycetes Basidiomycota Basidiomycetes Urediniomycetes Ustilaginomycetes
- 90. Kingdom: Fungi R. T. Moore, 1980 (Hibbett et al., 2007. A higher-level phylogenetic classification of the
- 92. Скачать презентацию

Место грибов в живом мире, строение, общая характеристика, разнообразие
Место грибов в живом мире, строение, общая характеристика, разнообразие

«Fungorum ordo chaos est»
C. Linnaeus
Наука о грибах называется микология (от
«Fungorum ordo chaos est»
C. Linnaeus
Наука о грибах называется микология (от

Критерии для группировки организмов
Структурно-морфологический
сравнительное морфологическое описание признаков организмов, обнаруживаемых визуально
Критерии для группировки организмов
Структурно-морфологический
сравнительное морфологическое описание признаков организмов, обнаруживаемых визуально
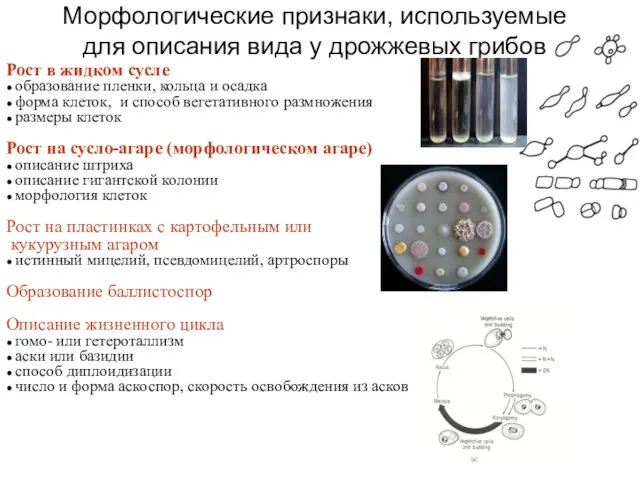
Морфологические признаки, используемые для описания вида у дрожжевых грибов
Рост в жидком
Морфологические признаки, используемые для описания вида у дрожжевых грибов
Рост в жидком

Физиолого-биохимические признаки, используемые для описания вида у дрожжевых грибов
● Брожение сахаров:
Физиолого-биохимические признаки, используемые для описания вида у дрожжевых грибов
● Брожение сахаров:

Филогенетический анализ позволяет установить родство организмов по генотипам ,
а не
Филогенетический анализ позволяет установить родство организмов по генотипам ,
а не
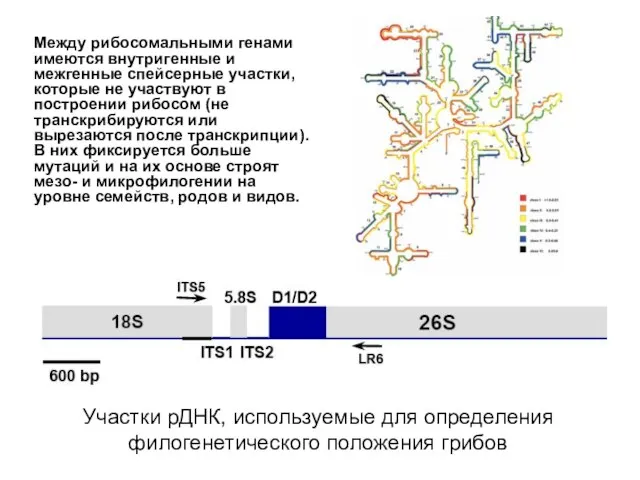
Участки рДНК, используемые для определения филогенетического положения грибов
Между рибосомальными генами имеются
Участки рДНК, используемые для определения филогенетического положения грибов
Между рибосомальными генами имеются

Филогенетическое дерево базидиомицетовых дрожжей
Филогенетическое дерево базидиомицетовых дрожжей

«Нужно всегда помнить, что любое построенное дерево является лишь филогенетической гипотезой,
«Нужно всегда помнить, что любое построенное дерево является лишь филогенетической гипотезой,

Карл Линней (1707-1778) “Система природы”, 1735
“ Minerals exist; plants exist
Карл Линней (1707-1778) “Система природы”, 1735
“ Minerals exist; plants exist

Э.Фриз в 1831 г. предложил выделить грибы в самостоятельное царство живого
Э.Фриз в 1831 г. предложил выделить грибы в самостоятельное царство живого
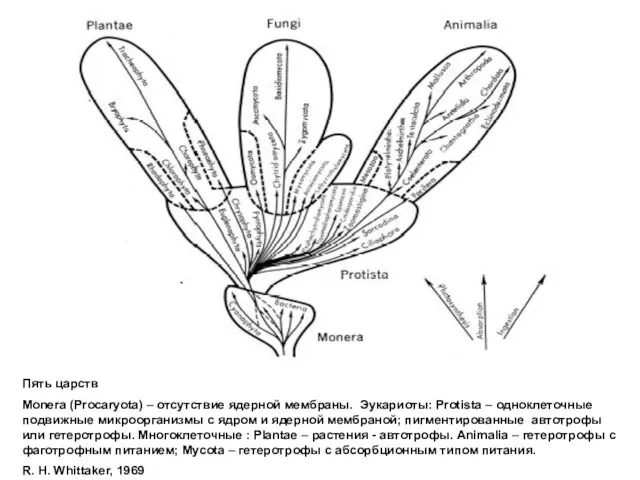
Пять царств
Monera (Procaryota) – отсутствие ядерной мембраны. Эукариоты: Protista –
Пять царств
Monera (Procaryota) – отсутствие ядерной мембраны. Эукариоты: Protista –

автотрофы гетеротрофы
(способны к синтезу органических (питаются готовыми органическими
веществ из
автотрофы гетеротрофы
(способны к синтезу органических (питаются готовыми органическими
веществ из

Грибы – эукариотические гетеротрофные организмы (не имеющие хлорофилла) с абсорционным (осмотрофным)
Грибы – эукариотические гетеротрофные организмы (не имеющие хлорофилла) с абсорционным (осмотрофным)

ГРИБЫ
Грибы имеют способ питания, отличающий их от других эукариотных организмов
ГРИБЫ
Грибы имеют способ питания, отличающий их от других эукариотных организмов
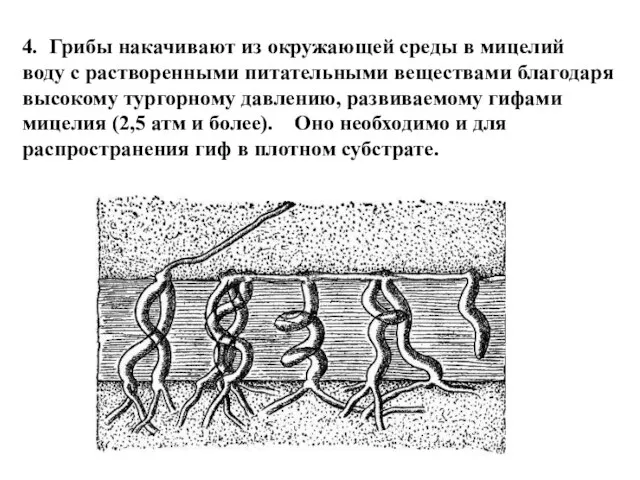
4. Грибы накачивают из окружающей среды в мицелий
воду с растворенными питательными
воду с растворенными питательными

5. Сахара, проникающие в клетку, быстро превращаются в шестиатомный спирт манит,
5. Сахара, проникающие в клетку, быстро превращаются в шестиатомный спирт манит,

7. Для распространения спор грибы образуют на поверхности субстрата различные органы
7. Для распространения спор грибы образуют на поверхности субстрата различные органы
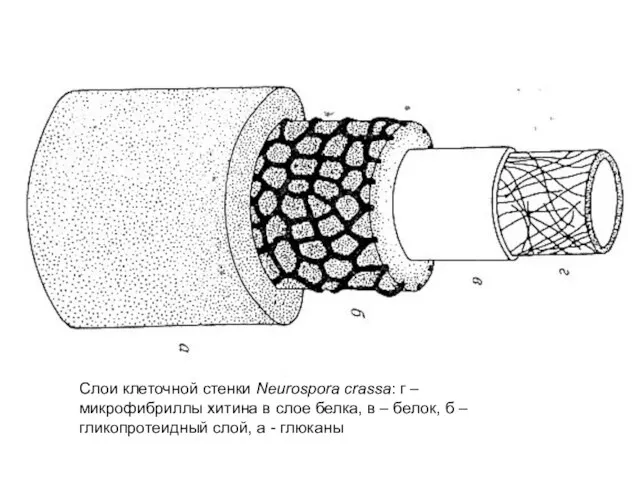
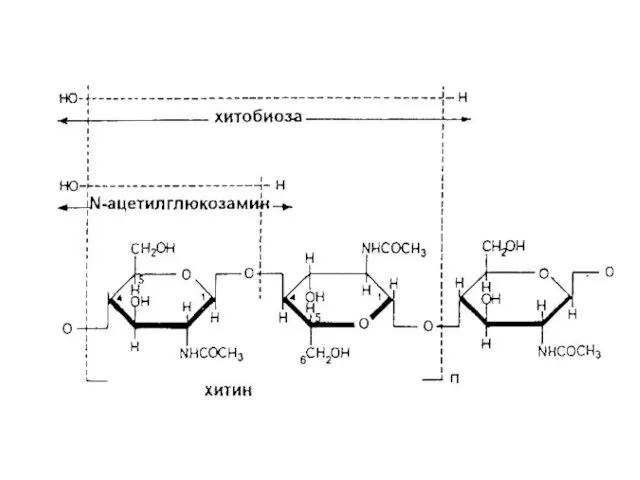
Слои клеточной стенки Neurospora crassa: г – микрофибриллы хитина в слое
Слои клеточной стенки Neurospora crassa: г – микрофибриллы хитина в слое
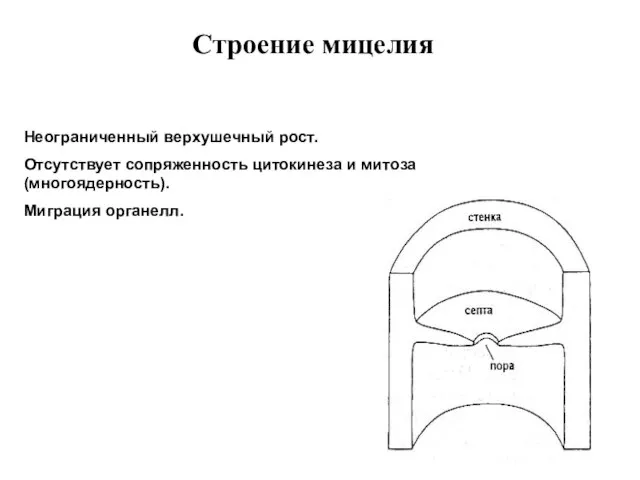
Строение мицелия
Неограниченный верхушечный рост.
Отсутствует сопряженность цитокинеза и митоза (многоядерность).
Миграция органелл.
Строение мицелия
Неограниченный верхушечный рост.
Отсутствует сопряженность цитокинеза и митоза (многоядерность).
Миграция органелл.
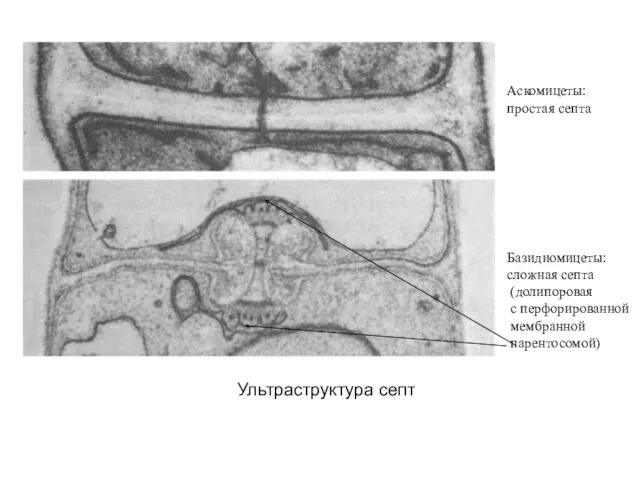
Ультраструктура септ
Аскомицеты:
простая септа
Базидиомицеты:
сложная септа
(долипоровая
с перфорированной
мембранной
парентосомой)
Ультраструктура септ
Аскомицеты:
простая септа
Базидиомицеты:
сложная септа
(долипоровая
с перфорированной
мембранной
парентосомой)
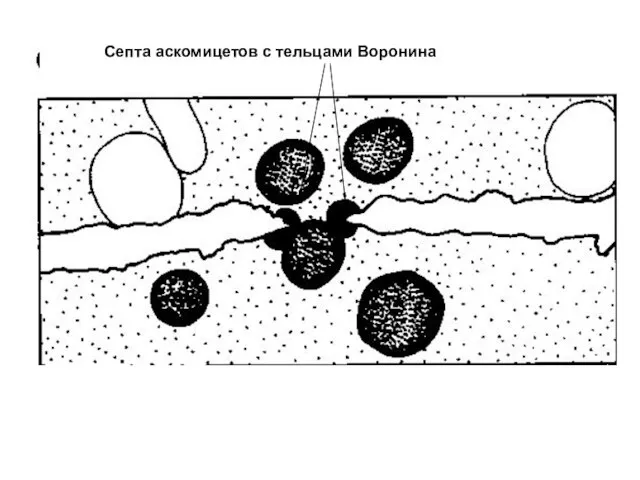
Септа аскомицетов с тельцами Воронина
Септа аскомицетов с тельцами Воронина
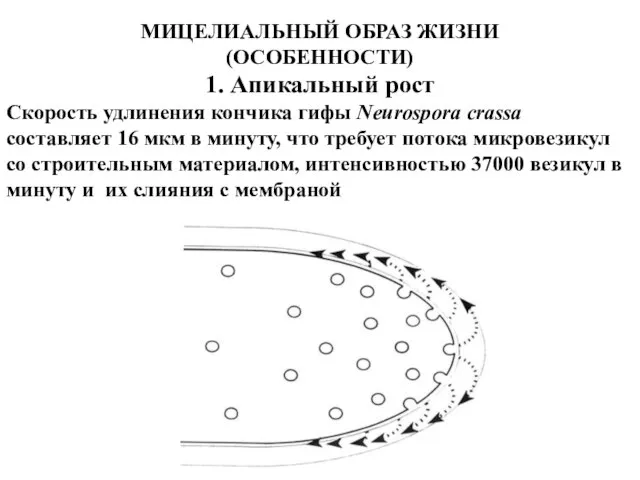
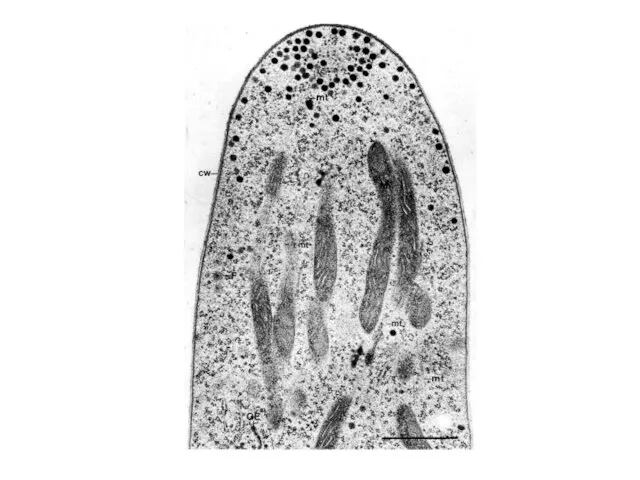
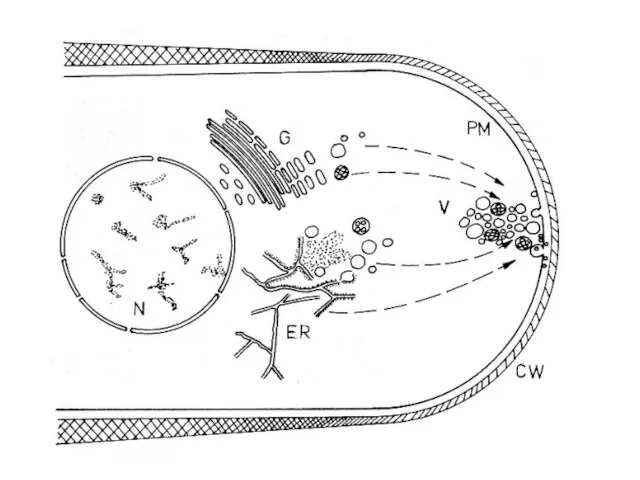
МИЦЕЛИАЛЬНЫЙ ОБРАЗ ЖИЗНИ
(ОСОБЕННОСТИ)
1. Апикальный рост
Скорость удлинения кончика гифы Neurospora crassa составляет
МИЦЕЛИАЛЬНЫЙ ОБРАЗ ЖИЗНИ
(ОСОБЕННОСТИ)
1. Апикальный рост
Скорость удлинения кончика гифы Neurospora crassa составляет

Покоящиеся и проводящие структуры
Склероции, хламидоспоры
Синнемы – сросшиеся боковыми сторонами гифы
Крупные синнемы
Покоящиеся и проводящие структуры
Склероции, хламидоспоры
Синнемы – сросшиеся боковыми сторонами гифы
Крупные синнемы
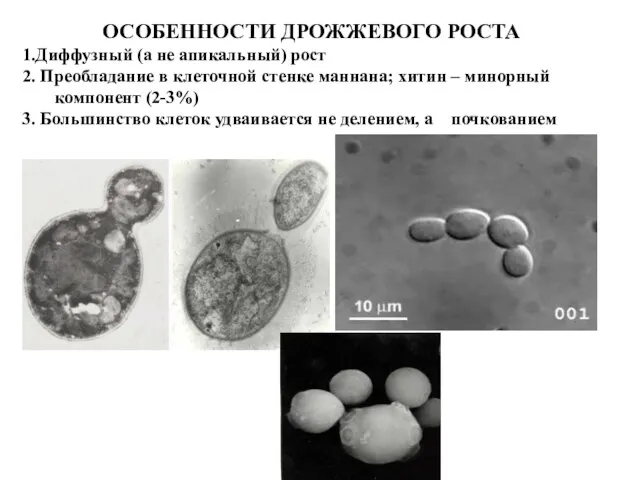
ОСОБЕННОСТИ ДРОЖЖЕВОГО РОСТА
1.Диффузный (а не апикальный) рост
2. Преобладание в клеточной стенке
ОСОБЕННОСТИ ДРОЖЖЕВОГО РОСТА
1.Диффузный (а не апикальный) рост
2. Преобладание в клеточной стенке
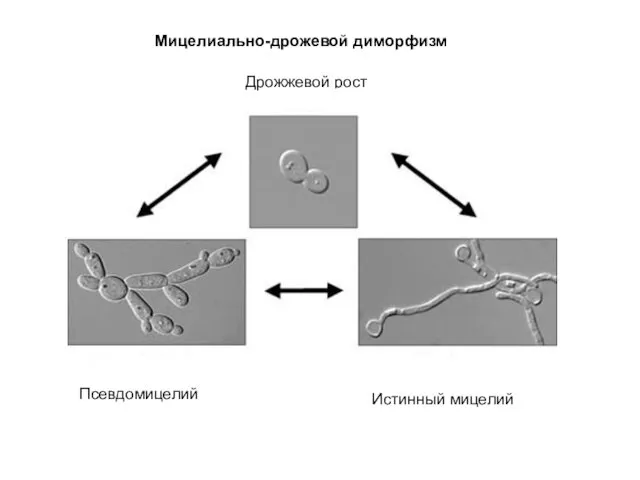
Мицелиально-дрожевой диморфизм
Истинный мицелий
Псевдомицелий
Дрожжевой рост
Мицелиально-дрожевой диморфизм
Истинный мицелий
Псевдомицелий
Дрожжевой рост

МИЦЕЛИАЛЬНО-ДРОЖЖЕВОЙ
ДИМОРФИЗМ ОБУСЛОВЛЕН:
Газовым составом среды (аэробные/анаэробные условия)
Состоянием ядер в
МИЦЕЛИАЛЬНО-ДРОЖЖЕВОЙ
ДИМОРФИЗМ ОБУСЛОВЛЕН:
Газовым составом среды (аэробные/анаэробные условия)
Состоянием ядер в

Фитопатогены
Грибы применяют ферменты для лизиса кутикулы и клеточной стенки
H. Strasser
Фитопатогены
Грибы применяют ферменты для лизиса кутикулы и клеточной стенки
H. Strasser
Fungi use mechanical force to penetrate host cuticle
H. Strasser
Fungi use mechanical force to penetrate host cuticle
H. Strasser
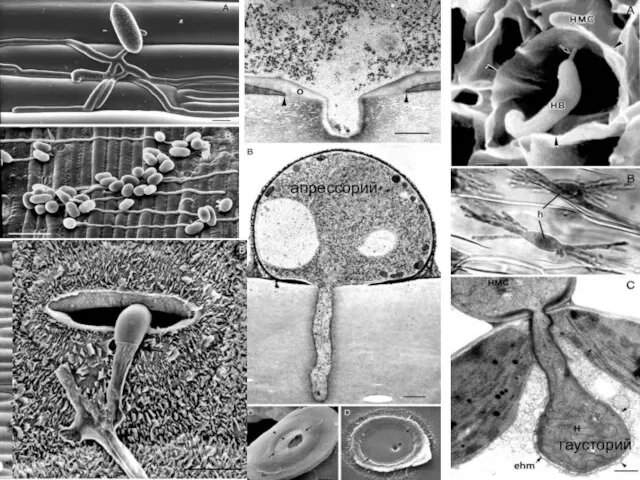
апрессорий
гаусторий
апрессорий
гаусторий

Митоз
Ядерная оболочка сохраняется во все фазы (закрытый митоз)
Центриоли только у жгутиковых
Митоз
Ядерная оболочка сохраняется во все фазы (закрытый митоз)
Центриоли только у жгутиковых

Геном у грибов – 25-40 млн. нуклеотидных пар.
Низкий процент повторяющихся последовательностей
Геном у грибов – 25-40 млн. нуклеотидных пар.
Низкий процент повторяющихся последовательностей

Размножение
Вегетативное (мицелий, оидии, хламидоспоры), бесполое (зооспоры, спорангиоспоры, конидии) и половое (смена
Размножение
Вегетативное (мицелий, оидии, хламидоспоры), бесполое (зооспоры, спорангиоспоры, конидии) и половое (смена

Явление разноядерности – наличие в клетках ядер, гетероаллельных по некоторым генам.
Заменяет
Явление разноядерности – наличие в клетках ядер, гетероаллельных по некоторым генам.
Заменяет
Секторность грибной колонии
Секторность грибной колонии

Парасексуальный (псевдополой) процесс – слияние гаплоидных ядер гетероаллельных по какому-то локусу
Парасексуальный (псевдополой) процесс – слияние гаплоидных ядер гетероаллельных по какому-то локусу

Анастомозы, происходящие между соседними гифами внутри колонии, армируют ее дают более
Анастомозы, происходящие между соседними гифами внутри колонии, армируют ее дают более
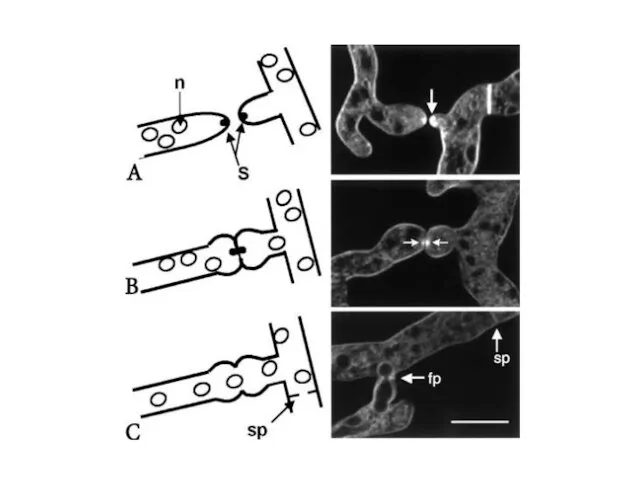
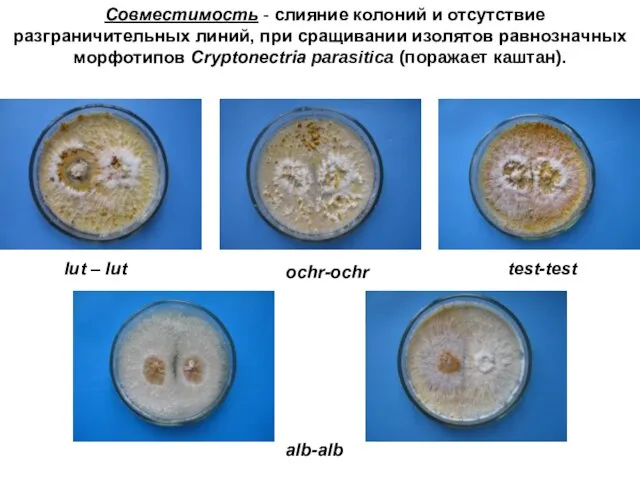
Совместимость - слияние колоний и отсутствие разграничительных линий, при сращивании
Совместимость - слияние колоний и отсутствие разграничительных линий, при сращивании
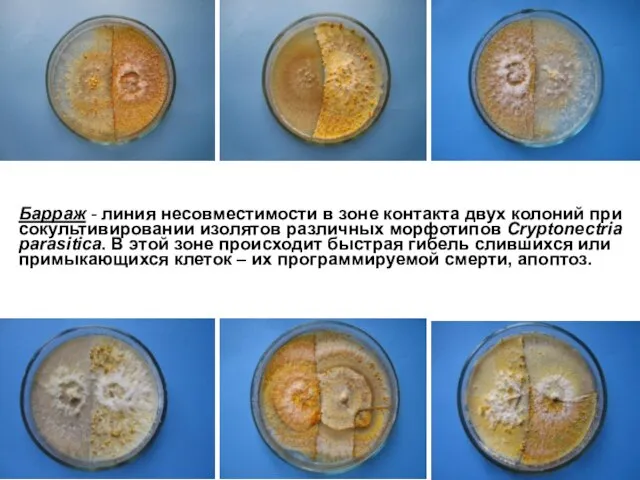
Барраж - линия несовместимости в зоне контакта двух колоний при сокультивировании
Барраж - линия несовместимости в зоне контакта двух колоний при сокультивировании

Вегетативная несовместимость – механизм защиты грибов от вирусной инфекции, так как
Вегетативная несовместимость – механизм защиты грибов от вирусной инфекции, так как

ОБЩАЯ ХАРАКТЕРИСТИКА ГРИБОВ
Углеводы клеточной стенки – хитин и глюканы (маннаны)
Синтез лизина
Углеводы клеточной стенки – хитин и глюканы (маннаны)
Синтез лизина

Что же дал филогенетический анализ для определения места грибов в системе
Что же дал филогенетический анализ для определения места грибов в системе
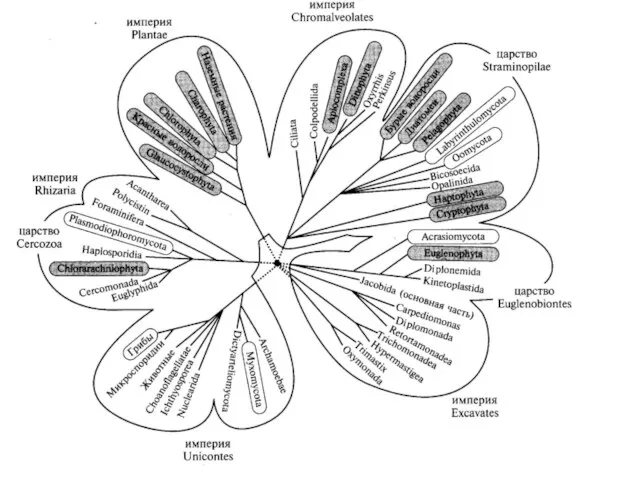

Организмы, изучаемые микологами, относятся к трем большим независимо эволюционирующим группам, которым
Организмы, изучаемые микологами, относятся к трем большим независимо эволюционирующим группам, которым
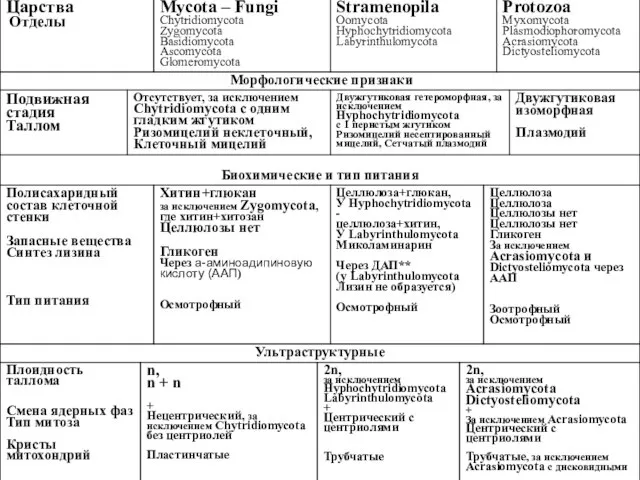
** – диаминопимелиновую кислоту
** – диаминопимелиновую кислоту
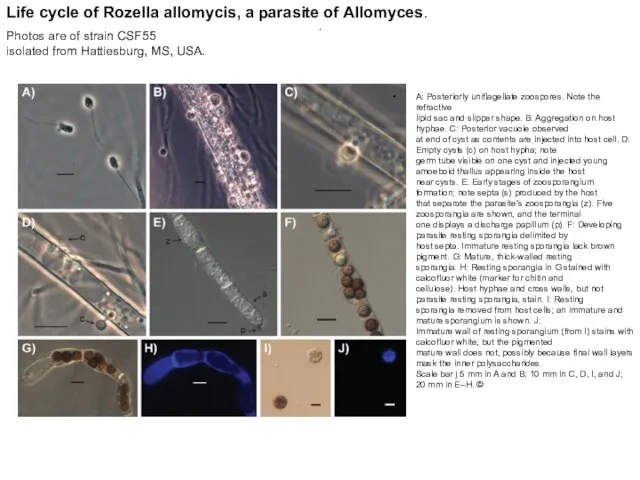
.
A: Posteriorly uniflagellate zoospores. Note the refractive
lipid sac and slipper shape.
.
A: Posteriorly uniflagellate zoospores. Note the refractive lipid sac and slipper shape.
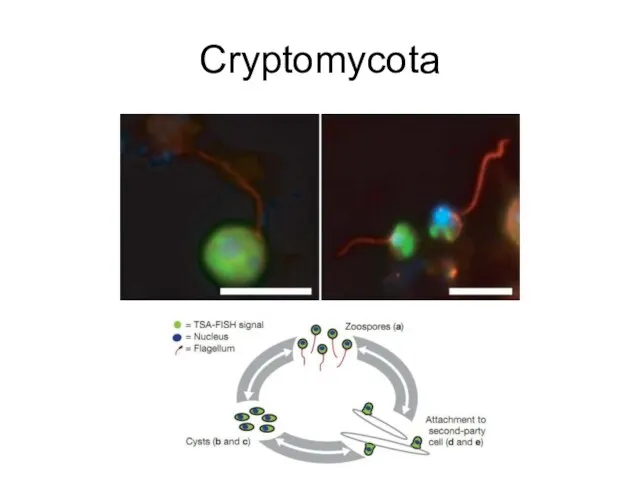
Cryptomycota
Cryptomycota
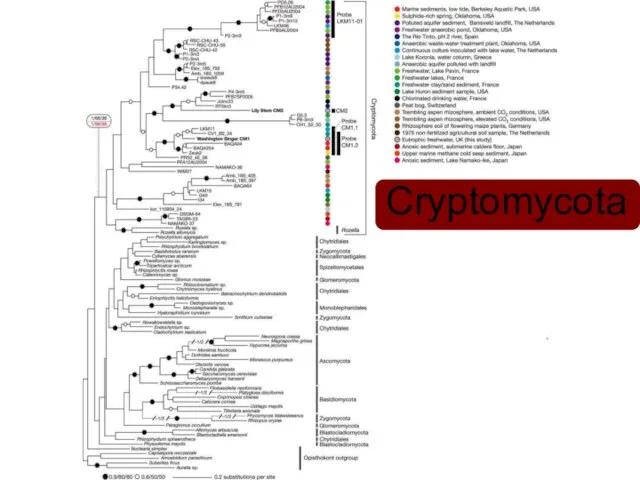
Cryptomycota
Cryptomycota
Cryptomycota
Cryptomycota
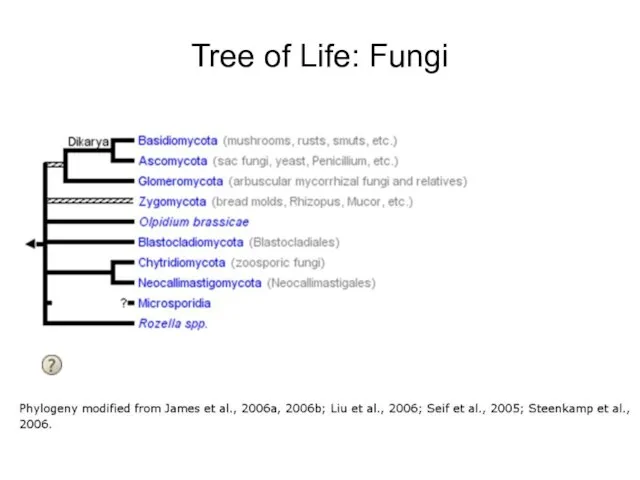
Tree of Life: Fungi
Tree of Life: Fungi
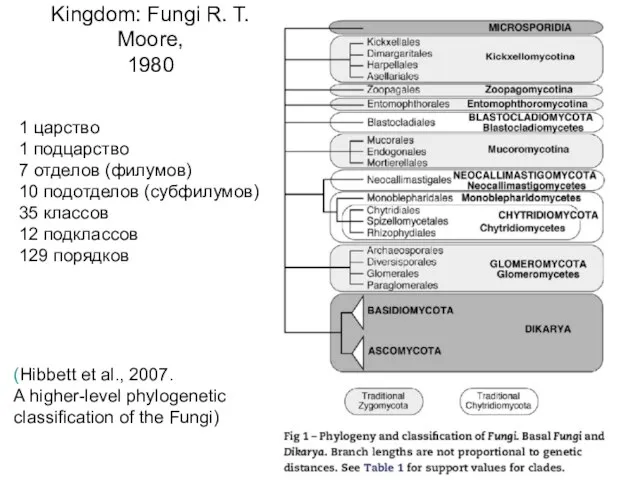
Kingdom: Fungi R. T. Moore,
1980
(Hibbett et al., 2007.
A higher-level
Kingdom: Fungi R. T. Moore,
1980
(Hibbett et al., 2007. A higher-level
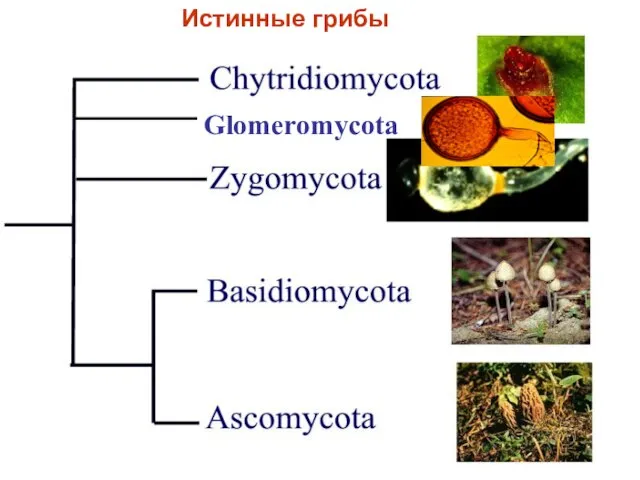
Glomeromycota
Glomeromycota
Истинные грибы
Glomeromycota
Glomeromycota
Истинные грибы

Иерархическая классификация
Regnum- Kingdom- Царство Fungi
Divisio- Phylum- Отдел Basidiomycota
Classis- Class-
Иерархическая классификация
Regnum- Kingdom- Царство Fungi
Divisio- Phylum- Отдел Basidiomycota
Classis- Class-

Agaricus bisporus (Lange) Imbach
Lange первым описал этот гриб как Coprinus bisporus
Imbach
Agaricus bisporus (Lange) Imbach
Lange первым описал этот гриб как Coprinus bisporus
Imbach
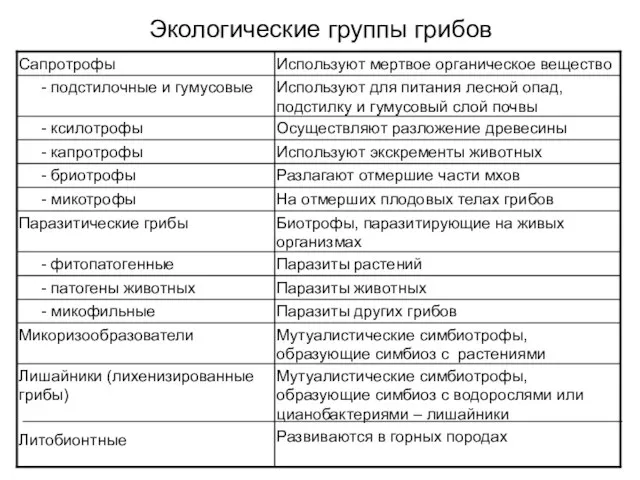
Экологические группы грибов
Экологические группы грибов

Сапротрофные грибы
Гифы грибов в почве в люминисцентном микроскопе
Посев из почвы на
Сапротрофные грибы
Гифы грибов в почве в люминисцентном микроскопе
Посев из почвы на

Ксилотрофные грибы – разрушители древесины
Домовой гриб (Serpula lacrimans)
Трутовые грибы (пор. Aphyllophorales)
Бурая
Ксилотрофные грибы – разрушители древесины
Домовой гриб (Serpula lacrimans)
Трутовые грибы (пор. Aphyllophorales)
Бурая

Фитопатогенные грибы
Фитопатология – одна из важнейших отраслей экологической и прикладной биологии
Раневые
Фитопатогенные грибы
Фитопатология – одна из важнейших отраслей экологической и прикладной биологии
Раневые

Микопаразитические грибы
Микопаразитические грибы, растущие на плодовых телах агариковых грибов
Микопаразитические грибы
Микопаразитические грибы, растущие на плодовых телах агариковых грибов

Грибы – паразиты животных
Энтомопатогенные грибы
Entomophthora (зигомицеты)
Beauveria bassiana
Энтомопатогенные грибы используются для производства
Грибы – паразиты животных
Энтомопатогенные грибы
Entomophthora (зигомицеты)
Beauveria bassiana
Энтомопатогенные грибы используются для производства
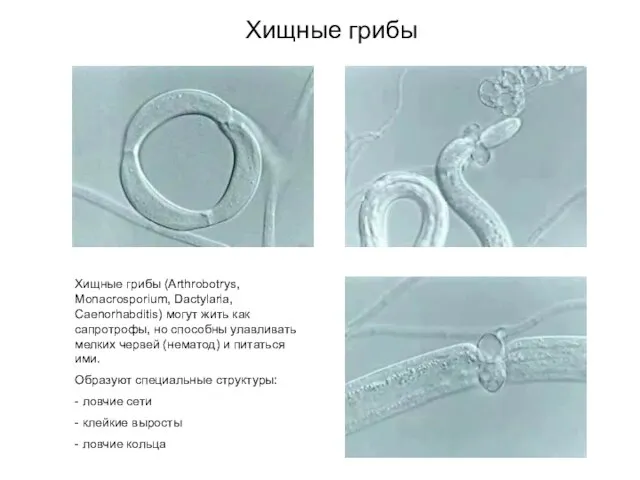
Хищные грибы
Хищные грибы (Arthrobotrys, Monacrosporium, Dactylaria, Caenorhabditis) могут жить как сапротрофы,
Хищные грибы
Хищные грибы (Arthrobotrys, Monacrosporium, Dactylaria, Caenorhabditis) могут жить как сапротрофы,

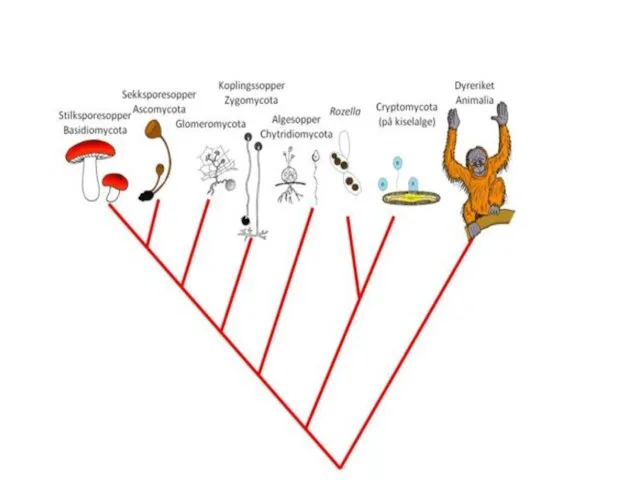
Царство Fungi (Eumycota)
Отделы:
Chytridiomycota
Образуют подвижные споры называемые зооспорами
Зигота, мейоз происходит в покоящемся
Царство Fungi (Eumycota)
Отделы:
Chytridiomycota
Образуют подвижные споры называемые зооспорами
Зигота, мейоз происходит в покоящемся

Какое количество видов грибов существует?
По разным источникам грибов от 100
Какое количество видов грибов существует?
По разным источникам грибов от 100

Оценка максимально возможного разнообразия грибов
Цветковых растений на Британских островах описано =
Оценка максимально возможного разнообразия грибов
Цветковых растений на Британских островах описано =
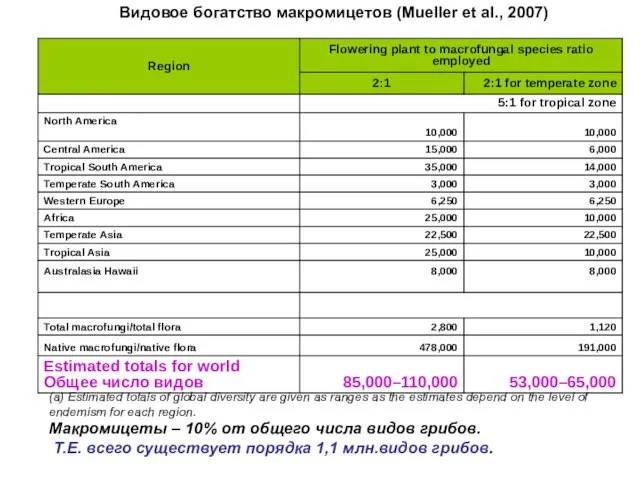
(a) Estimated totals of global diversity are given as ranges as
(a) Estimated totals of global diversity are given as ranges as

Потенциал исследований видового разнообразия грибов
Грибы в экстремальных местообитаниях
Обширные регионы не изучены
Потенциал исследований видового разнообразия грибов
Грибы в экстремальных местообитаниях
Обширные регионы не изучены
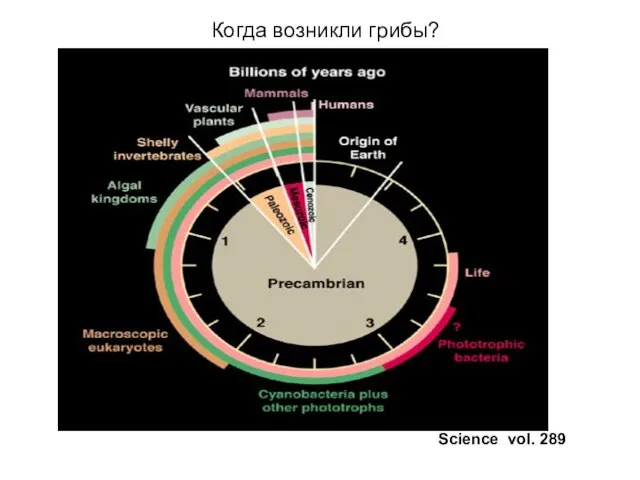
Когда возникли грибы?
Science vol. 289
Когда возникли грибы?
Science vol. 289

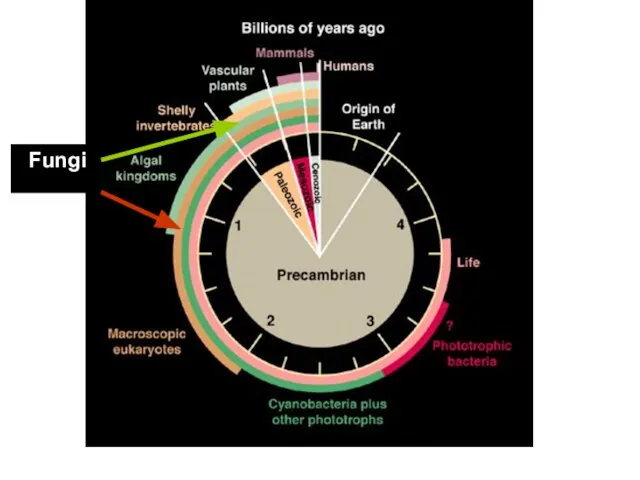
Грибы древние организмы
Многие виды грибов возникли 450 млн.лет назад, а, возможно,
Грибы древние организмы
Многие виды грибов возникли 450 млн.лет назад, а, возможно,
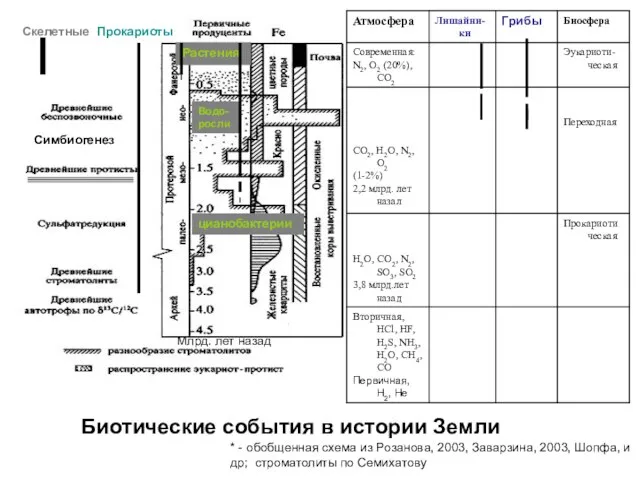
Скелетные
Биотические события в истории Земли
Симбиогенез
Прокариоты
цианобактерии
Растения
Водо-росли
Млрд. лет назад
* - обобщенная схема из
Скелетные
Биотические события в истории Земли
Симбиогенез
Прокариоты
цианобактерии
Растения
Водо-росли
Млрд. лет назад
* - обобщенная схема из

Berbee and Taylor, 2001
Расчеты на основе уточненной скорости замены нуклеотидных последовательностей
Berbee and Taylor, 2001
Расчеты на основе уточненной скорости замены нуклеотидных последовательностей
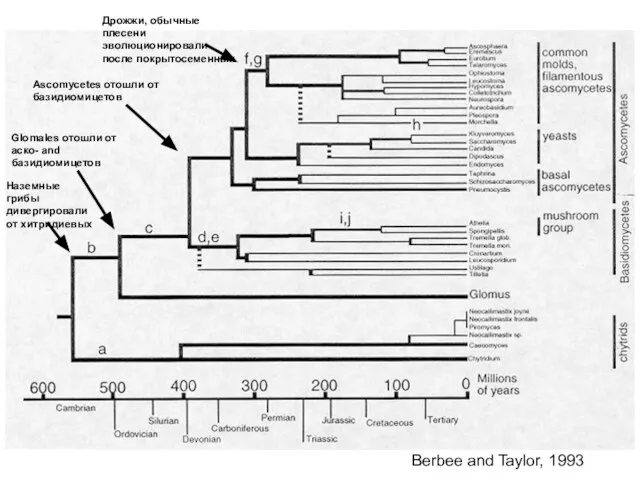
Berbee and Taylor, 1993
Ascomycetes отошли от базидиомицетов
Дрожжи, обычные плесени эволюционировали после
Berbee and Taylor, 1993
Ascomycetes отошли от базидиомицетов
Дрожжи, обычные плесени эволюционировали после

Heckman et al. (2001). Molecular Evidence for the Early Colonization of
Heckman et al. (2001). Molecular Evidence for the Early Colonization of

Грибы, из водоемов, стали переходить к наземной жизни около 1,3 млрд.
Грибы, из водоемов, стали переходить к наземной жизни около 1,3 млрд.
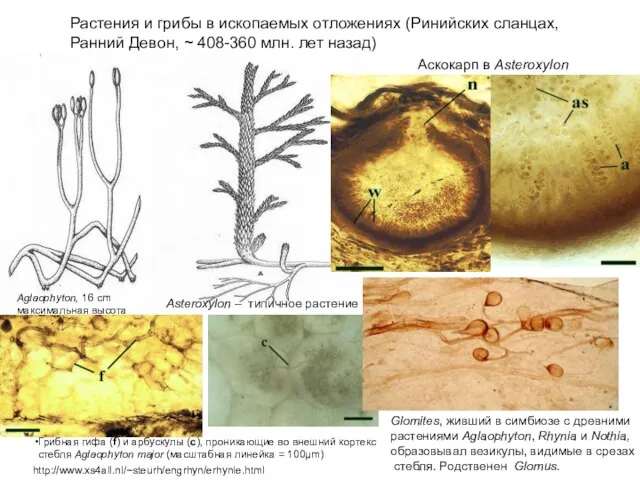
Aскокaрп в Asteroxylon
http://www.xs4all.nl/~steurh/engrhyn/erhynie.html
Растения и грибы в ископаемых отложениях (Ринийских сланцах, Ранний
Aскокaрп в Asteroxylon
http://www.xs4all.nl/~steurh/engrhyn/erhynie.html
Растения и грибы в ископаемых отложениях (Ринийских сланцах, Ранний
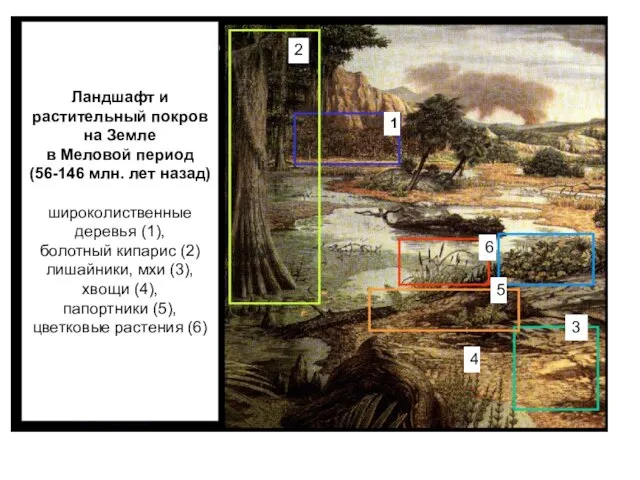
Ландшафт и
растительный покров
на Земле
в Меловой период
(56-146 млн.
Ландшафт и
растительный покров
на Земле
в Меловой период
(56-146 млн.
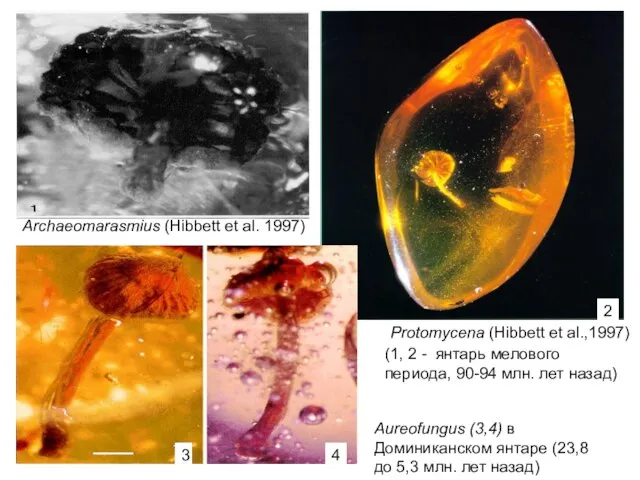
Archaeomarasmius (Hibbett et al. 1997)
Protomycena (Hibbett et al.,1997)
Aureofungus (3,4) в
Archaeomarasmius (Hibbett et al. 1997)
Protomycena (Hibbett et al.,1997)
Aureofungus (3,4) в
Fungi
Fungi

Продолжительность жизни грибов
От нескольких суток до тысячилетий.
Продолжительность жизни грибов
От нескольких суток до тысячилетий.
Л-2
Л-2

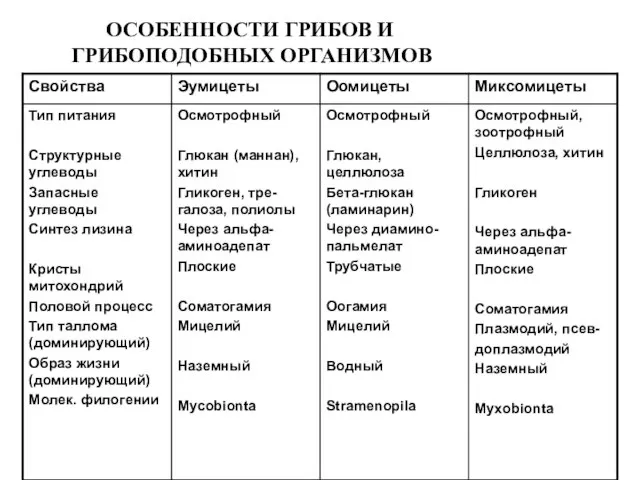
ОСОБЕННОСТИ ГРИБОВ И
ГРИБОПОДОБНЫХ ОРГАНИЗМОВ
ОСОБЕННОСТИ ГРИБОВ И
ГРИБОПОДОБНЫХ ОРГАНИЗМОВ

CНROMISTA
Hyphochytriomycota
Labyrinthulomycota
Oomycota
FUNGI
Ascomycota
CНROMISTA
Hyphochytriomycota
Labyrinthulomycota
Oomycota
FUNGI
Ascomycota

Kingdom: Fungi R. T. Moore,
1980
(Hibbett et al., 2007.
A higher-level
Kingdom: Fungi R. T. Moore,
1980
(Hibbett et al., 2007. A higher-level
 Типы тканей
Типы тканей Тип Моллюски или Мягкотелые
Тип Моллюски или Мягкотелые Игра Счастливый случай
Игра Счастливый случай Отряд ластоногие
Отряд ластоногие Животные Курской области
Животные Курской области Теория эволюции Откуда берутся новые формы живых систем?
Теория эволюции Откуда берутся новые формы живых систем?  Домашние растения Невидимые труженики
Домашние растения Невидимые труженики Тип Хордовые. Подтип Бесчерепные (7 класс)
Тип Хордовые. Подтип Бесчерепные (7 класс) Класс Двудольные
Класс Двудольные Соглашение о сохранении белых медведей 1973 года
Соглашение о сохранении белых медведей 1973 года 2.3. Матричные синтезы. Часть 1
2.3. Матричные синтезы. Часть 1 Нутрия
Нутрия Образование половых клеток. Мейоз
Образование половых клеток. Мейоз Происхождение жизни. Источники информации и логика рассуждений
Происхождение жизни. Источники информации и логика рассуждений Значение воды в жизнедеятельности растений Автор: Ерохнович Ростислав Школа № 42 класс 6 Г Учитель: Козлова В. М.
Значение воды в жизнедеятельности растений Автор: Ерохнович Ростислав Школа № 42 класс 6 Г Учитель: Козлова В. М. Мезофиты
Мезофиты Порода хаски
Порода хаски 05.1 Стрекающие
05.1 Стрекающие Schönheit der Natur. Durch Musik untermalt
Schönheit der Natur. Durch Musik untermalt Презентация на тему "Селекция растений" - скачать презентации по Биологии
Презентация на тему "Селекция растений" - скачать презентации по Биологии Зимующие птицы. Снегири: самец и самка
Зимующие птицы. Снегири: самец и самка Строение и работа сердца
Строение и работа сердца Цитоскелет
Цитоскелет Покормите птиц зимой
Покормите птиц зимой Тема урока: «Способы добывания пищи» разработка Жарковой Валентины Петровны МОУ СОШ №1 р.п. Лысые Горы Саратовской области
Тема урока: «Способы добывания пищи» разработка Жарковой Валентины Петровны МОУ СОШ №1 р.п. Лысые Горы Саратовской области  Физиология мышечного аппарата. (Лекция 3)
Физиология мышечного аппарата. (Лекция 3) Семейство Губоцветные. Смолина Ольга Викторовна Учитель биологии МБОУ СОШ №5, г. Торжок
Семейство Губоцветные. Смолина Ольга Викторовна Учитель биологии МБОУ СОШ №5, г. Торжок Умей сказать нет Мельникова Галина Викторовна МОУ Янгельская СОШ Им. Филатова А.К.
Умей сказать нет Мельникова Галина Викторовна МОУ Янгельская СОШ Им. Филатова А.К.