- Митохондрии, пероксисомы и пластиды

Содержание
- 2. Митохондриальный геном растений Геном митохондрий растений значительно больше генома животных митохондрий. Например, у арабидопсиса в 20
- 3. Свобода или беспорядок? Контроль ядра? Частые рекомбинации приводят к существованию мтх генома в форме множества колец
- 4. Пластидный геном гены организованы в форме оперонов есть интроны кольцевая хромосома около 100 генов размер: от
- 5. Митохондрии… что интересно? Нестабильный геном вследствие постоянных рекомбинаций Постоянное взаимодействие с ядром: импорт и… экспорт? Цитоплазматическая
- 6. Про мембраны митохондрий Внешняя мембрана. Проницаема для веществ с молекулярной массой до 10 000 через порины
- 7. Как белки импортируются в мтх С обеих сторон участвуют шапероны (HSP70) Транслоказы обеспечивают перетаскивание полипептидов На
- 8. РНК полимеразы в свете эволюции Наличие «фаговых» РНК полимераз с ядерным кодированием демонстрирует контроль ядра над
- 9. РНК-полимеразы пластид Гены, которые обслуживает ядерная РНК-полимераза. Гены, имеющие нестандартные промоторы. Такие промоторы свойственны лишь немногим
- 10. Кто регулирует АФК? Митохондрии являются сенсорами редокс-статуса клетки Митохондрии - одно из основных мест образования АФК
- 11. Митохондрии динамичны и «общительны» Считается, что «нестехиометричность» генома (т.е. его неравномерное распределение между митохондриями) может служить
- 12. Деление митохондрий Митохондрии делятся – факт. Это наблюдали многие. Деление происходит по «бактериальному сценарию», однако… …этот
- 13. Анализ мутантов показал… В делении участвуют динамины – семейство ГТФаз с многочисленными функциями типа «слияния и
- 14. Мутанты по делению В фенотипе наблюдаем либо увеличение размеров митохондрий, либо слияние их в сеть… David
- 15. Мтх в интерфазе Сканирующая лазерная микроскопия (CSLM) и 3D-СЭМ позволили наблюдать динамику хондриома в клеточном цикле
- 16. José M. Seguí-Simarro et al. Plant Physiol. 2008;148:1380 ©2008 by American Society of Plant Biologists Митохондрии
- 17. Зачем так сложно? Сейчас предполагают, что сложная динамика хондриома тесно связана с «геномной свободой», т.е. слияние
- 18. Митохондрии и свет МТХ в ответ на свет движутся вместе с хлоропластами. Они с ними связаны?
- 19. Подвижность мтх Митохондрии очень подвижны, передвигаются в основном по МФ (миозины). Средняя скорость около 0,5 µм/с,
- 20. Митохондрии-убийцы У животных и грибов мтх активно участвуют в апоптозе У растений канонического апоптоза нет, но
- 21. Li, Xing J. Exp. Bot. 2011.62(1):331-43 Митохондрии-убийцы AsA – аскорбиновая к-та
- 22. Пероксисомы – загадочные и отважные Пероксисомы – одномембранные органеллы с диаметром 0,1-1 µм. Основные функции: липидный
- 23. Маленькие и сложные В пероксисомах много оксидаз (↓), генерирующих H2O2, Ликвидация АФК с помощью каталазы и
- 24. Размножение пероксисом Могут образовываться de novo из субдоменов ЭПР, но также размножаются делением Pex11p обеспечивает удлинение,
- 25. Сходство или совпадение? Почему механизмы деления пероксисом и митохондрий так похожи? Ведь эти органеллы не родственны.
- 26. И пластиды тоже… Недавно был открыт DRP5 Это динамин, участник деления пластид и пероксисом, специфичный для
- 27. Пластиды. Начало. Самыми первыми были цианобактерии… Первыми пластидами были хлоропласты. У одноклеточных водорослей только они и
- 28. Фотосинтез Гравитропизм Восстановление неорганических субстратов (NO2) Окрашивание каротиноидами Запас крахмала Запас железа (фитоферритин) Синтез изопреноидов Пластидная
- 29. Хлоропласты Типичные хлоропласты линзовидной формы, 5-10 µм в диаметре, 2-4 µм в толщину. Типичная клетка листа
- 30. Хлоропласты Люмен тилакоидов образует единое пространство, топологически эквивалентное ММП Тилакоиды собираются в граны ФС II локализуется
- 31. ДНК-динамика Количество копий ДНК в зрелых хлоропластах очень велико. Каждый нуклеоид содержит в среднем 10 копий
- 32. Ещё немного о РНК-полимеразах Начало дифференцировки пропластиды в хлоропласт сопровождается активацией ядерной РНК-полимеразы (NEP) Она запускает
- 33. Другие пластиды «Пластида» – от слова «пластичность» Пропластиды в 10-100 раз меньше хлоропластов, и в клетке
- 34. Этиопласты и их будущее Этиопласты накапливают липиды для быстрой сборки тилакоидов в форме проламеллярных тел (75%
- 35. Лейкопласты Лейкопласты - бесцветные пластиды, вовлеченные в синтез изопреноидов, прежде всего моно- и сесквитерпенов, содержащихся в
- 36. Портретная галерея
- 37. Обмен информацией Поток информации от ядра к х/п называют антеградным, и он включает в себя как
- 38. Ретроградный сигналинг Сигнал о перевосстановленности цепи снижает транскрипцию генов ФС и повышает – генов антиоксидантных систем
- 39. Импорт белков
- 40. Двойное кодирование Большинство белковых комплексов х/п имеют двойное кодирование Самый известный пример – Рубиско. Большие субъединицы
- 42. Скачать презентацию

Митохондриальный геном растений
Геном митохондрий растений значительно больше генома животных митохондрий. Например,
Митохондриальный геном растений
Геном митохондрий растений значительно больше генома животных митохондрий. Например,
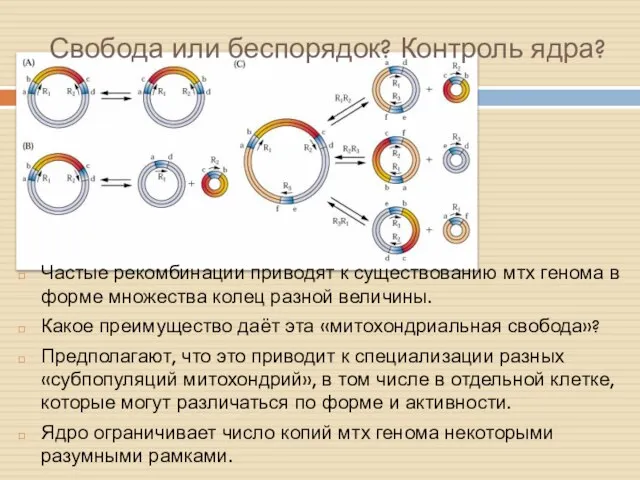
Свобода или беспорядок? Контроль ядра?
Частые рекомбинации приводят к существованию мтх генома
Свобода или беспорядок? Контроль ядра?
Частые рекомбинации приводят к существованию мтх генома

Пластидный геном
гены организованы в форме оперонов
есть интроны
кольцевая хромосома
Пластидный геном
гены организованы в форме оперонов
есть интроны
кольцевая хромосома

Митохондрии… что интересно?
Нестабильный геном вследствие постоянных рекомбинаций
Постоянное взаимодействие с ядром: импорт
Митохондрии… что интересно?
Нестабильный геном вследствие постоянных рекомбинаций
Постоянное взаимодействие с ядром: импорт

Про мембраны митохондрий
Внешняя мембрана. Проницаема для веществ с молекулярной массой до
Про мембраны митохондрий
Внешняя мембрана. Проницаема для веществ с молекулярной массой до
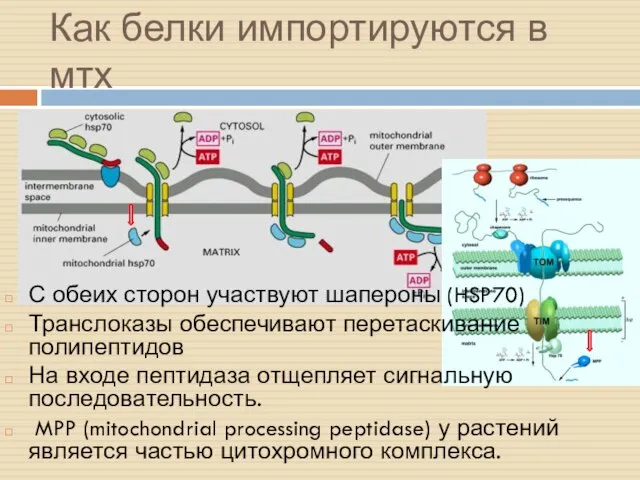
Как белки импортируются в мтх
С обеих сторон участвуют шапероны (HSP70)
Транслоказы обеспечивают
Как белки импортируются в мтх
С обеих сторон участвуют шапероны (HSP70)
Транслоказы обеспечивают
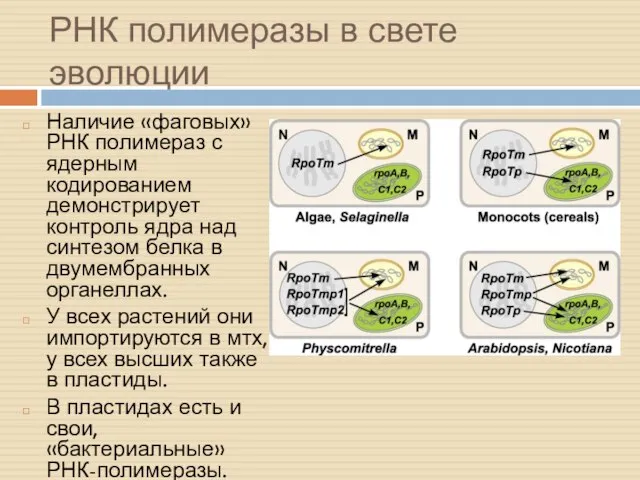
РНК полимеразы в свете эволюции
Наличие «фаговых» РНК полимераз с ядерным кодированием
РНК полимеразы в свете эволюции
Наличие «фаговых» РНК полимераз с ядерным кодированием

РНК-полимеразы пластид
Гены, которые обслуживает ядерная РНК-полимераза.
Гены, имеющие нестандартные промоторы. Такие промоторы
РНК-полимеразы пластид
Гены, которые обслуживает ядерная РНК-полимераза.
Гены, имеющие нестандартные промоторы. Такие промоторы

Кто регулирует АФК?
Митохондрии являются сенсорами редокс-статуса клетки
Митохондрии - одно из основных
Кто регулирует АФК?
Митохондрии являются сенсорами редокс-статуса клетки
Митохондрии - одно из основных

Митохондрии динамичны и «общительны»
Считается, что «нестехиометричность» генома (т.е. его неравномерное распределение
Митохондрии динамичны и «общительны»
Считается, что «нестехиометричность» генома (т.е. его неравномерное распределение
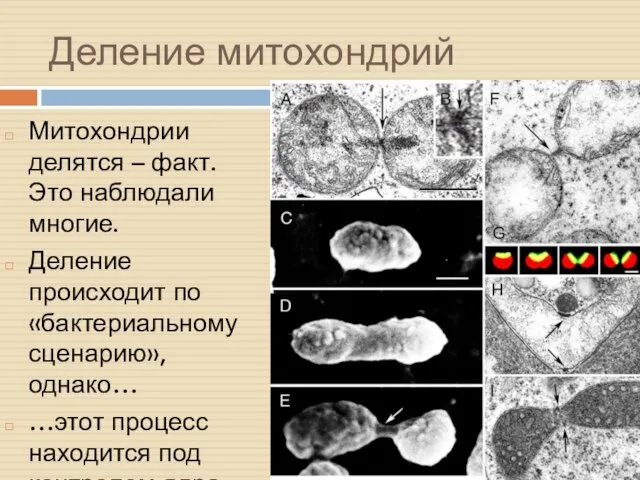
Деление митохондрий
Митохондрии делятся – факт. Это наблюдали многие.
Деление происходит по «бактериальному
Деление митохондрий
Митохондрии делятся – факт. Это наблюдали многие.
Деление происходит по «бактериальному
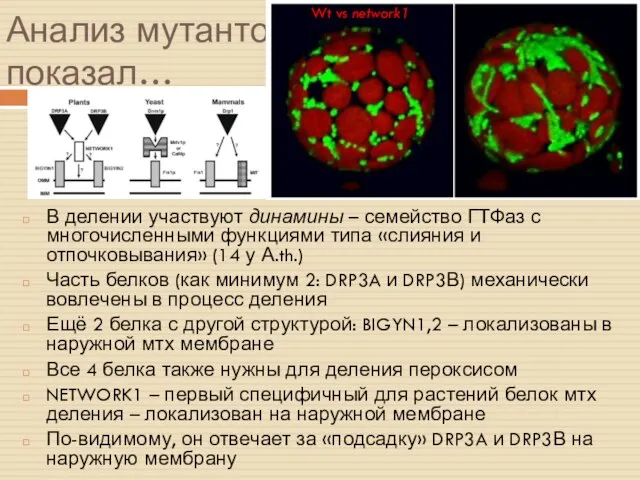
Анализ мутантов показал…
В делении участвуют динамины – семейство ГТФаз с многочисленными
Анализ мутантов показал…
В делении участвуют динамины – семейство ГТФаз с многочисленными

Мутанты по делению
В фенотипе наблюдаем либо увеличение размеров митохондрий, либо слияние
Мутанты по делению
В фенотипе наблюдаем либо увеличение размеров митохондрий, либо слияние

Мтх в интерфазе
Сканирующая лазерная микроскопия (CSLM) и 3D-СЭМ позволили наблюдать динамику
Мтх в интерфазе
Сканирующая лазерная микроскопия (CSLM) и 3D-СЭМ позволили наблюдать динамику
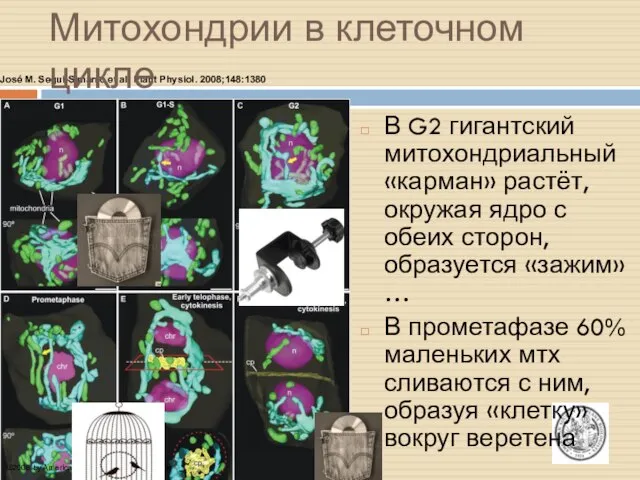
José M. Seguí-Simarro et al. Plant Physiol. 2008;148:1380
©2008 by American Society
José M. Seguí-Simarro et al. Plant Physiol. 2008;148:1380
©2008 by American Society
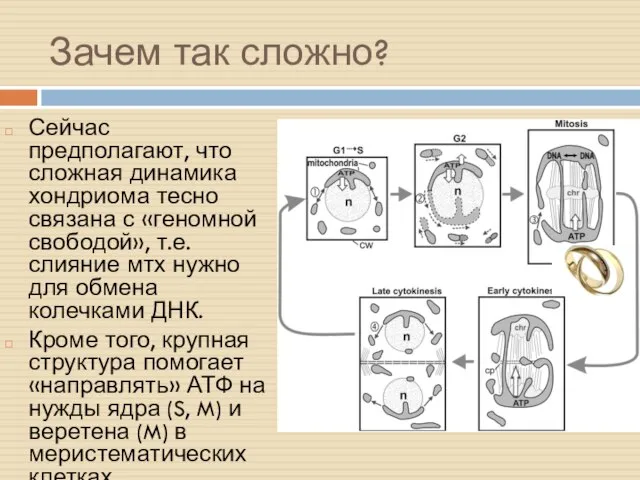
Зачем так сложно?
Сейчас предполагают, что сложная динамика хондриома тесно связана с
Зачем так сложно?
Сейчас предполагают, что сложная динамика хондриома тесно связана с
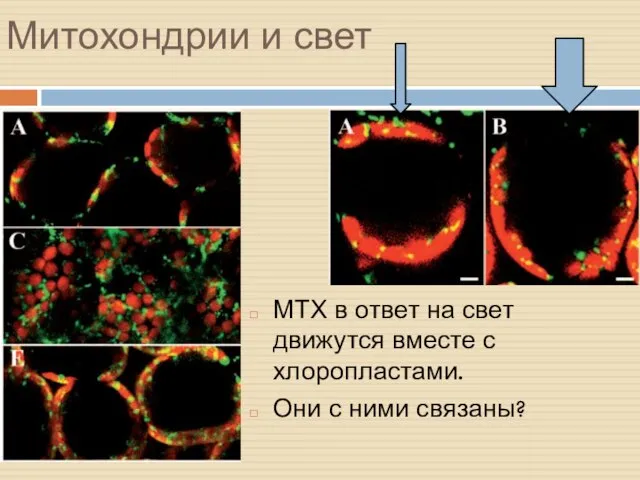
Митохондрии и свет
МТХ в ответ на свет движутся вместе с хлоропластами.
Митохондрии и свет
МТХ в ответ на свет движутся вместе с хлоропластами.

Подвижность мтх
Митохондрии очень подвижны, передвигаются в основном по МФ (миозины).
Средняя скорость
Подвижность мтх
Митохондрии очень подвижны, передвигаются в основном по МФ (миозины).
Средняя скорость
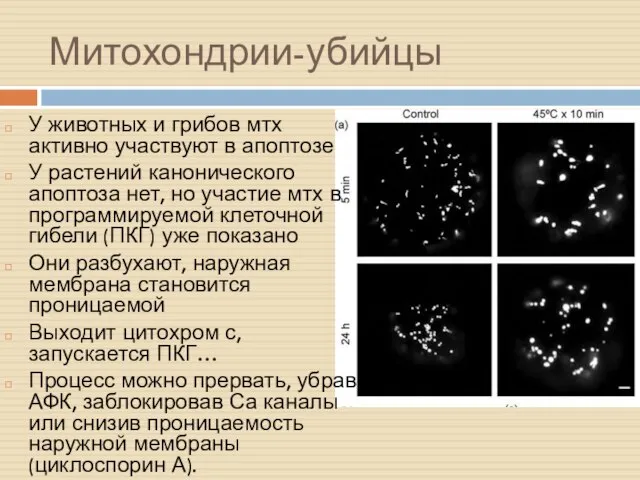
Митохондрии-убийцы
У животных и грибов мтх активно участвуют в апоптозе
У растений канонического
Митохондрии-убийцы
У животных и грибов мтх активно участвуют в апоптозе
У растений канонического
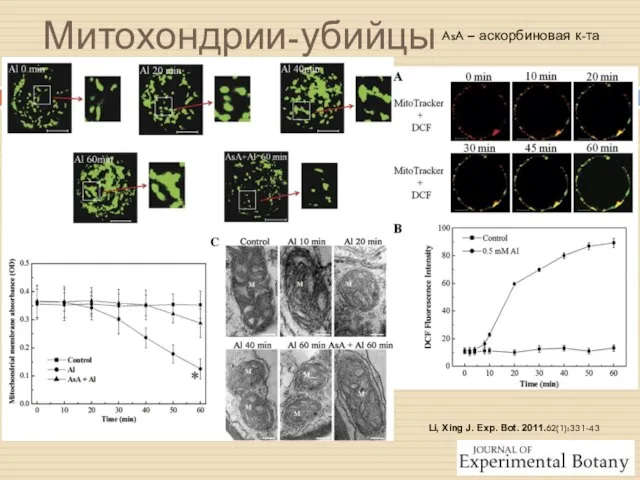
Li, Xing J. Exp. Bot. 2011.62(1):331-43
Митохондрии-убийцы
AsA – аскорбиновая к-та
Li, Xing J. Exp. Bot. 2011.62(1):331-43
Митохондрии-убийцы
AsA – аскорбиновая к-та

Пероксисомы – загадочные и отважные
Пероксисомы – одномембранные органеллы с диаметром 0,1-1
Пероксисомы – загадочные и отважные
Пероксисомы – одномембранные органеллы с диаметром 0,1-1
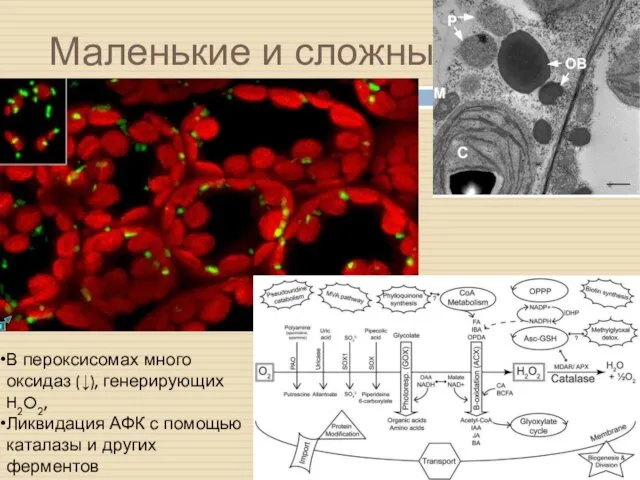
Маленькие и сложные
В пероксисомах много оксидаз (↓), генерирующих H2O2,
Ликвидация АФК с
Маленькие и сложные
В пероксисомах много оксидаз (↓), генерирующих H2O2,
Ликвидация АФК с

Размножение пероксисом
Могут образовываться de novo из субдоменов ЭПР, но также размножаются
Размножение пероксисом
Могут образовываться de novo из субдоменов ЭПР, но также размножаются

Сходство или совпадение?
Почему механизмы деления пероксисом и митохондрий так похожи? Ведь
Сходство или совпадение?
Почему механизмы деления пероксисом и митохондрий так похожи? Ведь
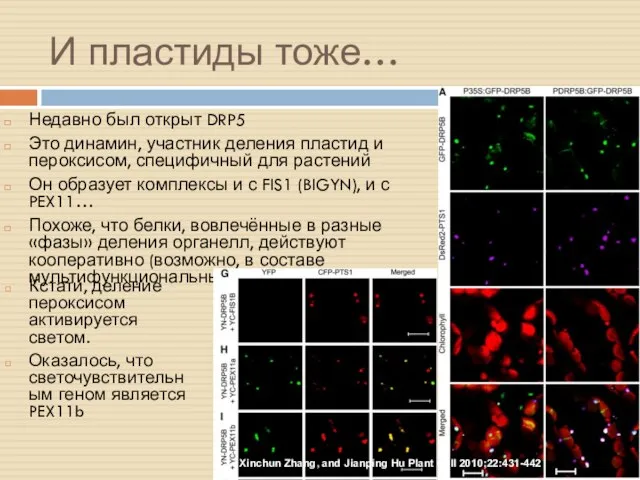
И пластиды тоже…
Недавно был открыт DRP5
Это динамин, участник деления пластид и
И пластиды тоже…
Недавно был открыт DRP5
Это динамин, участник деления пластид и
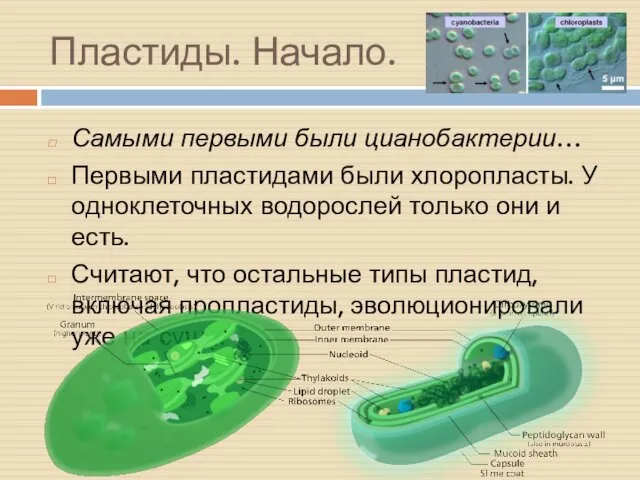
Пластиды. Начало.
Самыми первыми были цианобактерии…
Первыми пластидами были хлоропласты. У одноклеточных водорослей
Пластиды. Начало.
Самыми первыми были цианобактерии…
Первыми пластидами были хлоропласты. У одноклеточных водорослей

Фотосинтез
Гравитропизм
Восстановление неорганических субстратов (NO2)
Окрашивание каротиноидами
Запас крахмала
Запас железа (фитоферритин)
Синтез изопреноидов
Пластидная система
Фотосинтез
Гравитропизм
Восстановление неорганических субстратов (NO2)
Окрашивание каротиноидами
Запас крахмала
Запас железа (фитоферритин)
Синтез изопреноидов
Пластидная система
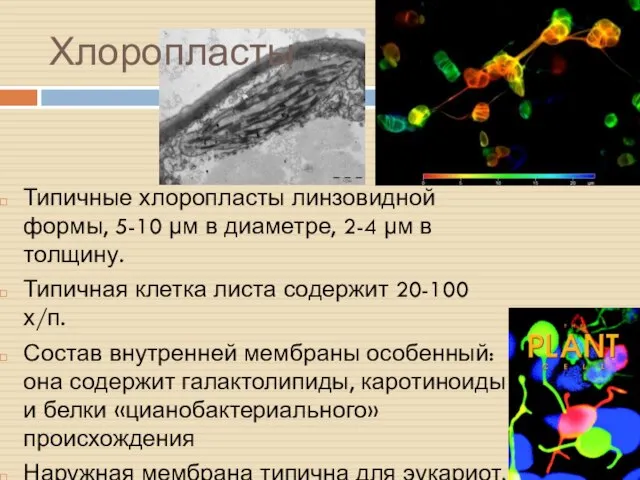
Хлоропласты
Типичные хлоропласты линзовидной формы, 5-10 µм в диаметре, 2-4 µм в
Хлоропласты
Типичные хлоропласты линзовидной формы, 5-10 µм в диаметре, 2-4 µм в

Хлоропласты
Люмен тилакоидов образует единое пространство, топологически эквивалентное ММП
Тилакоиды собираются в граны
ФС
Хлоропласты
Люмен тилакоидов образует единое пространство, топологически эквивалентное ММП
Тилакоиды собираются в граны
ФС
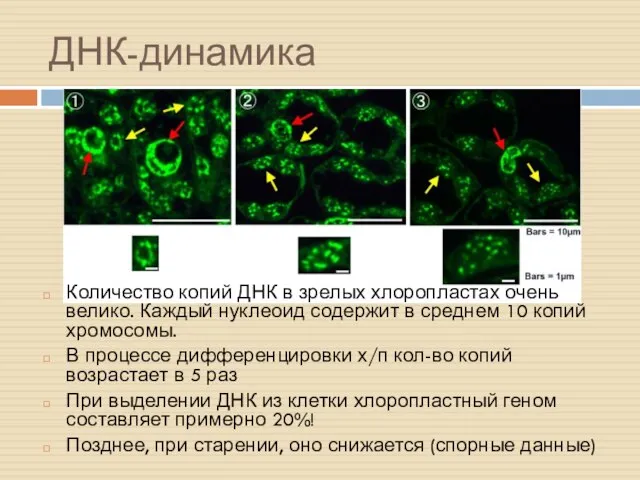
ДНК-динамика
Количество копий ДНК в зрелых хлоропластах очень велико. Каждый нуклеоид содержит
ДНК-динамика
Количество копий ДНК в зрелых хлоропластах очень велико. Каждый нуклеоид содержит

Ещё немного о РНК-полимеразах
Начало дифференцировки пропластиды в хлоропласт сопровождается активацией ядерной
Ещё немного о РНК-полимеразах
Начало дифференцировки пропластиды в хлоропласт сопровождается активацией ядерной

Другие пластиды
«Пластида» – от слова «пластичность»
Пропластиды в 10-100 раз меньше хлоропластов,
Другие пластиды
«Пластида» – от слова «пластичность»
Пропластиды в 10-100 раз меньше хлоропластов,
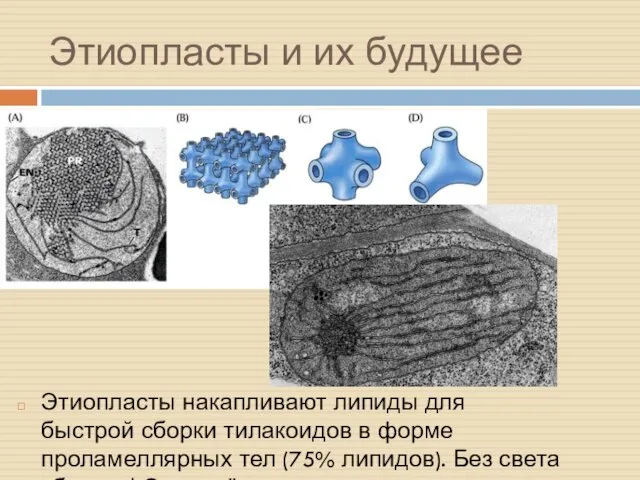
Этиопласты и их будущее
Этиопласты накапливают липиды для быстрой сборки тилакоидов в
Этиопласты и их будущее
Этиопласты накапливают липиды для быстрой сборки тилакоидов в

Лейкопласты
Лейкопласты - бесцветные пластиды, вовлеченные в синтез изопреноидов, прежде всего моно-
Лейкопласты
Лейкопласты - бесцветные пластиды, вовлеченные в синтез изопреноидов, прежде всего моно-
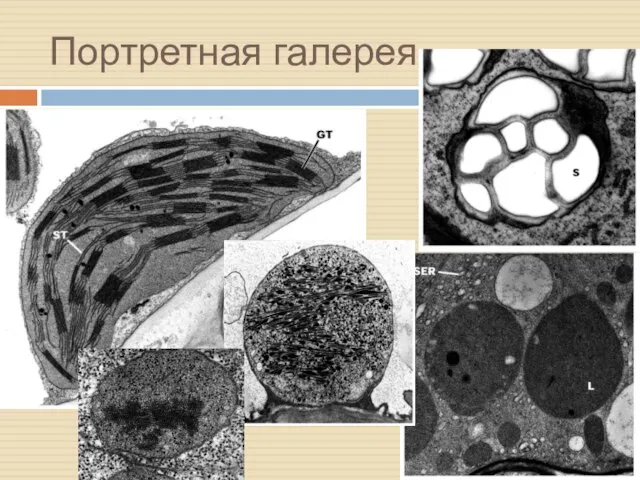
Портретная галерея
Портретная галерея

Обмен информацией
Поток информации от ядра к х/п называют антеградным, и он
Обмен информацией
Поток информации от ядра к х/п называют антеградным, и он
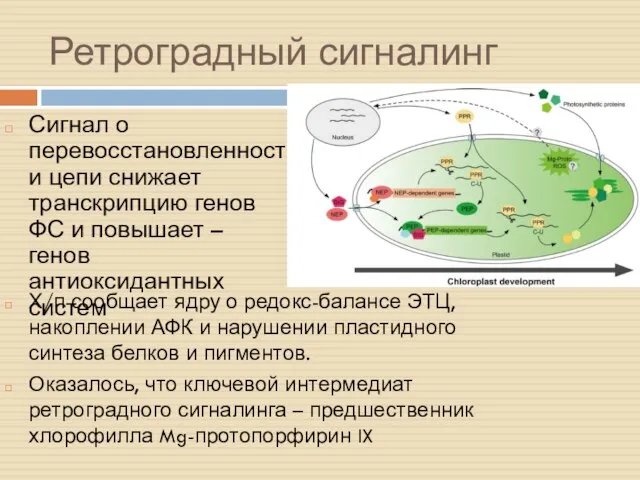
Ретроградный сигналинг
Сигнал о перевосстановленности цепи снижает транскрипцию генов ФС и повышает
Ретроградный сигналинг
Сигнал о перевосстановленности цепи снижает транскрипцию генов ФС и повышает
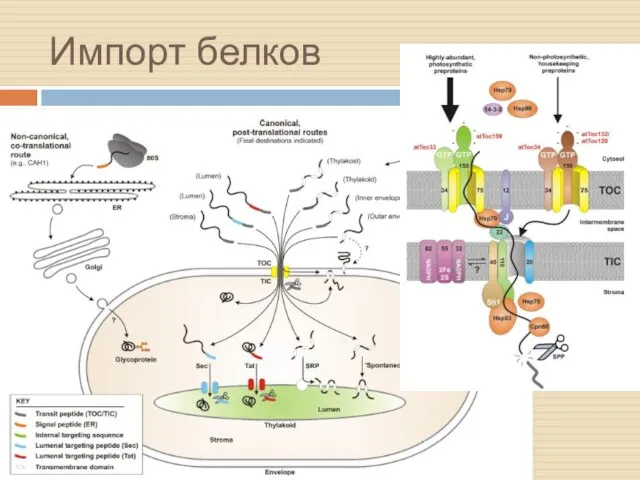
Импорт белков
Импорт белков
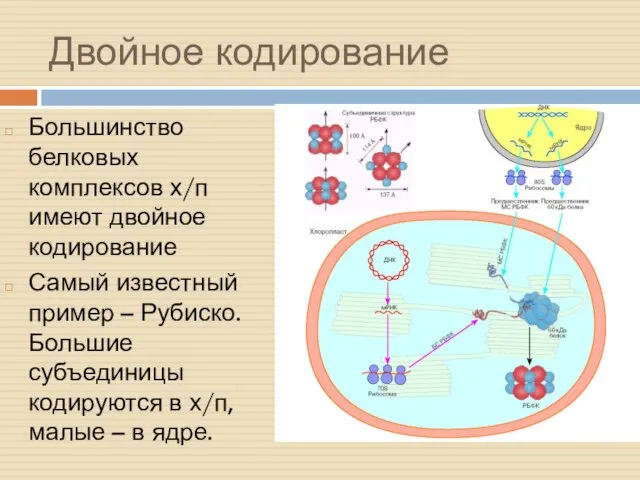
Двойное кодирование
Большинство белковых комплексов х/п имеют двойное кодирование
Самый известный пример –
Двойное кодирование
Большинство белковых комплексов х/п имеют двойное кодирование
Самый известный пример –
 Законы генетики
Законы генетики Нейрофизиологические основы поведения. Классификация форм научения
Нейрофизиологические основы поведения. Классификация форм научения Методы цитологии. Клеточная теория. Особенности химического состава клетки
Методы цитологии. Клеточная теория. Особенности химического состава клетки Физиология микроорганизмов
Физиология микроорганизмов Кириллова И.И. идентификатор : 248-130-269 Кириллова И.И. идентификатор : 248-130-269 Тема урока: «Ступеньки эволюционных идей»
Кириллова И.И. идентификатор : 248-130-269 Кириллова И.И. идентификатор : 248-130-269 Тема урока: «Ступеньки эволюционных идей»  Обрезка плодовых деревьев. Лекция 16
Обрезка плодовых деревьев. Лекция 16 Своя игра. (10 класс)
Своя игра. (10 класс) Сімейство котячих
Сімейство котячих Генотип - цілісна система. Цитоплазматична спадковість
Генотип - цілісна система. Цитоплазматична спадковість Класс насекомые. Отряд блохи
Класс насекомые. Отряд блохи Эволюция человека Подготовил учитель биологии высшей квалификационной категории Пету
Эволюция человека Подготовил учитель биологии высшей квалификационной категории Пету Кар Линней
Кар Линней Уровни организации живой природы
Уровни организации живой природы Клеточный уровень жизни Изучить структуру клеток Механизм устойчивости клетки как биосистемы Клетка –элементарная структур
Клеточный уровень жизни Изучить структуру клеток Механизм устойчивости клетки как биосистемы Клетка –элементарная структур Функциональная анатомия спинного и головного мозга
Функциональная анатомия спинного и головного мозга Болезни аквариумных рыб Выполнил: ученик 9 а класса Волкотрубенко Михаил.
Болезни аквариумных рыб Выполнил: ученик 9 а класса Волкотрубенко Михаил. Мир моря. Обитатели. Фото
Мир моря. Обитатели. Фото Аттестационная работа. Методическая разработка проекта Энергозатраты человека и пищевой рацион
Аттестационная работа. Методическая разработка проекта Энергозатраты человека и пищевой рацион Пищевые вещества
Пищевые вещества Уровни структурной организации организма человека. Организм человека как единое целое. Общие закономерности роста и развити
Уровни структурной организации организма человека. Организм человека как единое целое. Общие закономерности роста и развити Презентация урока Дыхание растений Выполнила: Задворова Л. А. – учитель биологии школы № 13
Презентация урока Дыхание растений Выполнила: Задворова Л. А. – учитель биологии школы № 13 Обмороження Виконала учениця 11-А класу Ковальова Анастасія
Обмороження Виконала учениця 11-А класу Ковальова Анастасія  Презентация Природные ресурсы и охрана природы
Презентация Природные ресурсы и охрана природы Внутреннее строение птицы Презентация к уроку биологии Отряскиной Т.А.
Внутреннее строение птицы Презентация к уроку биологии Отряскиной Т.А.  Популяция, как население и как интегрированная биологическая система
Популяция, как население и как интегрированная биологическая система Орган зрения - глаз
Орган зрения - глаз Отряд Бабочки или Чешуекрылые
Отряд Бабочки или Чешуекрылые Высшая Нервная Деятельность
Высшая Нервная Деятельность