- Нейрогенез (у высших животных: ЦНС - нервная трубка периферическая, НС - нервный гребень и плакоды)

Содержание
- 2. Основные производные эктодермального зародышевого листка позвоночных
- 3. Основные составлящие нервной системы
- 4. Нейроны – биполярные клетки, аксон растет от тела клетки
- 5. Глиальные клетки позвоночных изолируют аксоны, содержат миелиновые мебраны, обеспечивая передачу электрических импульсов от тела нейрона к
- 6. В течение развития нейральные стволовые клетки дают начало нейронам ЦНС Они также – источник всех типов
- 7. НЕЙРУЛЯЦИЯ 1. Начальная стадия нейрогенеза у позвоночных - нейруляция. У позвоночных участок эктодермы специфицируется как нейроэктодерма
- 8. Нейруляция у амфибий (первичная)
- 9. Нейроглия дрозофилы Edenfeld et al. 2005, Curr.Opin. Neuribiol. 15, 34-39
- 10. Виды нейроглии у насекомых Freeman, Doherty, 2006, Trends in Neurosci. 29, 82-90
- 11. Хотя миелиновая оболочка глии у многих беспозвоночных отсутствует, у некоторых беспозвоночных она встречается и отличается от
- 12. Нейруляция первичная и вторичная Роль вертикальных и планарных сигналов в индукции ЦНС Градиенты факторов, ответственных за
- 13. Нейруляция у куриного зародыша (вид сверху). А - плоская нейральная пластинка. B- нейральная пластинка (с подлежащей
- 14. Первичная нейруляция у куриного зародыша. Изгибание нейральной пластинки в районах контакта с хордой и эктодермой. Замыкание
- 15. Окончание нейруляции в головном отделе куриного зародыша 24-х часов развития. В каудальном отделе ещё происходит гаструляция
- 16. Особенности гаструляции у млекопитающих. Формирующаяся хорда у зародыша мыши контактирует с энтодермальными клетками первичной кишки (и,
- 17. Вторичная нейруляция в хвостовом районе (после 25-ого сомита) у куриного зародыша. Образование медуллярного тяжа из которого
- 18. Нейруляция у человека (формирование нервной трубки)
- 19. Нейруляция у человека. Важность своевременного закрытия переднего и заднего нейропоров для нормального развития. Дефекты, связанные с
- 20. Смена типов кадгерина в клетках эктодермы в ходе нейруляции обусловливает образование нервной трубки и нервного гребня
- 21. Основные производные эктодермального зародышевого листка позвоночных
- 22. Участие двух сигнальных центров: узелка (node) и антериорной висцеральной энтодермы (AVE) в формирование головы и головного
- 23. Региональная специфичность нервной трубки и её производных вдоль передне-задней оси (развитие мозговых структур). Стадии трёх и
- 24. Головной мозг позвоночных Forebrain Midbrain Hindbrain
- 25. Гомеотические (гомеозисные) гены контролируют спецификацию в передне-заднем направлении заднего отдела головного (ромбомеры) мозга и всего спинного
- 26. Факторы, участвующие в закладке различных отделов головного и спинного мозга млекопитающих
- 27. Участие гомеотических генов в спецификации нейральных структур вдоль передне-задней оси позвоночных и дрозофилы (ортологичные гены отмечены
- 28. Экспериментальное увеличение концентрации ретиноевой кислоты в заднем отделе головного мозга вызывает превращение ромбомеров одного вида в
- 29. Образование различных слоев нервной трубки и образование различных отделов мозга в ходе нейрогенеза
- 30. Пролиферация и миграция дифференцирующихся нейробластов в ЦНС млекопитающих (будущие головной и спинной отделы)
- 31. Градиент «изнутри наружу» миграции нейронов при образовании слоев коры переднего отдела мозга макаки-резус
- 32. Миграция нейрона из вентрикулярного слоя в один из верхних слоев по отростку глиальной клетки. А-схема, B-
- 33. Гены, ответственные за нейрогенез у беспозвоночных и позвоночных – весьма консервативны (ортологичные гены) Пронейральные гены Нейрогенные
- 34. Последовательная спецификация линии нейробластов (в виде кластеров клеток) у Drosophila вдоль дорсо-вентральной и передне-задней осей. За
- 35. Последовательная спецификация линии нейробластов (в виде кластеров клеток) у Drosophila вдоль дорсо-вентральной и передне-задней осей (продолжение).
- 38. Пронейральные гена дрозофилы кодируют белки-ТФ с характерным доменом: basic- Helix-Loop-Helix 4 гена A-S –Complex Нейрогенные гены:
- 39. Гены нейрональной дифференцировки дрозофилы (biparous/tap) Гены кодируют ТФ со структурой bHLH, но не проявляют большой гомологии
- 40. Пронейральные гены дрозофилы семейства achaete/scute
- 41. Часть нейрогенных генов дрозофилы могут экспрессироваться в ненейронных клетках и тогда их белковые продукты действуют как
- 42. Функции транскрипционных факторов и сигнальных молекул беспозвоночных, ортологичных генам позвоночных (см. ранее)
- 43. Спецификация и дифференцировка клеток спинного мозга
- 44. Нейруляция первичная и вторичная Роль вертикальных и планарных сигналов в индукции ЦНС Градиенты факторов, ответственных за
- 45. Осевое паттернирование нервной трубки. Указаны паракринные факторы, принимающие участие в нейруляции вдоль передне-задней оси зародыша.
- 46. Участие паракринных факторов в дорсо-вентральном паттернировании нервной трубки- последовательность индуцирующих воздействий хорды и эпидермиса на нейральную
- 47. Развитие спинного мозга человека. Дифференцировка слоев мозга (поперечный срез). Отметить эпендимный, маргинальный, мантийный слои, silcus limitans,
- 48. Образование новых сигнальных центров в пластинке дна (Shh) и крыши (BMP) нервной трубки под воздействием одноимённых
- 49. Поляризация нервной трубки доль дорсо-вентральной оси
- 50. Дорсо-вентральная спецификация нервной трубки (продолжение). Становление градиентов паракринных факторов в нервной трубке в дорсо-вентральном направлении, определяющие
- 51. Дорсо-вентральная спецификация нервной трубки (поперечный срез нервной трубки куриного зародыша). Экспрессия Shh (в нижней части) -маркирована
- 52. Каскадная индукция в вентральной части нервной трубки (вентральных V3 и моторных нейронов), инициированная участком хорды прилежащим
- 53. Региональная активация в нервной трубке различных факторов транскрипции (Pax3,6,7; Mash1,2, Lim1,2,3; Isl1,2) и сигнальных молекул (Delta,
- 54. Поляризация нервной трубки вдоль передне-задней оси (продолжение)
- 55. Поляризация нервной трубки вдоль проксимо-дистальной оси
- 56. Семейство факторов транскрипции LIM, включающих подсемейство Islet Lim transcription factors are responsive for axon target selection
- 57. Спецификация различных групп двигательных нейронов в передне-заднем и дорсо-вентральном направлениях в зависимости от активации в них
- 58. Организация двигательных (моторных) нейронов (спецификация LIM-содержащих нейронов спинного мозга.
- 59. Нервный гребень (только у позвоночных) Клетки нервного гребня позвоночных – предшественники периферической (парасимпатической и симпатической) нервной
- 60. Нервный гребень (НГ) – производное эмбриональной эктодермы (существует временно), закладывается по краям нейральной пластинки и окончательно
- 61. Нервный гребень (НГ) – эволюционно появляется у позвоночных. В эмбриогенезе формируется дорсолатерально в передне-заднем направлении и
- 62. Производные клеток нервного гребня (НГ)
- 63. Миграция клеток нервного гребня в туловищном отделе куриного зародыша (два пути миграции у птиц и млекопитающих,
- 64. Специфические маркеры НГ свидетельствуют о миграции клеток НГ куриного зародыша вентрально через антериорные отделы склеротомов (роль
- 65. Роль белков RhoB и Slug в клетках НГ, способствующих миграции клеток. (Rho участвует в упрочении цитоскелета
- 66. Последовательное ограничение путей дифференцировки клеток НГ (гипотетическая модель)
- 67. Дифференцировка клеток НГ определяется последовательными индуцирующими воздействиями паракринных факторов FGF и NGF (NGF-обусловливает миграцию клеток, действуя
- 68. Конечные стадии дифференцировки клеток нервного гребня туловищного отдела. Альтернативные варианты дифференцировки в хромоафинные клетки мозгового вещества
- 69. Черепные плакоды – третий источник нервных клеток (и не только их)
- 70. Черепные плакоды - временные локальные утолщения эктодермы между эпидермальной и нейральной эктодермой головы. Формирование плакод индуцируется
- 71. Черепные плакоды
- 72. Как растут аксоны?
- 73. Схематическое изображение моторного нейрона. Электрические импульсы приходят от дендритов и передаются по аксону. Длина аксона может
- 74. Внутриклеточные причины роста аксонов: взаимодействия актиновых и миозиновых структур
- 75. Внешние факторы управления движением конуса роста аксона (проецирование его на ткани-мишени) в принципе те же, что
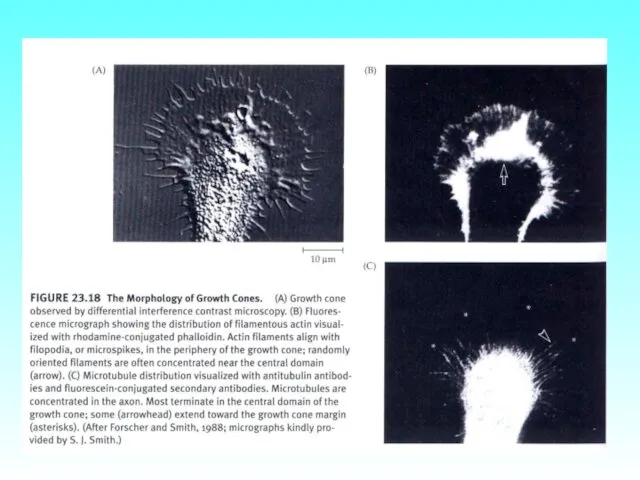
- 76. Конус роста аксона нейрональной клетки бражника. Видны микрошипики (филоподии, содержащие микрофиламенты, окрашены в зелёный цвет, и
- 78. Семафорины (наряду с эфринами) - мембранные или секреторные белки ряда (эпителиальных) клеток ограничивают рост аксонов в
- 79. Общая схема организации в зародыше направления роста аксонов нейрональных клеток мозга и нервного гребня (путём отталкивания
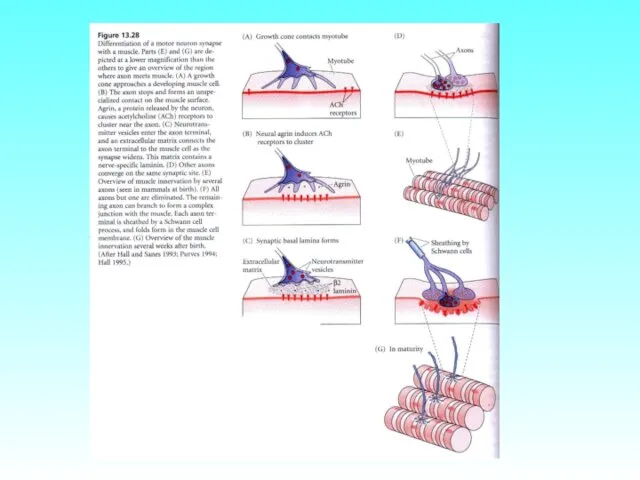
- 80. Формирование синапсов
- 83. Скачать презентацию
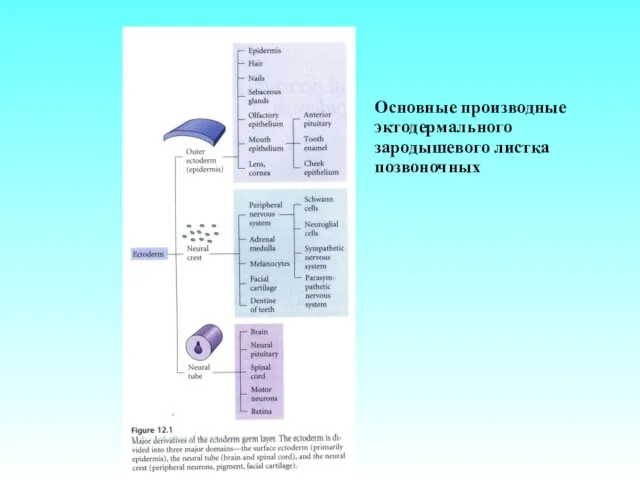
Основные производные эктодермального зародышевого листка позвоночных
Основные производные эктодермального зародышевого листка позвоночных
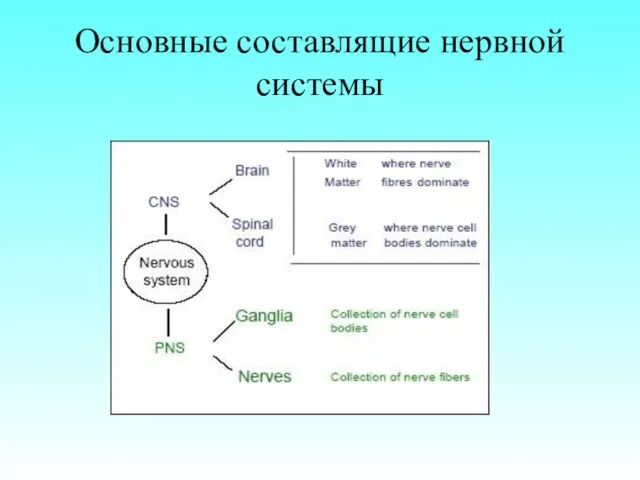
Основные составлящие нервной системы
Основные составлящие нервной системы
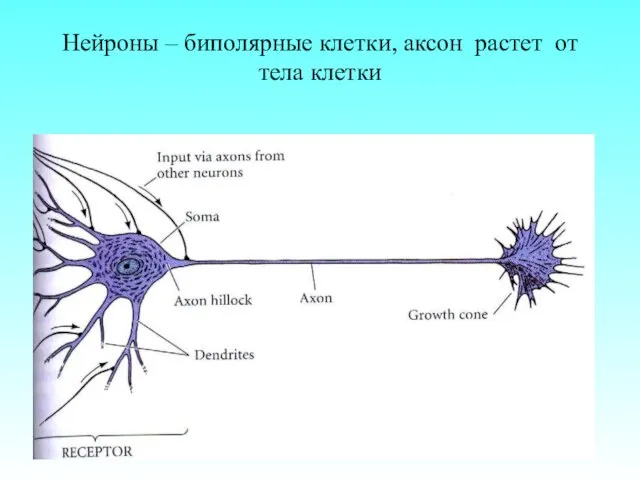
Нейроны – биполярные клетки, аксон растет от тела клетки
Нейроны – биполярные клетки, аксон растет от тела клетки
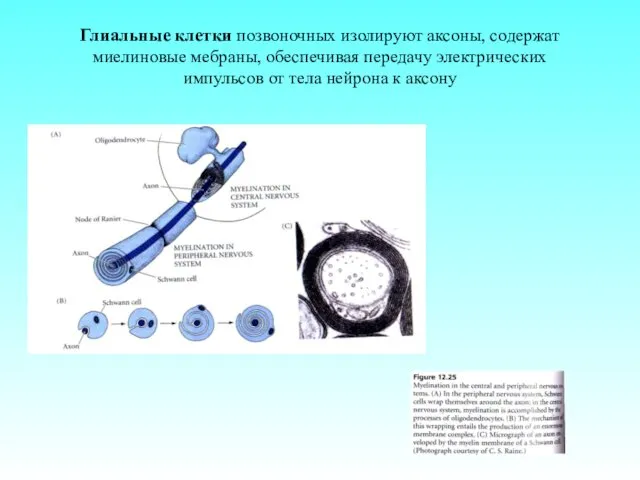
Глиальные клетки позвоночных изолируют аксоны, содержат миелиновые мебраны, обеспечивая передачу электрических
Глиальные клетки позвоночных изолируют аксоны, содержат миелиновые мебраны, обеспечивая передачу электрических

В течение развития нейральные стволовые клетки дают начало нейронам ЦНС
Они
В течение развития нейральные стволовые клетки дают начало нейронам ЦНС
Они

НЕЙРУЛЯЦИЯ
1. Начальная стадия нейрогенеза у позвоночных - нейруляция.
У позвоночных
НЕЙРУЛЯЦИЯ 1. Начальная стадия нейрогенеза у позвоночных - нейруляция. У позвоночных
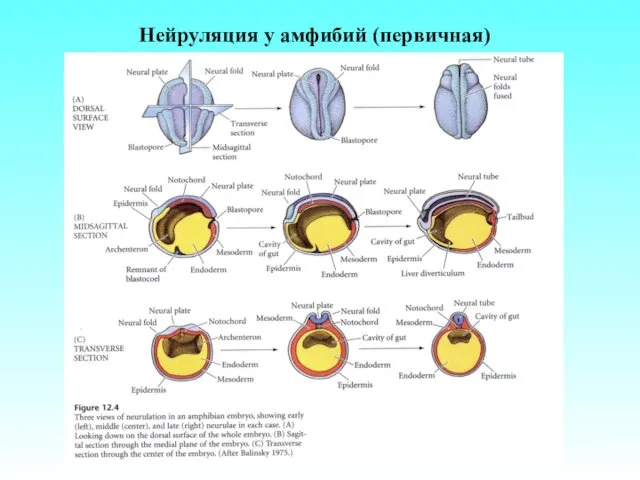
Нейруляция у амфибий (первичная)
Нейруляция у амфибий (первичная)
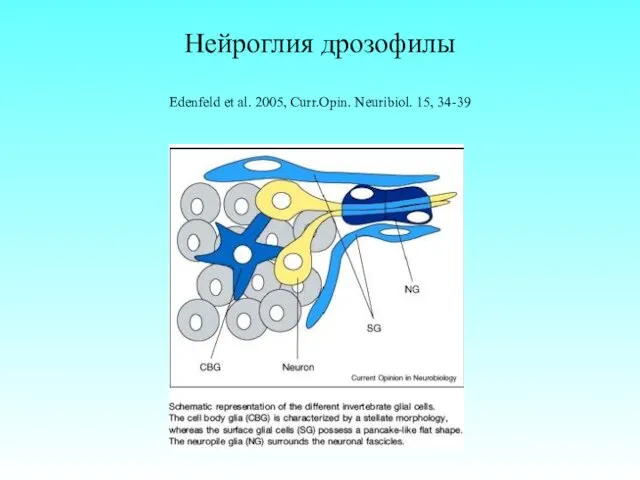
Нейроглия дрозофилы
Edenfeld et al. 2005, Curr.Opin. Neuribiol. 15, 34-39
Нейроглия дрозофилы
Edenfeld et al. 2005, Curr.Opin. Neuribiol. 15, 34-39

Виды нейроглии у насекомых
Freeman, Doherty, 2006, Trends in Neurosci. 29, 82-90
Виды нейроглии у насекомых
Freeman, Doherty, 2006, Trends in Neurosci. 29, 82-90
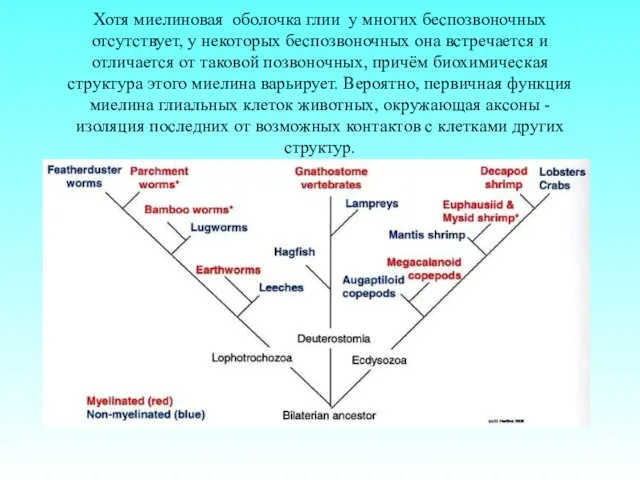
Хотя миелиновая оболочка глии у многих беспозвоночных отсутствует, у некоторых беспозвоночных
Хотя миелиновая оболочка глии у многих беспозвоночных отсутствует, у некоторых беспозвоночных
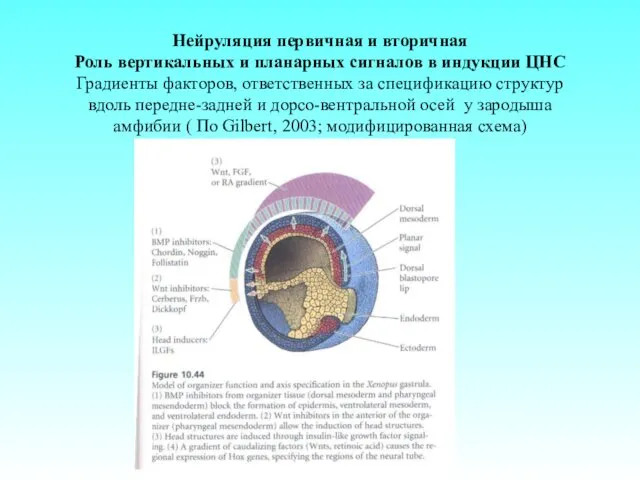
Нейруляция первичная и вторичная
Роль вертикальных и планарных сигналов в индукции ЦНС
Градиенты
Нейруляция первичная и вторичная Роль вертикальных и планарных сигналов в индукции ЦНС Градиенты
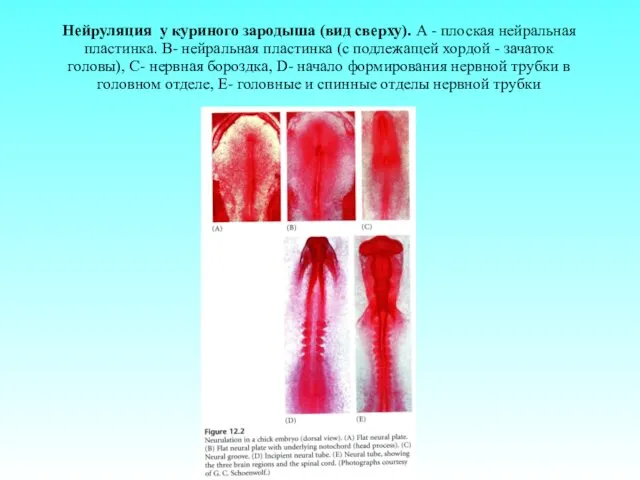
Нейруляция у куриного зародыша (вид сверху). А - плоская нейральная пластинка.
Нейруляция у куриного зародыша (вид сверху). А - плоская нейральная пластинка.
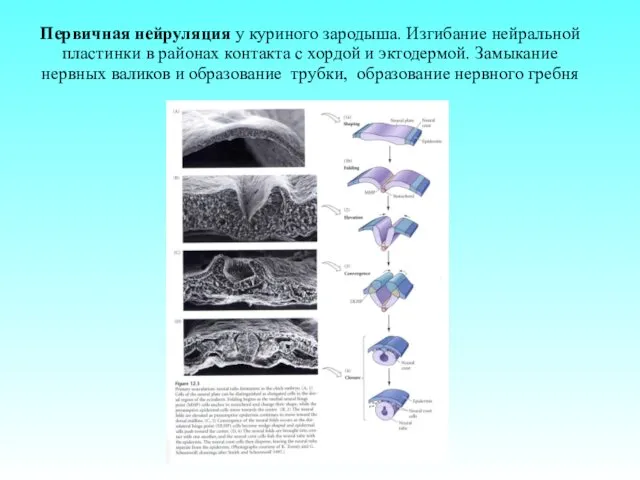
Первичная нейруляция у куриного зародыша. Изгибание нейральной пластинки в районах контакта
Первичная нейруляция у куриного зародыша. Изгибание нейральной пластинки в районах контакта
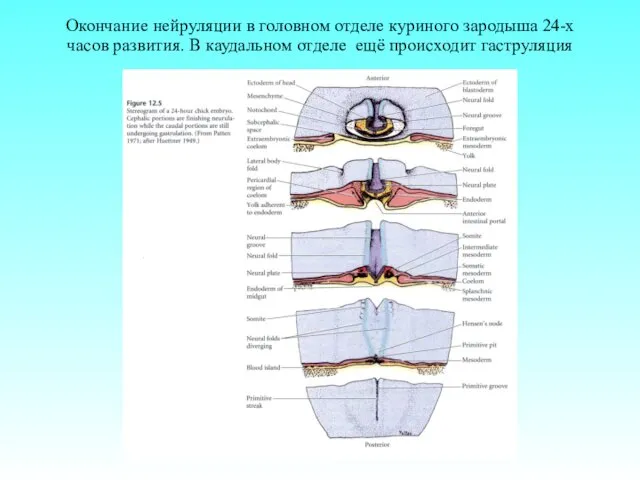
Окончание нейруляции в головном отделе куриного зародыша 24-х часов развития. В
Окончание нейруляции в головном отделе куриного зародыша 24-х часов развития. В

Особенности гаструляции у млекопитающих. Формирующаяся хорда у зародыша мыши контактирует с
Особенности гаструляции у млекопитающих. Формирующаяся хорда у зародыша мыши контактирует с
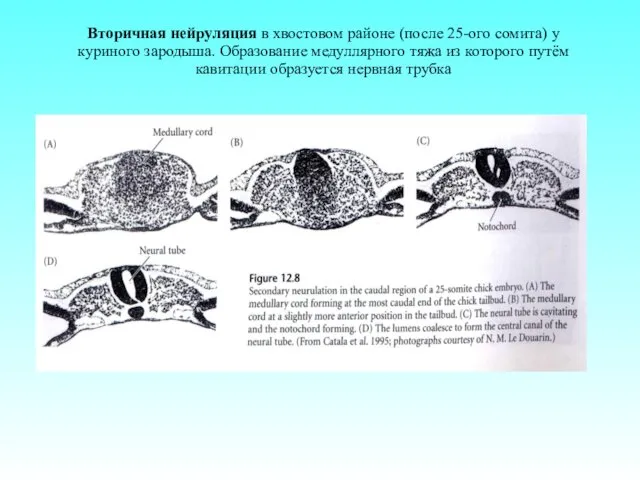
Вторичная нейруляция в хвостовом районе (после 25-ого сомита) у куриного зародыша.
Вторичная нейруляция в хвостовом районе (после 25-ого сомита) у куриного зародыша.
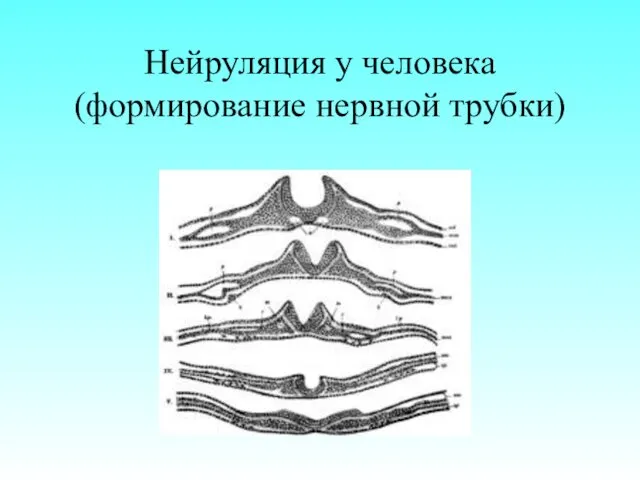
Нейруляция у человека (формирование нервной трубки)
Нейруляция у человека (формирование нервной трубки)
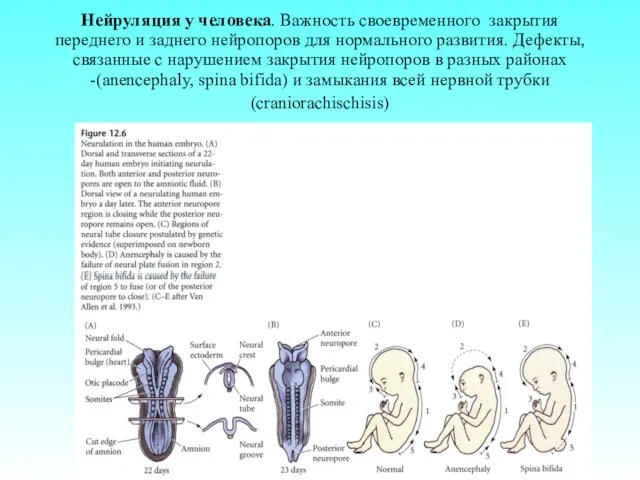
Нейруляция у человека. Важность своевременного закрытия переднего и заднего нейропоров для
Нейруляция у человека. Важность своевременного закрытия переднего и заднего нейропоров для
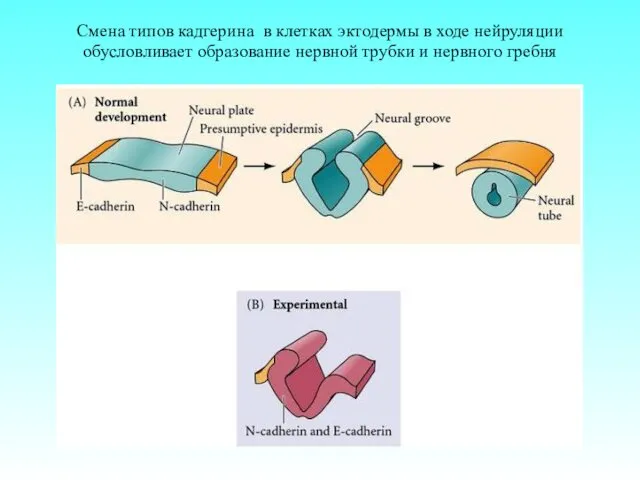
Смена типов кадгерина в клетках эктодермы в ходе нейруляции обусловливает образование
Смена типов кадгерина в клетках эктодермы в ходе нейруляции обусловливает образование
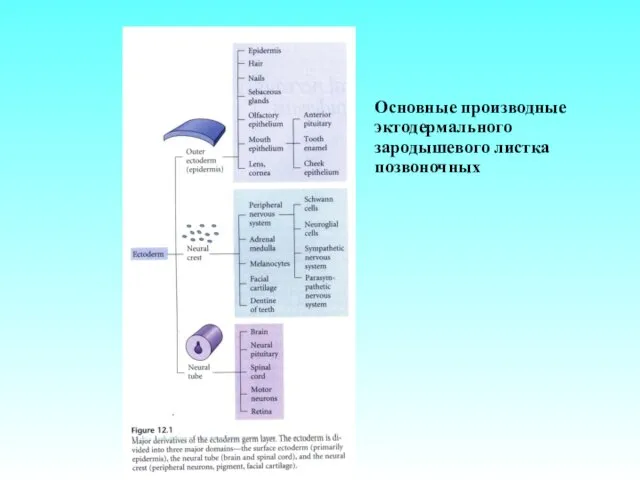
Основные производные эктодермального зародышевого листка позвоночных
Основные производные эктодермального зародышевого листка позвоночных
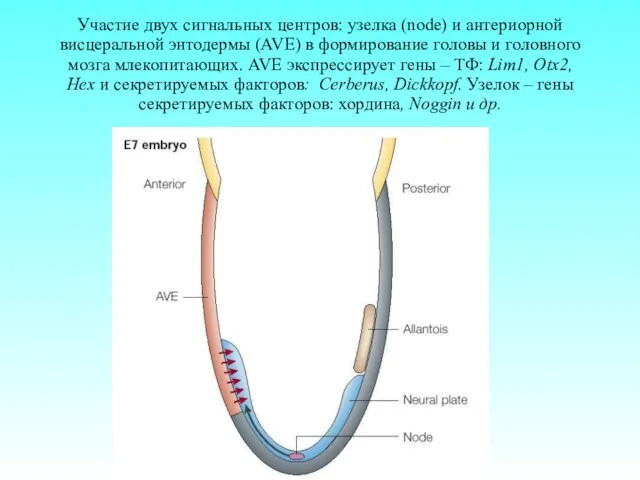
Участие двух сигнальных центров: узелка (node) и антериорной висцеральной энтодермы (AVE)
Участие двух сигнальных центров: узелка (node) и антериорной висцеральной энтодермы (AVE)
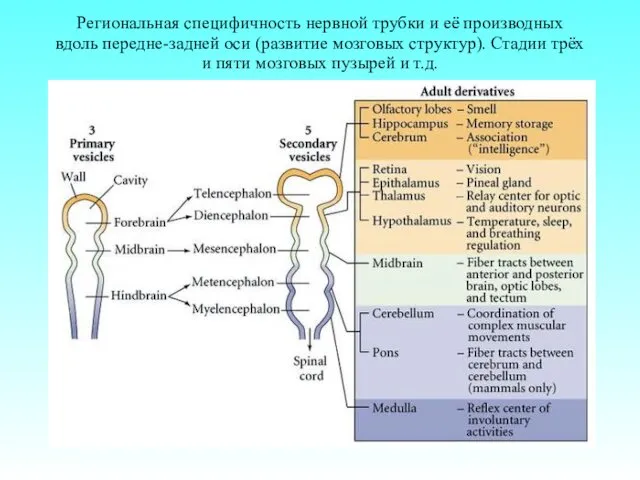
Региональная специфичность нервной трубки и её производных вдоль передне-задней оси (развитие
Региональная специфичность нервной трубки и её производных вдоль передне-задней оси (развитие
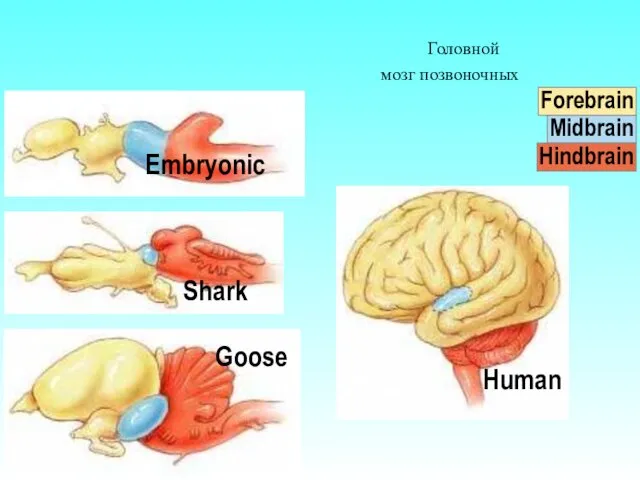
Головной мозг позвоночных
Forebrain
Midbrain
Hindbrain
Головной мозг позвоночных
Forebrain
Midbrain
Hindbrain
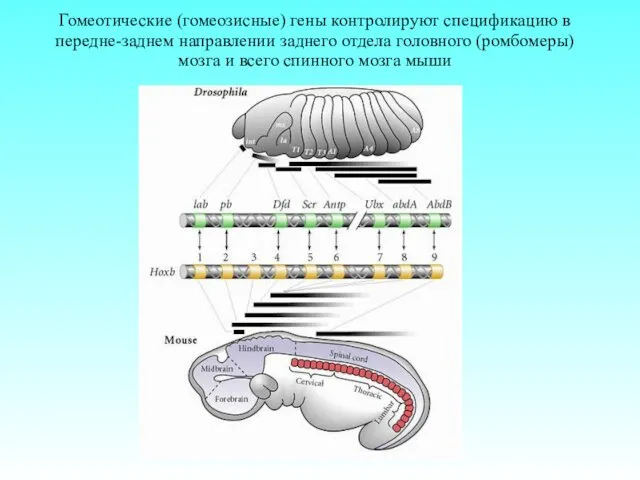
Гомеотические (гомеозисные) гены контролируют спецификацию в передне-заднем направлении заднего отдела головного
Гомеотические (гомеозисные) гены контролируют спецификацию в передне-заднем направлении заднего отдела головного
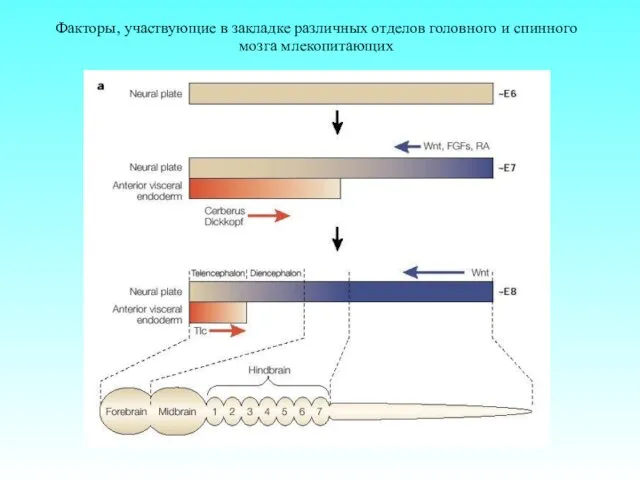
Факторы, участвующие в закладке различных отделов головного и спинного мозга млекопитающих
Факторы, участвующие в закладке различных отделов головного и спинного мозга млекопитающих
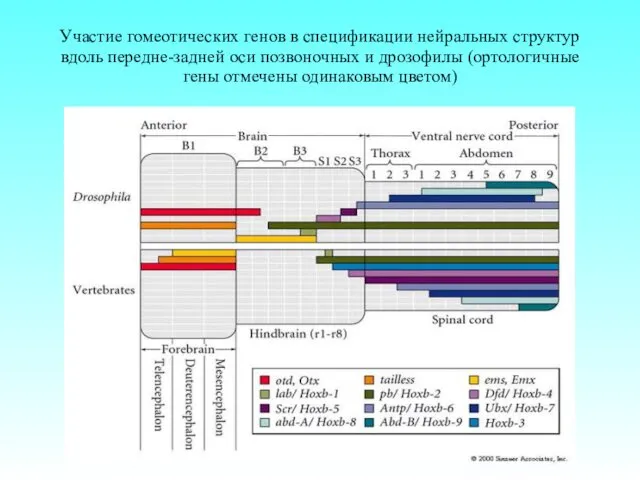
Участие гомеотических генов в спецификации нейральных структур вдоль передне-задней оси позвоночных
Участие гомеотических генов в спецификации нейральных структур вдоль передне-задней оси позвоночных
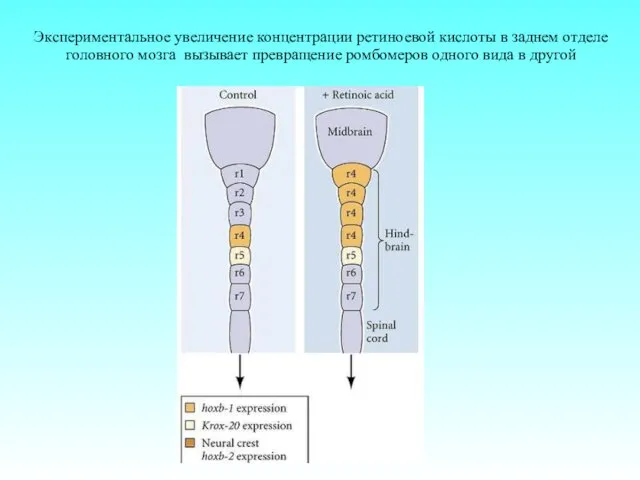
Экспериментальное увеличение концентрации ретиноевой кислоты в заднем отделе головного мозга вызывает
Экспериментальное увеличение концентрации ретиноевой кислоты в заднем отделе головного мозга вызывает

Образование различных слоев нервной трубки и образование различных отделов мозга в
Образование различных слоев нервной трубки и образование различных отделов мозга в
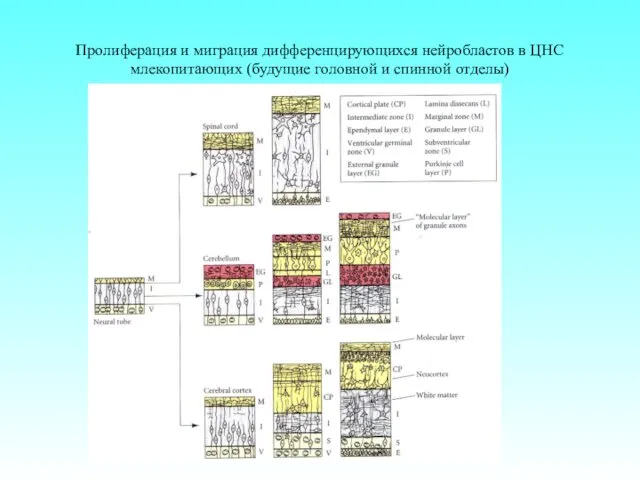
Пролиферация и миграция дифференцирующихся нейробластов в ЦНС млекопитающих (будущие головной и
Пролиферация и миграция дифференцирующихся нейробластов в ЦНС млекопитающих (будущие головной и
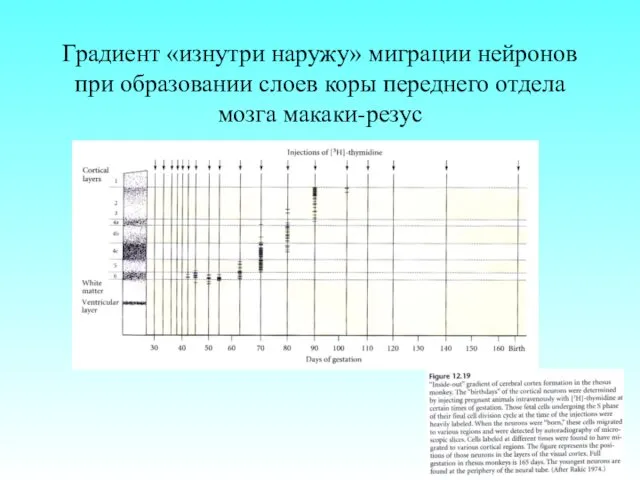
Градиент «изнутри наружу» миграции нейронов при образовании слоев коры переднего отдела
Градиент «изнутри наружу» миграции нейронов при образовании слоев коры переднего отдела
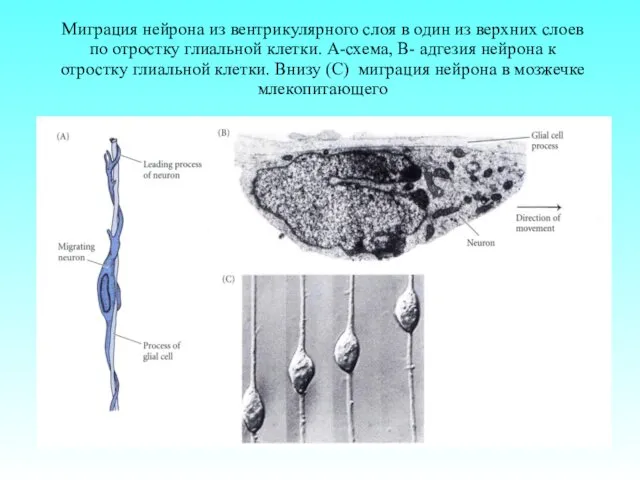
Миграция нейрона из вентрикулярного слоя в один из верхних слоев по
Миграция нейрона из вентрикулярного слоя в один из верхних слоев по

Гены, ответственные за нейрогенез у беспозвоночных и позвоночных – весьма консервативны
Гены, ответственные за нейрогенез у беспозвоночных и позвоночных – весьма консервативны
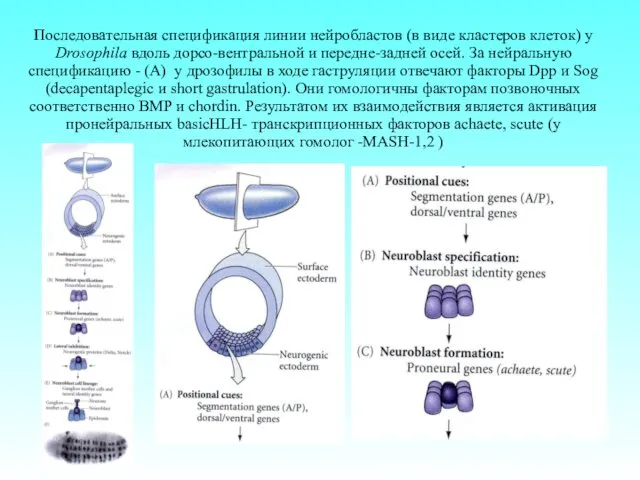
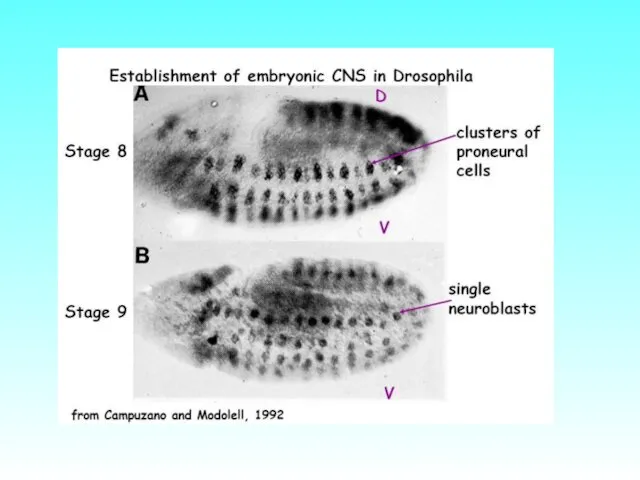
Последовательная спецификация линии нейробластов (в виде кластеров клеток) у Drosophila вдоль
Последовательная спецификация линии нейробластов (в виде кластеров клеток) у Drosophila вдоль
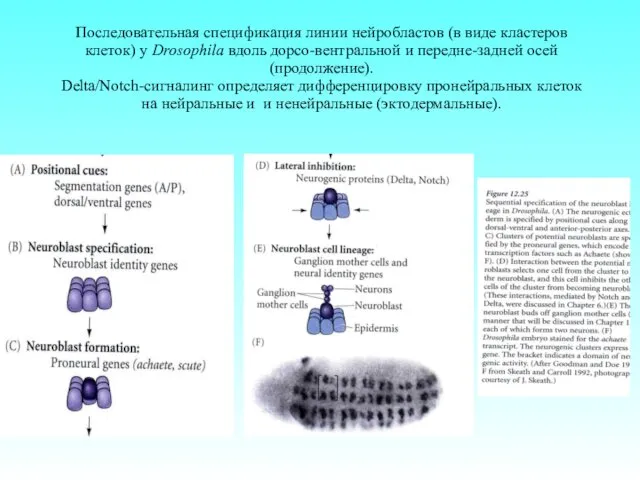
Последовательная спецификация линии нейробластов (в виде кластеров клеток) у Drosophila вдоль
Последовательная спецификация линии нейробластов (в виде кластеров клеток) у Drosophila вдоль


Пронейральные гена дрозофилы кодируют белки-ТФ с характерным доменом: basic- Helix-Loop-Helix
4 гена
Пронейральные гена дрозофилы кодируют белки-ТФ с характерным доменом: basic- Helix-Loop-Helix
4 гена

Гены нейрональной дифференцировки дрозофилы (biparous/tap)
Гены кодируют ТФ со структурой bHLH, но
Гены нейрональной дифференцировки дрозофилы (biparous/tap) Гены кодируют ТФ со структурой bHLH, но
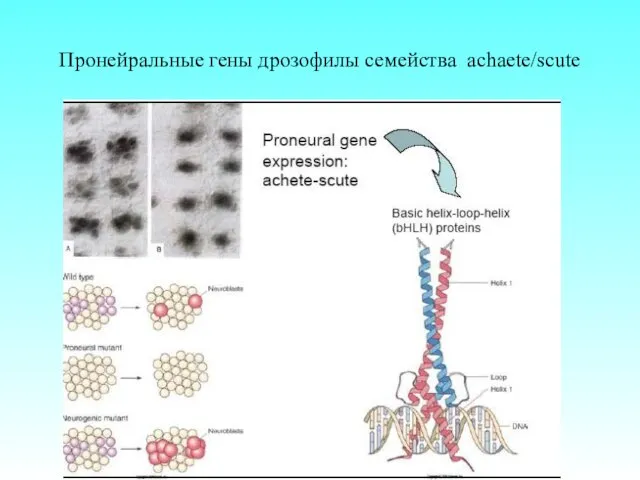
Пронейральные гены дрозофилы семейства achaete/scute
Пронейральные гены дрозофилы семейства achaete/scute

Часть нейрогенных генов дрозофилы могут экспрессироваться в ненейронных клетках и тогда
Часть нейрогенных генов дрозофилы могут экспрессироваться в ненейронных клетках и тогда
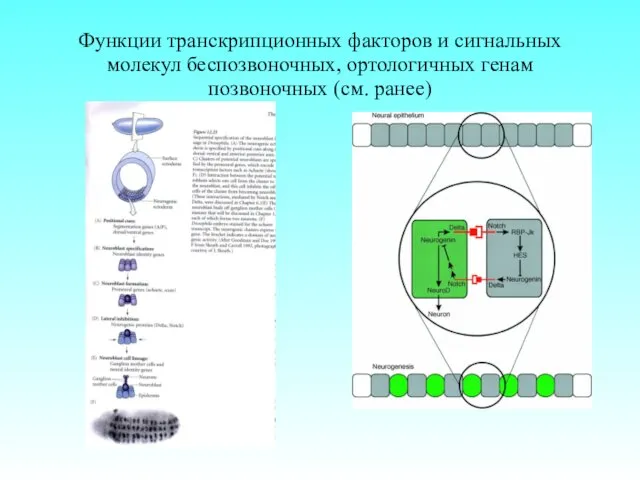
Функции транскрипционных факторов и сигнальных молекул беспозвоночных, ортологичных генам позвоночных (см.
Функции транскрипционных факторов и сигнальных молекул беспозвоночных, ортологичных генам позвоночных (см.

Спецификация и дифференцировка клеток спинного мозга
Спецификация и дифференцировка клеток спинного мозга
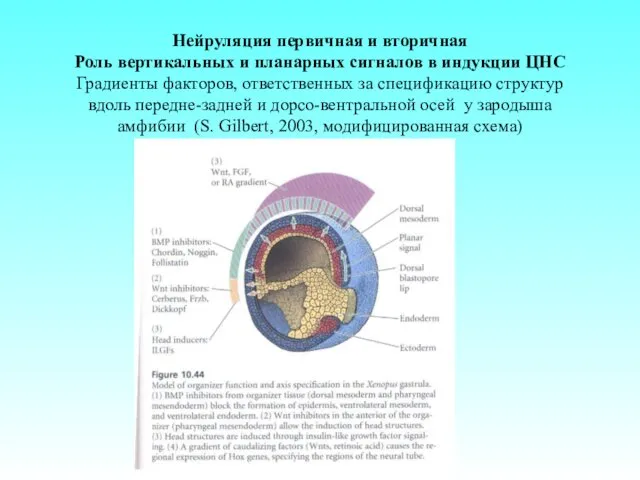
Нейруляция первичная и вторичная
Роль вертикальных и планарных сигналов в индукции ЦНС
Градиенты
Нейруляция первичная и вторичная Роль вертикальных и планарных сигналов в индукции ЦНС Градиенты
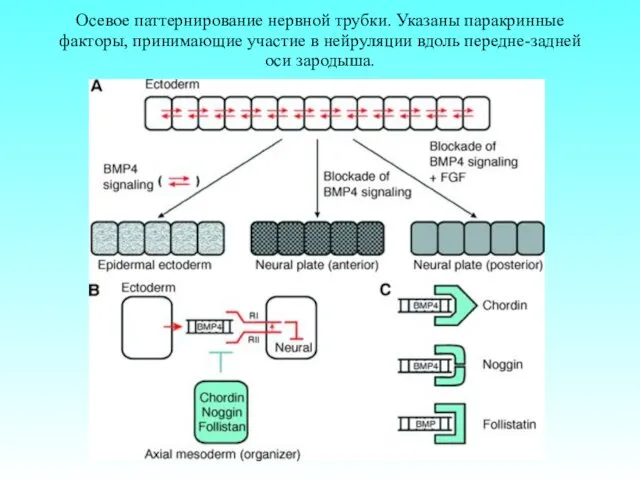
Осевое паттернирование нервной трубки. Указаны паракринные факторы, принимающие участие в нейруляции
Осевое паттернирование нервной трубки. Указаны паракринные факторы, принимающие участие в нейруляции
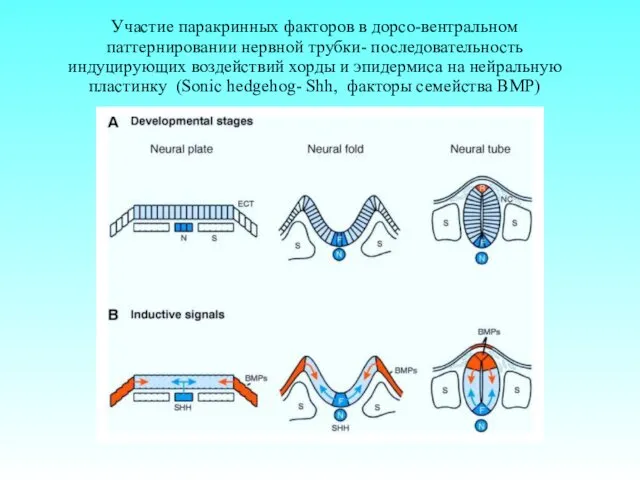
Участие паракринных факторов в дорсо-вентральном паттернировании нервной трубки- последовательность индуцирующих воздействий
Участие паракринных факторов в дорсо-вентральном паттернировании нервной трубки- последовательность индуцирующих воздействий
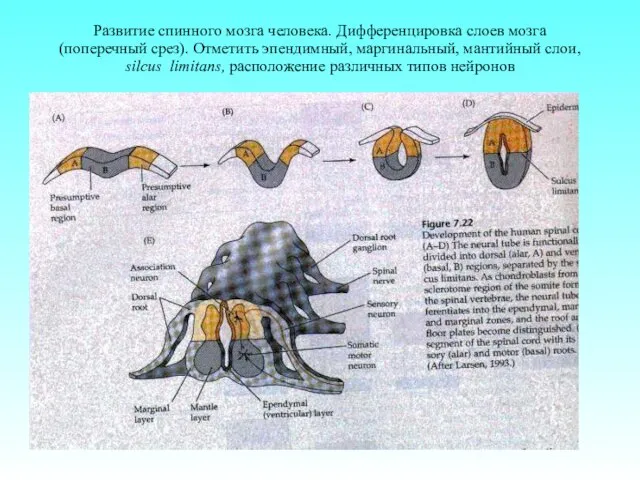
Развитие спинного мозга человека. Дифференцировка слоев мозга (поперечный срез). Отметить эпендимный,
Развитие спинного мозга человека. Дифференцировка слоев мозга (поперечный срез). Отметить эпендимный,
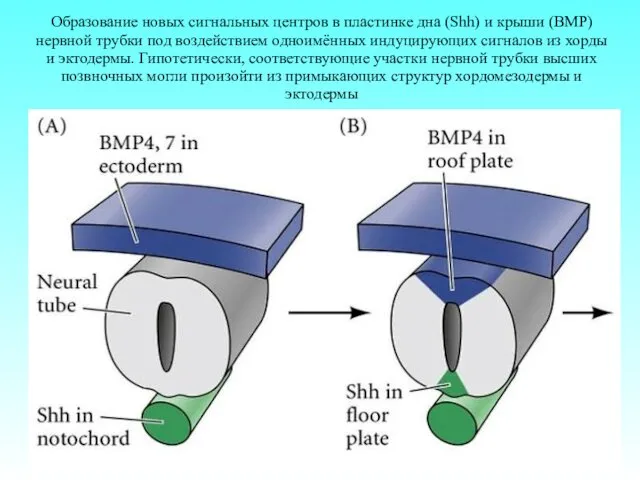
Образование новых сигнальных центров в пластинке дна (Shh) и крыши (BMP)
Образование новых сигнальных центров в пластинке дна (Shh) и крыши (BMP)

Поляризация нервной трубки доль дорсо-вентральной оси
Поляризация нервной трубки доль дорсо-вентральной оси
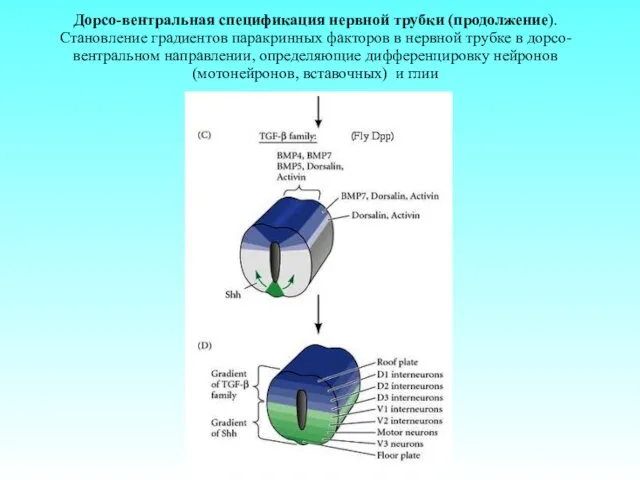
Дорсо-вентральная спецификация нервной трубки (продолжение). Становление градиентов паракринных факторов в нервной
Дорсо-вентральная спецификация нервной трубки (продолжение). Становление градиентов паракринных факторов в нервной
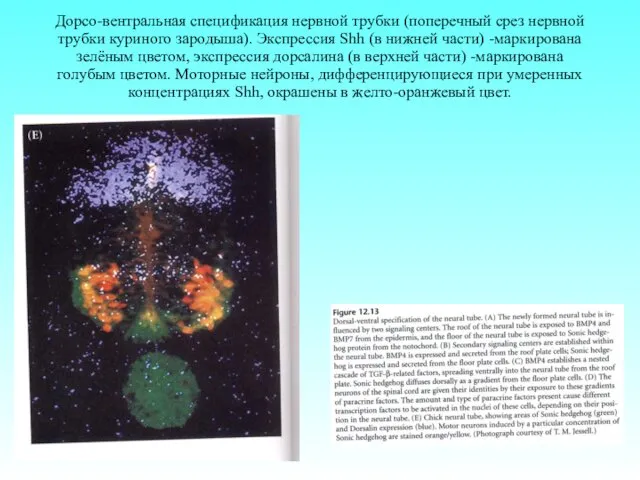
Дорсо-вентральная спецификация нервной трубки (поперечный срез нервной трубки куриного зародыша). Экспрессия
Дорсо-вентральная спецификация нервной трубки (поперечный срез нервной трубки куриного зародыша). Экспрессия
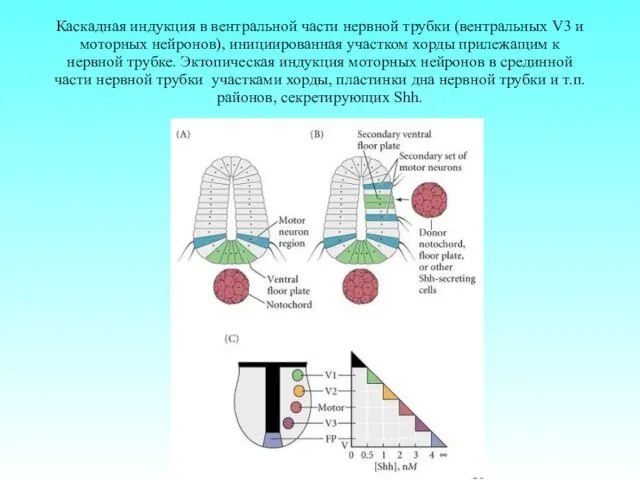
Каскадная индукция в вентральной части нервной трубки (вентральных V3 и моторных
Каскадная индукция в вентральной части нервной трубки (вентральных V3 и моторных
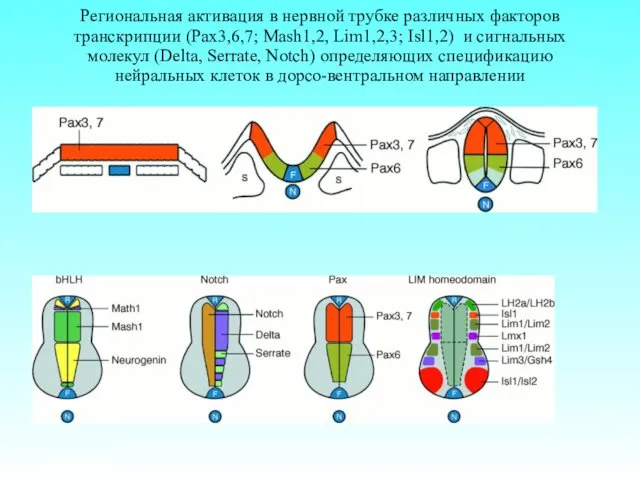
Региональная активация в нервной трубке различных факторов транскрипции (Pax3,6,7; Mash1,2, Lim1,2,3;
Региональная активация в нервной трубке различных факторов транскрипции (Pax3,6,7; Mash1,2, Lim1,2,3;
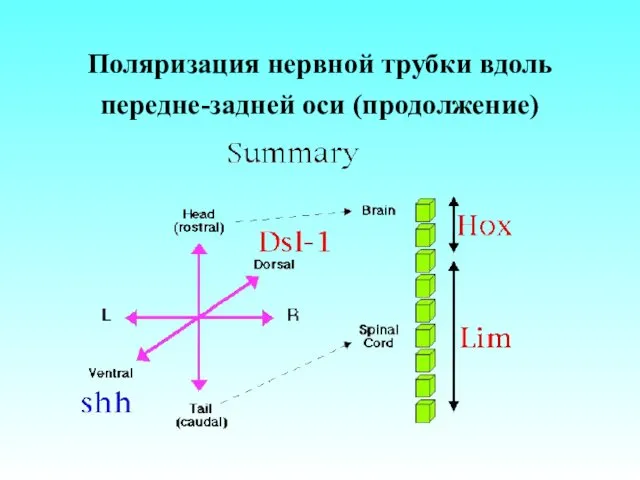
Поляризация нервной трубки вдоль передне-задней оси (продолжение)
Поляризация нервной трубки вдоль передне-задней оси (продолжение)
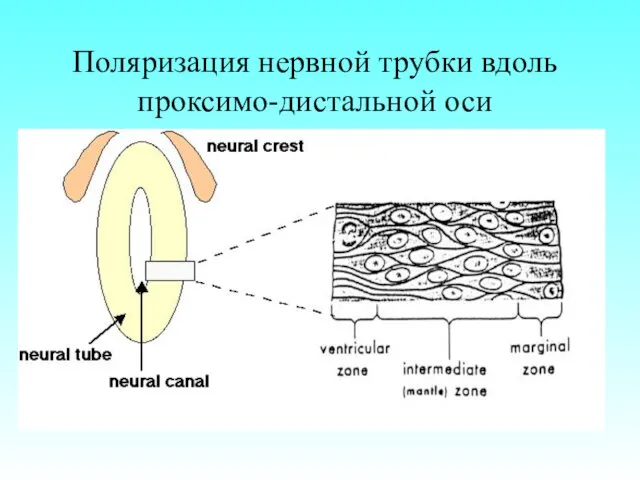
Поляризация нервной трубки вдоль проксимо-дистальной оси
Поляризация нервной трубки вдоль проксимо-дистальной оси
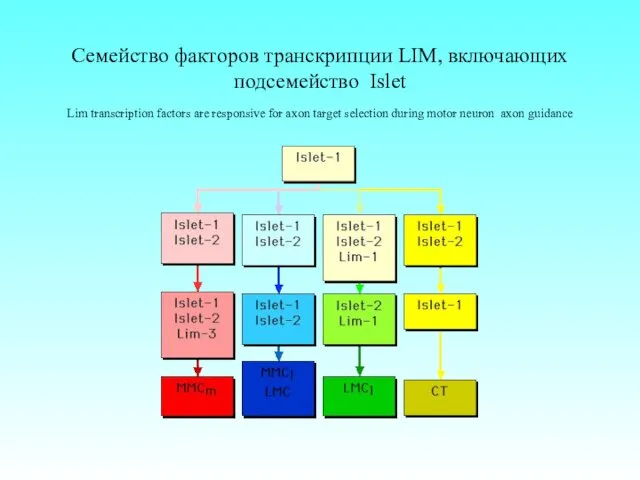
Семейство факторов транскрипции LIM, включающих подсемейство Islet
Lim transcription factors are responsive
Семейство факторов транскрипции LIM, включающих подсемейство Islet Lim transcription factors are responsive

Спецификация различных групп двигательных нейронов в передне-заднем и дорсо-вентральном направлениях
Спецификация различных групп двигательных нейронов в передне-заднем и дорсо-вентральном направлениях
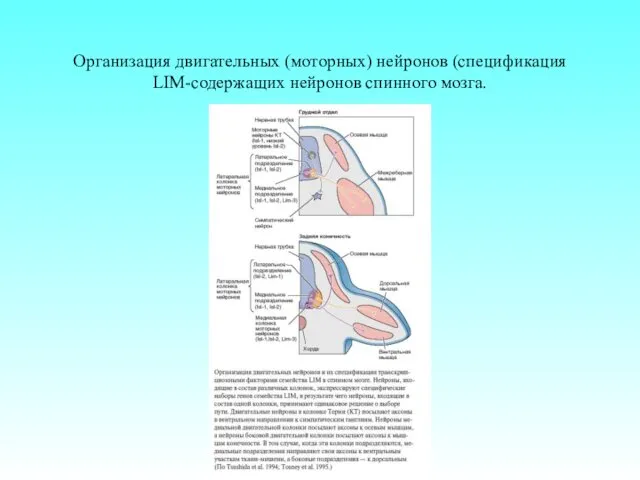
Организация двигательных (моторных) нейронов (спецификация LIM-содержащих нейронов спинного мозга.
Организация двигательных (моторных) нейронов (спецификация LIM-содержащих нейронов спинного мозга.

Нервный гребень
(только у позвоночных)
Клетки нервного гребня позвоночных – предшественники периферической
Нервный гребень
(только у позвоночных)
Клетки нервного гребня позвоночных – предшественники периферической

Нервный гребень (НГ) – производное эмбриональной эктодермы (существует временно), закладывается по
Нервный гребень (НГ) – производное эмбриональной эктодермы (существует временно), закладывается по

Нервный гребень (НГ) – эволюционно появляется у позвоночных. В эмбриогенезе формируется
Нервный гребень (НГ) – эволюционно появляется у позвоночных. В эмбриогенезе формируется

Производные клеток нервного гребня (НГ)
Производные клеток нервного гребня (НГ)
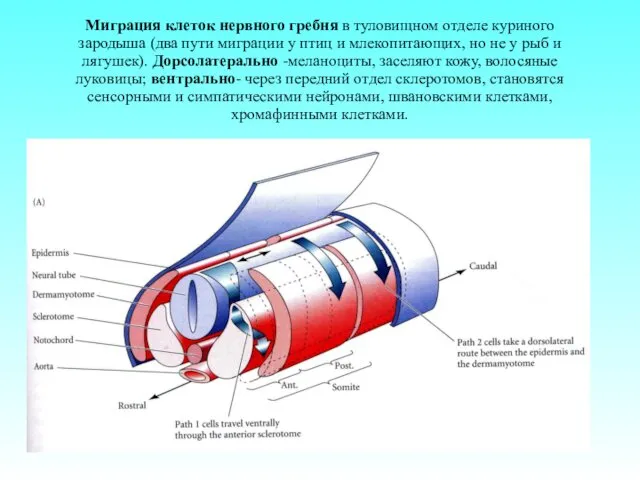
Миграция клеток нервного гребня в туловищном отделе куриного зародыша (два пути
Миграция клеток нервного гребня в туловищном отделе куриного зародыша (два пути
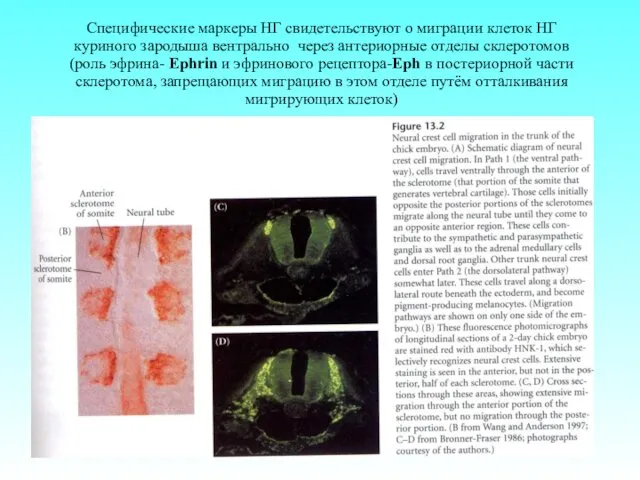
Специфические маркеры НГ свидетельствуют о миграции клеток НГ куриного зародыша вентрально
Специфические маркеры НГ свидетельствуют о миграции клеток НГ куриного зародыша вентрально
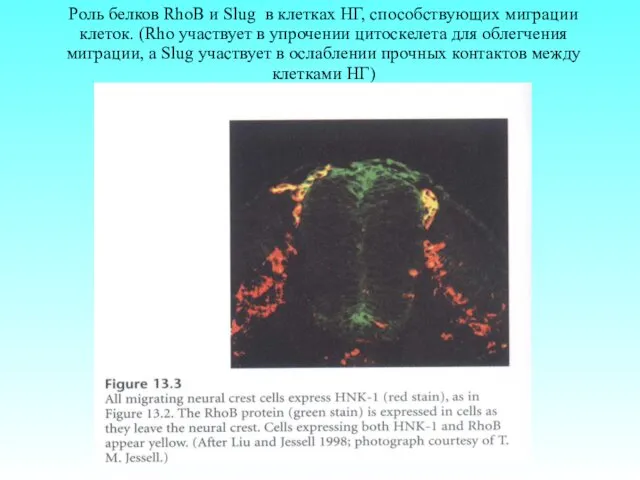
Роль белков RhoB и Slug в клетках НГ, способствующих миграции клеток.
Роль белков RhoB и Slug в клетках НГ, способствующих миграции клеток.
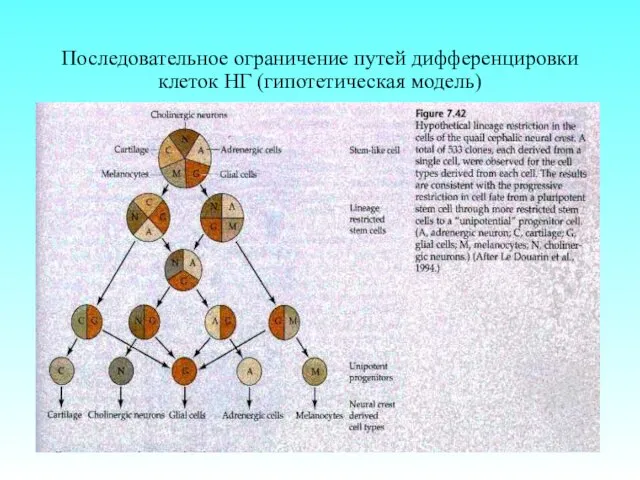
Последовательное ограничение путей дифференцировки клеток НГ (гипотетическая модель)
Последовательное ограничение путей дифференцировки клеток НГ (гипотетическая модель)
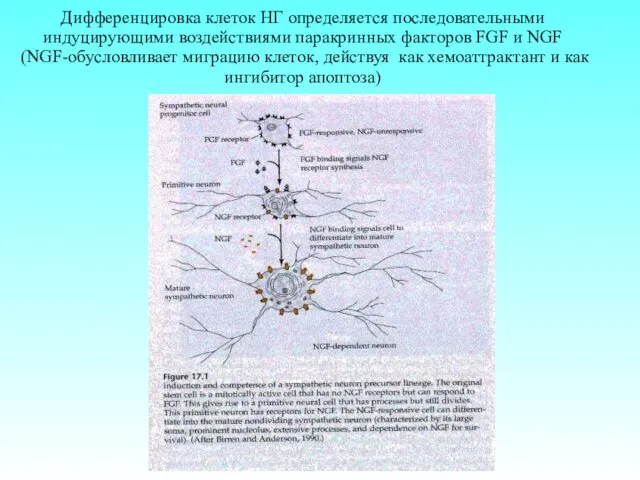
Дифференцировка клеток НГ определяется последовательными индуцирующими воздействиями паракринных факторов FGF и
Дифференцировка клеток НГ определяется последовательными индуцирующими воздействиями паракринных факторов FGF и
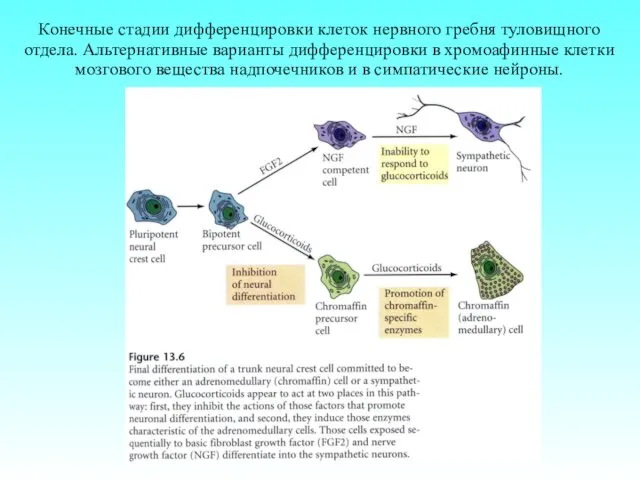
Конечные стадии дифференцировки клеток нервного гребня туловищного отдела. Альтернативные варианты дифференцировки
Конечные стадии дифференцировки клеток нервного гребня туловищного отдела. Альтернативные варианты дифференцировки

Черепные плакоды – третий источник нервных клеток (и не только их)
Черепные плакоды – третий источник нервных клеток (и не только их)
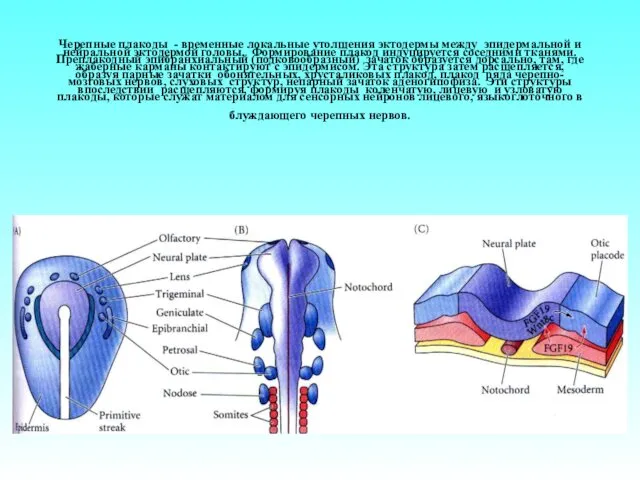
Черепные плакоды - временные локальные утолщения эктодермы между эпидермальной и нейральной
Черепные плакоды - временные локальные утолщения эктодермы между эпидермальной и нейральной
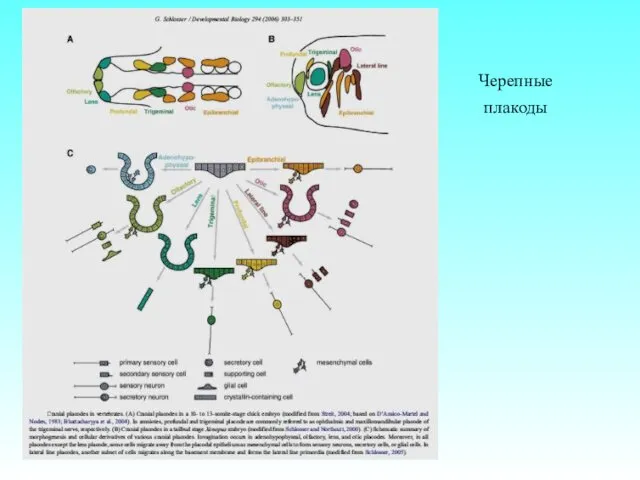
Черепные плакоды
Черепные плакоды

Как растут аксоны?
Как растут аксоны?
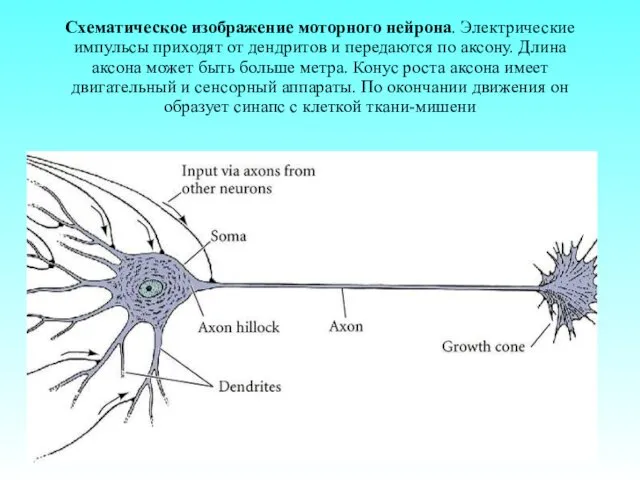
Схематическое изображение моторного нейрона. Электрические импульсы приходят от дендритов и передаются
Схематическое изображение моторного нейрона. Электрические импульсы приходят от дендритов и передаются
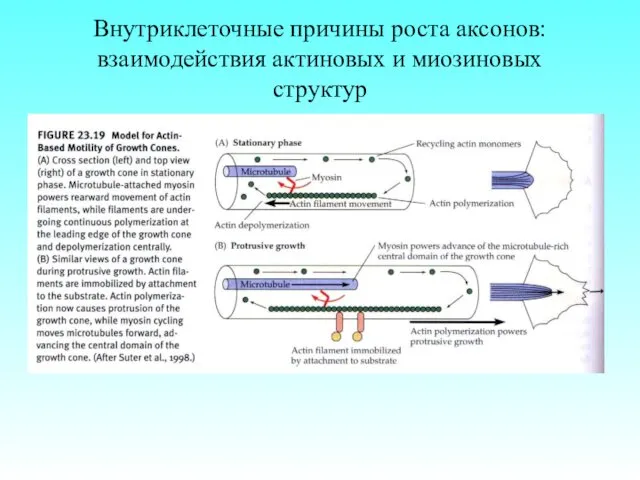
Внутриклеточные причины роста аксонов: взаимодействия актиновых и миозиновых структур
Внутриклеточные причины роста аксонов: взаимодействия актиновых и миозиновых структур

Внешние факторы управления движением конуса роста аксона (проецирование его на ткани-мишени)
Внешние факторы управления движением конуса роста аксона (проецирование его на ткани-мишени)
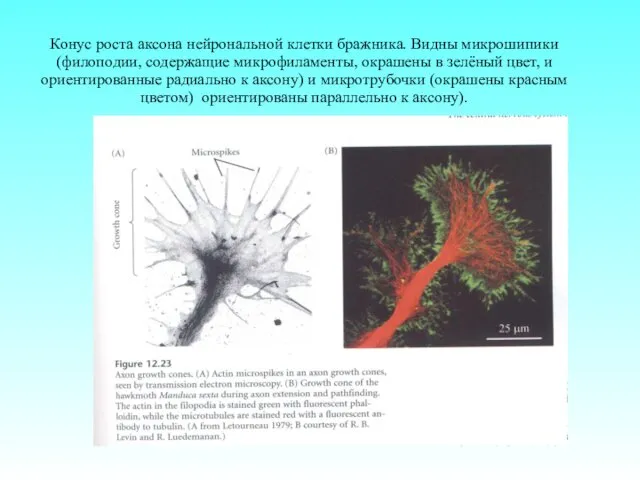
Конус роста аксона нейрональной клетки бражника. Видны микрошипики (филоподии, содержащие микрофиламенты,
Конус роста аксона нейрональной клетки бражника. Видны микрошипики (филоподии, содержащие микрофиламенты,
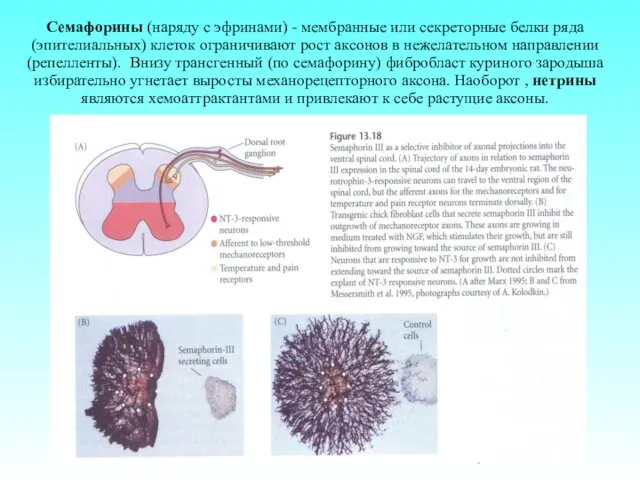
Семафорины (наряду с эфринами) - мембранные или секреторные белки ряда (эпителиальных)
Семафорины (наряду с эфринами) - мембранные или секреторные белки ряда (эпителиальных)
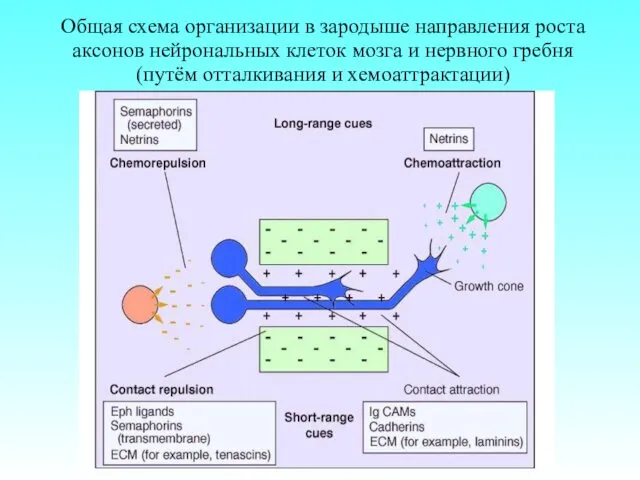
Общая схема организации в зародыше направления роста аксонов нейрональных клеток мозга
Общая схема организации в зародыше направления роста аксонов нейрональных клеток мозга

 Презентация на тему "Гипотезы происхождения Жизни" - скачать презентации по Биологии
Презентация на тему "Гипотезы происхождения Жизни" - скачать презентации по Биологии Тема: Основи безпеки харчування. Хвороби, до яких призводить неякісне, незбалансоване харчування. Перша допомога при харчових
Тема: Основи безпеки харчування. Хвороби, до яких призводить неякісне, незбалансоване харчування. Перша допомога при харчових  Презентация на тему "Зрительный анализатор" - скачать презентации по Биологии
Презентация на тему "Зрительный анализатор" - скачать презентации по Биологии Презентация на тему Птицы леса
Презентация на тему Птицы леса  Портреты, полученные из ДНК
Портреты, полученные из ДНК Презентация на тему "МНОГООБРАЗИЕ КУЛЬТУРНЫХ РАСТЕНИЙ" - скачать бесплатно презентации по Биологии
Презентация на тему "МНОГООБРАЗИЕ КУЛЬТУРНЫХ РАСТЕНИЙ" - скачать бесплатно презентации по Биологии Мухи (лат. Musca)
Мухи (лат. Musca) Презентация кормушки
Презентация кормушки Санитарная микробиология
Санитарная микробиология Достижения отечественной селекции
Достижения отечественной селекции Понятие дизайна и фитодизайна. Фитоэргономика
Понятие дизайна и фитодизайна. Фитоэргономика Презентация на тему "Спинной мозг" - скачать бесплатно презентации по Биологии
Презентация на тему "Спинной мозг" - скачать бесплатно презентации по Биологии Молюски Виконала учениця 8 класу ЗОШ 1-3 ст. №5 м.Ковеля Хіняєва Олена
Молюски Виконала учениця 8 класу ЗОШ 1-3 ст. №5 м.Ковеля Хіняєва Олена  Женская половая система
Женская половая система Корень. Строение и функции. Видоизменения корней
Корень. Строение и функции. Видоизменения корней Група №2 Кущенко К. Беспала С. Дмитренко А. Бандура А. Часник Є.
Група №2 Кущенко К. Беспала С. Дмитренко А. Бандура А. Часник Є.  Його вклад в генетику Олександр Сергійович Серебровський
Його вклад в генетику Олександр Сергійович Серебровський  Обмен веществ. Ассимиляция. Диссимиляция
Обмен веществ. Ассимиляция. Диссимиляция Частная микробиология. Pseudomonas aeruginosa, род proteus
Частная микробиология. Pseudomonas aeruginosa, род proteus Проект на тему Першозвіри
Проект на тему Першозвіри Семейство крестоцветные КАПУСТА
Семейство крестоцветные КАПУСТА  Генетика пола
Генетика пола ВИТАМИНЫ А и С
ВИТАМИНЫ А и С Инструкционно–технологическая карта на выполнение практической работы по выгонки луковичных растений (лука)
Инструкционно–технологическая карта на выполнение практической работы по выгонки луковичных растений (лука) Частная остеология
Частная остеология Животные и растения тропических лесов
Животные и растения тропических лесов Зәр шығару жүйесі
Зәр шығару жүйесі Аттестационная работа. Образовательная программа внеурочной деятельности Мудрый совенок
Аттестационная работа. Образовательная программа внеурочной деятельности Мудрый совенок