- Нуклеиновые кислоты

Содержание
- 2. Локализация НК
- 3. Потоки информации I. Репликация ДНК Информация удваивается и в процессе деления (митоза или мейоза) передаётся в
- 4. Потоки генетической информации
- 5. Основная догма молекулярной биологии Потоки информации направляются от ДНК к ДНК или определённому белку ДНК ↑
- 6. Функции нуклеиновых кислот 1. Хранение генетической информации на основе генетического кода ДНК. 2. Передача генетической информации
- 7. Пространственная структура и локализация нуклеиновых кислот
- 8. Схема структурной организации ядра 1. Оболочка ядра; 2. Внешняя мембрана; 3. Внутренняя мембрана; 4. Гетерохроматин; 5.
- 9. Хроматин Генетический материал в интерфазном ядре находится в виде хроматиновых нитей. Переплетаясь внутри ядра они образуют
- 10. Организация хроматина Белки представлены основными – гистонами и кислыми (или нейтральными) – негистоновыми белками. Известно пять
- 11. Схема, иллюстрирующая образование нуклеосомной частицы В каждой нуклеосомной частице фрагмент двойной спирали ДНК, имеющий в длину
- 12. Структура нуклеосом
- 13. Хроматин обладает следующими свойствами: - Высокой стабильностью структуры, что обеспечивает постоянство генома из поколение в поколение.
- 14. Хроматин выполняет следующие функции: - Хранение генетической наследственной информации в виде строгой последовательности нуклеотидов ДНК, стабилизованной
- 15. Хромосомы В интерфазном ядре генетический материал представлен хроматином. Хромосомы (от гр. Chroma - цвет, soma -
- 16. Схема нескольких уровней упаковки ДНК, которые являются последовательными этапами формирования плотно конденсированной метафазной хромосомы. 1 –
- 17. Схема основных процессов превращения и распределения генетического материала во время клеточного цикла. G1 – хромосомы представлены
- 19. Митоз Существует сложный механизм, по которому генетический материал в ядре вначале удваивается (репликация ДНК) и затем
- 20. Все циклины делят на 2 подсемейства: G1-циклины (D, Е) и митотические циклины (А и В). Любой
- 21. Циклины и циклинзависимые киназы, регулирующие прохождение клеточного цикла
- 22. Четыре основных элемента последовательностей нуклеотидов ДНК, необходимых для образования и функционирования эукариотических хромосом
- 23. Схема распределение сегментов в каждой из хромосом, составляющих человеческий кариотип на стадии метафазы (первая хроматида) и
- 24. Формы хромосом в период метафазы. а) метацентрическая; б) субметацентрическая; в) акроцентрическая; г) телоцентрическая
- 25. Схема хромосомного цикла
- 26. Разновидность и биологическая роль нуклеиновых кислот
- 27. Сравнительная характеристика нуклеиновых кислот
- 28. Молекулярная масса, Д
- 29. Нуклеиновая кислота (полинуклеотид) + НОН Мононуклеотиды + НОН Нуклеозиды + Н3РО4 + НОН Азотистые основания +
- 30. Структурные компоненты нуклеиновых кислот 1. Азотистые основания: аденин, гуанин, цитозин, урацил, тимин и др. 2. Пентозы:
- 31. Состав нуклеиновых кислот
- 32. Азотистое основание + пентоза --- нуклеозид: Аденин + (дезокси)рибоза ------ аденозин Гуанин + (дезокси)рибоза -------- гуанозин
- 33. Производные пурина
- 34. Производные пиримидина
- 35. Пентозы β-Д-рибоза β-Д-дезоксирибоза
- 41. Физиологически активные мононуклеотиды АМФ, АДФ, АТФ, Г ТФ, ЦТФ, ц АМФ, ц ГМФ, КоА, Флавиномононуклеоид Динуклеотиды
- 43. Пуриновые и пиримидиновые основания
- 44. Номенклатура нуклеотидов
- 45. Пентозы
- 46. Пурин - пиримидиновые пары оснований в ДНК
- 47. Гибридизация нуклеиновых кислот
- 51. Антипараллельность цепей ДНК
- 53. Макромолекула ДНК представляет собой две длинные полимерные цепи, состоящие из мономеров дезоксирибонуклеотидов, прочно соединенных между собой
- 54. Схематическое строение молекулы ДНК
- 55. Нуклеотиды В нуклеотиде пуриновое или пиримидиновое основание связано с дезоксирибозой и остатком фосфорной кислоты. В ДНК
- 56. Правила Э. Чаргаффа Изучив химический состав ДНК в 1950 году, Чаргафф обнаружил, что в молекуле ДНК:
- 58. Схематичес-кое изображе-ние двухце-почечной спиральной структуры молекулы ДНК
- 59. Уровни организации ДНК. В молекуле ДНК можно выделить первичную структуру – последовательность нуклеотидов в цепи, вторичную
- 60. ДНК - это очень длинная (10 мм и более), тонкая, накрученная на нуклеосомы, закрученная в спираль
- 61. Две полинуклеотидные цепи ДНК не являются идентичными, но они комплементарны друг другу. Это связано со строгим
- 62. Организация ДНК в клетке. ДНК в клетке находится в ядре, где организовано в виде хроматина (комплекс
- 63. Свойства ДНК Молекула ДНК обладает некоторыми уникальными свойствами: 1. Она удваивается. Этот процесс называется репликацией. Он
- 64. Комплементарность – строгое соответствие азотистых оснований одной цепи ДНК другой (А – Т, Г – Ц)
- 65. Генетический код – строгая последовательность триплетов (кодонов) в ДНК и м-РНК, контролирует порядок расположения аминокислот в
- 66. 1. Триплетность. 2. Специфичность. 3. Неперекрываемость. 4. Универсальность. 5. Избыточность. 6. Колинеарность. 7. Однонаправленность. 8. Наличие
- 67. Колинеарность – свойство, обусловливающее соответствие между последовательностями триплетов нуклеотидов (кодонов) нуклеиновых кислот и аминокислот полипептидных цепей.
- 69. Кольцевая молекула митохондриальной ДНК
- 70. Общая характеристика РНК У прокариотических и эукариотических организмов наследственная информация закодирована в молекуле ДНК. Однако ДНК,
- 71. РНК – полимер, состоящий из рибонуклеотидов, которые содержат азотистое основание (аденин, гуанин, цитозин, урацил), рибозу и
- 74. Характеристика РНК м-РНК – является матрицей, на которой строится полипептид. Имеют линейную структуру. Содержит 300-3000 нуклеотидов.
- 75. Матричная РНК (мРНК) Молекулы мРНК образуются на специфических участках ДНК, называемых структурными генами. Они несут в
- 76. Процессинг – молекулярные механизмы «созревания» м-РНК ДНК промотор экзон интрон экзон интрон экзон терминатор Транскрипция экзон
- 77. Транспортная РНК (тРНК) Молекулы тРНК образуются на особых генах ДНК. тРНК короткие, однонитевые, имеют форму клеверного
- 80. Строение транспортных РНК
- 81. Молекулы тРНК имеют четыре ключевые области: а) несущий конец. В этом месте к ней присоединяется специфическая
- 82. Рибосомальная РНК (рРНК) Рибосомальная РНК образуется на особых генах ДНК в ядрышке. рРНК - крупная одноцепочечная
- 83. Строение эукариотических и прокариотических рибосом
- 84. Репликация ДНК - сложный, многоступенчатый процесс, требующий вовлечения большого числа специальных белков и ферментов. Скорость репликации
- 85. Общая характеристика процесса репликации Уникальное свойство молекулы ДНК удваиваться перед делением клетки называется репликацией. Это свойство
- 86. Полуконсервативный механизм репликации ДНК. Каждая из двух цепей родительской молекулы ДНК используется в качестве матрицы для
- 87. Полуконсервативная репликация
- 88. Репликация ДНК – удвоение молекулы перед делением клетки. Образуются две дочерние молекулы, идентичные материнской. Каждая состоит
- 89. Инициация репликации Активация дезоксирибонуклеотидов. Монофосфаты дезоксирибонуклеотидов (АМФ, ГМФ, ЦМФ, ТМФ) находятся в состоянии "свободного плавания" в
- 90. Распознавание точки инициации Раскручивание ДНК начинается с определенной точки ее цепи. Такая особая точка называется точкой
- 91. Распознавание точки репликации Репликация обеих цепей двухцепочной ДНК идет одновременно и непрерывно
- 92. Схема процесса репликации ДНК и главные типы белков, действующих в области репликационной вилки
- 93. Образование одиночных нитей ДНК Двойная спираль ДНК раскручивается и разворачивается в одиночные нити ДНК путем разрыва
- 94. Элонгация полинуклеотидной цепи Спаривание оснований. Свободные трифосфаты дезоксирибонуклеотидов своими азотистыми основаниями с помощью ДНК-полимеразы присоединяются водородными
- 95. Элонгация – основной процесс при синтезе новых молекул ДНК. Это добавление дезоксирибонук-леотида к 3' – концу
- 96. Образование новых цепей ДНК Соседствующие нуклеотиды связываются между собой фосфорными остатками и образуют новую цепь ДНК.
- 97. Схема образо-вания прайме-ров второй цепи ДНК и син-теза отста-ющей дочерней цепи
- 98. Образование праймеров. На отстающей нити сначала образуется короткая цепь РНК по шаблону ДНК (рис. 4.10). Она
- 99. Образование двойной спирали. Терминация. Дочерние цепи ДНК образуются отдельными фрагментами по длине хромосомы. Такой отдельный фрагмент
- 100. Значение репликации: а) процесс обеспечивает точное удвоение генетическое информации; б) процесс является важнейшим молекулярным механизмом, лежащим
- 101. Трансляция – перенос генетической информации с РНК на упорядоченую структуру АМ в полипептидной цепи. Синтез белков
- 103. Скачать презентацию
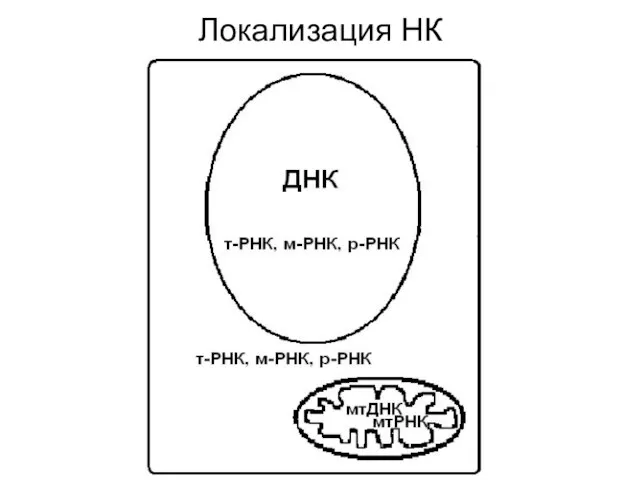
Локализация НК
Локализация НК

Потоки информации
I. Репликация ДНК
Информация удваивается и в процессе деления (митоза или
Потоки информации
I. Репликация ДНК
Информация удваивается и в процессе деления (митоза или
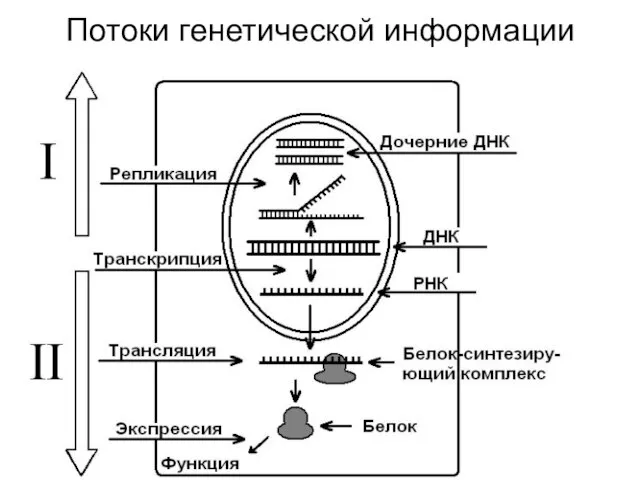
Потоки генетической информации
Потоки генетической информации

Основная догма молекулярной биологии
Потоки информации направляются от ДНК к ДНК или
Основная догма молекулярной биологии
Потоки информации направляются от ДНК к ДНК или

Функции нуклеиновых кислот
1. Хранение генетической информации на основе генетического кода ДНК.
2.
Функции нуклеиновых кислот
1. Хранение генетической информации на основе генетического кода ДНК.
2.

Пространственная структура и локализация нуклеиновых кислот
Пространственная структура и локализация нуклеиновых кислот
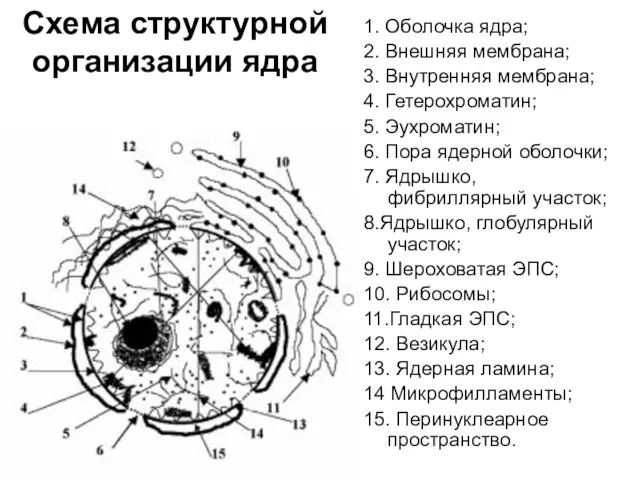
Схема структурной организации ядра
1. Оболочка ядра;
2. Внешняя мембрана;
3. Внутренняя
Схема структурной организации ядра
1. Оболочка ядра;
2. Внешняя мембрана;
3. Внутренняя

Хроматин
Генетический материал в интерфазном ядре находится в виде хроматиновых нитей. Переплетаясь
Хроматин
Генетический материал в интерфазном ядре находится в виде хроматиновых нитей. Переплетаясь

Организация хроматина
Белки представлены основными – гистонами и кислыми (или нейтральными) –
Организация хроматина
Белки представлены основными – гистонами и кислыми (или нейтральными) –
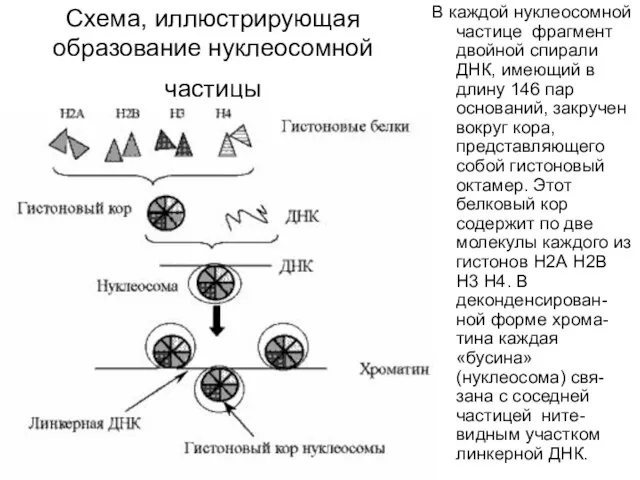
Схема, иллюстрирующая образование нуклеосомной частицы
В каждой нуклеосомной частице фрагмент двойной
Схема, иллюстрирующая образование нуклеосомной частицы
В каждой нуклеосомной частице фрагмент двойной
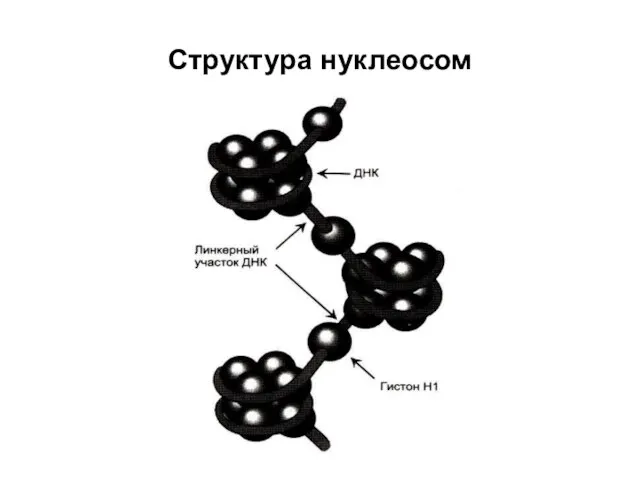
Структура нуклеосом
Структура нуклеосом

Хроматин обладает следующими свойствами:
- Высокой стабильностью структуры, что обеспечивает постоянство генома
Хроматин обладает следующими свойствами:
- Высокой стабильностью структуры, что обеспечивает постоянство генома

Хроматин выполняет следующие функции:
- Хранение генетической наследственной информации в виде строгой
Хроматин выполняет следующие функции:
- Хранение генетической наследственной информации в виде строгой

Хромосомы
В интерфазном ядре генетический материал представлен хроматином.
Хромосомы (от гр. Chroma
Хромосомы
В интерфазном ядре генетический материал представлен хроматином.
Хромосомы (от гр. Chroma

Схема нескольких уровней упаковки ДНК, которые являются последовательными этапами формирования плотно
Схема нескольких уровней упаковки ДНК, которые являются последовательными этапами формирования плотно

Схема основных процессов превращения и распределения генетического материала во время клеточного
Схема основных процессов превращения и распределения генетического материала во время клеточного
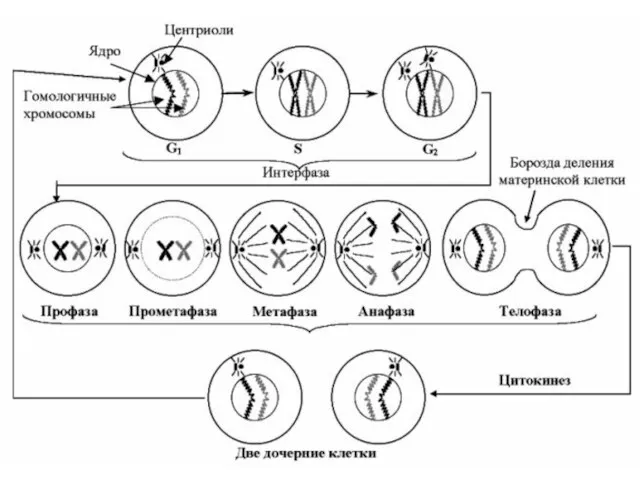

Митоз
Существует сложный механизм, по которому генетический материал в ядре вначале удваивается
Митоз
Существует сложный механизм, по которому генетический материал в ядре вначале удваивается

Все циклины делят на 2 подсемейства: G1-циклины (D, Е) и митотические
Все циклины делят на 2 подсемейства: G1-циклины (D, Е) и митотические
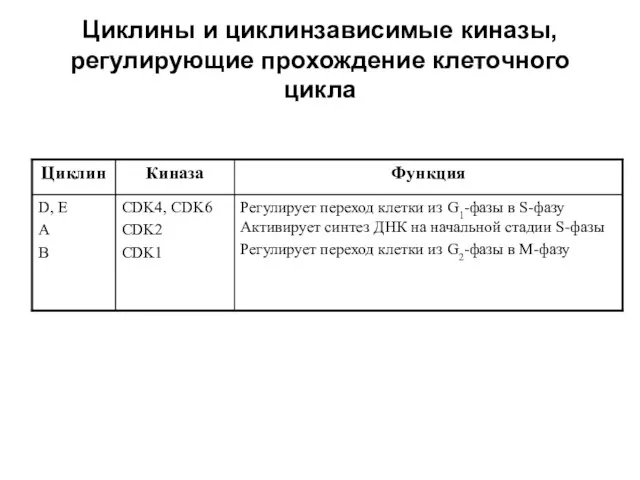
Циклины и циклинзависимые киназы, регулирующие прохождение клеточного цикла
Циклины и циклинзависимые киназы, регулирующие прохождение клеточного цикла
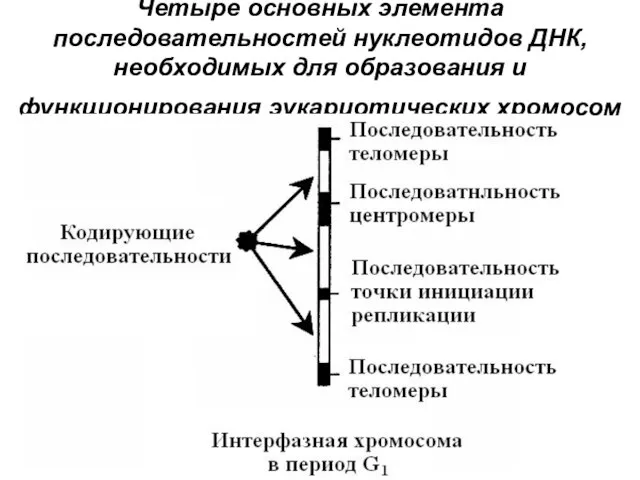
Четыре основных элемента последовательностей нуклеотидов ДНК, необходимых для образования и функционирования
Четыре основных элемента последовательностей нуклеотидов ДНК, необходимых для образования и функционирования

Схема распределение сегментов в каждой из хромосом, составляющих человеческий кариотип на
Схема распределение сегментов в каждой из хромосом, составляющих человеческий кариотип на
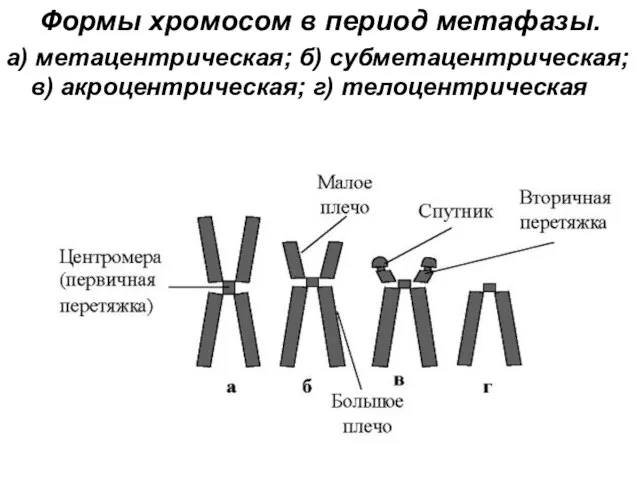
Формы хромосом в период метафазы.
а) метацентрическая; б) субметацентрическая; в) акроцентрическая;
Формы хромосом в период метафазы.
а) метацентрическая; б) субметацентрическая; в) акроцентрическая;
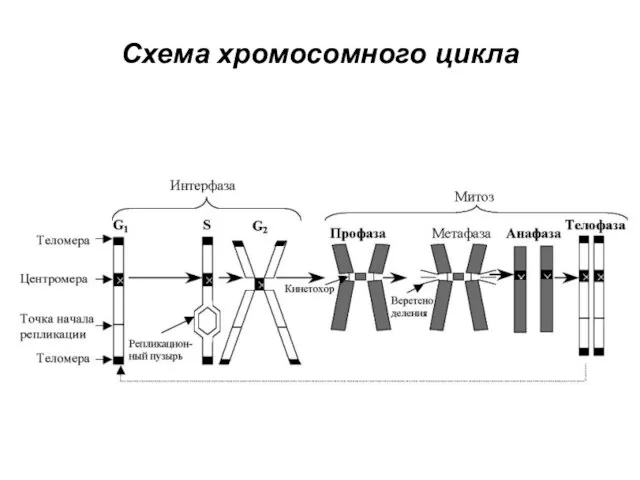
Схема хромосомного цикла
Схема хромосомного цикла
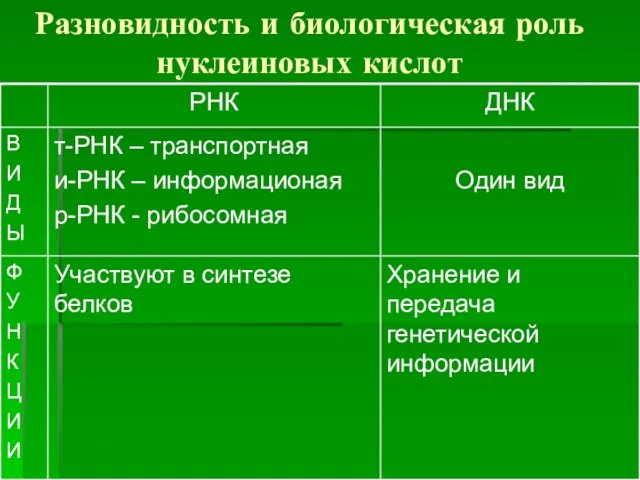
Разновидность и биологическая роль нуклеиновых кислот
Разновидность и биологическая роль нуклеиновых кислот
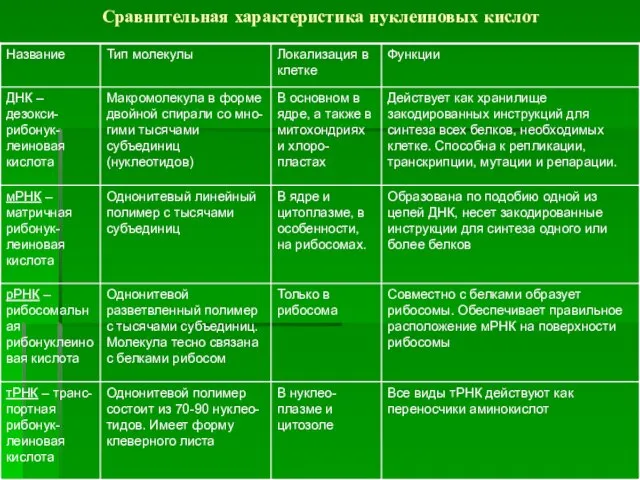
Сравнительная характеристика нуклеиновых кислот
Сравнительная характеристика нуклеиновых кислот

Молекулярная масса, Д
Молекулярная масса, Д

Нуклеиновая кислота
(полинуклеотид)
+ НОН
Мононуклеотиды
+ НОН
Нуклеозиды + Н3РО4
+ НОН
Азотистые основания
+
пентозы
Нуклеиновая кислота
(полинуклеотид)
+ НОН
Мононуклеотиды
+ НОН
Нуклеозиды + Н3РО4
+ НОН
Азотистые основания
+
пентозы

Структурные компоненты нуклеиновых кислот
1. Азотистые основания: аденин, гуанин, цитозин, урацил, тимин
Структурные компоненты нуклеиновых кислот
1. Азотистые основания: аденин, гуанин, цитозин, урацил, тимин
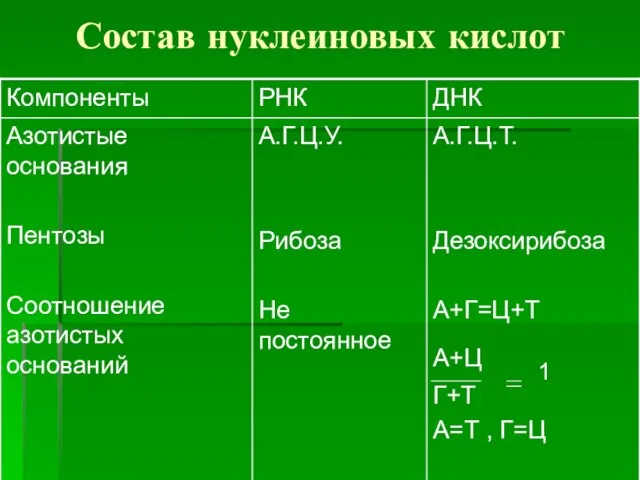
Состав нуклеиновых кислот
Состав нуклеиновых кислот

Азотистое основание + пентоза --- нуклеозид:
Аденин + (дезокси)рибоза ------ аденозин
Гуанин +
Азотистое основание + пентоза --- нуклеозид:
Аденин + (дезокси)рибоза ------ аденозин
Гуанин +
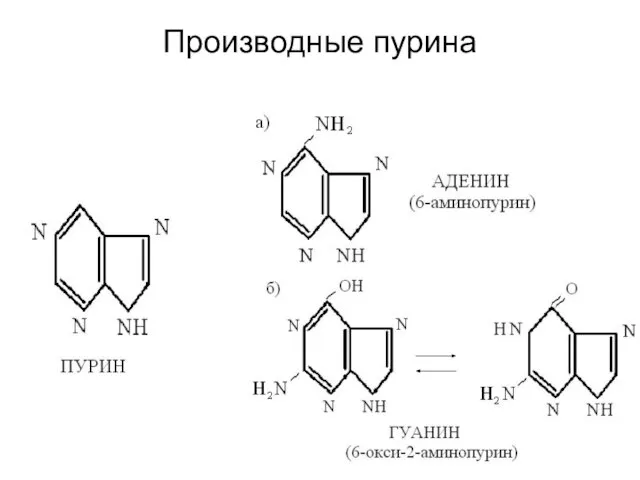
Производные пурина
Производные пурина
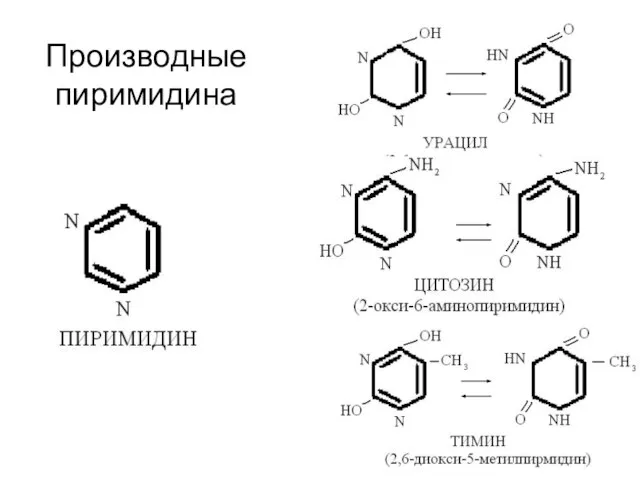
Производные
пиримидина
Производные
пиримидина
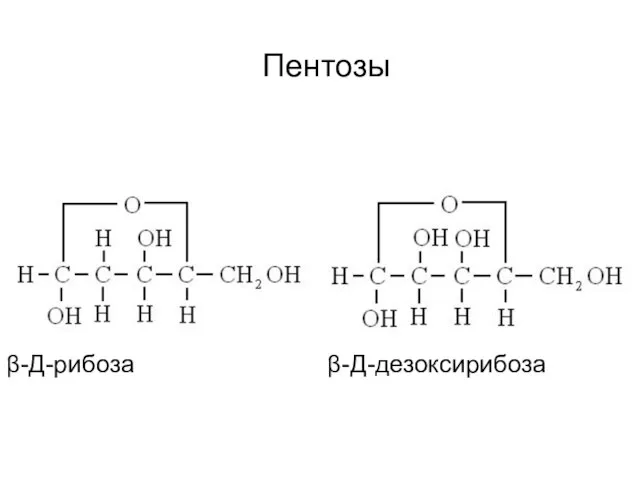
Пентозы
β-Д-рибоза β-Д-дезоксирибоза
Пентозы
β-Д-рибоза β-Д-дезоксирибоза



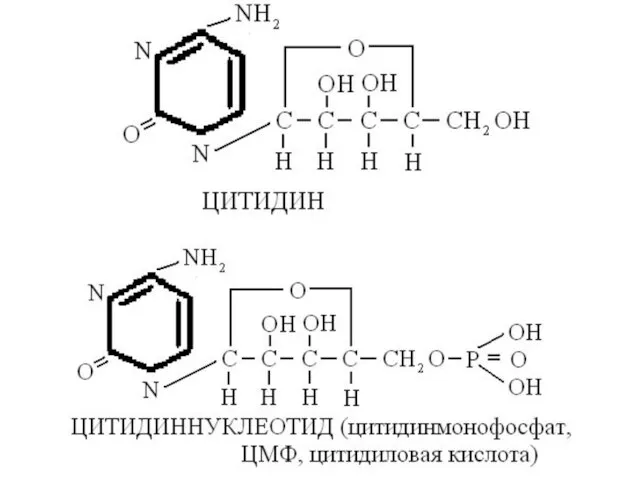


Физиологически активные мононуклеотиды
АМФ, АДФ, АТФ, Г ТФ, ЦТФ, ц АМФ, ц
Физиологически активные мононуклеотиды
АМФ, АДФ, АТФ, Г ТФ, ЦТФ, ц АМФ, ц

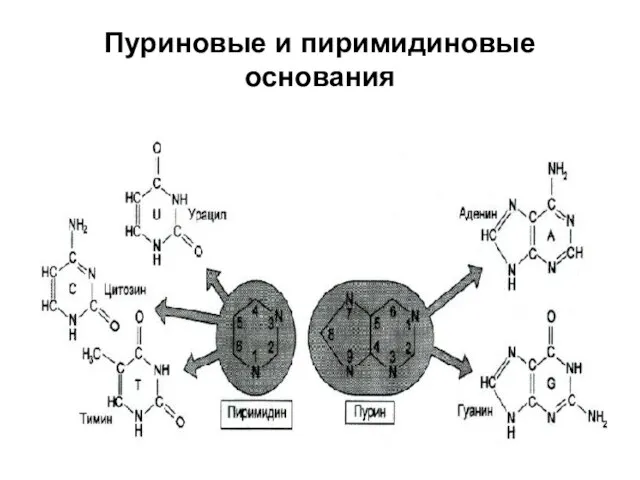
Пуриновые и пиримидиновые основания
Пуриновые и пиримидиновые основания
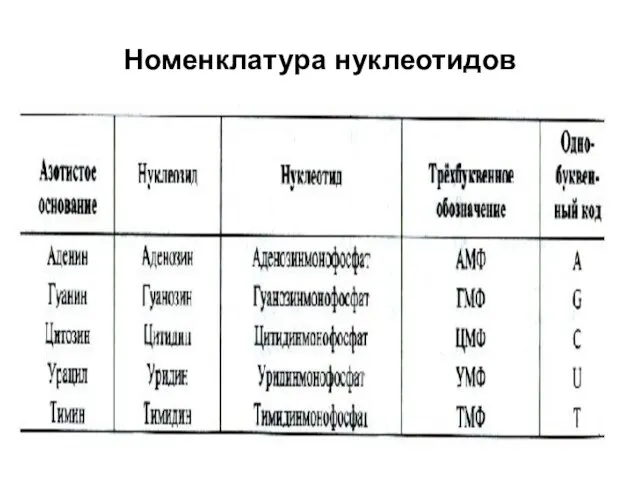
Номенклатура нуклеотидов
Номенклатура нуклеотидов
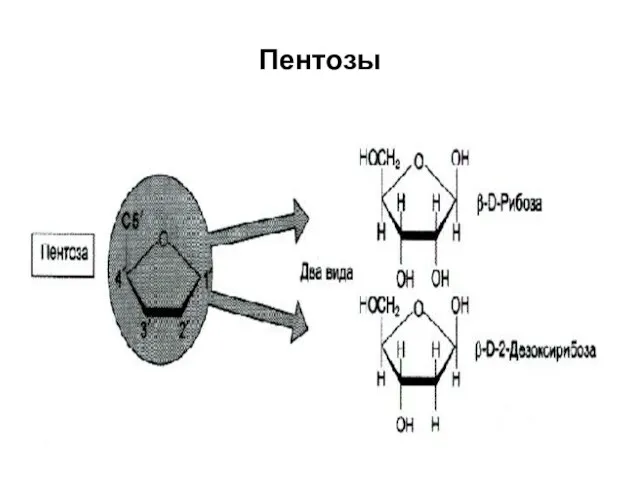
Пентозы
Пентозы
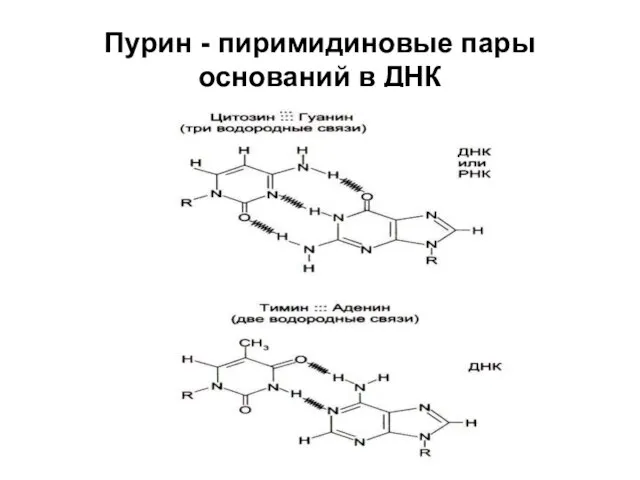
Пурин - пиримидиновые пары оснований в ДНК
Пурин - пиримидиновые пары оснований в ДНК
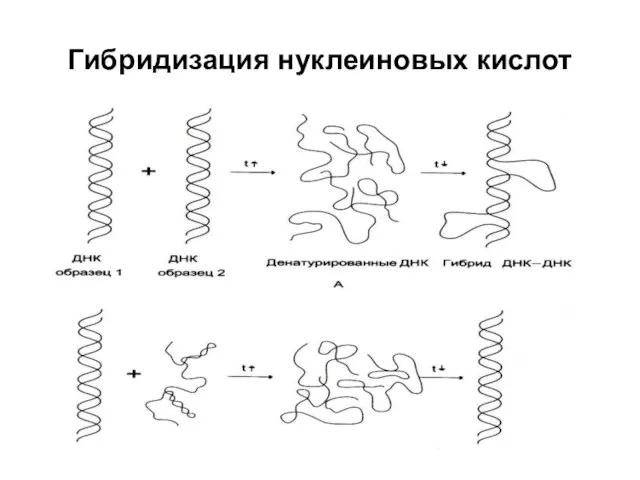
Гибридизация нуклеиновых кислот
Гибридизация нуклеиновых кислот

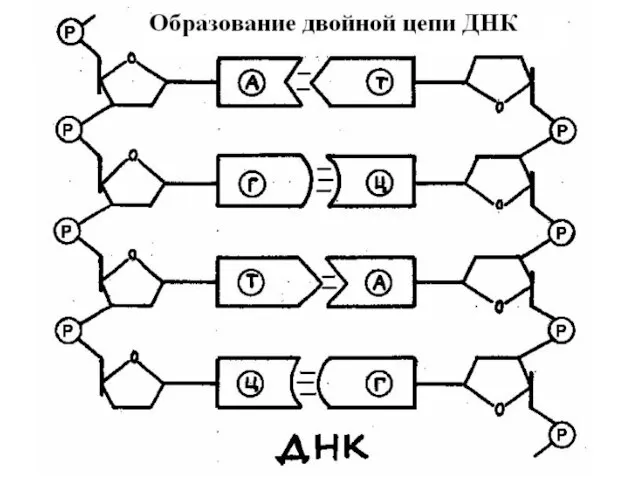

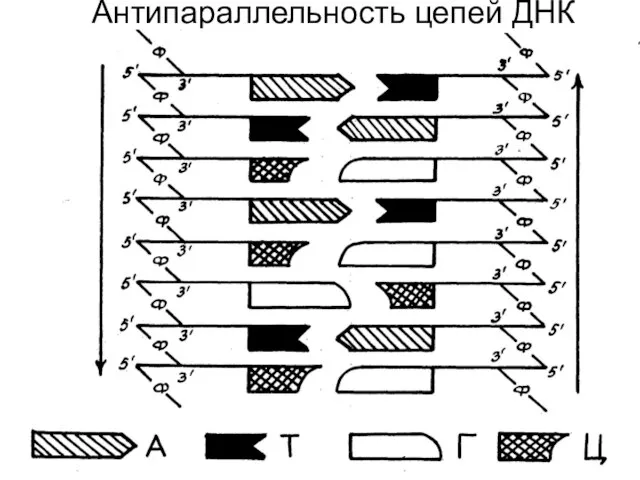
Антипараллельность цепей ДНК
Антипараллельность цепей ДНК


Макромолекула ДНК представляет собой две длинные полимерные цепи, состоящие из мономеров
Макромолекула ДНК представляет собой две длинные полимерные цепи, состоящие из мономеров
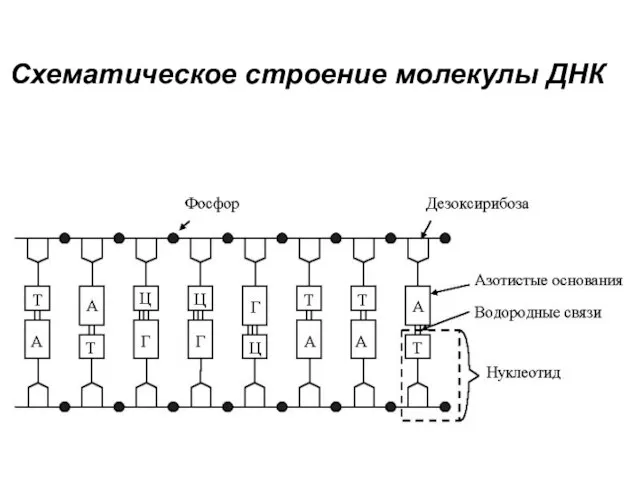
Схематическое строение молекулы ДНК
Схематическое строение молекулы ДНК

Нуклеотиды
В нуклеотиде пуриновое или пиримидиновое основание связано с дезоксирибозой и
Нуклеотиды
В нуклеотиде пуриновое или пиримидиновое основание связано с дезоксирибозой и

Правила Э. Чаргаффа
Изучив химический состав ДНК в 1950 году, Чаргафф
Правила Э. Чаргаффа
Изучив химический состав ДНК в 1950 году, Чаргафф
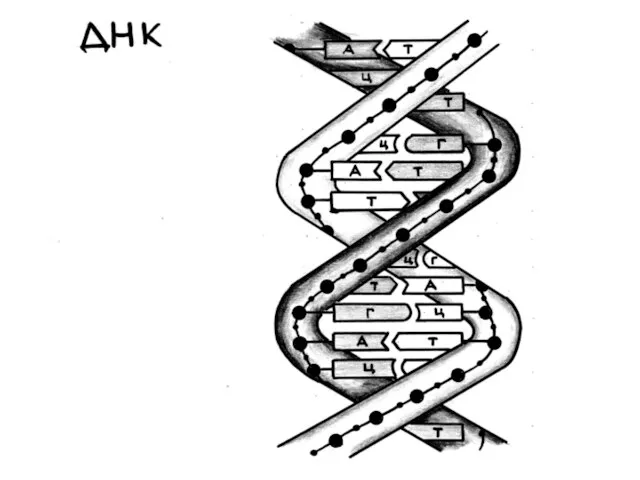
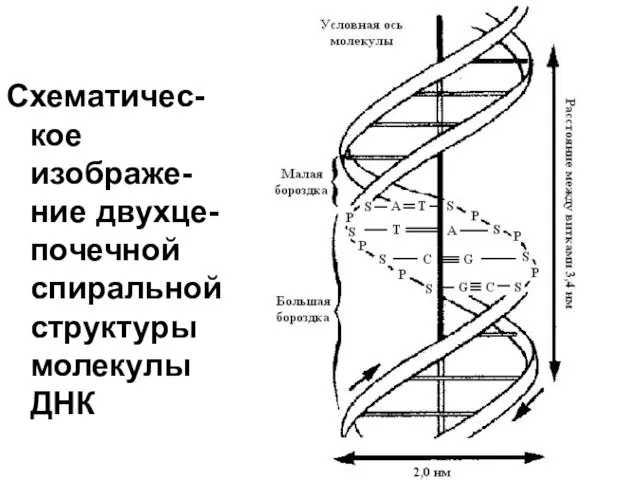
Схематичес-кое изображе-ние двухце-почечной спиральной структуры молекулы ДНК
Схематичес-кое изображе-ние двухце-почечной спиральной структуры молекулы ДНК

Уровни организации ДНК.
В молекуле ДНК можно выделить первичную структуру –
Уровни организации ДНК.
В молекуле ДНК можно выделить первичную структуру –

ДНК - это очень длинная (10 мм и более), тонкая, накрученная
ДНК - это очень длинная (10 мм и более), тонкая, накрученная

Две полинуклеотидные цепи ДНК не являются идентичными, но они комплементарны друг
Две полинуклеотидные цепи ДНК не являются идентичными, но они комплементарны друг

Организация ДНК в клетке.
ДНК в клетке находится в ядре, где организовано
Организация ДНК в клетке.
ДНК в клетке находится в ядре, где организовано

Свойства ДНК
Молекула ДНК обладает некоторыми уникальными свойствами:
1. Она удваивается. Этот
Свойства ДНК
Молекула ДНК обладает некоторыми уникальными свойствами:
1. Она удваивается. Этот

Комплементарность
– строгое соответствие азотистых оснований одной цепи ДНК другой (А
Комплементарность
– строгое соответствие азотистых оснований одной цепи ДНК другой (А

Генетический код
– строгая последовательность триплетов (кодонов) в ДНК и м-РНК,
Генетический код
– строгая последовательность триплетов (кодонов) в ДНК и м-РНК,

1. Триплетность.
2. Специфичность.
3. Неперекрываемость.
4. Универсальность.
5. Избыточность.
6. Колинеарность.
7. Однонаправленность.
8. Наличие «старт»
1. Триплетность.
2. Специфичность.
3. Неперекрываемость.
4. Универсальность.
5. Избыточность.
6. Колинеарность.
7. Однонаправленность.
8. Наличие «старт»

Колинеарность – свойство, обусловливающее соответствие между последовательностями триплетов нуклеотидов (кодонов) нуклеиновых
Колинеарность – свойство, обусловливающее соответствие между последовательностями триплетов нуклеотидов (кодонов) нуклеиновых
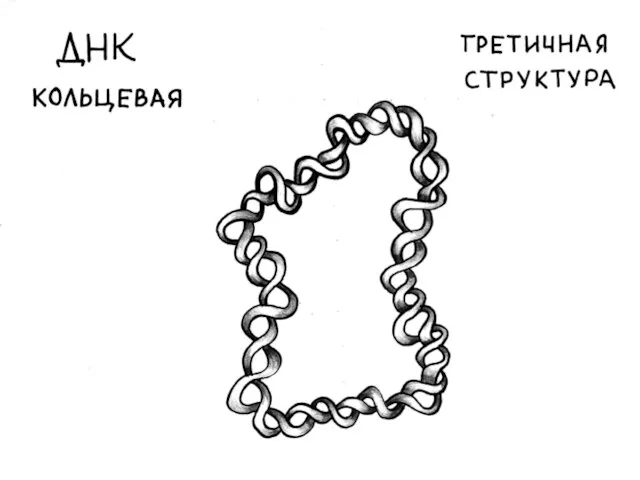
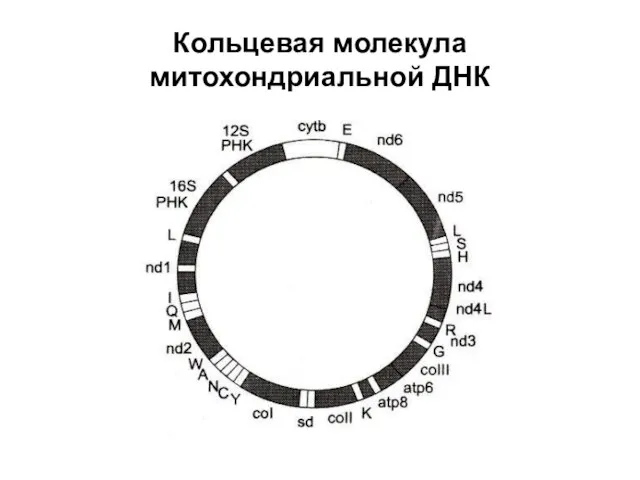
Кольцевая молекула митохондриальной ДНК
Кольцевая молекула митохондриальной ДНК

Общая характеристика РНК
У прокариотических и эукариотических организмов наследственная информация закодирована в
Общая характеристика РНК
У прокариотических и эукариотических организмов наследственная информация закодирована в

РНК – полимер, состоящий из рибонуклеотидов, которые содержат азотистое основание (аденин,
РНК – полимер, состоящий из рибонуклеотидов, которые содержат азотистое основание (аденин,
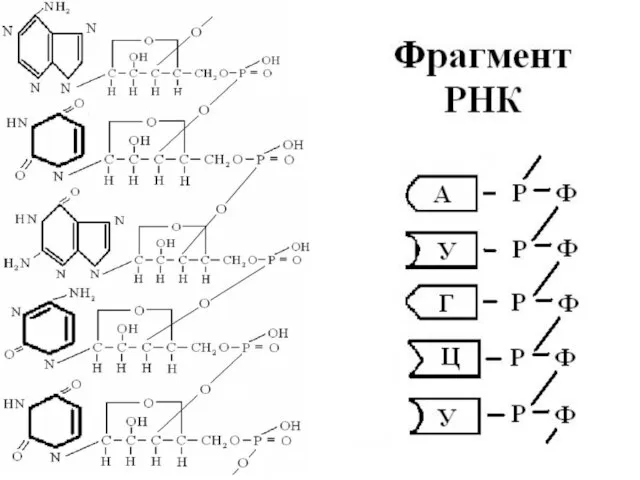
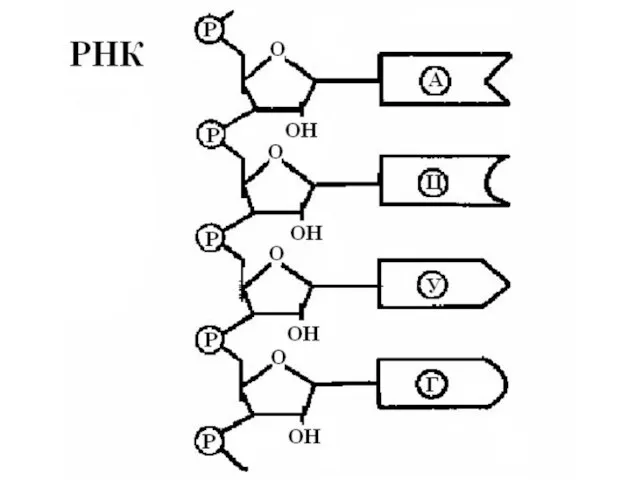

Характеристика РНК
м-РНК – является матрицей, на которой строится полипептид. Имеют линейную
Характеристика РНК
м-РНК – является матрицей, на которой строится полипептид. Имеют линейную

Матричная РНК (мРНК)
Молекулы мРНК образуются на специфических участках ДНК, называемых
Матричная РНК (мРНК)
Молекулы мРНК образуются на специфических участках ДНК, называемых

Процессинг – молекулярные механизмы «созревания» м-РНК ДНК
промотор экзон интрон экзон интрон
Процессинг – молекулярные механизмы «созревания» м-РНК ДНК
промотор экзон интрон экзон интрон

Транспортная РНК (тРНК)
Молекулы тРНК образуются на особых генах ДНК. тРНК короткие,
Транспортная РНК (тРНК)
Молекулы тРНК образуются на особых генах ДНК. тРНК короткие,

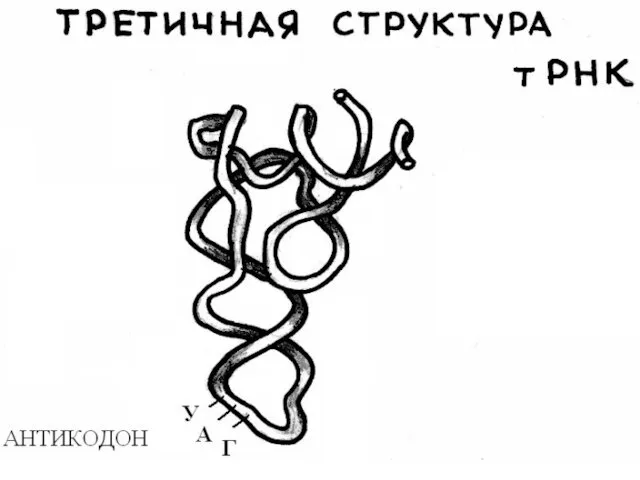
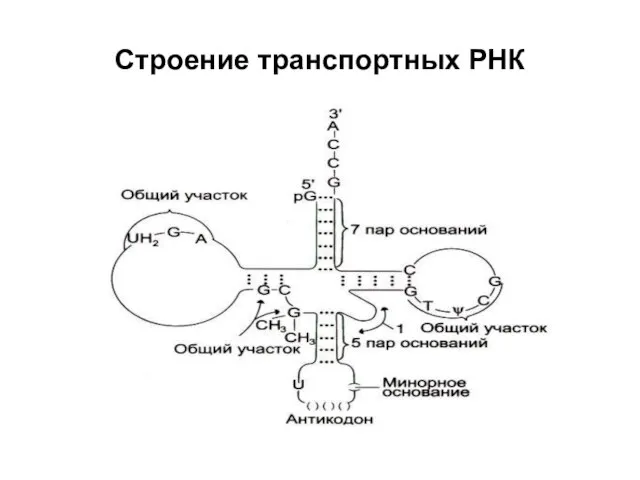
Строение транспортных РНК
Строение транспортных РНК

Молекулы тРНК имеют четыре ключевые области:
а) несущий конец. В этом
Молекулы тРНК имеют четыре ключевые области:
а) несущий конец. В этом

Рибосомальная РНК (рРНК)
Рибосомальная РНК образуется на особых генах ДНК в
Рибосомальная РНК (рРНК)
Рибосомальная РНК образуется на особых генах ДНК в
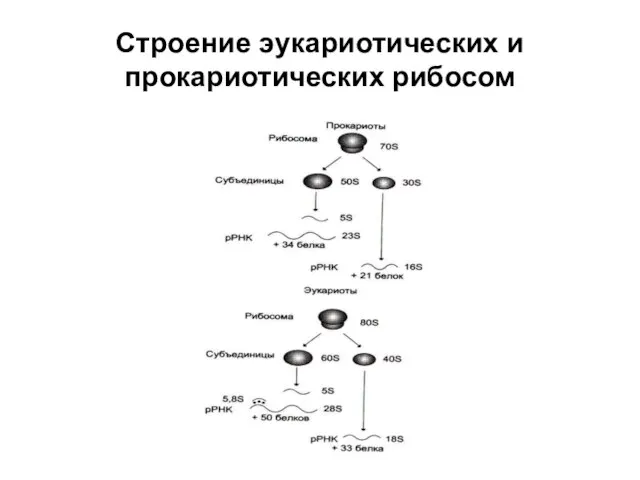
Строение эукариотических и прокариотических рибосом
Строение эукариотических и прокариотических рибосом

Репликация ДНК - сложный, многоступенчатый процесс, требующий вовлечения большого числа специальных
Репликация ДНК - сложный, многоступенчатый процесс, требующий вовлечения большого числа специальных

Общая характеристика процесса репликации
Уникальное свойство молекулы ДНК удваиваться перед делением клетки
Общая характеристика процесса репликации
Уникальное свойство молекулы ДНК удваиваться перед делением клетки
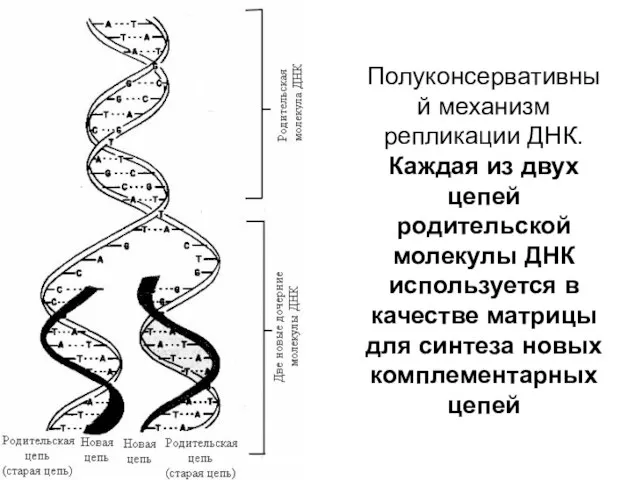
Полуконсервативный механизм репликации ДНК.
Каждая из двух цепей родительской молекулы ДНК
Полуконсервативный механизм репликации ДНК. Каждая из двух цепей родительской молекулы ДНК
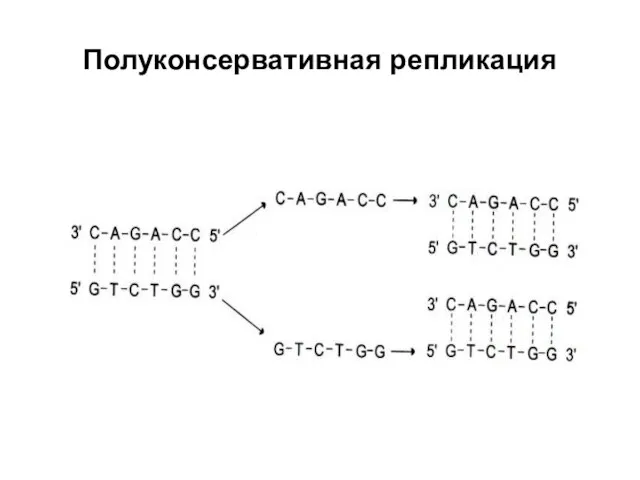
Полуконсервативная репликация
Полуконсервативная репликация

Репликация ДНК – удвоение молекулы перед делением клетки.
Образуются две дочерние молекулы,
Репликация ДНК – удвоение молекулы перед делением клетки.
Образуются две дочерние молекулы,

Инициация репликации
Активация дезоксирибонуклеотидов. Монофосфаты дезоксирибонуклеотидов (АМФ, ГМФ, ЦМФ, ТМФ) находятся в
Инициация репликации
Активация дезоксирибонуклеотидов. Монофосфаты дезоксирибонуклеотидов (АМФ, ГМФ, ЦМФ, ТМФ) находятся в

Распознавание точки инициации
Раскручивание ДНК начинается с определенной точки ее цепи.
Распознавание точки инициации
Раскручивание ДНК начинается с определенной точки ее цепи.
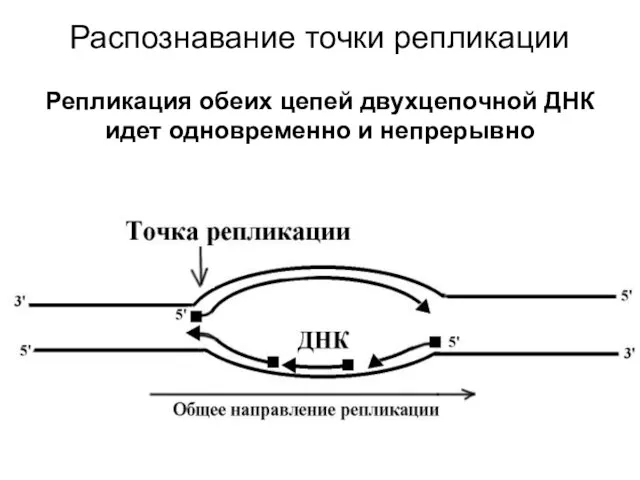
Распознавание точки репликации
Репликация обеих цепей двухцепочной ДНК идет одновременно и
Распознавание точки репликации Репликация обеих цепей двухцепочной ДНК идет одновременно и
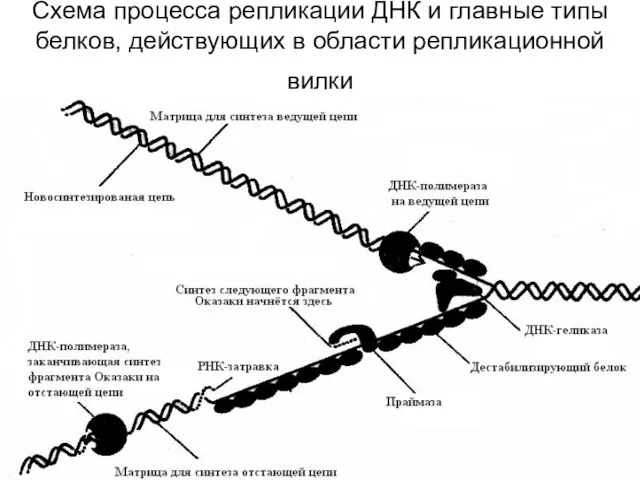
Схема процесса репликации ДНК и главные типы белков, действующих в области
Схема процесса репликации ДНК и главные типы белков, действующих в области

Образование одиночных нитей ДНК
Двойная спираль ДНК раскручивается и разворачивается в одиночные
Образование одиночных нитей ДНК
Двойная спираль ДНК раскручивается и разворачивается в одиночные

Элонгация полинуклеотидной цепи
Спаривание оснований. Свободные трифосфаты дезоксирибонуклеотидов своими азотистыми основаниями с
Элонгация полинуклеотидной цепи
Спаривание оснований. Свободные трифосфаты дезоксирибонуклеотидов своими азотистыми основаниями с
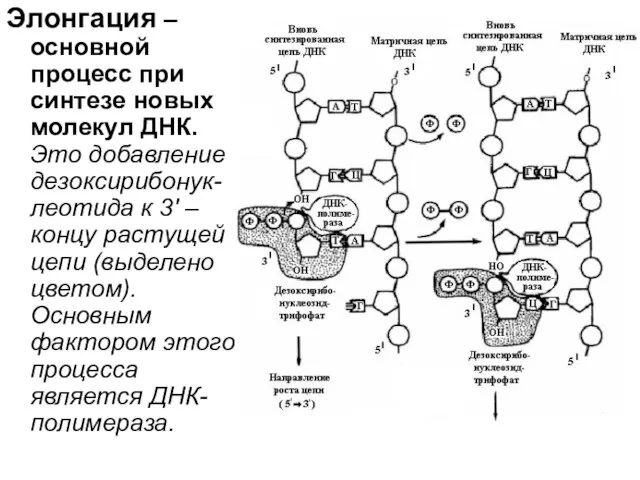
Элонгация – основной процесс при синтезе новых молекул ДНК. Это добавление
Элонгация – основной процесс при синтезе новых молекул ДНК. Это добавление

Образование новых цепей ДНК
Соседствующие нуклеотиды связываются между собой фосфорными остатками
Образование новых цепей ДНК
Соседствующие нуклеотиды связываются между собой фосфорными остатками
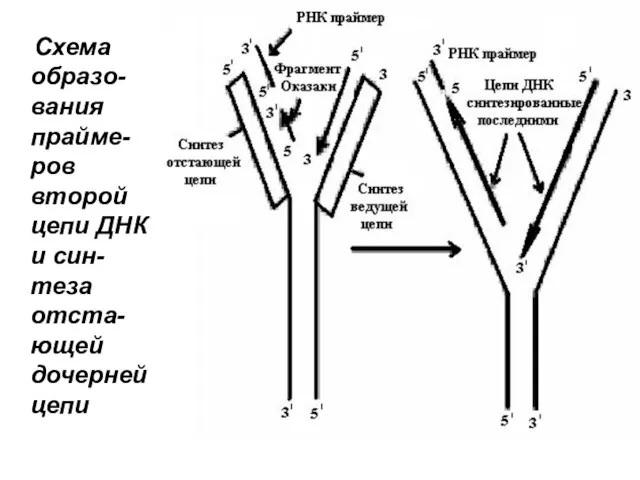
Схема образо-вания прайме-ров второй цепи ДНК и син-теза отста-ющей дочерней
Схема образо-вания прайме-ров второй цепи ДНК и син-теза отста-ющей дочерней

Образование праймеров. На отстающей нити сначала образуется короткая цепь РНК по
Образование праймеров. На отстающей нити сначала образуется короткая цепь РНК по

Образование двойной спирали. Терминация.
Дочерние цепи ДНК образуются отдельными фрагментами по
Образование двойной спирали. Терминация.
Дочерние цепи ДНК образуются отдельными фрагментами по

Значение репликации:
а) процесс обеспечивает точное удвоение генетическое информации;
б)
Значение репликации:
а) процесс обеспечивает точное удвоение генетическое информации;
б)

Трансляция – перенос генетической информации с РНК на упорядоченую структуру АМ
Трансляция – перенос генетической информации с РНК на упорядоченую структуру АМ
 Концепция эволюции в биологии.
Концепция эволюции в биологии. Костная система
Костная система Особенности строения и жизнедеятельности рыб
Особенности строения и жизнедеятельности рыб Презентация на тему Строение корневища Молодой побег
Презентация на тему Строение корневища Молодой побег Размножение алоэ
Размножение алоэ Репродуктивні органи рослин. Квітка
Репродуктивні органи рослин. Квітка Презентация на тему Осторожно клещи
Презентация на тему Осторожно клещи Why do leaves fall in autumn
Why do leaves fall in autumn Органы чувств
Органы чувств Презентация на тему Отряд Насекомоядные
Презентация на тему Отряд Насекомоядные  Презентация на тему "Орган вкуса" - скачать презентации по Биологии
Презентация на тему "Орган вкуса" - скачать презентации по Биологии Презентация на тему "ткани человека" - скачать бесплатно презентации по Биологии
Презентация на тему "ткани человека" - скачать бесплатно презентации по Биологии Лекция 3 Размножение на клеточном и организменном уровне
Лекция 3 Размножение на клеточном и организменном уровне Circulation and Gas Exchange. Chapter 42
Circulation and Gas Exchange. Chapter 42 Путешествие по эукариотической клетке: урок-игра
Путешествие по эукариотической клетке: урок-игра Презентация по биологии Внутренняя среда организма
Презентация по биологии Внутренняя среда организма  Neurology. Spinal cord compression
Neurology. Spinal cord compression Микропрепараты. Растительные ткани
Микропрепараты. Растительные ткани Тема: «Изучение зависимости урожайности картофеля от разных способов посадки»
Тема: «Изучение зависимости урожайности картофеля от разных способов посадки» Методы электронной микроскопии
Методы электронной микроскопии Презентация на тему "Різноманітність птахів" - скачать бесплатно презентации по Биологии
Презентация на тему "Різноманітність птахів" - скачать бесплатно презентации по Биологии Братья наши меньшие. Весёлая энцинклопедия для детей
Братья наши меньшие. Весёлая энцинклопедия для детей Ученые-ботаники, исследователи Донского и Азово-Черноморского региона
Ученые-ботаники, исследователи Донского и Азово-Черноморского региона Презентация на тему "Энергетический обмен" - скачать презентации по Биологии
Презентация на тему "Энергетический обмен" - скачать презентации по Биологии Synthetic theory of evolution
Synthetic theory of evolution Обеспечение жизнедеятельности человека при автономном существовании в природных условиях
Обеспечение жизнедеятельности человека при автономном существовании в природных условиях Стебель, его строение и значение
Стебель, его строение и значение Презентация по биологии Внутренняя среда
Презентация по биологии Внутренняя среда