- Обмен нуклеотидов. Строение мононуклеотида

Содержание
- 2. История открытия нуклеиновых кислот и доказательство их генетической роли В 1869 г. швейцарский биохимик Иоганн Фридрих
- 3. Нуклеоти́ды (нуклеозидфосфаты) фосфорные эфиры нуклеозидов. Свободные нуклеотиды, в частности АТФ, цАМФ, АДФ, играют важную роль в
- 4. Классификация нуклеотидов По характеру углевода-пентозы они могут быть рибонуклеотидами ( содержат рибозу ) или же дезоксирибонуклеотидами
- 5. ПИРИМИДИНОВЫЕ ОСНОВАНИЯ НУКЛЕИНОВЫХ КИСЛОТ 5
- 6. УГЛЕВОДЫ НУКЛЕОТИДОВ 6
- 7. По частоте встречаемости в составе нуклеиновых кислот ГЛАВНЫЕ МИНОРНЫЕ К минорным нуклеотидам относятся те нуклеотиды, количество
- 8. Некоторые минорные (модифицированные) азотистые основания. 8
- 9. Пуриновые и пиримидиновые основания, входящие в молекул нуклеиновых кислот 9
- 10. Образование дезоксирибонуклеотида путём соединения фосфата, дезоксирибозы и азотистого основания. 10
- 11. Схема строения рибонуклеотида 11
- 12. Биологическая роль нуклеотидов Рибонуклеотиды пуринового или пиримидинового рядов (АМФ, ГМФ,УМФ и ЦМФ и их минорные производные)
- 13. Гидролиз нуклеопротеинов 13
- 14. БИОСИНТЕЗ ПУРИНОВЫХ НУКЛЕОТИДОВ Происхождение атомов С и N в пуриновом основании 14
- 15. СИНТЕЗ 5-ФОСФОРИБОЗИЛ-1-АМИНА И ОБРАЗОВАНИЕ ИМФ СКОРОСТЬ - ЛИМИТИРУЮЩЕЙ И РЕГУЛЯТОРНОЙ СТАДИЕЙ ПРОЦЕССА ЯВЛЯЕТСЯ ОБРАЗОВАНИЕ 5-ФОСФОРИБОЗИЛ-1-АМИНА ,
- 16. Синтез АМФ и ГМФ из ИМФ 16
- 17. ЗАПАСНЫЕ ПУТИ СИНТЕЗА ПУРИНОВЫХ НУКЛЕОТИДОВ Потребность в большом количестве нуклеотидов привела к развитию «запасных» путей синтеза
- 18. ПРОИСХОЖДЕНИЕ АТОМОВ ПИРИМИДИНОВОГО КОЛЬЦА И СИНТЕЗ УМФ 18
- 19. Регуляторным ферментом в синтезе пиримидиновых нуклеотидов является полифункциональный КАД-фермент, который катализирует реакции 1, 2, 3; Дигидрооротатдегидрогеназа
- 20. 20
- 21. ПЕРЕВАРИВАНИЕ НУКЛЕИНОВЫХ КИСЛОТ ПИЩИ В ЖЕЛУДОЧНО-КИШЕЧНОМ ТРАКТЕ 21
- 22. КАТАБОЛИЗМ ПУРИНОВЫХ НУКЛЕОТИДОВ 22
- 23. Катаболизм пуриновых нуклеотидов до мочевой кислоты. 23
- 24. КАТАБОЛИЗМ ПУРИНОВЫХ ОСНОВАНИЙ 24
- 25. Мочевая кислота - основной продукт катаболизма пуриновых нуклеотидов у человека. 25
- 26. ГИПЕРУРИКЕМИЯ И ПОДАГРА Когда в плазме крови концентрация мочевой кислоты превышает норму, то возникает гиперурикемия. Вследствие
- 27. Как правило, подагра генетически детерминирована и носит семейный характер. Она вызвана нарушениями в работе ФРДФ синтетазы
- 28. Гиперурикемическая (подагрическая) стимуляция умственной активности Свою разгадку повышенная частота подагриков среди гениев нашла в 1955 году
- 29. Первым подагриком, зарегистрированным в истории, был Иудейский царь, мудрый Аза. Подагрой болели многие греческие вожди, участвовавшие
- 30. Эфроимсон приводит следующую статистику: «крупные выборки гениев и выдающихся талантов дают цифру 5-10% (подагриков), малые выборки
- 31. Наследственное заболевание, характеризующееся увеличением синтеза мочевой кислоты (у детей) вызванное дефектом фермента гипоксантин-гуанинфосфорибозилтрансферазы, который катализирует реутилизацию
- 33. Скачать презентацию

История открытия нуклеиновых кислот и доказательство их генетической роли
В 1869 г.
История открытия нуклеиновых кислот и доказательство их генетической роли
В 1869 г.
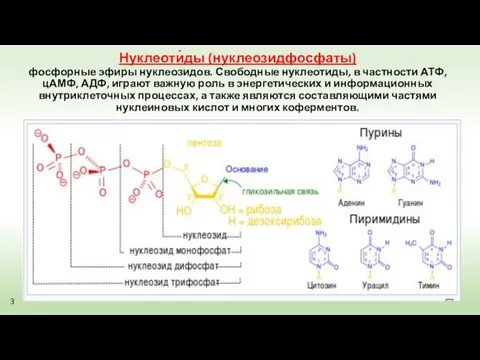
Нуклеоти́ды (нуклеозидфосфаты)
фосфорные эфиры нуклеозидов. Свободные нуклеотиды, в частности АТФ, цАМФ,
Нуклеоти́ды (нуклеозидфосфаты) фосфорные эфиры нуклеозидов. Свободные нуклеотиды, в частности АТФ, цАМФ,

Классификация нуклеотидов
По характеру углевода-пентозы они могут быть рибонуклеотидами ( содержат рибозу
Классификация нуклеотидов
По характеру углевода-пентозы они могут быть рибонуклеотидами ( содержат рибозу
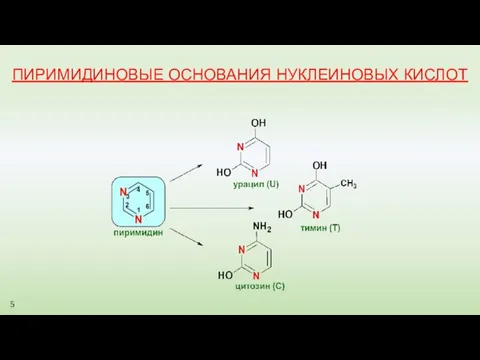
ПИРИМИДИНОВЫЕ ОСНОВАНИЯ НУКЛЕИНОВЫХ КИСЛОТ
5
ПИРИМИДИНОВЫЕ ОСНОВАНИЯ НУКЛЕИНОВЫХ КИСЛОТ
5
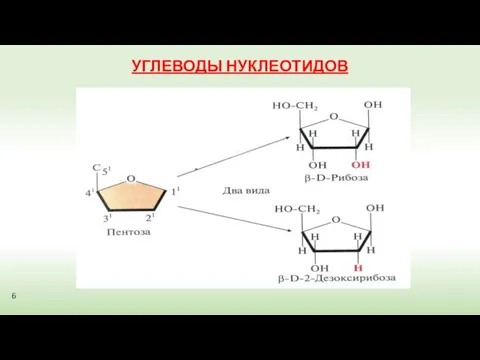
УГЛЕВОДЫ НУКЛЕОТИДОВ
6
УГЛЕВОДЫ НУКЛЕОТИДОВ
6

По частоте встречаемости в составе нуклеиновых кислот
ГЛАВНЫЕ
МИНОРНЫЕ
К минорным нуклеотидам относятся
По частоте встречаемости в составе нуклеиновых кислот
ГЛАВНЫЕ
МИНОРНЫЕ
К минорным нуклеотидам относятся
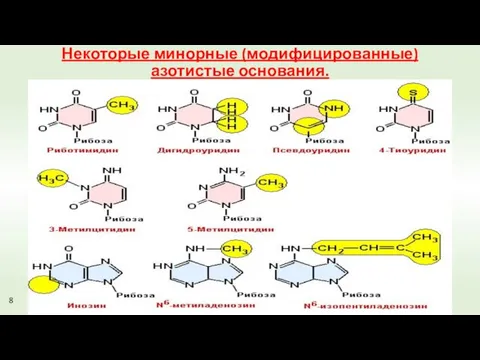
Некоторые минорные (модифицированные) азотистые основания.
8
Некоторые минорные (модифицированные) азотистые основания.
8
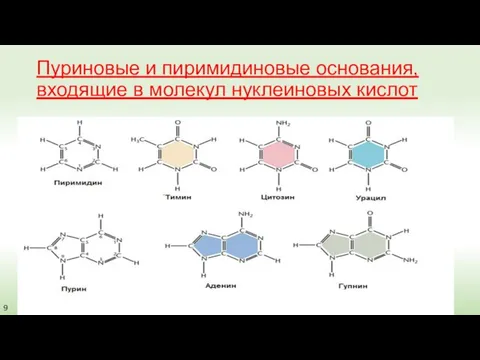
Пуриновые и пиримидиновые основания, входящие в молекул нуклеиновых кислот
9
Пуриновые и пиримидиновые основания, входящие в молекул нуклеиновых кислот
9
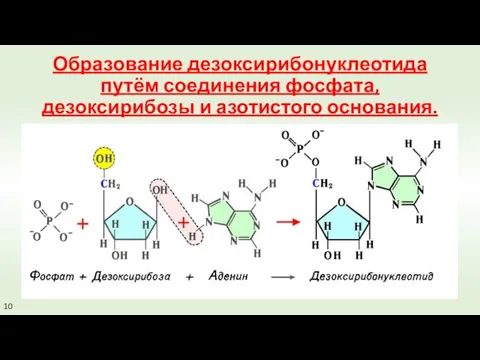
Образование дезоксирибонуклеотида путём соединения фосфата, дезоксирибозы и азотистого основания.
10
Образование дезоксирибонуклеотида путём соединения фосфата, дезоксирибозы и азотистого основания.
10
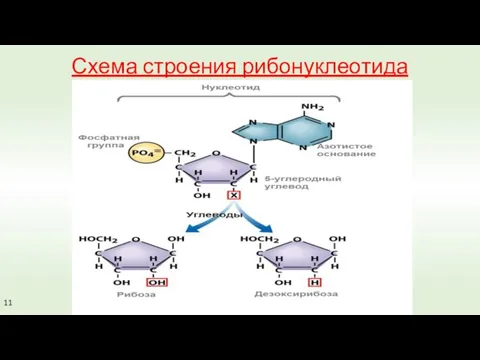
Схема строения рибонуклеотида
11
Схема строения рибонуклеотида
11

Биологическая роль нуклеотидов
Рибонуклеотиды пуринового или пиримидинового рядов (АМФ, ГМФ,УМФ и
Биологическая роль нуклеотидов
Рибонуклеотиды пуринового или пиримидинового рядов (АМФ, ГМФ,УМФ и

Гидролиз нуклеопротеинов
13
Гидролиз нуклеопротеинов
13
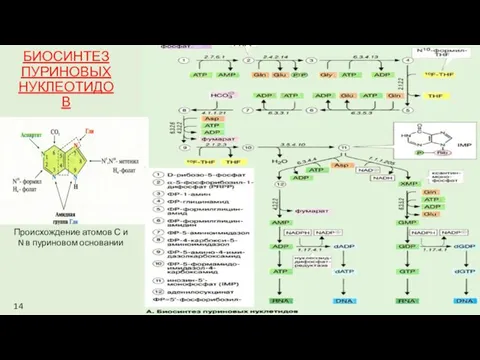
БИОСИНТЕЗ ПУРИНОВЫХ НУКЛЕОТИДОВ
Происхождение атомов С и N в пуриновом основании
14
БИОСИНТЕЗ ПУРИНОВЫХ НУКЛЕОТИДОВ
Происхождение атомов С и N в пуриновом основании
14
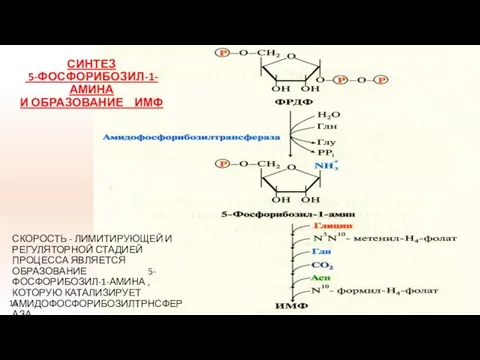
СИНТЕЗ
5-ФОСФОРИБОЗИЛ-1-АМИНА
И ОБРАЗОВАНИЕ ИМФ
СКОРОСТЬ - ЛИМИТИРУЮЩЕЙ И РЕГУЛЯТОРНОЙ СТАДИЕЙ ПРОЦЕССА
СИНТЕЗ
5-ФОСФОРИБОЗИЛ-1-АМИНА
И ОБРАЗОВАНИЕ ИМФ
СКОРОСТЬ - ЛИМИТИРУЮЩЕЙ И РЕГУЛЯТОРНОЙ СТАДИЕЙ ПРОЦЕССА
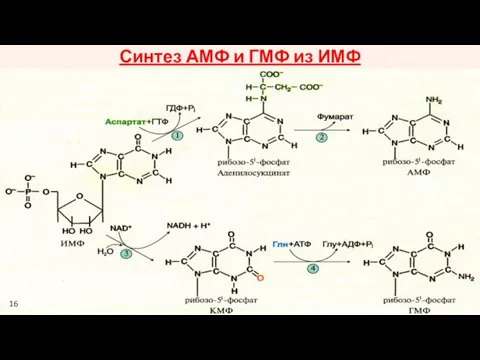
Синтез АМФ и ГМФ из ИМФ
16
Синтез АМФ и ГМФ из ИМФ
16

ЗАПАСНЫЕ ПУТИ СИНТЕЗА ПУРИНОВЫХ НУКЛЕОТИДОВ
Потребность в большом количестве нуклеотидов привела к
ЗАПАСНЫЕ ПУТИ СИНТЕЗА ПУРИНОВЫХ НУКЛЕОТИДОВ
Потребность в большом количестве нуклеотидов привела к
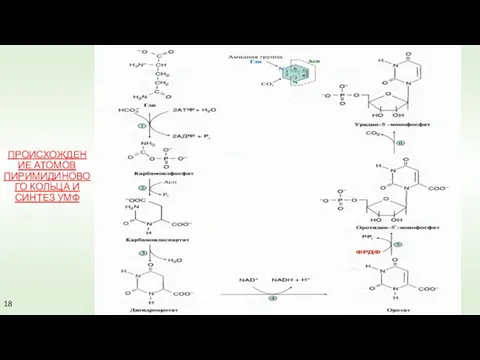
ПРОИСХОЖДЕНИЕ АТОМОВ ПИРИМИДИНОВОГО КОЛЬЦА И СИНТЕЗ УМФ
18
ПРОИСХОЖДЕНИЕ АТОМОВ ПИРИМИДИНОВОГО КОЛЬЦА И СИНТЕЗ УМФ
18

Регуляторным ферментом в синтезе пиримидиновых нуклеотидов является полифункциональный
КАД-фермент, который катализирует
Регуляторным ферментом в синтезе пиримидиновых нуклеотидов является полифункциональный
КАД-фермент, который катализирует

20
20
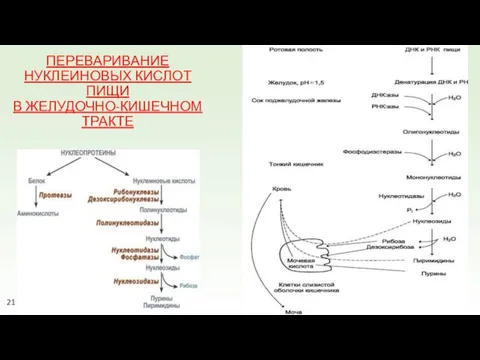
ПЕРЕВАРИВАНИЕ НУКЛЕИНОВЫХ КИСЛОТ ПИЩИ
В ЖЕЛУДОЧНО-КИШЕЧНОМ ТРАКТЕ
21
ПЕРЕВАРИВАНИЕ НУКЛЕИНОВЫХ КИСЛОТ ПИЩИ
В ЖЕЛУДОЧНО-КИШЕЧНОМ ТРАКТЕ
21
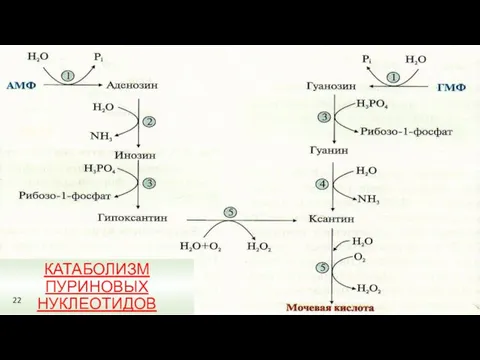
КАТАБОЛИЗМ ПУРИНОВЫХ НУКЛЕОТИДОВ
22
КАТАБОЛИЗМ ПУРИНОВЫХ НУКЛЕОТИДОВ
22
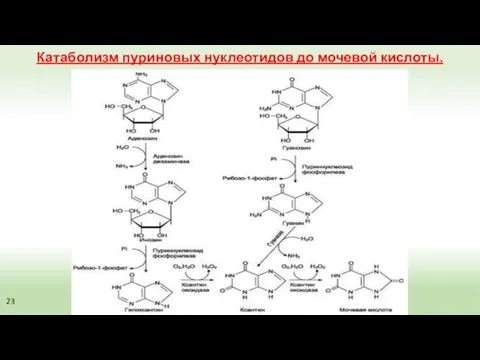
Катаболизм пуриновых нуклеотидов до мочевой кислоты.
23
Катаболизм пуриновых нуклеотидов до мочевой кислоты.
23

КАТАБОЛИЗМ ПУРИНОВЫХ ОСНОВАНИЙ
24
КАТАБОЛИЗМ ПУРИНОВЫХ ОСНОВАНИЙ
24

Мочевая кислота - основной продукт катаболизма пуриновых нуклеотидов у человека.
25
Мочевая кислота - основной продукт катаболизма пуриновых нуклеотидов у человека.
25
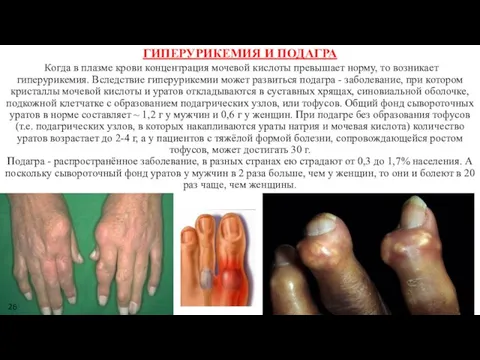
ГИПЕРУРИКЕМИЯ И ПОДАГРА
Когда в плазме крови концентрация мочевой кислоты превышает
ГИПЕРУРИКЕМИЯ И ПОДАГРА Когда в плазме крови концентрация мочевой кислоты превышает
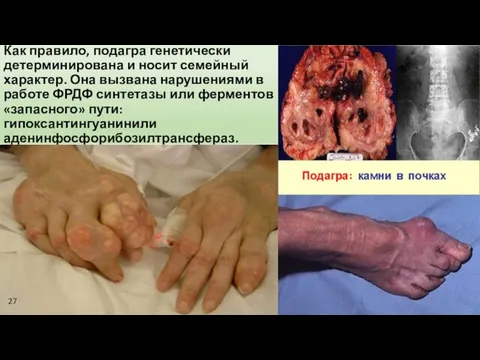
Как правило, подагра генетически детерминирована и носит семейный характер. Она вызвана
Как правило, подагра генетически детерминирована и носит семейный характер. Она вызвана

Гиперурикемическая (подагрическая) стимуляция умственной активности
Свою разгадку повышенная частота подагриков среди гениев
Гиперурикемическая (подагрическая) стимуляция умственной активности
Свою разгадку повышенная частота подагриков среди гениев

Первым подагриком, зарегистрированным в истории, был Иудейский царь, мудрый Аза.
Подагрой болели
Первым подагриком, зарегистрированным в истории, был Иудейский царь, мудрый Аза.
Подагрой болели
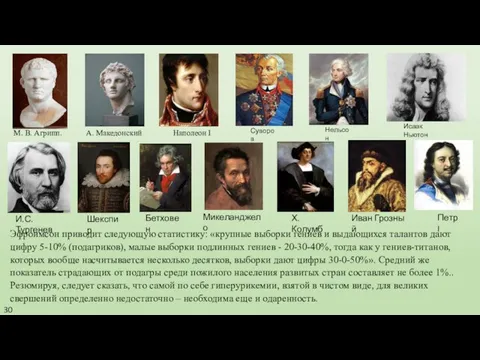
Эфроимсон приводит следующую статистику: «крупные выборки гениев и выдающихся талантов дают
Эфроимсон приводит следующую статистику: «крупные выборки гениев и выдающихся талантов дают
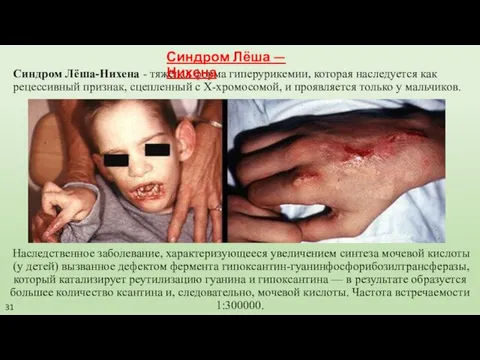
Наследственное заболевание, характеризующееся увеличением синтеза мочевой кислоты
(у детей) вызванное
Наследственное заболевание, характеризующееся увеличением синтеза мочевой кислоты (у детей) вызванное
 Wild animals
Wild animals Тип Плоские черви
Тип Плоские черви Структура, функции и биофизические свойства биологических мембран. Транспорт веществ в мембранах
Структура, функции и биофизические свойства биологических мембран. Транспорт веществ в мембранах Органы и системы органов
Органы и системы органов Биотехнология растений. Клеточная селекция
Биотехнология растений. Клеточная селекция Тетерев
Тетерев Немного про акул
Немного про акул  Птицы леса НСО
Птицы леса НСО Цитоплазма и органеллы
Цитоплазма и органеллы Общая характеристика процессов метаболизма
Общая характеристика процессов метаболизма Кровеносная система животных. Кровь
Кровеносная система животных. Кровь Физиологический механизм и условия формирования условных рефлексов
Физиологический механизм и условия формирования условных рефлексов Полимеразная цепная реакция (ПЦР) и методы, основанные на ПЦР
Полимеразная цепная реакция (ПЦР) и методы, основанные на ПЦР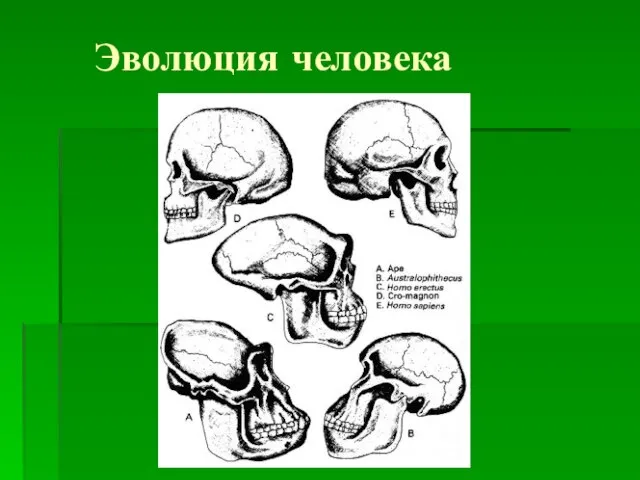 Эволюция человека
Эволюция человека Комодский варан
Комодский варан Переваривание углеводов. Метаболизм гликогена
Переваривание углеводов. Метаболизм гликогена Древние растения и животные Кропачева Марина Васильевна учитель начальных классов МАОУ «СОШ №12 с углубленным изучением немец
Древние растения и животные Кропачева Марина Васильевна учитель начальных классов МАОУ «СОШ №12 с углубленным изучением немец Как живут животные
Как живут животные Семейства Бобовые, Лилейные, Злаковые
Семейства Бобовые, Лилейные, Злаковые Экосистемы Земли
Экосистемы Земли  Класс Млекопитающие или Звери Отряд Ластоногие
Класс Млекопитающие или Звери Отряд Ластоногие Автор Лазарева О. В.
Автор Лазарева О. В.  Материк Антарктида
Материк Антарктида Что такое память?
Что такое память? Многообразие насекомых, их роль в природе, жизни человека и охрана
Многообразие насекомых, их роль в природе, жизни человека и охрана САНИТАРНАЯ ОХРАНА АТМОСФЕРНОГО ВОЗДУХА Доцент С.Н. Носков
САНИТАРНАЯ ОХРАНА АТМОСФЕРНОГО ВОЗДУХА Доцент С.Н. Носков  Немного о кошках
Немного о кошках Анатомо-морфологические особенности и основные физиологические функции организма
Анатомо-морфологические особенности и основные физиологические функции организма