- Основы медицинской экологии

Содержание
- 2. ВЕГЕТАТИВНАЯ (АВТОНОМНАЯ) НЕРВНАЯ СИСТЕМА
- 3. Еще в начале XIX века французский физиолог М. Биша разделил функции животного организма на животные (анимальные,
- 4. ОТЛИЧИЯ ВЕГЕТАТИВНОЙ И СОМАТИЧЕСКОЙ СИСТЕМЫ Вегетативная н.с. непроизвольная Иннервирует внутренние органы, железы внешней и внутренней секреции,
- 5. ОТЛИЧИЯ ВЕГЕТАТИВНОЙ И СОМАТИЧЕСКОЙ СИСТЕМЫ Волокна вегетативной нервной системы выходят из ЦНС только на определенных участках
- 6. Вегетативные нервные волокна имеют меньший диаметр, чем соматические. Волокна типа В покрыты тонкой миелиновой оболочкой, типа
- 7. В вегетативной нервной системе выделяют симпатический и парасимпатический отделы. Эти отделы имеют центральную и периферические части.
- 8. Многие внутренние органы наряду с симпатической и парасимпатической иннервациями имеют собственный местный нервный механизм регуляции функций,
- 9. ФУНКЦИИ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ 1. Управление деятельностью висцеральных органов: а) пусковая функция активация специфической для органа
- 10. а) усиление функциональных ответов на раздражители за счет использования энергетических ресурсов (эрготропная функция); б) коррекция и
- 12. Симпатический отдел вегетативной нервной системы Центры симпатической нервной системы представлены ядрами, расположенными в боковых рогах серого
- 14. Часть преганглионарных волокон, выходящих из ядер спинного мозга, проходит через вертебральные ганглии, не прерываясь, и переключаются
- 15. Окончания преганглионарных волокон вырабатывают ацетилхолин, постганглионарных — в основном норадреналин. Исключение составляют постганглионарные волокна, иннервирующие потовые
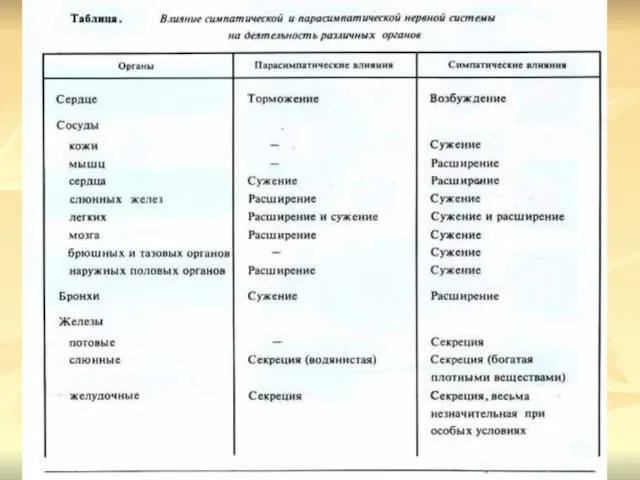
- 16. ДЕЙСТВИЕ СИМПАТИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ Симпатическая нервная система иннервирует все органы и ткани организма, в том числе
- 17. Симпатическая нервная система не только регулирует работу внутренних органов, но и оказывает влияние на обменные процессы,
- 18. Путем раздражения двигательного нерва вызывали сокращения мышцы и доводили ее до степени утомления. Раздражение симпатического нерва
- 19. Симпатическая нервная система отвечает на любой стресс. Ее возбуждение приводит к увеличению активности мозгового вещества надпочечников
- 20. ПАРАСИМПАТИЧЕСКИЙ ОТДЕЛ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ Центрами парасимпатического отдела автономной нервной системы являются ядра, находящиеся в среднем
- 21. Ганглии парасимпатической нервной системы располагаются вблизи иннервируемых органов или внутри них. Поэтому преганглионарные волокна парасимпатического отдела
- 23. Постганглионарные парасимпатические волокна иннервируют глазные мышцы, слезные и слюнные железы, мускулатуру и железы пищеварительного тракта, трахею,
- 24. Действие парасимпатической нервной системы направлено на восстановление и поддержание постоянства состава внутренней среды организма, нарушенного в
- 25. ВНУТРИОРГАННЫЙ ОТДЕЛ (ЭНТЕРАЛЬНЫЙ, МЕТАСИМПАТИЧЕСКИЙ) К этому отделу относятся интрамуральные системы всех полых висцеральных органов, обладающих собственной
- 26. Этот отдел отличается более строгой автономностью, т.е. независимостью от ЦНС, так как не имеет прямых синаптических
- 27. В сфере управления этого отдела находятся гладкие мышцы, всасывающий и секретирующий эпителий, локальный кровоток, местные эндокринные
- 28. Синаптическая передача В вегетативной нервной системе существует три вида синаптической передачи: химическая, электрическая и смешанная. Основным
- 29. Кроме постсинаптических рецепторов выделяют пре- и внесинаптические рецепторы. Пресинаптические рецепторы непосредственно на функции органов и тканей
- 30. Локализация и количество любых рецепторов на мембране клетки детерминировано генетическим аппаратом. Однако это количество может меняться
- 31. Медиаторы и рецепторы вегетативной нервной системы Ацетилхолин высвобождается в окончаниях холинергических парасимпатических и симпатических волокон. Инактивация
- 32. Н-ХОЛИНОРЕЦЕПТОРЫ в периферических отделах вегетативной нервной системы расположены в ганглионарных синапсах симпатического и парасимпатического отделов, в
- 33. М-ХОЛИНОРЕЦЕПТОРЫ подразделяются на несколько типов: М1-, М2- и М3-холинорецепторы. Все они блокируются атропином. М1-холинорецепторы находятся на
- 34. М3-холинорецепторы локализованы в основном в гладких мышцах некоторых внутренних органов и экзокринных железах. Взаимодействие ацетилхолина с
- 35. Возбуждение М3-холинорецепторов экзокринных желез вызывает слезотечение, усиление потоотделения, выделение обильной бедной белком слюны, выделение желудочного сока.
- 36. Норадреналин Обеспечивает химическую передачу нервного импульса в норадренергических синапсах. Норадреналин относится к катехоламинам. Он синтезируется из
- 37. Действие норадреналина на клетку опосредуется адренорецепторами. Адренорецепторы находятся в различных тканях организма и воспринимают действие норадреналина
- 38. Возбуждение α1-адренорецепторов приводит к: сужению радиальной мышцы глаза и расширению зрачка (мидриаз), сужению соответствующих сосудов и
- 39. Среди α2-адренорецепторов выделяют пре-, пост- и внесинаптические. Возбуждение пресинаптических α2-адренорецепторов по механизму отрицательной обратной связи уменьшает
- 40. Β1-адренорецепторы (постсинаптические) выявлены в основном в проводящей системе сердца и гладкой мышце кишечника. Их возбуждение приводит
- 41. Стимуляция пресинаптических β2-адренорецепторов по механизму положительной обратной связи вызывает выделение норадреналина при его недостатке в синаптической
- 42. Возбуждение β2-адренорецепторов вызывает : расширение соответствующих сосудов и понижение АД, расслабление бронхов и матки, усиление в
- 43. ДОФАМИН осуществляет химическую передачу нервных импульсов не только в дофаминергических синапсах ЦНС, но и во вставочных
- 44. Д-рецепторы выявлены на гладкомышечных клетках кишечника, сосудов почек, аорты, паращиторидных железах, канальцах почек. Возбуждение этих рецепторов
- 45. АТФ может играть роль не только макроергического соединения, но и медиатора. Местом его локализации является пресинаптические
- 46. Одним из медиаторов внутриорганного отдела вегетативной нервной системы является серотонин, или 5-окситриптамин, который выполняет также медиаторную
- 47. Периферические S1-рецепторы (или 5-НТ1) в основном обнаружены в гладких мышцах желудочно-кишечного тракта, сосудах скелетных мышц и
- 48. Роль медиатора в вегетативной нервной системе может играть гистамин. Наибольшее количество его находится в постганглионарных симпатических
- 49. Возбуждение Н1-рецепторов сопровождается: спазмом бронхов повышением тонуса и перистальтики кишечника сужением крупных сосудов, но расширением артериол,
- 50. Возбуждение Н2-рецепторов приводит к: повышению секреции кислоты в желудке и секреции бронхиальных желез уменьшению высвобождения гистамина
- 51. Функцию медиаторов синаптической передачи во внутриорганном отделе вегетативной нервной системы выполняют и некоторые аминокислоты, регуляторные нейропептиды,
- 52. Вегетативные (автономные) рефлексы Различают висцеро-висцеральные, висцеросоматические, висцеросенсорные рефлексы. Классическим примером висцеро-висцерального рефлекса является рефлекс Гольца, показывающий,
- 53. К висцеросоматическим рефлексам относятся : торможение общей двигательной активности организма при раздражении хемо- и механорецепторов каротидной
- 54. При висцеросенсорных рефлексах в ответ на раздражение вегетативных чувствительных волокон возникают не только реакции во внутренних
- 55. Центры регуляции вегетативных функций Центры регуляции вегетативных функций разделяются на: спинальные, стволовые (бульбарные, мезэнцефалические), гипоталамические, мозжечковые,
- 56. Спинальные центры. На уровне спинного мозга происходит: регуляция просвета зрачка, величины глазной щели, сосудистого тонуса, потоотделения.
- 57. Стволовые центры. Эти центры находятся в продолговатом мозге, мосту, среднем мозге. За счет ядер блуждающих нервов
- 58. В продолговатом мозге находятся центры, с помощью которых осуществляются такие сложные рефлексы, как сосание, жевание, глотание,
- 59. Гипоталамические центры. Гипоталамус является главным подкорковым центром интеграции висцеральных процессов, что обеспечивается вегетативными, соматическими и эндокринными
- 60. Стимуляция передних ядер гипоталамуса приводит к эффектам, подобным раздражению парасимпатической нервной системы: сужение зрачков и глазных
- 61. Средняя группа ядер гипоталамуса обеспечивает регуляцию метаболизма и водного баланса. Вентромедиальные ядра отвечают за насыщение, латеральные
- 62. Центры лимбической системы. Эти центры отвечают за формирование вегетативного компонента эмоциональных реакций, пищевое, сексуальное, оборонительное поведение,
- 64. Скачать презентацию

ВЕГЕТАТИВНАЯ (АВТОНОМНАЯ) НЕРВНАЯ СИСТЕМА
ВЕГЕТАТИВНАЯ (АВТОНОМНАЯ) НЕРВНАЯ СИСТЕМА

Еще в начале XIX века французский физиолог М. Биша разделил
Еще в начале XIX века французский физиолог М. Биша разделил

ОТЛИЧИЯ ВЕГЕТАТИВНОЙ И СОМАТИЧЕСКОЙ СИСТЕМЫ
Вегетативная н.с. непроизвольная
Иннервирует внутренние органы, железы внешней
ОТЛИЧИЯ ВЕГЕТАТИВНОЙ И СОМАТИЧЕСКОЙ СИСТЕМЫ
Вегетативная н.с. непроизвольная
Иннервирует внутренние органы, железы внешней

ОТЛИЧИЯ ВЕГЕТАТИВНОЙ И СОМАТИЧЕСКОЙ СИСТЕМЫ
Волокна вегетативной нервной системы выходят из ЦНС
ОТЛИЧИЯ ВЕГЕТАТИВНОЙ И СОМАТИЧЕСКОЙ СИСТЕМЫ
Волокна вегетативной нервной системы выходят из ЦНС

Вегетативные нервные волокна имеют меньший диаметр, чем соматические. Волокна типа В
Вегетативные нервные волокна имеют меньший диаметр, чем соматические. Волокна типа В

В вегетативной нервной системе выделяют симпатический и парасимпатический отделы. Эти отделы
В вегетативной нервной системе выделяют симпатический и парасимпатический отделы. Эти отделы

Многие внутренние органы наряду с симпатической и парасимпатической иннервациями имеют собственный
Многие внутренние органы наряду с симпатической и парасимпатической иннервациями имеют собственный

ФУНКЦИИ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
1. Управление деятельностью висцеральных органов:
а) пусковая функция
ФУНКЦИИ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
1. Управление деятельностью висцеральных органов: а) пусковая функция

а) усиление функциональных ответов на раздражители за счет использования энергетических
а) усиление функциональных ответов на раздражители за счет использования энергетических

Симпатический отдел вегетативной нервной системы
Центры симпатической нервной системы представлены ядрами, расположенными
Симпатический отдел вегетативной нервной системы
Центры симпатической нервной системы представлены ядрами, расположенными
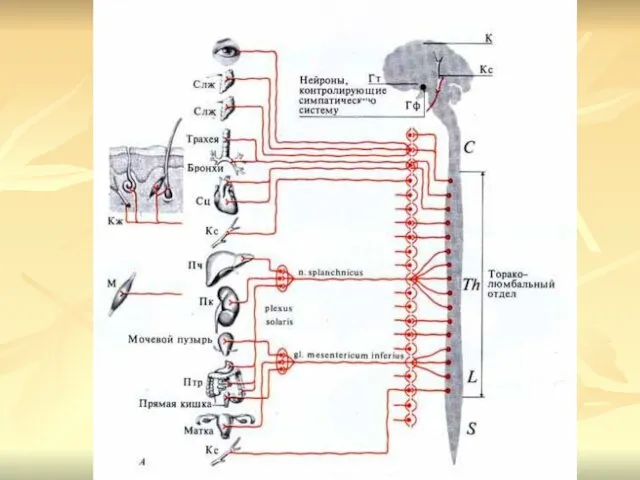

Часть преганглионарных волокон, выходящих из ядер спинного мозга, проходит через вертебральные
Часть преганглионарных волокон, выходящих из ядер спинного мозга, проходит через вертебральные

Окончания преганглионарных волокон вырабатывают ацетилхолин, постганглионарных — в основном норадреналин. Исключение
Окончания преганглионарных волокон вырабатывают ацетилхолин, постганглионарных — в основном норадреналин. Исключение

ДЕЙСТВИЕ СИМПАТИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ Симпатическая нервная система иннервирует все органы
ДЕЙСТВИЕ СИМПАТИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ Симпатическая нервная система иннервирует все органы

Симпатическая нервная система не только регулирует работу внутренних органов, но
Симпатическая нервная система не только регулирует работу внутренних органов, но

Путем раздражения двигательного нерва вызывали сокращения мышцы и доводили ее до
Путем раздражения двигательного нерва вызывали сокращения мышцы и доводили ее до

Симпатическая нервная система отвечает на любой стресс. Ее возбуждение приводит к
Симпатическая нервная система отвечает на любой стресс. Ее возбуждение приводит к

ПАРАСИМПАТИЧЕСКИЙ ОТДЕЛ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
Центрами парасимпатического отдела автономной нервной системы
ПАРАСИМПАТИЧЕСКИЙ ОТДЕЛ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
Центрами парасимпатического отдела автономной нервной системы

Ганглии парасимпатической нервной системы располагаются вблизи иннервируемых органов или внутри
Ганглии парасимпатической нервной системы располагаются вблизи иннервируемых органов или внутри
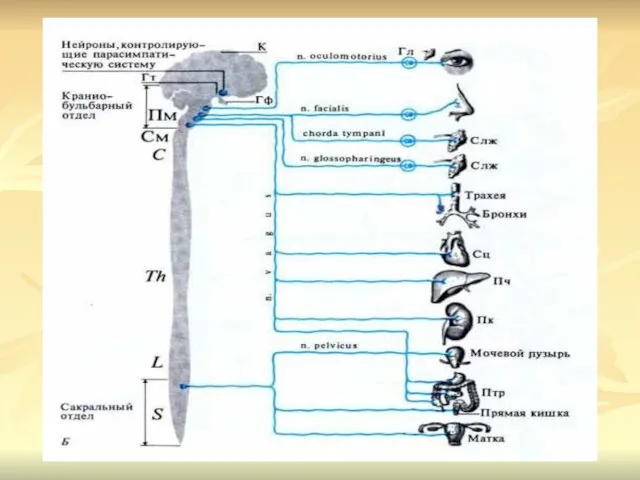

Постганглионарные парасимпатические волокна иннервируют глазные мышцы, слезные и слюнные железы, мускулатуру
Постганглионарные парасимпатические волокна иннервируют глазные мышцы, слезные и слюнные железы, мускулатуру

Действие парасимпатической нервной системы направлено на восстановление и поддержание постоянства
Действие парасимпатической нервной системы направлено на восстановление и поддержание постоянства

ВНУТРИОРГАННЫЙ ОТДЕЛ (ЭНТЕРАЛЬНЫЙ, МЕТАСИМПАТИЧЕСКИЙ)
К этому отделу относятся интрамуральные системы всех полых
ВНУТРИОРГАННЫЙ ОТДЕЛ (ЭНТЕРАЛЬНЫЙ, МЕТАСИМПАТИЧЕСКИЙ)
К этому отделу относятся интрамуральные системы всех полых

Этот отдел отличается более строгой автономностью, т.е. независимостью от ЦНС, так
Этот отдел отличается более строгой автономностью, т.е. независимостью от ЦНС, так

В сфере управления этого отдела находятся гладкие мышцы, всасывающий и секретирующий
В сфере управления этого отдела находятся гладкие мышцы, всасывающий и секретирующий

Синаптическая передача
В вегетативной нервной системе существует три вида синаптической передачи: химическая,
Синаптическая передача
В вегетативной нервной системе существует три вида синаптической передачи: химическая,

Кроме постсинаптических рецепторов выделяют пре- и внесинаптические рецепторы. Пресинаптические рецепторы непосредственно
Кроме постсинаптических рецепторов выделяют пре- и внесинаптические рецепторы. Пресинаптические рецепторы непосредственно

Локализация и количество любых рецепторов на мембране клетки детерминировано генетическим аппаратом.
Локализация и количество любых рецепторов на мембране клетки детерминировано генетическим аппаратом.

Медиаторы и рецепторы вегетативной нервной системы
Ацетилхолин высвобождается в окончаниях
Медиаторы и рецепторы вегетативной нервной системы
Ацетилхолин высвобождается в окончаниях

Н-ХОЛИНОРЕЦЕПТОРЫ в периферических отделах вегетативной нервной системы расположены в ганглионарных
Н-ХОЛИНОРЕЦЕПТОРЫ в периферических отделах вегетативной нервной системы расположены в ганглионарных

М-ХОЛИНОРЕЦЕПТОРЫ подразделяются на несколько типов: М1-, М2- и М3-холинорецепторы. Все
М-ХОЛИНОРЕЦЕПТОРЫ подразделяются на несколько типов: М1-, М2- и М3-холинорецепторы. Все

М3-холинорецепторы локализованы в основном в гладких мышцах некоторых внутренних органов
М3-холинорецепторы локализованы в основном в гладких мышцах некоторых внутренних органов

Возбуждение М3-холинорецепторов экзокринных желез вызывает слезотечение, усиление потоотделения, выделение обильной
Возбуждение М3-холинорецепторов экзокринных желез вызывает слезотечение, усиление потоотделения, выделение обильной

Норадреналин
Обеспечивает химическую передачу нервного импульса в норадренергических синапсах. Норадреналин относится
Норадреналин
Обеспечивает химическую передачу нервного импульса в норадренергических синапсах. Норадреналин относится

Действие норадреналина на клетку опосредуется адренорецепторами. Адренорецепторы находятся в различных
Действие норадреналина на клетку опосредуется адренорецепторами. Адренорецепторы находятся в различных

Возбуждение α1-адренорецепторов приводит к:
сужению радиальной мышцы глаза и расширению
Возбуждение α1-адренорецепторов приводит к:
сужению радиальной мышцы глаза и расширению

Среди α2-адренорецепторов выделяют пре-, пост- и внесинаптические. Возбуждение пресинаптических α2-адренорецепторов
Среди α2-адренорецепторов выделяют пре-, пост- и внесинаптические. Возбуждение пресинаптических α2-адренорецепторов

Β1-адренорецепторы (постсинаптические) выявлены в основном в проводящей системе сердца и
Β1-адренорецепторы (постсинаптические) выявлены в основном в проводящей системе сердца и

Стимуляция пресинаптических β2-адренорецепторов по механизму положительной обратной связи вызывает выделение
Стимуляция пресинаптических β2-адренорецепторов по механизму положительной обратной связи вызывает выделение

Возбуждение β2-адренорецепторов вызывает :
расширение соответствующих сосудов и понижение АД,
расслабление
Возбуждение β2-адренорецепторов вызывает :
расширение соответствующих сосудов и понижение АД,
расслабление

ДОФАМИН осуществляет химическую передачу нервных импульсов не только в дофаминергических
ДОФАМИН осуществляет химическую передачу нервных импульсов не только в дофаминергических

Д-рецепторы выявлены на гладкомышечных клетках кишечника, сосудов почек, аорты, паращиторидных
Д-рецепторы выявлены на гладкомышечных клетках кишечника, сосудов почек, аорты, паращиторидных

АТФ может играть роль не только макроергического соединения, но и
АТФ может играть роль не только макроергического соединения, но и

Одним из медиаторов внутриорганного отдела вегетативной нервной системы является серотонин,
Одним из медиаторов внутриорганного отдела вегетативной нервной системы является серотонин,

Периферические S1-рецепторы (или 5-НТ1) в основном обнаружены в гладких мышцах
Периферические S1-рецепторы (или 5-НТ1) в основном обнаружены в гладких мышцах

Роль медиатора в вегетативной нервной системе может играть гистамин. Наибольшее количество
Роль медиатора в вегетативной нервной системе может играть гистамин. Наибольшее количество

Возбуждение Н1-рецепторов сопровождается:
спазмом бронхов
повышением тонуса и перистальтики кишечника
Возбуждение Н1-рецепторов сопровождается:
спазмом бронхов
повышением тонуса и перистальтики кишечника

Возбуждение Н2-рецепторов приводит к:
повышению секреции кислоты в желудке и секреции
Возбуждение Н2-рецепторов приводит к:
повышению секреции кислоты в желудке и секреции

Функцию медиаторов синаптической передачи во внутриорганном отделе вегетативной нервной системы
Функцию медиаторов синаптической передачи во внутриорганном отделе вегетативной нервной системы

Вегетативные (автономные) рефлексы
Различают висцеро-висцеральные, висцеросоматические, висцеросенсорные рефлексы. Классическим примером висцеро-висцерального
Вегетативные (автономные) рефлексы
Различают висцеро-висцеральные, висцеросоматические, висцеросенсорные рефлексы. Классическим примером висцеро-висцерального

К висцеросоматическим рефлексам относятся :
торможение общей двигательной активности организма при
К висцеросоматическим рефлексам относятся :
торможение общей двигательной активности организма при

При висцеросенсорных рефлексах в ответ на раздражение вегетативных чувствительных волокон
При висцеросенсорных рефлексах в ответ на раздражение вегетативных чувствительных волокон

Центры регуляции вегетативных функций
Центры регуляции вегетативных функций разделяются на:
спинальные,
стволовые
Центры регуляции вегетативных функций
Центры регуляции вегетативных функций разделяются на:
спинальные,
стволовые

Спинальные центры. На уровне спинного мозга происходит:
регуляция просвета зрачка,
Спинальные центры. На уровне спинного мозга происходит:
регуляция просвета зрачка,

Стволовые центры. Эти центры находятся в продолговатом мозге, мосту, среднем
Стволовые центры. Эти центры находятся в продолговатом мозге, мосту, среднем

В продолговатом мозге находятся центры, с помощью которых осуществляются такие сложные
В продолговатом мозге находятся центры, с помощью которых осуществляются такие сложные

Гипоталамические центры. Гипоталамус является главным подкорковым центром интеграции висцеральных процессов, что
Гипоталамические центры. Гипоталамус является главным подкорковым центром интеграции висцеральных процессов, что

Стимуляция передних ядер гипоталамуса приводит к эффектам, подобным раздражению парасимпатической нервной
Стимуляция передних ядер гипоталамуса приводит к эффектам, подобным раздражению парасимпатической нервной

Средняя группа ядер гипоталамуса обеспечивает регуляцию метаболизма и водного баланса.
Средняя группа ядер гипоталамуса обеспечивает регуляцию метаболизма и водного баланса.

Центры лимбической системы. Эти центры отвечают за формирование вегетативного компонента эмоциональных
Центры лимбической системы. Эти центры отвечают за формирование вегетативного компонента эмоциональных
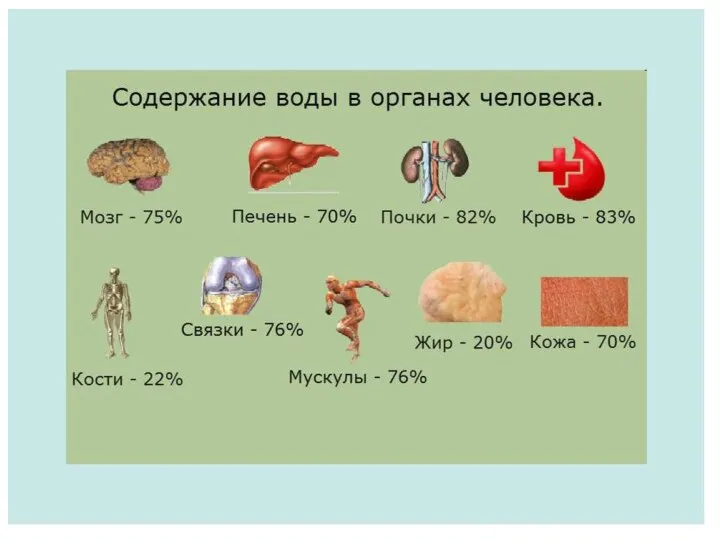 Определение содержания воды в организме
Определение содержания воды в организме Системы органов многоклеточного организма. 11класс
Системы органов многоклеточного организма. 11класс Презентация Таймырский заповедник
Презентация Таймырский заповедник  Учение о клетке Курс Общая биология 9 класс 2009
Учение о клетке Курс Общая биология 9 класс 2009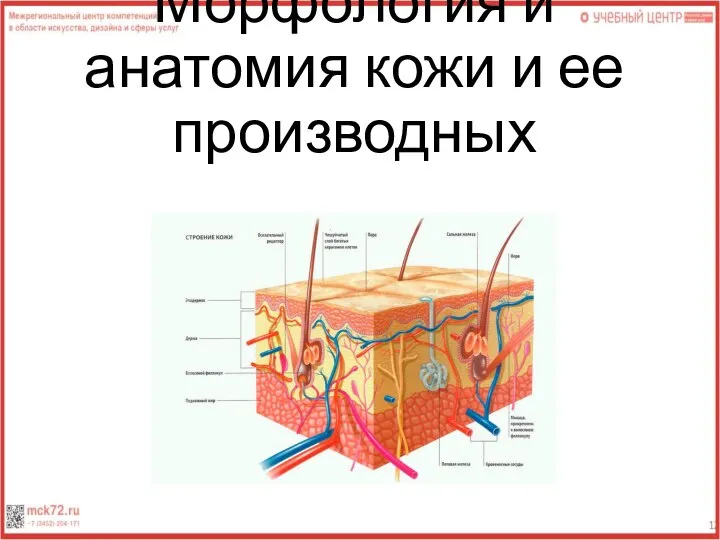 Морфология и анатомия кожи и ее производных
Морфология и анатомия кожи и ее производных Отряд Перепончатокрылые
Отряд Перепончатокрылые Клетка. Учение о тканях. Виды тканей
Клетка. Учение о тканях. Виды тканей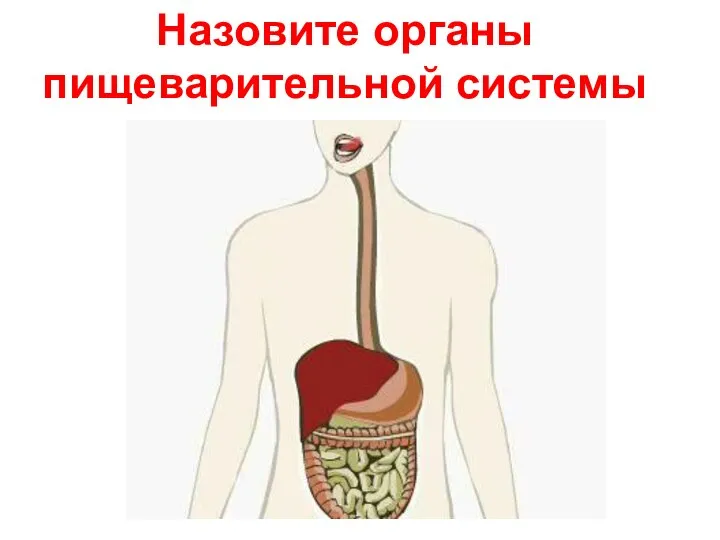 Пищеварение в желудке и в кишечнике
Пищеварение в желудке и в кишечнике Скелет человека Осевой скелет и скелет конечностей. Автор: Орлова Наталья Алексеевна Учитель биологии и химии М
Скелет человека Осевой скелет и скелет конечностей. Автор: Орлова Наталья Алексеевна Учитель биологии и химии М ТИП Плоские черви Класс Ресничные черви Класс Сосальщики (Трематоды) Класс Ленточные черви (Цестоды) Презентация учителя био
ТИП Плоские черви Класс Ресничные черви Класс Сосальщики (Трематоды) Класс Ленточные черви (Цестоды) Презентация учителя био Pathologic Protozoa
Pathologic Protozoa Эфирные масла и их биологическое действие
Эфирные масла и их биологическое действие Степная Пищуха из Красной книги Челябинской области
Степная Пищуха из Красной книги Челябинской области Биосферные резерваты
Биосферные резерваты 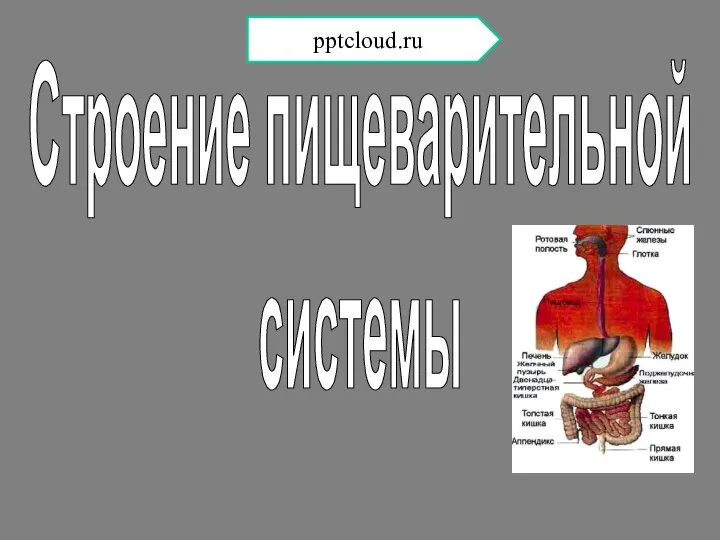 Строение пищеварительной системы
Строение пищеварительной системы Дерево какао
Дерево какао Видообразование. Микроэволюция и макроэволюция
Видообразование. Микроэволюция и макроэволюция Кольчатые черви
Кольчатые черви  «Строение цветка и его особенности».
«Строение цветка и его особенности». Презентация на тему "Воздействие человека и его деятельности на животных" - скачать бесплатно презентации по Биологии
Презентация на тему "Воздействие человека и его деятельности на животных" - скачать бесплатно презентации по Биологии Презентация на тему "Красная книга растений (5 класс)" - скачать бесплатно презентации по Биологии
Презентация на тему "Красная книга растений (5 класс)" - скачать бесплатно презентации по Биологии Строение клетки. 5 класс
Строение клетки. 5 класс Размножение и развитие пресмыкающихся
Размножение и развитие пресмыкающихся Алгоритм исследования Овощи
Алгоритм исследования Овощи Ми қыртысының қалыптасуы. Анализаторлар туралы түсінік
Ми қыртысының қалыптасуы. Анализаторлар туралы түсінік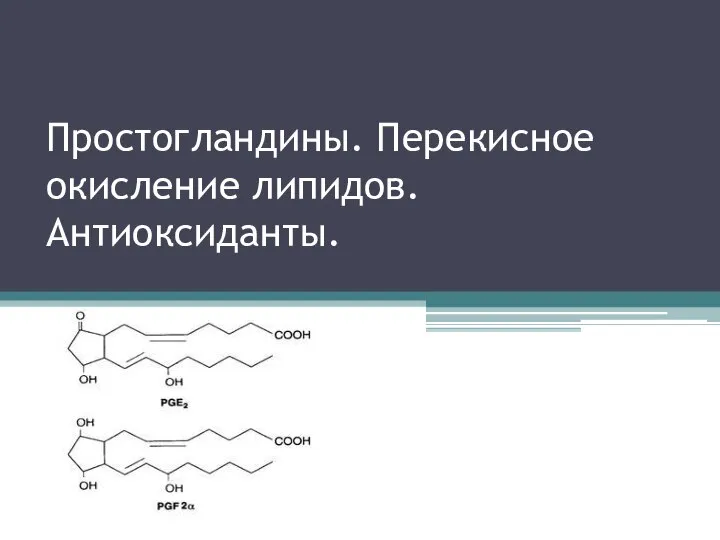 Простогландины. Перекисное окисление липидов. Антиоксиданты
Простогландины. Перекисное окисление липидов. Антиоксиданты Запліднення. Періоди онтогенезу багатоклітинних організмів: ембріогенез і постембріональний розвиток
Запліднення. Періоди онтогенезу багатоклітинних організмів: ембріогенез і постембріональний розвиток Генетика. Основные этапы развития генетики
Генетика. Основные этапы развития генетики