- Развитие нервной системы живых организмов

Содержание
- 2. Происхождение нервных клеток -Evolution of nervous systems Нейронная теория - Neuron doctrine Гены раннего нейрогенеза –neural
- 3. Существование живых организмов зависит от умения общаться с внешним миром. Эукариоты. Многоклеточные. (540-620 млн лет.) У
- 4. Появление нервных клеток в организме связано непосредственно с реактивностью на внешние раздражения, т.е. с функцией, которая
- 5. Теории происхождения и эволюции нервных клеток 1. Теория Н. Клейненберга (1872)- нейроны возникли от единого нервно-мышечного
- 6. Молекулярно-генетические исследования показывают, что в возникновении нервной системы принимают участие транскрипционные регуляторы семейства Sox (SoxB), сигнальные
- 7. Sponges (губки) Есть сократительные клетки, но нет нервных. Cnidarian (гидра) – появляются нервные клетки, которые не
- 8. Цефализация и формирование ганглиев Плоские черви (головной конец, рот, анус). Планария - билатеральная НС. Между нейронами
- 9. Развитие и усложнение сегментарной нервной системы наблюдается у высших беспозвоночных животных - насекомых. Мозговой отдел воспринимает
- 10. НС позвоночных в отличие от беспозвоночных -занимает спинное положение, развивается из спинной эктодермы зародыша, представлена трубкой.
- 11. Parallel evolution and diversity of nervous systems Brain Behav Evol 2009;74:177–190 (гребневики) (губки) (кораллы, медузы…) (вторичноротые)
- 12. Два сценария эволюции нервных клеток: полифилия- независимое происхождение и монофилия- от единого предка Moroz L.L, Kohn
- 13. Nature. ; 2012. 483(7389): 289–294 Генетическая программа гомологичная трем сигнальным центрам позвоночных (the anterior neural ridge
- 14. Геном человека содержит примерно 30000 генов и около 50% из них экспрессируются в ткани мозга. В
- 15. Neurogenetics (2006) 7: 67–80 Gene expression analyses reveal molecular relationships among 20 regions of the human
- 16. MBP myelin basic protein glial high affinity glutamate transporter GRM3 glutamate receptor, metabotropic 3 (рецептор возбуждающего
- 17. анализаторную (анализ поступающей по сенсорным системам информации), интегративную (объединение, связь сигналов от различных сенсорных и моторных
- 18. Как устроена нервная система? Теория сети Герлах, Гельд, Мейнерт и Гольджи Нервная ткань - своеобразный синцитий,
- 19. S. Ramon y Cajal 1852-1934 Создатель «Нейронной теории» 1. Каждый нейрон является клеточной единицей самостоятельной в
- 21. В основе развития нервной системы лежат процессы: Индукция эктодермы Детерминация и позиционная информация Пролиферация клеток Миграция
- 22. Развитие нервной системы позвоночных I. Мезодермальная индукция – Vg1 (TGF-B), activin, BMP2,4, FGF, WNT II. Нейральная
- 23. Мозг, базальная пластинка, нотохорд Передняя эндодерма - граница c ЦНС Сигнальные центры у Xenopus BCNE-blastula chordin,
- 24. Neural Induction in Xenopus PLoS Biology May 2004 | Volume 2 | Issue 5 | Page
- 25. Нервная индукция (т.н. по умолчанию) - блокада BMP4 в части клеток эктодермы приводит к их нейрализации
- 26. У млекопитающих результатом взаимодействия BMP4 и chordin является активация пронейральных basicHLH транскрипционных факторов- MASH-1,2 +FGF передняя
- 27. Механизмы нейральной индукции Smad(s) - внутриклеточный передатчик сигнала BMPs
- 28. Факторы аксиальной мезодермы (организатора) CBP / p300
- 29. Нокаут генов noggin (В) и noggin+chordin (С) у мышей приводит к разным дефектам в развитии головы.
- 30. Детерминация и позиционная информация Определение судьбы нейральных клеток начинается со стадии бластулы и далее активируются SoxB
- 31. Модели развития нервной системы: Мозаичная модель – детерминирована судьба отдельной клетки (беспозвоночные). Регуляционная модель – случайное
- 32. Мозаичная модель – детерминирована судьба отдельной клетки (беспозвоночные).
- 33. Аm Al Al Pm Pm Pl Pl Клональная модель развития мозга - М. Джекобсон, Г. Хирозе
- 34. Int. J. Dev. BioI. 411:735-743 (1996) Гены ранней паттернизации передней части нервной пластинки Dll, Wnt, POU,
- 35. Бластула Гаструла Нейрула Детерминация Вероятно, процесс детерминации начинается в бластуле, а закрепляется при появление осевых структур
- 36. Они имеют сходную пространственную организацию и сходный порядок вдоль хромосом Нох гены консервативны в эволюционном ряду.
- 37. Дорзо-вентральный паттерн в нервной трубке устанавливают BMPs и WNT со стороны эпидермиса и Shh (Sonic hedgehog),
- 38. NIH PA 2011 Градиенты многих морфогенов и транскрипционных факторов координируют структурную организацию мозга FGF + Уровень
- 39. Распределение транскрипционных факторов и сигнальных молекул в развивающейся нервной трубке меняется во времени.
- 40. Важную роль в развитии и формировании структур переднего мозга играют регуляторные гены кодирующие транскрипционные факторы Pax6
- 41. Pax6 – Paired box gene 6 играет важную роль в развитии НС, «мастер ген» для развития
- 42. Pax6 в развитии разных структур мозга
- 43. Empty-spiracles family gene - Emx2 экспрессия появляется в вентрикулярной герминальной зоне, ранний маркер неокортекса (Е8.5), играет
- 44. Nature.Rev. 434 | JUNE 2007 | VOLUME 8 У мышей нокаутных по Emx2/Pax6 не формируется неокортекс
- 45. Транскрипционные факторы Emx2 и Pax6 определяют специфичность представительства сенсорных модальностей по коре мозга. Дикий тип Градиенты
- 46. Orthodenticle family genes - Otx2 ген в развитии переднего мозга и передней части головы. Делеция гена
- 47. На эктодерме дрозофилы установлено, что Notch/Delta сигнальная система определяет дифференцировку в нейробласты. Судьба клетки зависит от
- 48. Numb ингибитор Notch. Ассиметричное распределение Numb определяет судьбу клетки при делении стволовых нейроэпителиальных клеток и нейробластов
- 49. Дифференцировка нейрона регулируется множеством факторов
- 50. Дифференцировка нервных клеток в развивающейся нервной системе происходит в разных структурах в разное время. Это явление
- 51. Neuron. 2011 October 6; 72(1): 22–40. Дискретные транскрипционные факторы в пространстве и времени регулируют полярность нейрона
- 52. Путями для миграции конуса роста могут быть границы компартментов Аксоны пионеры (pioneer axon) прокладывают пути на
- 53. Навигация аксона- пионера с помощью нейронов ориентиров Нейроны Ti 1 лапки кузнечика посылают аксоны в ЦНС
- 54. На развивающейся личинке дрозофилы установлена роль САМ- фасциклина II в процессе фасцикуляции аксонов (Grenningloh et al.
- 55. Соответствующие по времени воздействия комплексов специфических молекулярных факторов, являются критичными для правильного развития структур мозга и
- 56. У млекопитающих нервные клетки формируются из двух источников: 1.Нервной трубки 2. Ганглиозной пластинки (нервного гребня)
- 57. Из нервной трубки формируется головной и спинной мозг.
- 58. Судьба клеток нервного гребня. Из нервного гребня формируются клетки периферической нервной системы, вегетативные ганглии, клетки мягкой
- 59. Время выхода в дифференцировку разных типов клеток нервной системы
- 60. Развитие коры мозга. Факторы bHLH контролируют пролиферацию и дифференцировку нервных клеток (NeuroD, Ngns, Mash, Olig и
- 61. Радиальная миграция нейробластов по отросткам клеток радиальной глии. Sidman and Rakic (1973)
- 62. Nature.Rev. 434 | JUNE 2007 | VOLUME 8 Генерация нейронов в неокортексе. Транскрипционные факторы определяющие разные
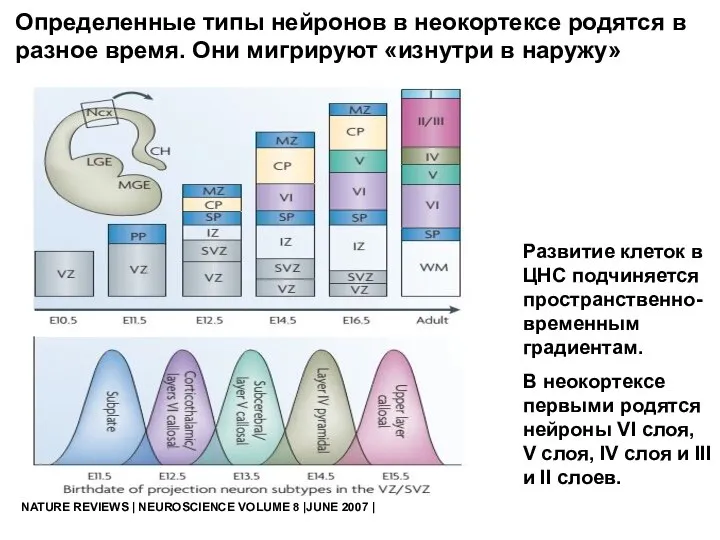
- 63. NATURE REVIEWS | NEUROSCIENCE VOLUME 8 |JUNE 2007 | Развитие клеток в ЦНС подчиняется пространственно-временным градиентам.
- 64. Белок Reelin регулирует клеточные взаимодействия, миграцию клеток и организацию кортикальной пластинки. (Reeler mice). Reelin- гликопротеин, регулирует
- 65. Мутация по гену reeler приводит к дезорганизации неокортекса
- 66. Конус роста аксона Конус роста аксона имеет двигательный и сенсорный аппараты. По окончании движения он образует
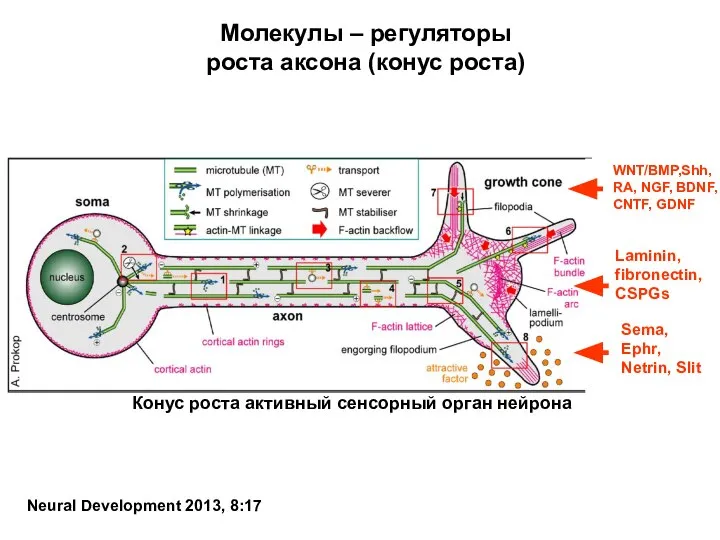
- 67. Молекулы – регуляторы роста аксона (конус роста) Neural Development 2013, 8:17
- 68. Навигация аксонов осуществляется за счет отталкивания и аттракции
- 69. Хемотрофические молекулы навигаторы являются членами консервативных семейств лиганд/рецепторных сигнальных систем. 1. Semaphorins и их рецепторы Plexin-
- 70. Формирование связей между нейронами включает несколько этапов. Эпигенетические влияния. Как зрительные (V), так и моторные (M)
- 71. Элиминация нейронов. В процессе развития нервной системы происходит элиминация «ненужных» нейронов. В зависимости от региона мозга
- 72. TUNEL Апоптоз при развитии неокортекса
- 74. Скачать презентацию

Происхождение нервных клеток -Evolution of nervous systems
Нейронная теория - Neuron doctrine
Гены
Происхождение нервных клеток -Evolution of nervous systems
Нейронная теория - Neuron doctrine
Гены
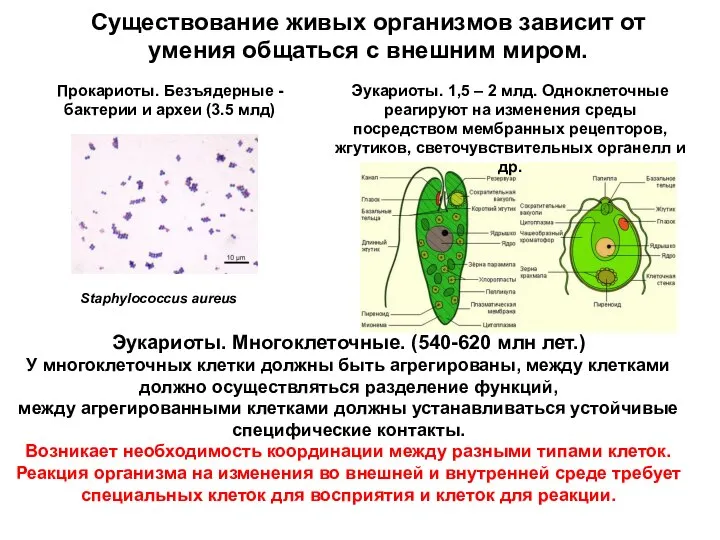
Существование живых организмов зависит от умения общаться с внешним миром.
Эукариоты. Многоклеточные.
Существование живых организмов зависит от умения общаться с внешним миром.
Эукариоты. Многоклеточные.

Появление нервных клеток в организме связано непосредственно с реактивностью на внешние
Появление нервных клеток в организме связано непосредственно с реактивностью на внешние

Теории происхождения и эволюции нервных клеток
1. Теория Н. Клейненберга (1872)- нейроны
Теории происхождения и эволюции нервных клеток
1. Теория Н. Клейненберга (1872)- нейроны
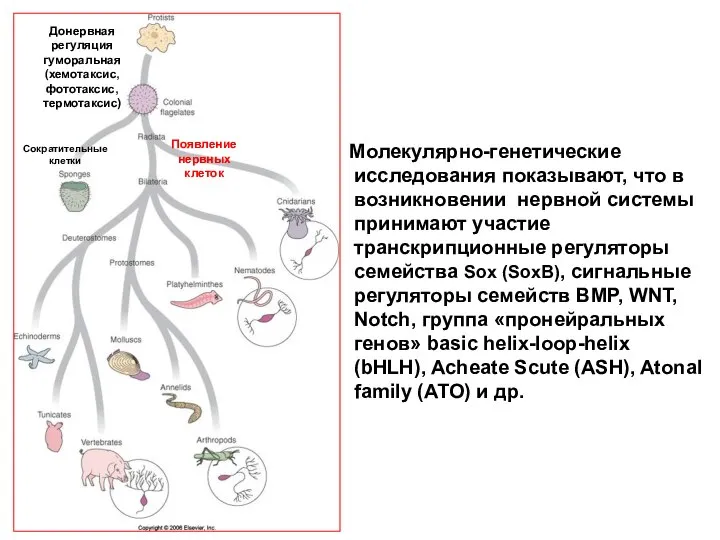
Молекулярно-генетические исследования показывают, что в возникновении нервной системы принимают участие
Молекулярно-генетические исследования показывают, что в возникновении нервной системы принимают участие

Sponges (губки)
Есть сократительные клетки, но нет нервных.
Cnidarian (гидра) – появляются нервные
Sponges (губки)
Есть сократительные клетки, но нет нервных.
Cnidarian (гидра) – появляются нервные
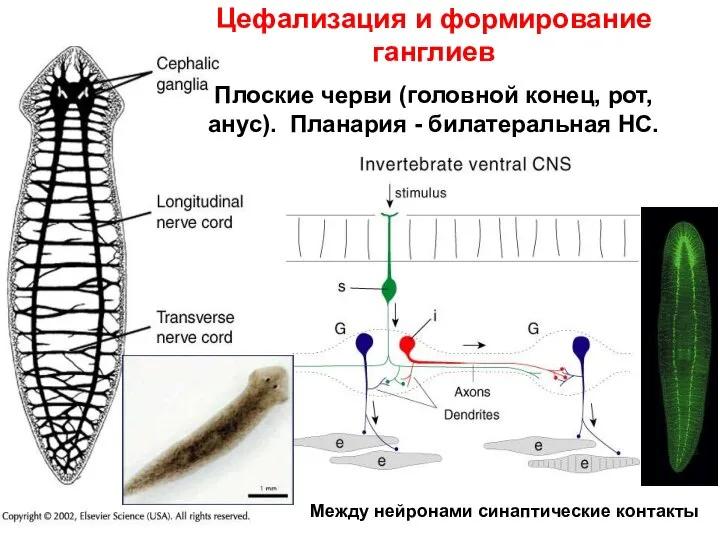
Цефализация и формирование ганглиев
Плоские черви (головной конец, рот, анус). Планария -
Цефализация и формирование ганглиев
Плоские черви (головной конец, рот, анус). Планария -
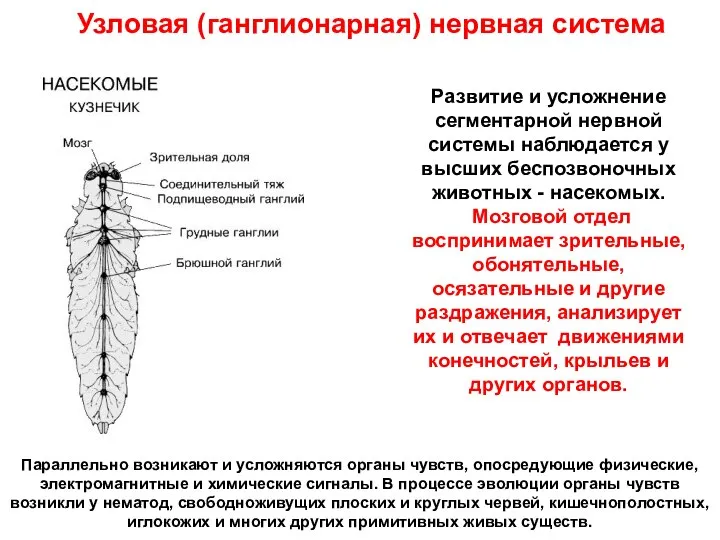
Развитие и усложнение сегментарной нервной системы наблюдается у высших беспозвоночных животных
Развитие и усложнение сегментарной нервной системы наблюдается у высших беспозвоночных животных
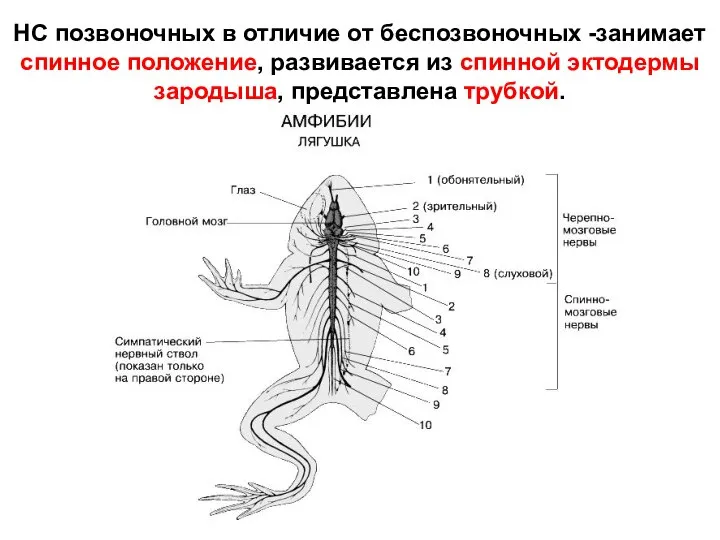
НС позвоночных в отличие от беспозвоночных -занимает спинное положение, развивается из
НС позвоночных в отличие от беспозвоночных -занимает спинное положение, развивается из
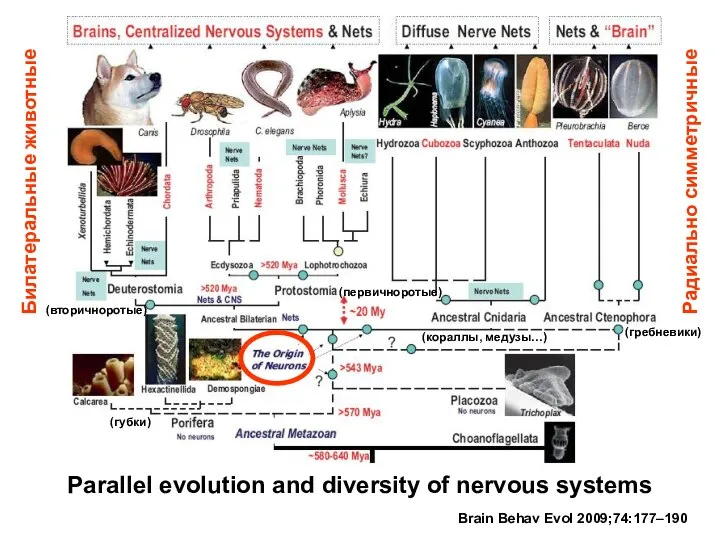
Parallel evolution and diversity of nervous systems
Brain Behav Evol 2009;74:177–190
(гребневики)
(губки)
(кораллы, медузы…)
(вторичноротые)
(первичноротые)
Билатеральные
Parallel evolution and diversity of nervous systems
Brain Behav Evol 2009;74:177–190
(гребневики)
(губки)
(кораллы, медузы…)
(вторичноротые)
(первичноротые)
Билатеральные
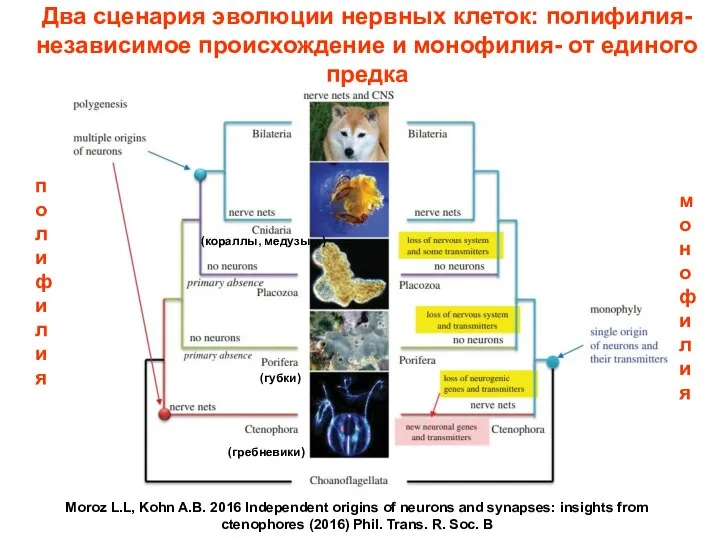
Два сценария эволюции нервных клеток: полифилия- независимое происхождение и монофилия- от
Два сценария эволюции нервных клеток: полифилия- независимое происхождение и монофилия- от
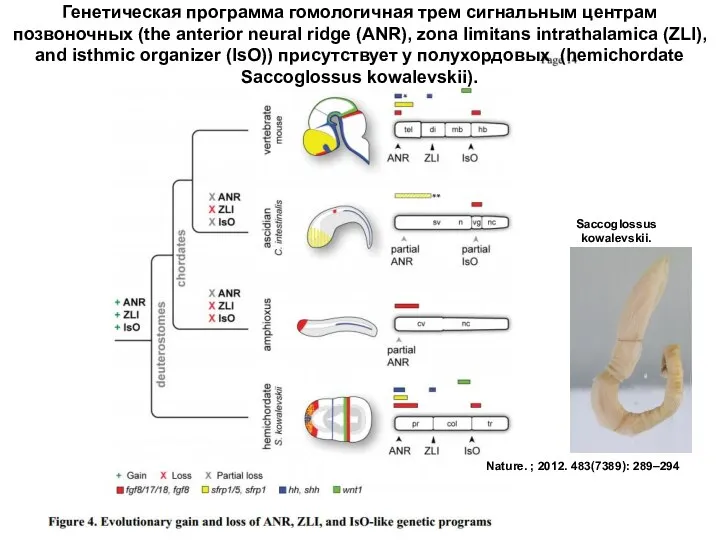
Nature. ; 2012. 483(7389): 289–294
Генетическая программа гомологичная трем сигнальным центрам позвоночных
Nature. ; 2012. 483(7389): 289–294
Генетическая программа гомологичная трем сигнальным центрам позвоночных

Геном человека содержит примерно 30000 генов и около 50% из них
Геном человека содержит примерно 30000 генов и около 50% из них
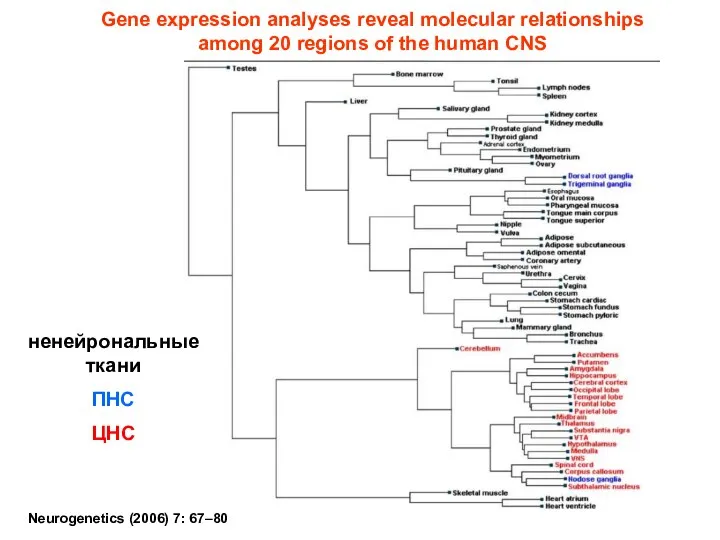
Neurogenetics (2006) 7: 67–80
Gene expression analyses reveal molecular relationships
among 20 regions
Neurogenetics (2006) 7: 67–80
Gene expression analyses reveal molecular relationships
among 20 regions
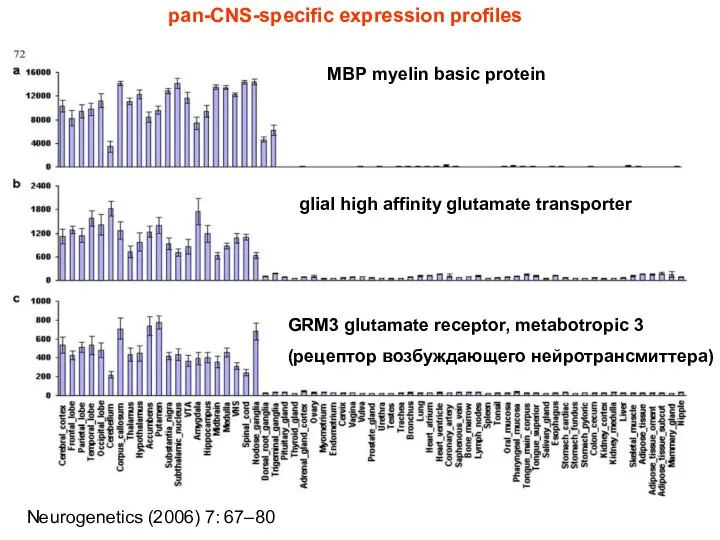
MBP myelin basic protein
glial high affinity glutamate transporter
GRM3 glutamate receptor, metabotropic
MBP myelin basic protein
glial high affinity glutamate transporter
GRM3 glutamate receptor, metabotropic

анализаторную (анализ поступающей по сенсорным системам информации),
интегративную (объединение, связь сигналов
анализаторную (анализ поступающей по сенсорным системам информации),
интегративную (объединение, связь сигналов
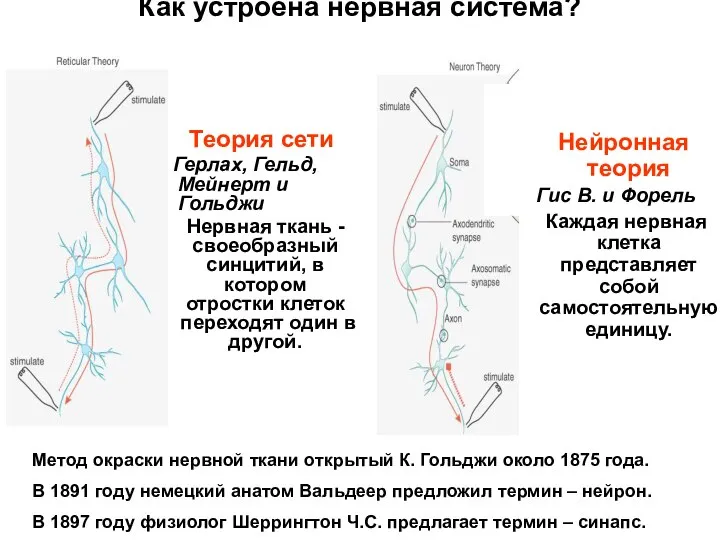
Как устроена нервная система?
Теория сети
Герлах, Гельд, Мейнерт и Гольджи
Как устроена нервная система?
Теория сети
Герлах, Гельд, Мейнерт и Гольджи

S. Ramon y Cajal
1852-1934
Создатель «Нейронной теории»
1. Каждый нейрон является
S. Ramon y Cajal
1852-1934
Создатель «Нейронной теории»
1. Каждый нейрон является


В основе развития нервной системы лежат процессы:
Индукция эктодермы
Детерминация и позиционная
В основе развития нервной системы лежат процессы:
Индукция эктодермы
Детерминация и позиционная
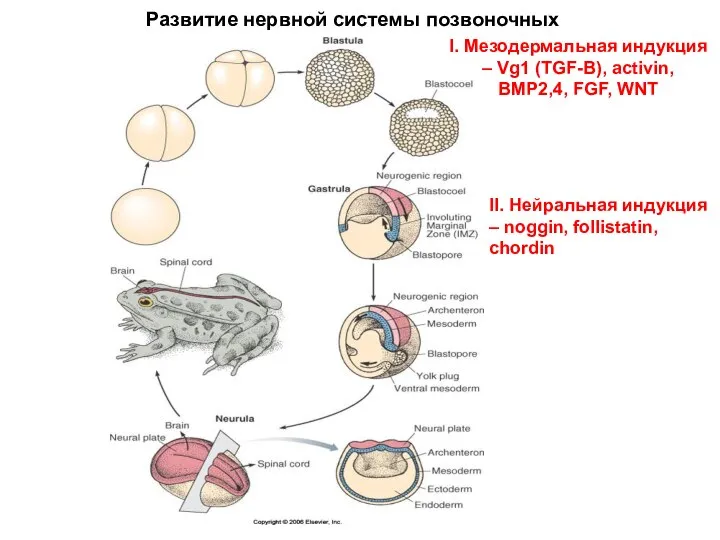
Развитие нервной системы позвоночных
I. Мезодермальная индукция – Vg1 (TGF-B), activin, BMP2,4,
Развитие нервной системы позвоночных
I. Мезодермальная индукция – Vg1 (TGF-B), activin, BMP2,4,
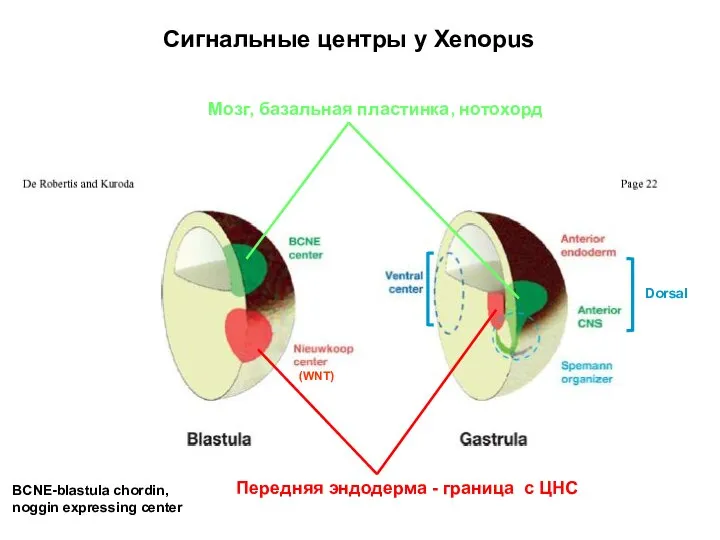
Мозг, базальная пластинка, нотохорд
Передняя эндодерма - граница c ЦНС
Сигнальные центры у
Мозг, базальная пластинка, нотохорд
Передняя эндодерма - граница c ЦНС
Сигнальные центры у
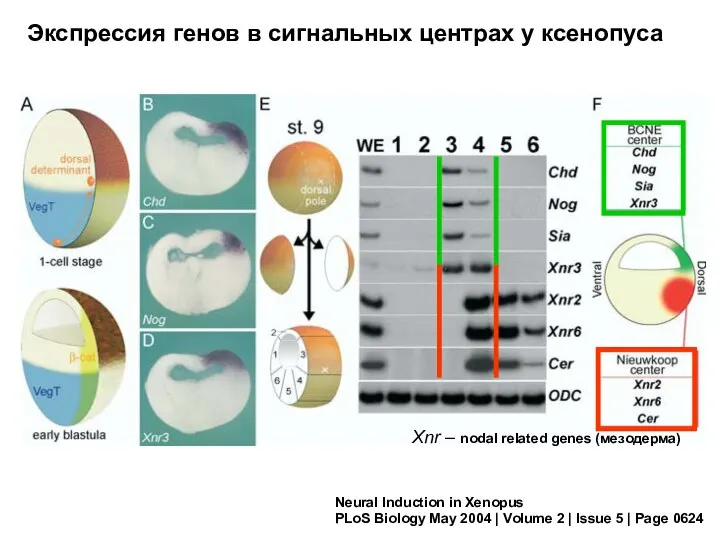
Neural Induction in Xenopus
PLoS Biology May 2004 | Volume 2
Neural Induction in Xenopus
PLoS Biology May 2004 | Volume 2
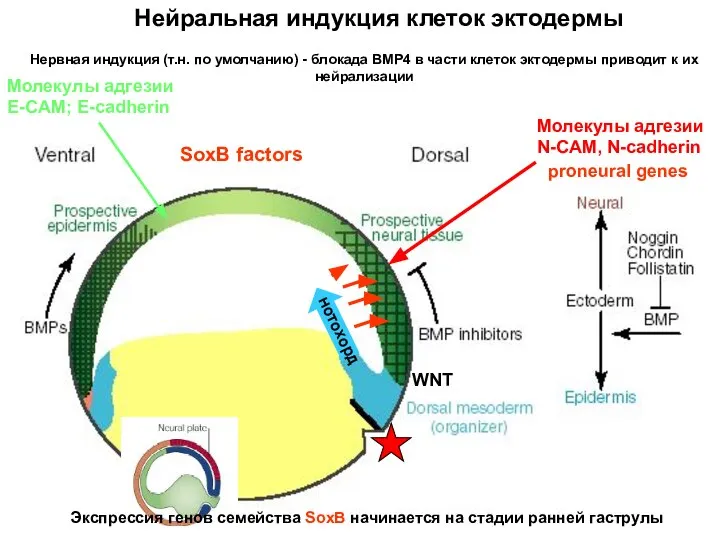
Нервная индукция (т.н. по умолчанию) - блокада BMP4 в части клеток
Нервная индукция (т.н. по умолчанию) - блокада BMP4 в части клеток
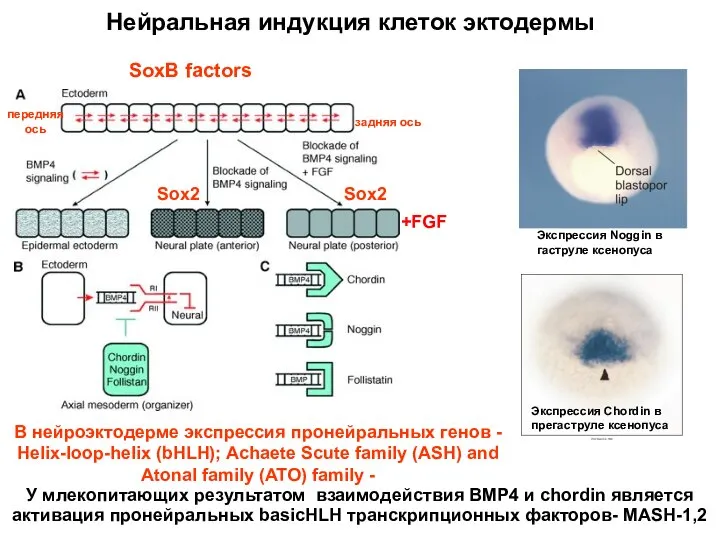
У млекопитающих результатом взаимодействия BMP4 и chordin является активация пронейральных basicHLH
У млекопитающих результатом взаимодействия BMP4 и chordin является активация пронейральных basicHLH
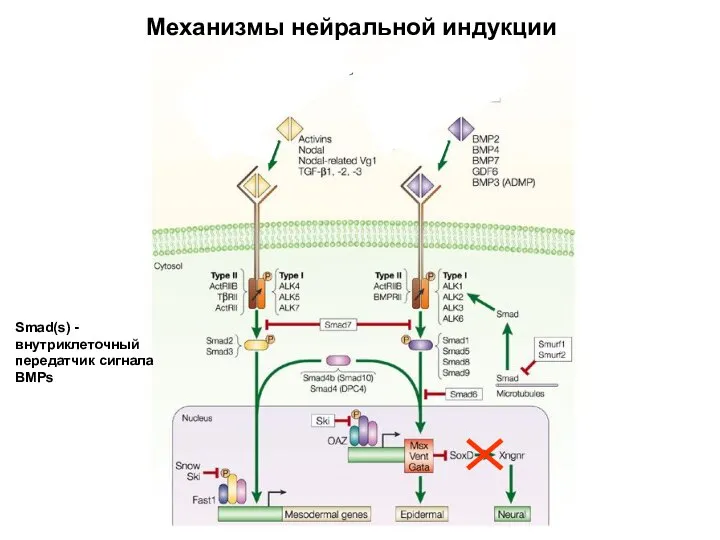
Механизмы нейральной индукции
Smad(s) - внутриклеточный передатчик сигнала BMPs
Механизмы нейральной индукции
Smad(s) - внутриклеточный передатчик сигнала BMPs
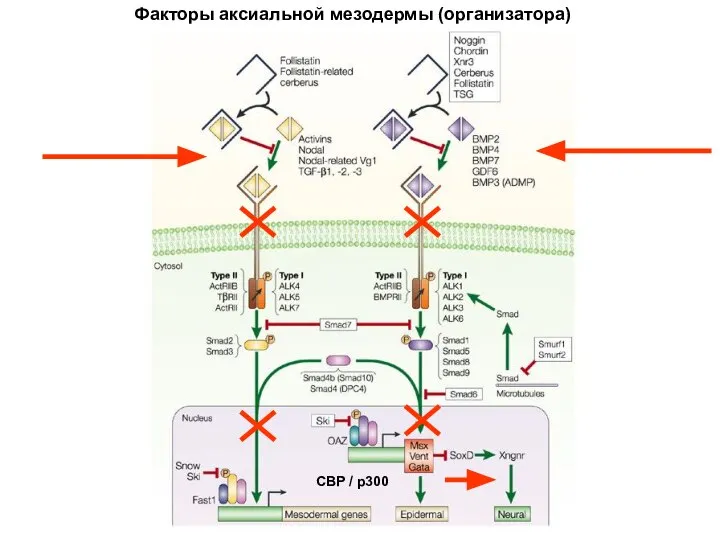
Факторы аксиальной мезодермы (организатора)
CBP / p300
Факторы аксиальной мезодермы (организатора)
CBP / p300
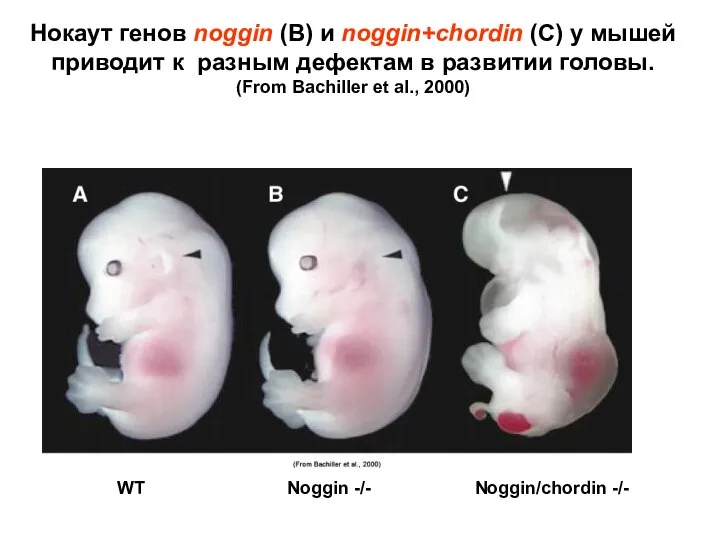
Нокаут генов noggin (В) и noggin+chordin (С) у мышей приводит к
Нокаут генов noggin (В) и noggin+chordin (С) у мышей приводит к
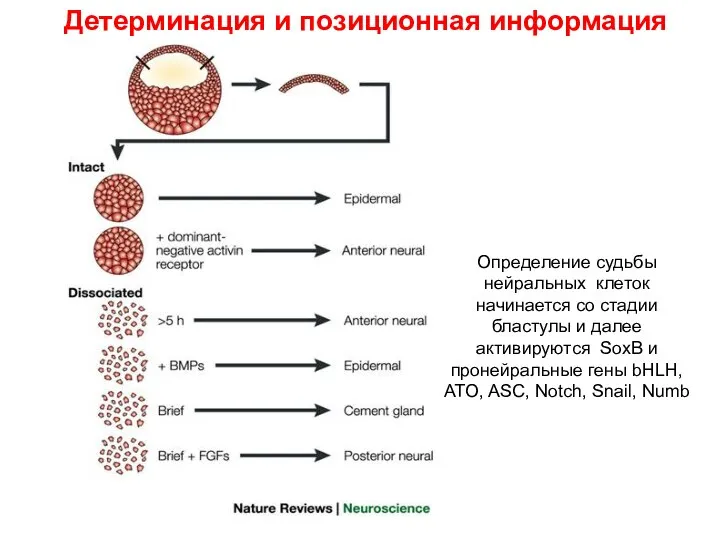
Детерминация и позиционная информация
Определение судьбы нейральных клеток начинается со стадии
Детерминация и позиционная информация
Определение судьбы нейральных клеток начинается со стадии

Модели развития нервной системы:
Мозаичная модель – детерминирована
судьба отдельной клетки (беспозвоночные).
Регуляционная
Модели развития нервной системы:
Мозаичная модель – детерминирована
судьба отдельной клетки (беспозвоночные).
Регуляционная
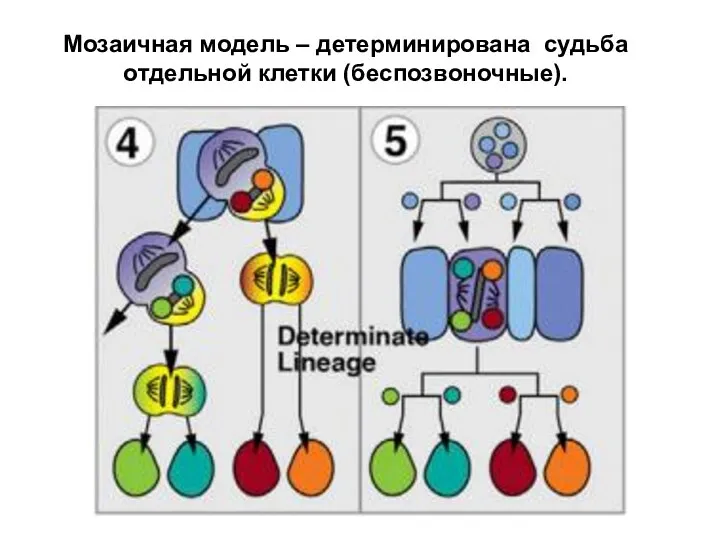
Мозаичная модель – детерминирована судьба отдельной клетки (беспозвоночные).
Мозаичная модель – детерминирована судьба отдельной клетки (беспозвоночные).
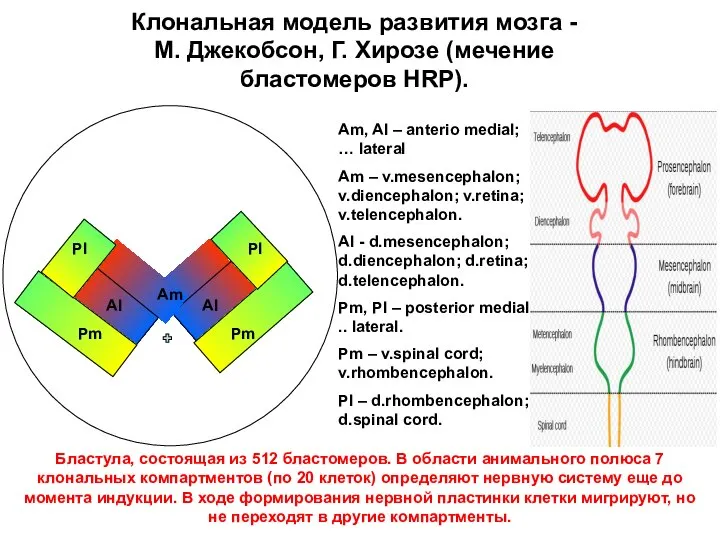
Аm
Al
Al
Pm
Pm
Pl
Pl
Клональная модель развития мозга - М. Джекобсон, Г. Хирозе (мечение бластомеров
Аm
Al
Al
Pm
Pm
Pl
Pl
Клональная модель развития мозга - М. Джекобсон, Г. Хирозе (мечение бластомеров
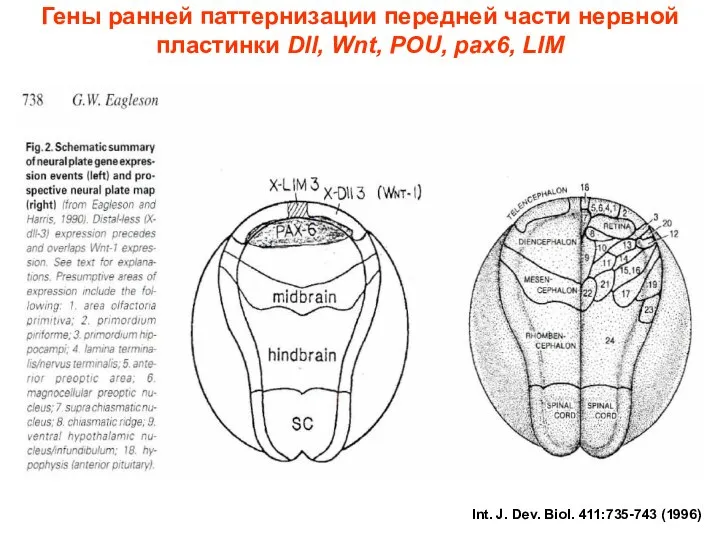
Int. J. Dev. BioI. 411:735-743 (1996)
Гены ранней паттернизации передней части нервной
Int. J. Dev. BioI. 411:735-743 (1996)
Гены ранней паттернизации передней части нервной
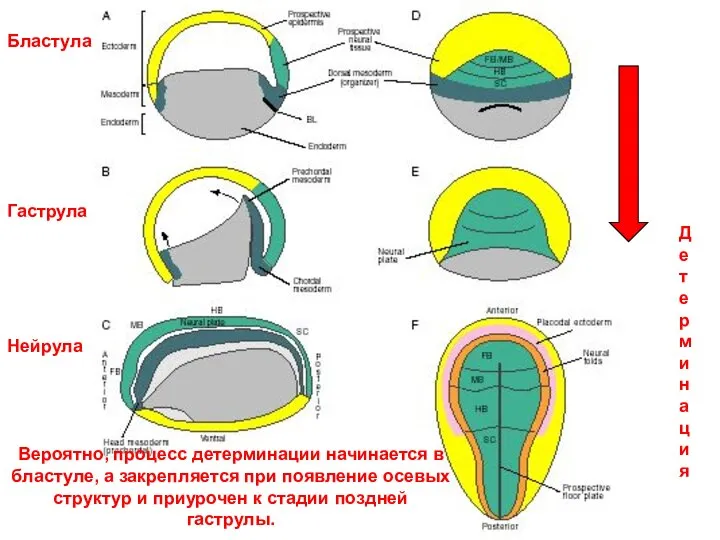
Бластула
Гаструла
Нейрула
Детерминация
Вероятно, процесс детерминации начинается в бластуле, а закрепляется при появление осевых
Бластула
Гаструла
Нейрула
Детерминация
Вероятно, процесс детерминации начинается в бластуле, а закрепляется при появление осевых
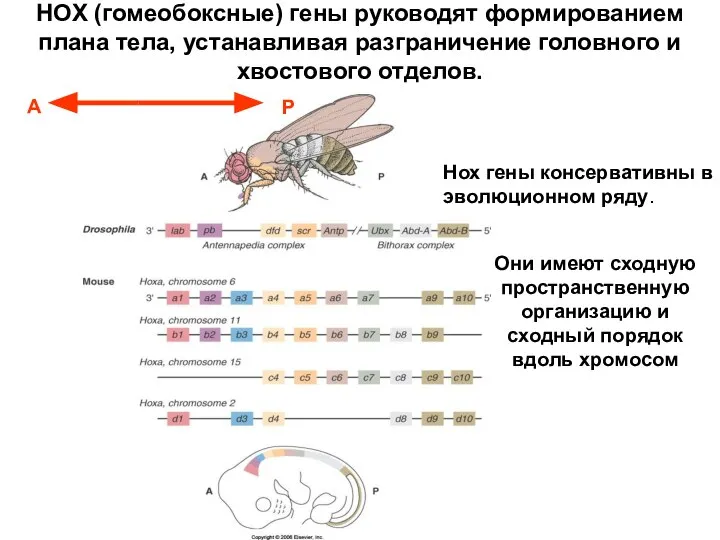
Они имеют сходную пространственную организацию и сходный порядок вдоль хромосом
Нох
Они имеют сходную пространственную организацию и сходный порядок вдоль хромосом
Нох
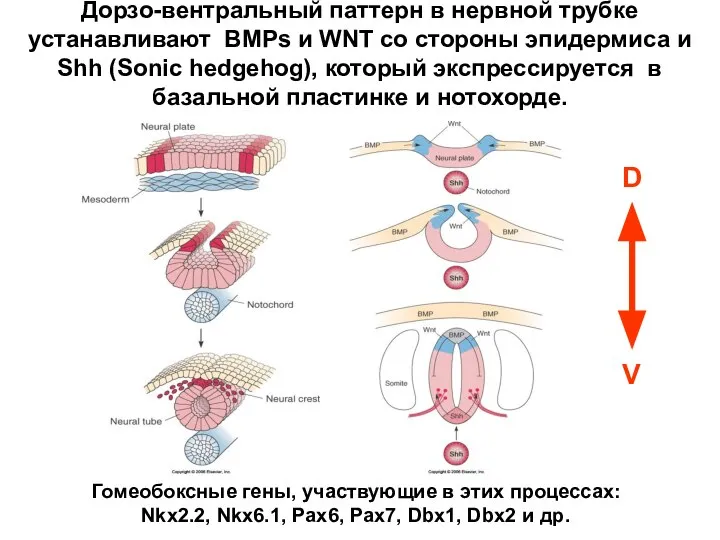
Дорзо-вентральный паттерн в нервной трубке устанавливают BMPs и WNT со стороны
Дорзо-вентральный паттерн в нервной трубке устанавливают BMPs и WNT со стороны
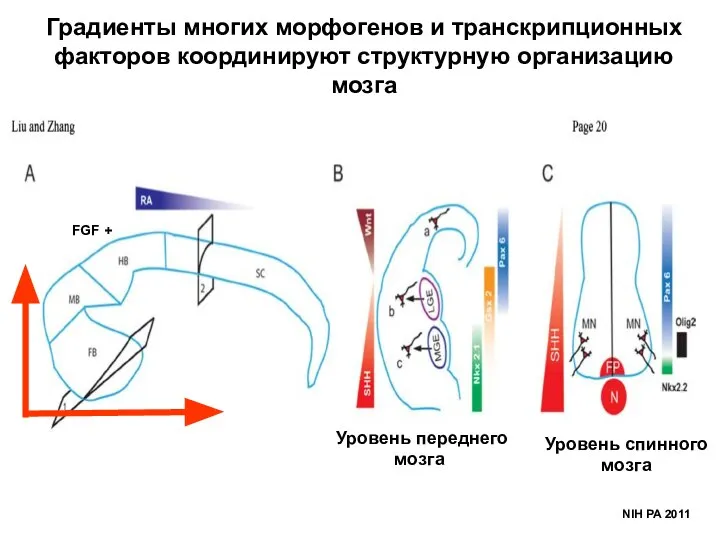
NIH PA 2011
Градиенты многих морфогенов и транскрипционных факторов координируют структурную организацию
NIH PA 2011
Градиенты многих морфогенов и транскрипционных факторов координируют структурную организацию
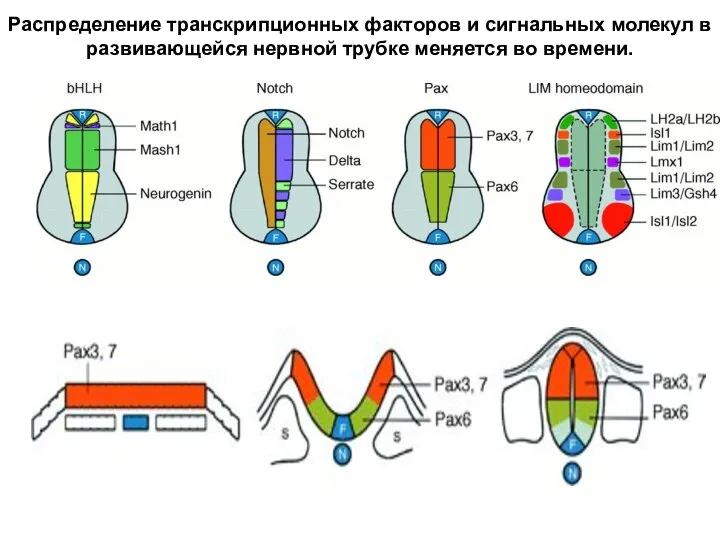
Распределение транскрипционных факторов и сигнальных молекул в развивающейся нервной трубке меняется
Распределение транскрипционных факторов и сигнальных молекул в развивающейся нервной трубке меняется

Важную роль в развитии и формировании структур переднего мозга играют регуляторные
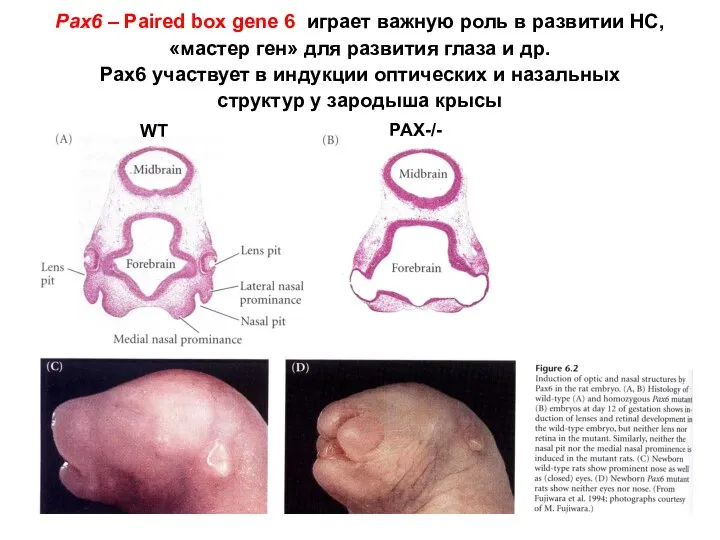
Pax6 – Paired box gene 6 играет важную роль в развитии
Pax6 – Paired box gene 6 играет важную роль в развитии
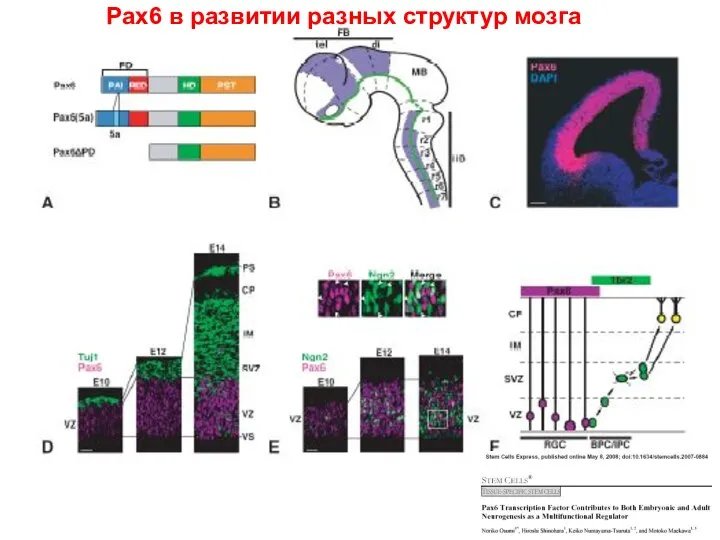
Pax6 в развитии разных структур мозга
Pax6 в развитии разных структур мозга
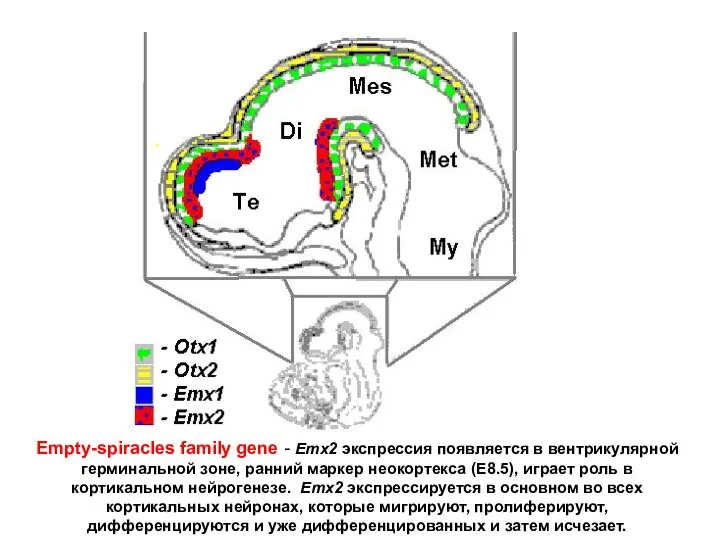
Empty-spiracles family gene - Emx2 экспрессия появляется в вентрикулярной герминальной зоне,
Empty-spiracles family gene - Emx2 экспрессия появляется в вентрикулярной герминальной зоне,
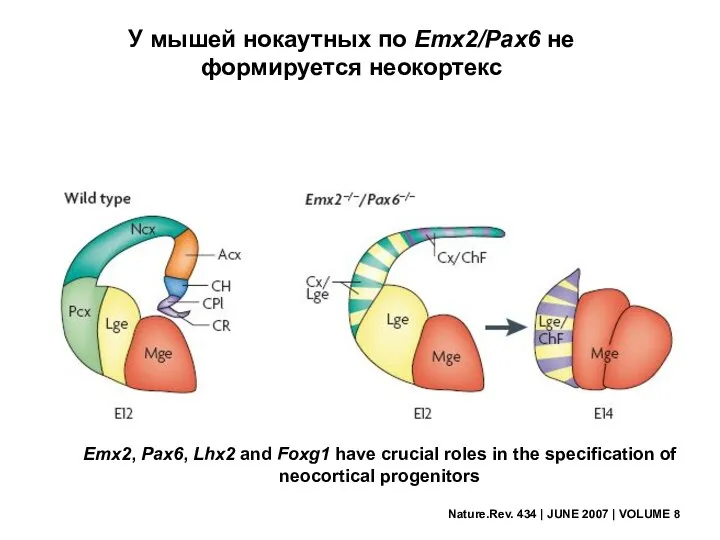
Nature.Rev. 434 | JUNE 2007 | VOLUME 8
У мышей нокаутных по
Nature.Rev. 434 | JUNE 2007 | VOLUME 8
У мышей нокаутных по
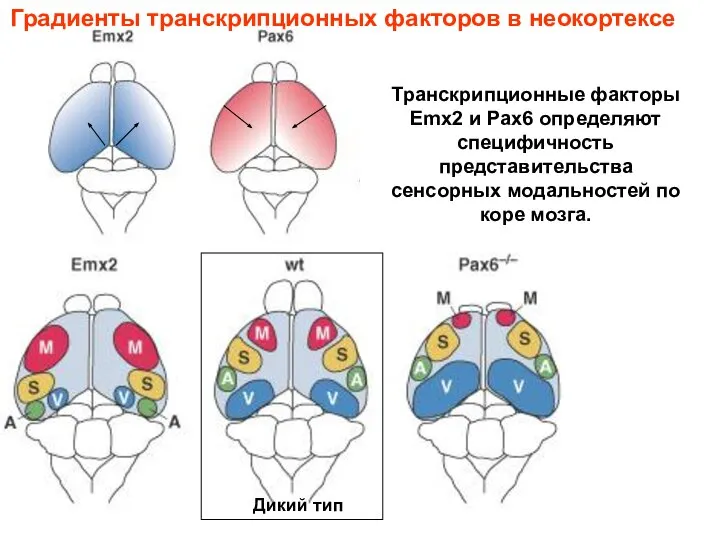
Транскрипционные факторы Emx2 и Pax6 определяют специфичность представительства сенсорных модальностей по
Транскрипционные факторы Emx2 и Pax6 определяют специфичность представительства сенсорных модальностей по
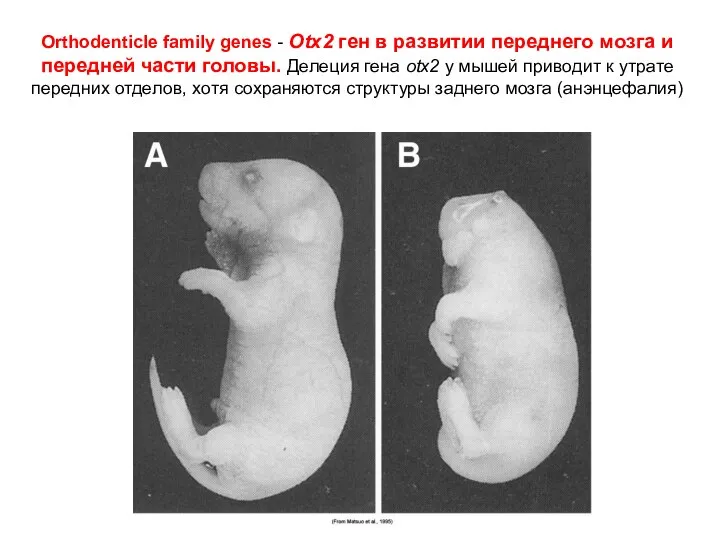
Orthodenticle family genes - Otx2 ген в развитии переднего мозга и
Orthodenticle family genes - Otx2 ген в развитии переднего мозга и
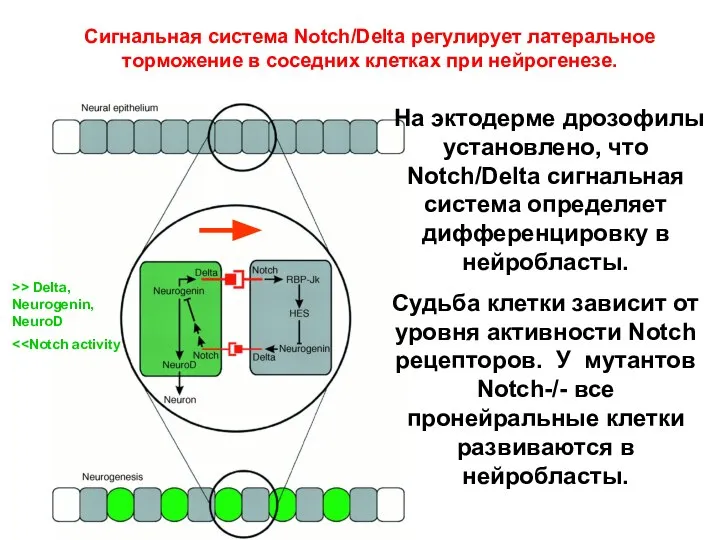
На эктодерме дрозофилы установлено, что Notch/Delta сигнальная система определяет дифференцировку в
На эктодерме дрозофилы установлено, что Notch/Delta сигнальная система определяет дифференцировку в
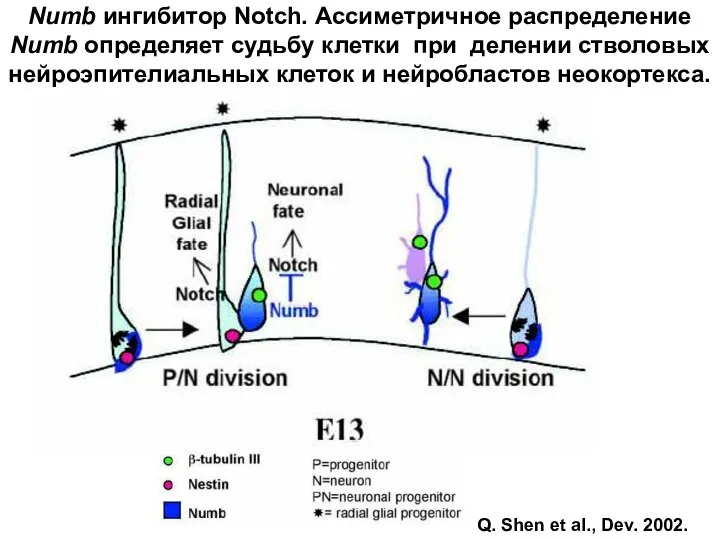
Numb ингибитор Notch. Ассиметричное распределение Numb определяет судьбу клетки при делении
Numb ингибитор Notch. Ассиметричное распределение Numb определяет судьбу клетки при делении
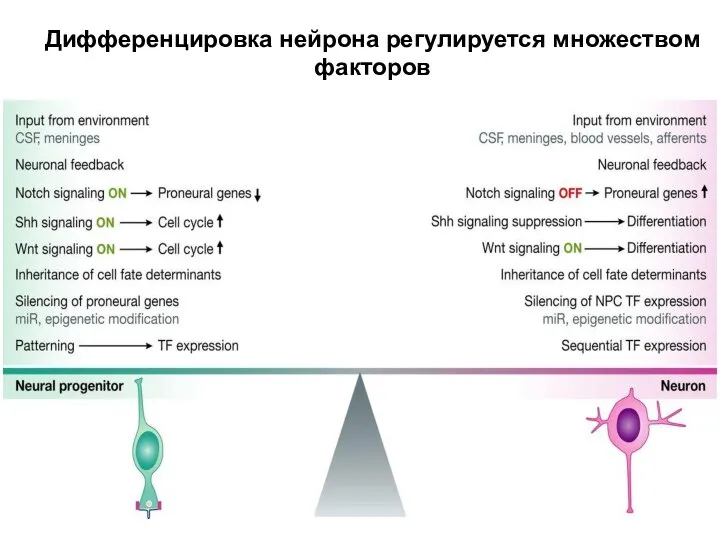
Дифференцировка нейрона регулируется множеством факторов
Дифференцировка нейрона регулируется множеством факторов
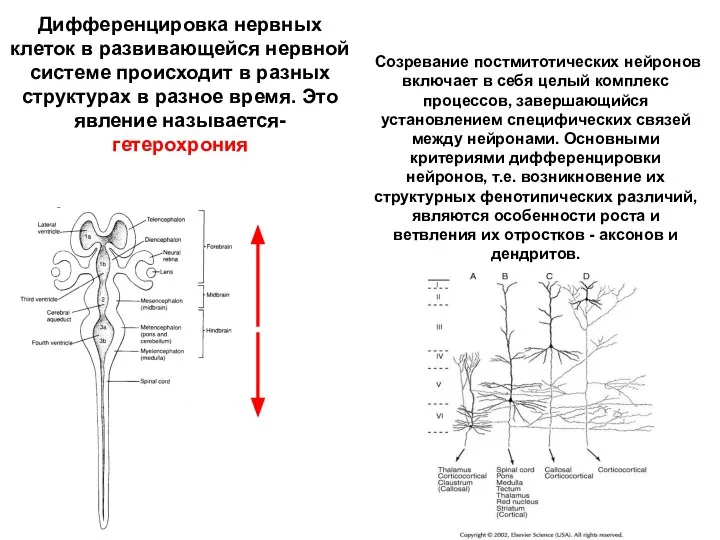
Дифференцировка нервных клеток в развивающейся нервной системе происходит в разных структурах
Дифференцировка нервных клеток в развивающейся нервной системе происходит в разных структурах
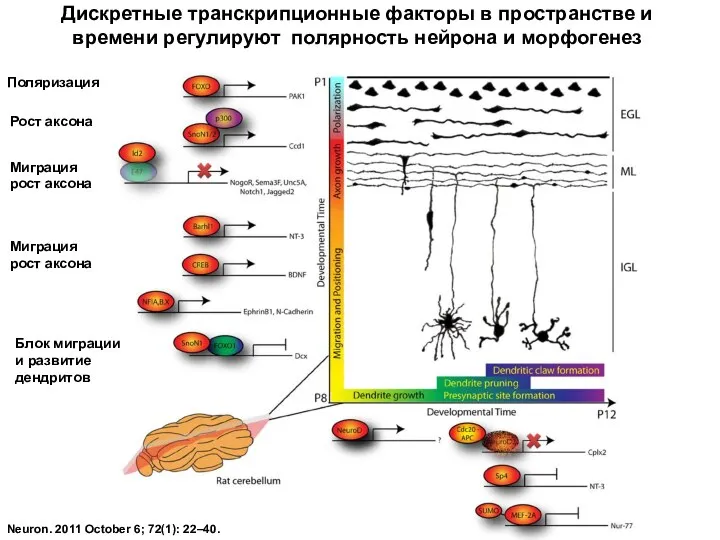
Neuron. 2011 October 6; 72(1): 22–40.
Дискретные транскрипционные факторы в пространстве и
Neuron. 2011 October 6; 72(1): 22–40.
Дискретные транскрипционные факторы в пространстве и
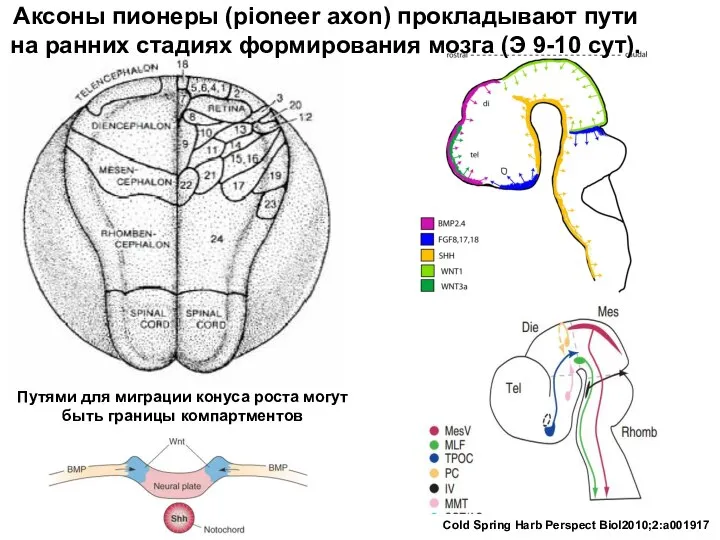
Путями для миграции конуса роста могут быть границы компартментов
Аксоны пионеры (pioneer
Путями для миграции конуса роста могут быть границы компартментов
Аксоны пионеры (pioneer
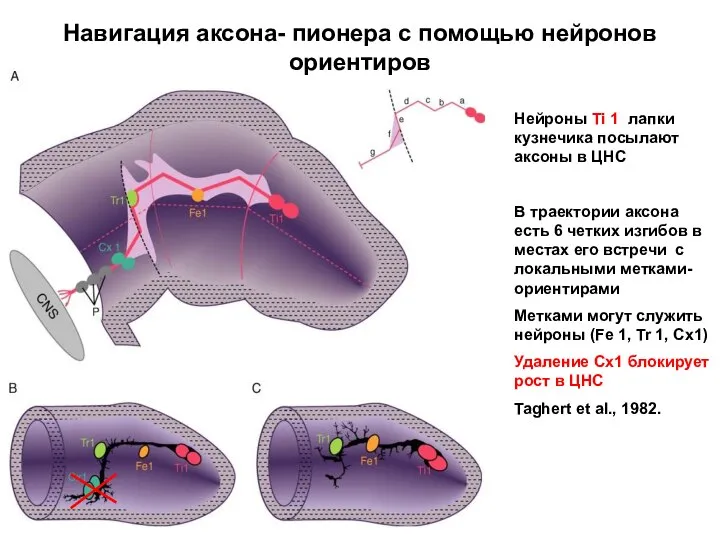
Навигация аксона- пионера с помощью нейронов ориентиров
Нейроны Ti 1 лапки кузнечика
Навигация аксона- пионера с помощью нейронов ориентиров
Нейроны Ti 1 лапки кузнечика
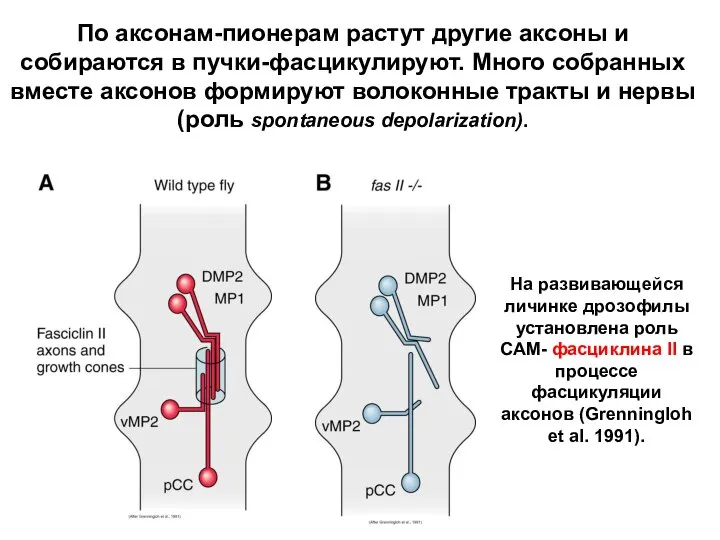
На развивающейся личинке дрозофилы установлена роль САМ- фасциклина II в процессе
На развивающейся личинке дрозофилы установлена роль САМ- фасциклина II в процессе

Соответствующие по времени воздействия комплексов специфических молекулярных факторов, являются критичными для
Соответствующие по времени воздействия комплексов специфических молекулярных факторов, являются критичными для

У млекопитающих нервные клетки формируются из двух источников:
1.Нервной трубки
2. Ганглиозной пластинки
У млекопитающих нервные клетки формируются из двух источников:
1.Нервной трубки
2. Ганглиозной пластинки
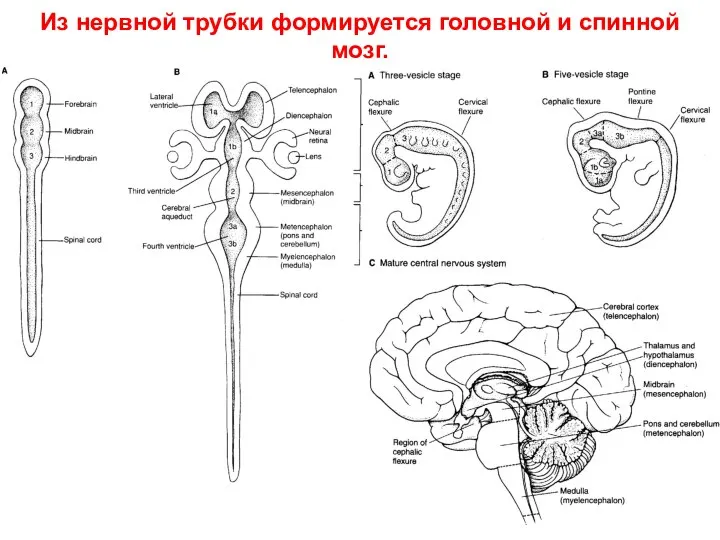
Из нервной трубки формируется головной и спинной мозг.
Из нервной трубки формируется головной и спинной мозг.
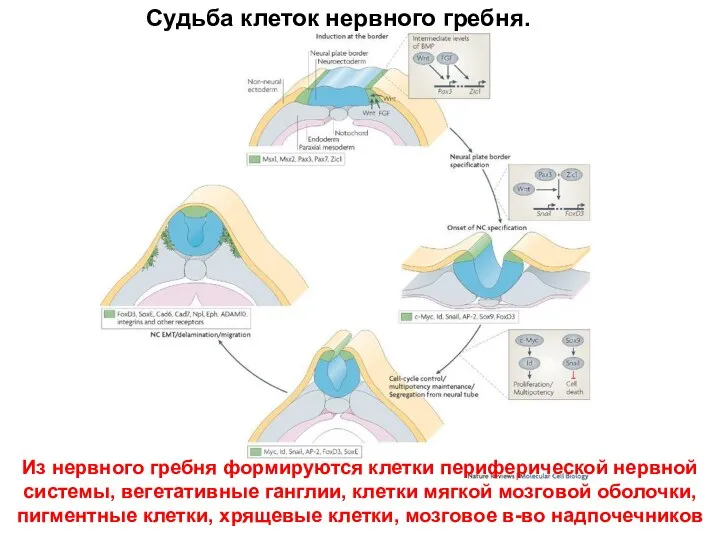
Судьба клеток нервного гребня.
Из нервного гребня формируются клетки периферической нервной системы,
Судьба клеток нервного гребня.
Из нервного гребня формируются клетки периферической нервной системы,
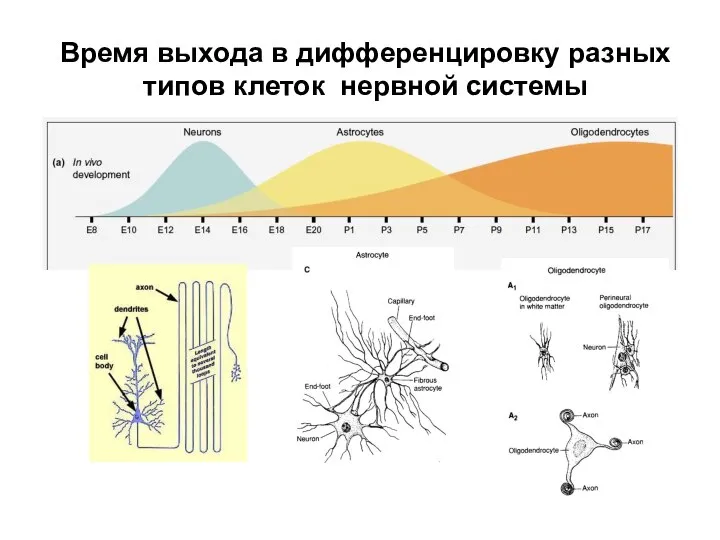
Время выхода в дифференцировку разных типов клеток нервной системы
Время выхода в дифференцировку разных типов клеток нервной системы
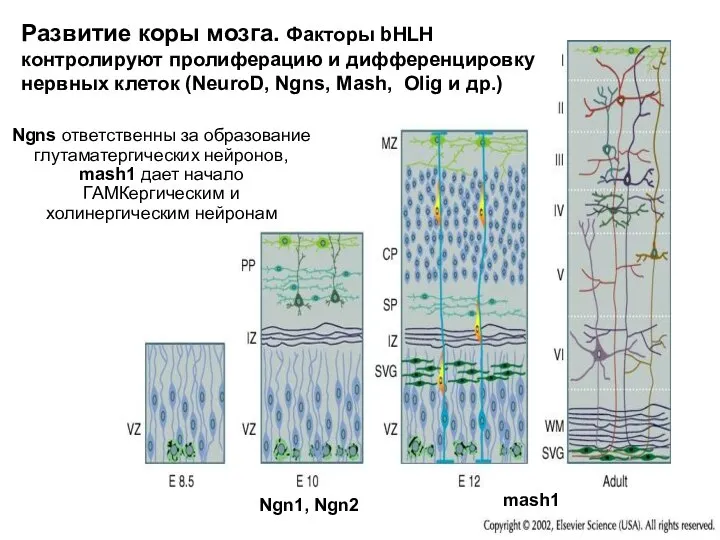
Развитие коры мозга. Факторы bHLH контролируют пролиферацию и дифференцировку нервных клеток
Развитие коры мозга. Факторы bHLH контролируют пролиферацию и дифференцировку нервных клеток
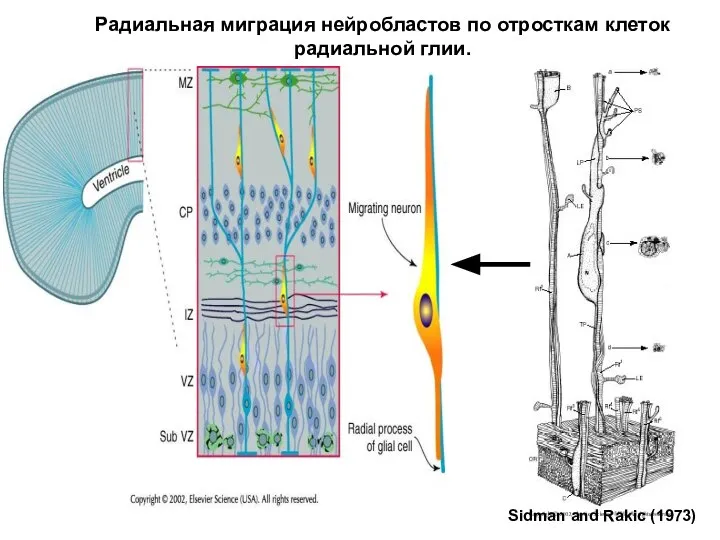
Радиальная миграция нейробластов по отросткам клеток радиальной глии.
Sidman and Rakic (1973)
Радиальная миграция нейробластов по отросткам клеток радиальной глии.
Sidman and Rakic (1973)
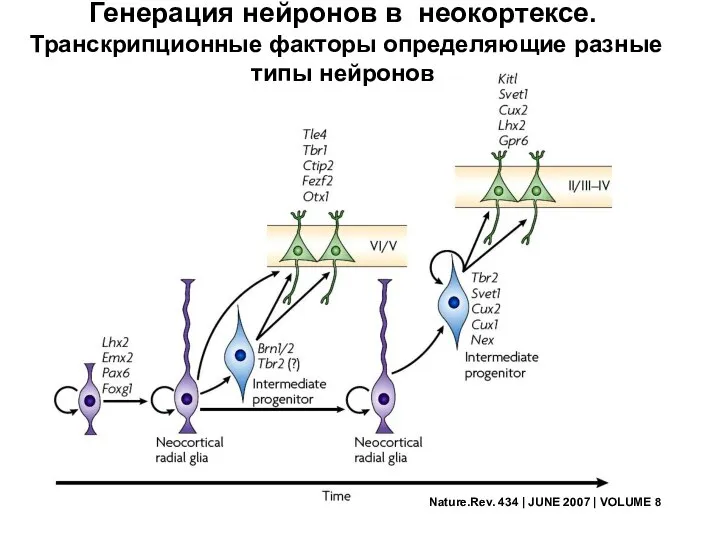
Nature.Rev. 434 | JUNE 2007 | VOLUME 8
Генерация нейронов в неокортексе.
Nature.Rev. 434 | JUNE 2007 | VOLUME 8
Генерация нейронов в неокортексе.
NATURE REVIEWS | NEUROSCIENCE VOLUME 8 |JUNE 2007 |
Развитие клеток в
NATURE REVIEWS | NEUROSCIENCE VOLUME 8 |JUNE 2007 |
Развитие клеток в
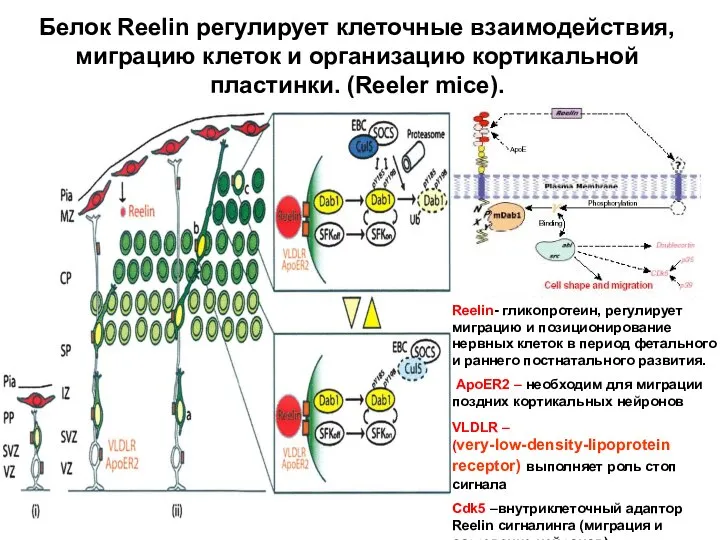
Белок Reelin регулирует клеточные взаимодействия, миграцию клеток и организацию кортикальной пластинки.
Белок Reelin регулирует клеточные взаимодействия, миграцию клеток и организацию кортикальной пластинки.
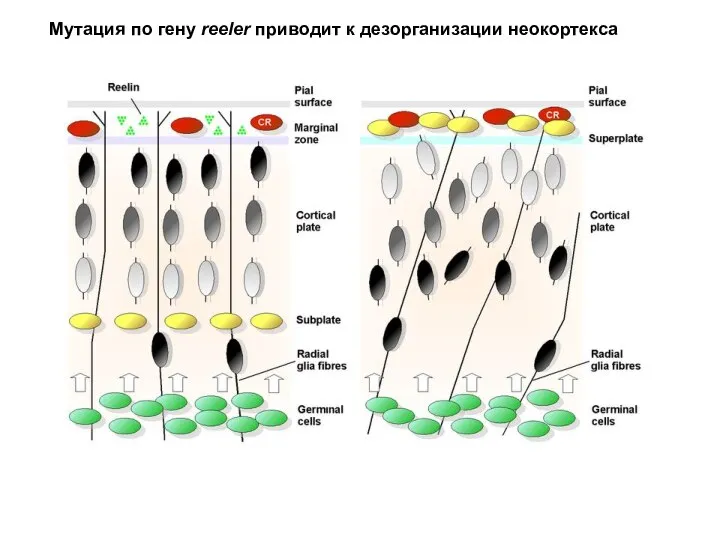
Мутация по гену reeler приводит к дезорганизации неокортекса
Мутация по гену reeler приводит к дезорганизации неокортекса
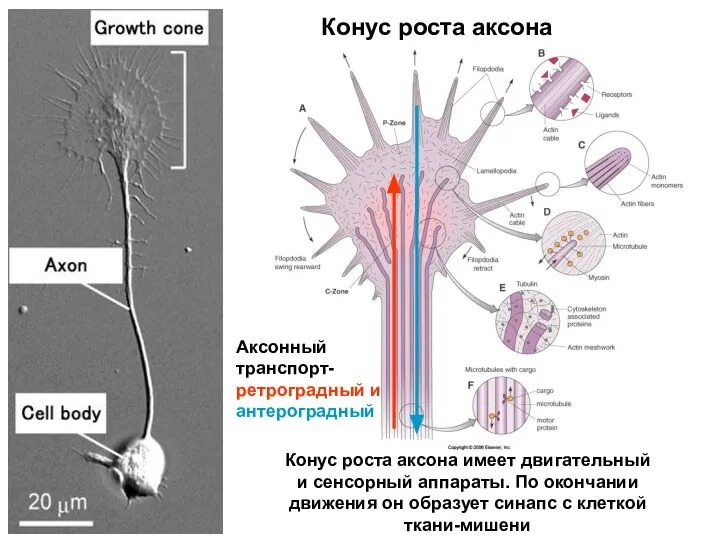
Конус роста аксона
Конус роста аксона имеет двигательный и сенсорный аппараты. По
Конус роста аксона
Конус роста аксона имеет двигательный и сенсорный аппараты. По
Молекулы – регуляторы роста аксона (конус роста)
Neural Development 2013, 8:17
Молекулы – регуляторы роста аксона (конус роста)
Neural Development 2013, 8:17
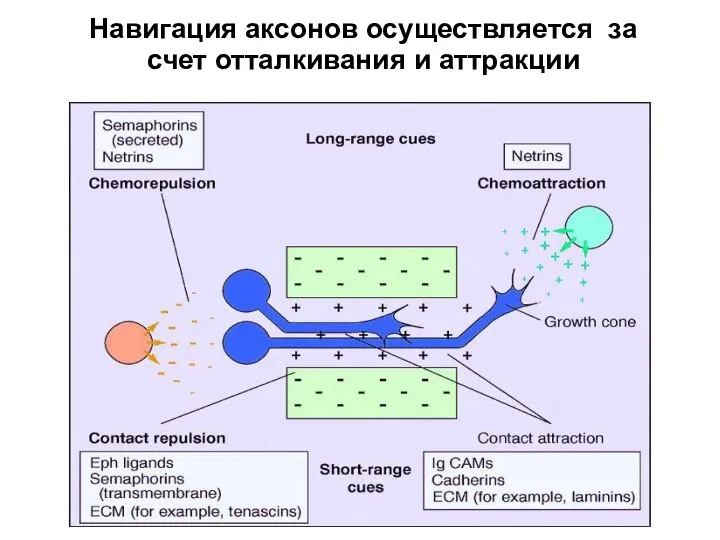
Навигация аксонов осуществляется за счет отталкивания и аттракции
Навигация аксонов осуществляется за счет отталкивания и аттракции

Хемотрофические молекулы навигаторы являются членами консервативных семейств лиганд/рецепторных сигнальных систем.
1.
Хемотрофические молекулы навигаторы являются членами консервативных семейств лиганд/рецепторных сигнальных систем.
1.
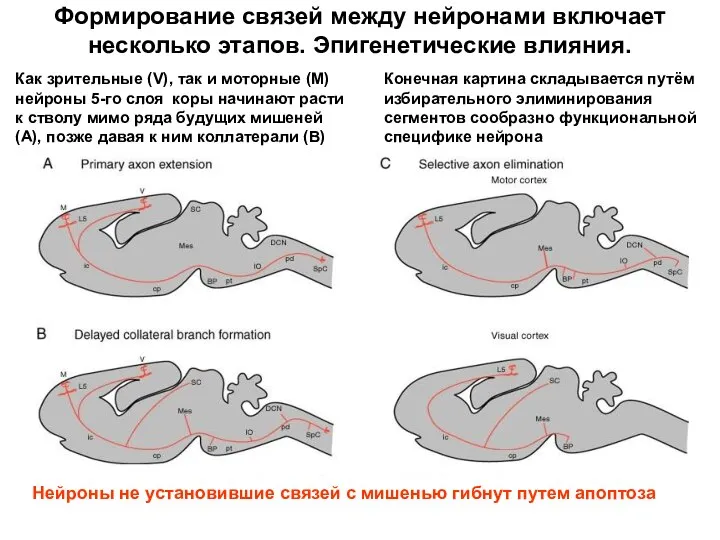
Формирование связей между нейронами включает несколько этапов. Эпигенетические влияния.
Как зрительные (V),
Формирование связей между нейронами включает несколько этапов. Эпигенетические влияния.
Как зрительные (V),

Элиминация нейронов.
В процессе развития нервной системы происходит элиминация «ненужных» нейронов. В
Элиминация нейронов.
В процессе развития нервной системы происходит элиминация «ненужных» нейронов. В
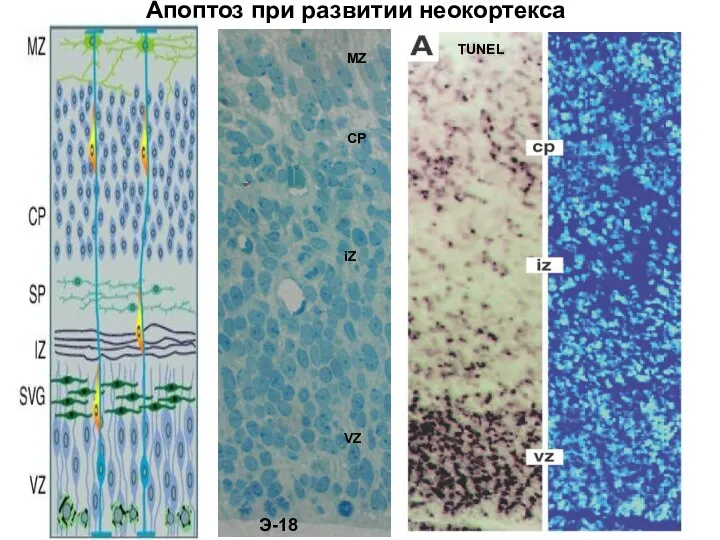
TUNEL
Апоптоз при развитии неокортекса
TUNEL
Апоптоз при развитии неокортекса
 Мутация. Мутацияның түрлері. Мутация тудыратын аурулар
Мутация. Мутацияның түрлері. Мутация тудыратын аурулар Презентация на тему Осенние и зимние явления в жизни растений
Презентация на тему Осенние и зимние явления в жизни растений  Презентация на тему Природа и безопасность . Ядовитые растения
Презентация на тему Природа и безопасность . Ядовитые растения БТ БФ Секвенирование
БТ БФ Секвенирование Окно в мир птиц Для тех, кто любит природу и неравнодушен к тому, что вокруг нас растет, бегает, плавает и щебечет, и для тех, кто не
Окно в мир птиц Для тех, кто любит природу и неравнодушен к тому, что вокруг нас растет, бегает, плавает и щебечет, и для тех, кто не Опорно-двигательная система. Значение опорно-двигательной системы. Строение костей
Опорно-двигательная система. Значение опорно-двигательной системы. Строение костей Строение и функции головного мозга
Строение и функции головного мозга Самый длинный язык у животных Язык у каждого вида животных имеет свое особое строение и определенную длину. Он очень важен для
Самый длинный язык у животных Язык у каждого вида животных имеет свое особое строение и определенную длину. Он очень важен для  История микробиологии в лицах
История микробиологии в лицах Клонування організмів
Клонування організмів Презентация на тему "Стебель" - скачать презентации по Биологии
Презентация на тему "Стебель" - скачать презентации по Биологии Вегетативное размножение растений
Вегетативное размножение растений Эти удивительные насекомые – бабочки
Эти удивительные насекомые – бабочки Подготовили: Анастасия Вайтехович, Анастасия Фатина, Светлана Лебедева Ученицы 9 «А» Руководитель: Пашутина Г.А. школы №63
Подготовили: Анастасия Вайтехович, Анастасия Фатина, Светлана Лебедева Ученицы 9 «А» Руководитель: Пашутина Г.А. школы №63 КУРИТЬ ИЛИ ЖИТЬ…? Курить всегда, курить везде.... И в морге, и на кладбище! Нас всюду никотин достанет. Найдет и по мозгам ударит. А может в сердце прямо пнет, А может в легкие воткнет.
КУРИТЬ ИЛИ ЖИТЬ…? Курить всегда, курить везде.... И в морге, и на кладбище! Нас всюду никотин достанет. Найдет и по мозгам ударит. А может в сердце прямо пнет, А может в легкие воткнет. Бактерии
Бактерии Генетика в кошках. Курс занимательной молекулярной генетики. Лекция I
Генетика в кошках. Курс занимательной молекулярной генетики. Лекция I Популяция элементарная единица эволюции. Изменчивость природных популяций
Популяция элементарная единица эволюции. Изменчивость природных популяций Комнатная герань
Комнатная герань Уход за волосами. Линия BIONIKA
Уход за волосами. Линия BIONIKA Презентация на тему "Внешнее строение корня" - скачать презентации по Биологии
Презентация на тему "Внешнее строение корня" - скачать презентации по Биологии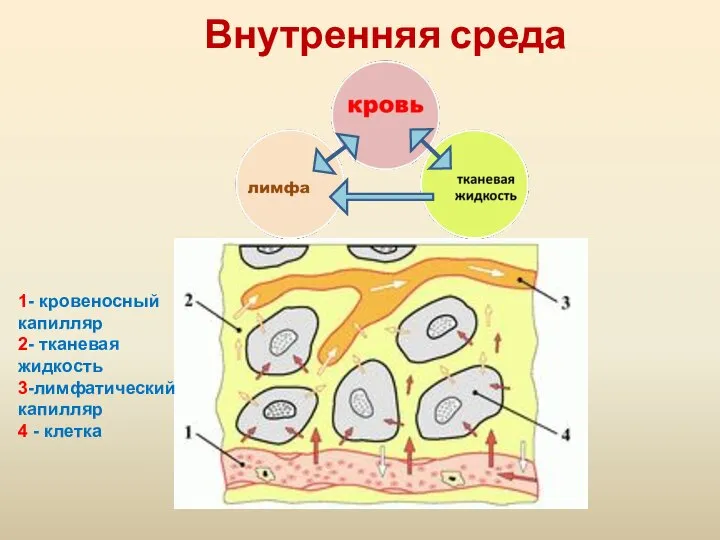 Внутренняя среда
Внутренняя среда Виды и разновидности фиалок
Виды и разновидности фиалок Как животные готовятся к зиме
Как животные готовятся к зиме Анализ пищевых продуктов на токсины
Анализ пищевых продуктов на токсины Домашнее животное кошка
Домашнее животное кошка Физиология нервов и нервно-мышечных синапсов
Физиология нервов и нервно-мышечных синапсов Ашыту өндірістерінде микроорганизмдерден практикалық бағалы өнім алуға пайдалану және олардың биохимиялық іс әрекеттері
Ашыту өндірістерінде микроорганизмдерден практикалық бағалы өнім алуға пайдалану және олардың биохимиялық іс әрекеттері