- Топография, структура и типы жировой ткани ,особенности метаболизма глюкозы в белой жировой ткани

Содержание
- 2. Жировая ткань представляет собой совокупность клеток адипоцитов (лат. adeps - жир, и kytos - полый пузырек),
- 3. Выделяют два типа расположения: центральный (абдоминальный, андроидный, тип яблока) и периферический (гиноидный, тип груши). 1.При центральном
- 4. Адипоциты белой жировой ткани - крупные клетки сферической формы, обычно диаметром от 30 до 70 мкм,
- 5. Функции: 1.Термоизолирующая - развитая жировая ткань образует теплоизоляционный слой, защищающий организм от действия очень низких температур.
- 6. Белые адипоциты - это резерв энергии! По сравнению с углеводами для целей накопления энергии жиры имеют
- 7. Запасание жира - липогенез Запасы триацилглицеринов в адипоцитах пополняются за счет использования жирных кислот , поступающих
- 8. Схема метаболизма глюкозы и липидов в адипоците.
- 9. Липолиз - новое прочтение Мобилизация жира В настоящее время, после открытия в 2004 году адипоцитарной триглицеридлипазы(ATGL),
- 10. В спокойном адипоците ATGL отщепляет жирные кислоты (С16-С18) во 2-м положении триацилглицерола с образованием продукта 1,3-ДАГ.
- 11. Регуляция липолиза в адипоците Основную роль в липолизе играет изменение состояния перилипина под воздействием протеинкиназы А.
- 12. Субстратный цикл Базальный распад и синтез триацилглицеролов в жировой ткани объединены в субстратный цикл, который заключается
- 13. Метаболические процессы белой жировой ткани Взрослые организмы практически не содержат бурого жира, его полностью заменяет жир
- 14. Если высокий уровень неэтерифицированных жирных кислот держится долгое время, нарушается тканевая чувствительность к инсулину и нарушается
- 15. Терморегуляция у новорожденных. Чертой реактивности новорожденных детей в течение первых недель после рождения является большая интенсивность
- 16. Особенности терморегуляции у новорожденных: 1.Новорожденные обеспечивают свои потребности в термогенезе с помощью недрожательного механизма, который невозможно
- 18. Скачать презентацию

Жировая ткань представляет собой совокупность клеток адипоцитов (лат. adeps - жир, и kytos - полый пузырек), главной функцией
Жировая ткань представляет собой совокупность клеток адипоцитов (лат. adeps - жир, и kytos - полый пузырек), главной функцией

Выделяют два типа расположения:
центральный (абдоминальный, андроидный, тип яблока) и периферический (гиноидный, тип груши).
Выделяют два типа расположения:
центральный (абдоминальный, андроидный, тип яблока) и периферический (гиноидный, тип груши).

Адипоциты белой жировой ткани - крупные клетки сферической формы, обычно диаметром от

Функции:
1.Термоизолирующая - развитая жировая ткань образует теплоизоляционный слой, защищающий организм
Функции:
1.Термоизолирующая - развитая жировая ткань образует теплоизоляционный слой, защищающий организм

Белые адипоциты - это резерв энергии!
По сравнению с углеводами для
Белые адипоциты - это резерв энергии!
По сравнению с углеводами для

Запасание жира - липогенез
Запасы триацилглицеринов в адипоцитах пополняются за счет
Запасание жира - липогенез
Запасы триацилглицеринов в адипоцитах пополняются за счет
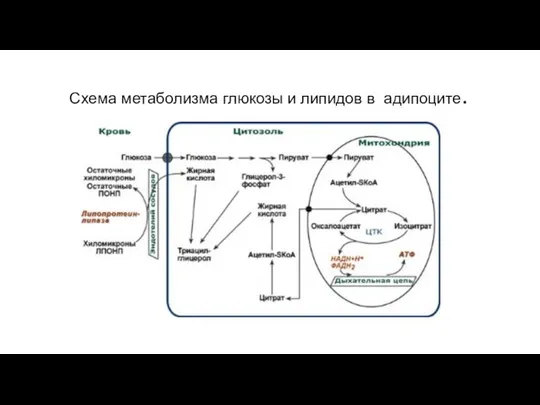
Схема метаболизма глюкозы и липидов в адипоците.
Схема метаболизма глюкозы и липидов в адипоците.

Липолиз - новое прочтение
Мобилизация жира
В настоящее время, после открытия
Липолиз - новое прочтение
Мобилизация жира
В настоящее время, после открытия

В спокойном адипоците
ATGL отщепляет жирные кислоты (С16-С18) во 2-м положении
В спокойном адипоците
ATGL отщепляет жирные кислоты (С16-С18) во 2-м положении
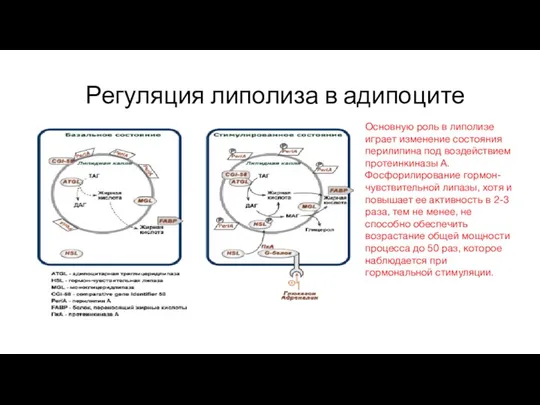
Регуляция липолиза в адипоците
Основную роль в липолизе играет изменение
Регуляция липолиза в адипоците
Основную роль в липолизе играет изменение

Субстратный цикл
Базальный распад и синтез триацилглицеролов в жировой ткани объединены в субстратный цикл, который
Субстратный цикл
Базальный распад и синтез триацилглицеролов в жировой ткани объединены в субстратный цикл, который

Метаболические процессы белой жировой ткани
Взрослые организмы практически не содержат бурого жира,
Метаболические процессы белой жировой ткани
Взрослые организмы практически не содержат бурого жира,

Если высокий уровень неэтерифицированных жирных кислот держится долгое время, нарушается тканевая

Терморегуляция у новорожденных.
Чертой реактивности новорожденных детей в течение первых недель
Терморегуляция у новорожденных.
Чертой реактивности новорожденных детей в течение первых недель

Особенности терморегуляции у новорожденных:
1.Новорожденные обеспечивают свои потребности в термогенезе с
Особенности терморегуляции у новорожденных:
1.Новорожденные обеспечивают свои потребности в термогенезе с
 Отличие человека от других живых существ
Отличие человека от других живых существ Водно-солевой обмен у животных
Водно-солевой обмен у животных Определение пола в животном мире Курс «Общая биология» Тема «Основы генетики» 9 класс Москва - 2009
Определение пола в животном мире Курс «Общая биология» Тема «Основы генетики» 9 класс Москва - 2009 Эукариотическая клетка. Цитоплазма
Эукариотическая клетка. Цитоплазма Вклад отечественных ученых в разработку учения о высшей нервной деятельности
Вклад отечественных ученых в разработку учения о высшей нервной деятельности Презентация на тему Класс Земноводные
Презентация на тему Класс Земноводные  Грибное место
Грибное место Презентация на тему "Химия в сельском хозяйстве" - скачать презентации по Биологии
Презентация на тему "Химия в сельском хозяйстве" - скачать презентации по Биологии Семя. Строение семян двудольных и однодольных растений
Семя. Строение семян двудольных и однодольных растений Презентация на тему Аристотель Заслуги в биологии
Презентация на тему Аристотель Заслуги в биологии Борьба за существование Формы борьбы за существование
Борьба за существование Формы борьбы за существование  Немембранные органоиды
Немембранные органоиды Презентация на тему "Презентація проекту: «Жири»" - скачать бесплатно презентации по Биологии
Презентация на тему "Презентація проекту: «Жири»" - скачать бесплатно презентации по Биологии Жизнь бабочек
Жизнь бабочек Биогеоценозы и биоценозы. Презентация Климовой Ирины Генриховны учителя биологии I квалификационной категории МОУ СОШ №11 г.Севе
Биогеоценозы и биоценозы. Презентация Климовой Ирины Генриховны учителя биологии I квалификационной категории МОУ СОШ №11 г.Севе Явление плазмолиза и деплазмолиза в клетках кожицы чешуи лука
Явление плазмолиза и деплазмолиза в клетках кожицы чешуи лука Презентация на тему "Породи гусей на території України" - скачать бесплатно презентации по Биологии
Презентация на тему "Породи гусей на території України" - скачать бесплатно презентации по Биологии Презентация на тему "Лес просит о помощи!" - презентации по Биологии
Презентация на тему "Лес просит о помощи!" - презентации по Биологии Семейства Бобовые, Лилейные, Злаковые
Семейства Бобовые, Лилейные, Злаковые Дендрологические парки
Дендрологические парки Понятие об обмене веществ и энергии. Методы изучения обмена энергии. Прямая и непрямая калориметрия
Понятие об обмене веществ и энергии. Методы изучения обмена энергии. Прямая и непрямая калориметрия Растения и их классификация
Растения и их классификация Презентация на тему "Красная книга растений (5 класс)" - скачать бесплатно презентации по Биологии
Презентация на тему "Красная книга растений (5 класс)" - скачать бесплатно презентации по Биологии Мхи
Мхи Презентация на тему "Половое и бесполое размножение организмов" - скачать презентации по Биологии
Презентация на тему "Половое и бесполое размножение организмов" - скачать презентации по Биологии Основы цитологии и гистологии человека
Основы цитологии и гистологии человека Тип губки. Многоклеточные организмы
Тип губки. Многоклеточные организмы Полярная медуза
Полярная медуза