- Трансгенная мышь с геном гормона роста

Содержание
- 2. Как определить экспрессию гена Найти мРНК данного гена с помощью молекулярного зонда Обнаружить белок с помощью
- 3. Регуляция экспрессии генов на разных уровнях
- 4. 1 уровень регуляции активности генов Это формирование активно транскрибируемого эухроматина и молчащего, компактизованного гетерохроматина Гетерохроматин подразделяется
- 5. Роль модификации и ремоделирования нуклеосом в регуляции генетических процессов Нуклеосома – октамер гистонов с намотанной ДНК
- 6. Модификация гистонов В регуляции активности генов
- 7. Нуклеосома – это октамер гистонов и 1,7 витка суперспирали фрагмента ДНК длиной 146 пар оснований В
- 8. Ремоделирование нуклеосом – это изменение их связывания с ДНК. Осуществляют белки Перемещение нуклеосом Изменение расстояния между
- 9. Ковалентная модификация включает – Ацетилирование – деацетилирование Фосфорилирование – дефосфорилирование Метилирование убиквитирование Метилирование лизина по положению
- 10. Гистоновый код- Аминокислотный остаток – разные АМК могут Подвергаться модификации – Лиз, Арг, Сер, Тре 2.
- 11. Регуляция изменением количества генов и структуры ДНК Утрата генетического материала –диминуция хроматина, разрушение ядра у эритроцитов
- 12. Активный хроматин – эухроматин Нуклеосомная упаковка изменена или отсутствует Длинные участки, чувствительные к ДНК-азе 1, указывающие
- 13. Метилирование ДНК Открыто еще до открытия Уотсона и Крика, но до сих пор остается много интригующих
- 14. Загадки- Отсутствует у дрозофилы и др. Обеспечивает накопление мутаций
- 15. У Dr дрожжейи C.elegans метилирования нет. Нарушение метилирования у человека приводит к остановке эмбрионального развития. Метилирование
- 16. Неактивный хроматин – гетерохроматин Конститутивный - центромера, теломера, интеркалярный Факультативный - при гаметогенезе в Х-хромосоме гетерохроматин
- 17. Транспозоны имеют рег элементы Транспозоны ограничены инвертированными повторами. При вырезании Т они сближаются и точно отрезаются
- 18. Величина генома может бстро увеличиваться за счет распространения транспозонов 4 хр-ма Др. буквально набита Т У-хр-ма
- 19. Регуляция на уровне репликации – В ядрышках ооцитов образуются экстрахромосомные копии генов рРНК. Этим достигается усиление
- 20. Регуляция генной активности на уровне транскрипции Регуляция транскрипции у прокариот( лактозный оперон) Позиционирование нуклеосом Регуляция транскрипции
- 21. Структура факторов транскрипции ДНК-связывающий домен Домены активации транскрипции Антирепрессорные домены Домены, связывающие лиганды Лиганды-индукторы –гормоны, ретиноевая
- 22. Типы факторов транскрипции
- 23. Факторы транскрипции – главные регуляторы генной активности
- 24. Транскрипция с разных промоторов В гене коллагена 43 экзона и 2 промотора варианты транскрипции приводят к
- 25. Регуляция экспрессии генов у прокариот – на примере лактозного оперона
- 26. В lac-опероне оператор перекрывает промотор и 5-конец 1 структурного гена
- 27. Метаболизм лактозы
- 28. При отсутствии в среде лактозы белок-репрессор связывается с оператором и препятствует соединению РНК-полимеразы с промотором и
- 29. При появлении лактозы репрессор связывается с ней, а не с оператором, транскрипция идет
- 32. Инсулятор блокирует действие энхансера, если инсулятор находится между энхансером и промотором
- 36. Активация транскрипции за счет связывания сap и сAMP
- 37. Регуляция транскрипции транспозонами (Т) Поскольку Т несут в своем составе регуляторные сигналы для Тр, (промоторы, энхансеры,
- 38. Т может изменить границы петель. Некоторые Т могут нести последовательности инсулятора, с которым связываются белки. Например,
- 39. Эукариотические мРНК довольно стабильны (часы и сутки)До выхода в цитоплазму они проходят процессинг. Поэтому часто регуляция
- 40. Посттранскрипционный уровень регуляции генной активности Транспорт мРНК из ядра Процессинг мРНК Альтернативный сплайсинг Редактирование мРНК (
- 41. Альтернативный сплайсинг как регулятор активности генов
- 42. Редактирование РНК – термин предложен Р. Бенни для феномена встраивания 4 У . В митохондриальный транскрипт
- 43. Биологические последствия редактирования РНК Образование пригодной для трансляции РНК И редактированная и нередактированная РНК могут быть
- 44. Регуляция генной активности на уровне трансляции Инициация трансляции с альтернативных сайтов Регуляция активности матрицы. Очень активные
- 45. Регуляция синтеза ферритина если железа в среде мало, то соответствующая мРНК не транслируется.Ингибирование происходит на стадии
- 46. Дискриминация мРНК- инициирующие участки РНК имеют разное сродство к рибосомам и поэтому с разной эффективностью связывают
- 47. Трансляционная репрессия – белок репрессор связывается с участком инициации трансляции и препятствует связыванию инициирующей рибосомы. Часто
- 48. Маскирование мРНК – осуществляется белком связывающимся с 3’ НТО. Как связывание с хвостом затыкает рот всей
- 49. Рнк не выходит из ядра, пока не закончится ее процессинг. У ВИЧ есть регулируемый ядерный транспорт.
- 50. Посттрансляционный уровень регуляции Фолдинг белка Роль гликозилирования, фосфорилирование, ацетилирование и др и др. модификации белка Удаление
- 51. Механизмы сайленсинга Гипотеза гистонового кода – ковалентные модификации ДНК и гистонов гипоацетилирование, убиквитирование и фосфорилирование гистонов
- 53. Скачать презентацию

Как определить экспрессию гена
Найти мРНК данного гена с помощью
молекулярного зонда
Обнаружить
Как определить экспрессию гена
Найти мРНК данного гена с помощью
молекулярного зонда
Обнаружить
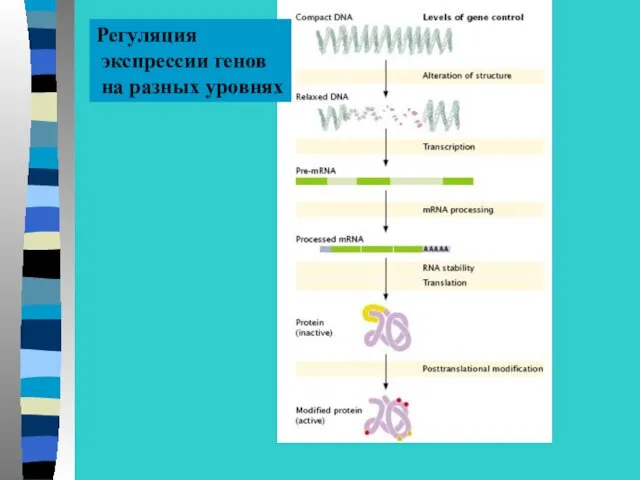
Регуляция
экспрессии генов
на разных уровнях
Регуляция
экспрессии генов
на разных уровнях

1 уровень регуляции активности генов
Это формирование активно транскрибируемого
эухроматина и молчащего,
1 уровень регуляции активности генов
Это формирование активно транскрибируемого
эухроматина и молчащего,

Роль модификации и ремоделирования нуклеосом
в регуляции генетических процессов
Нуклеосома – октамер
Роль модификации и ремоделирования нуклеосом
в регуляции генетических процессов
Нуклеосома – октамер
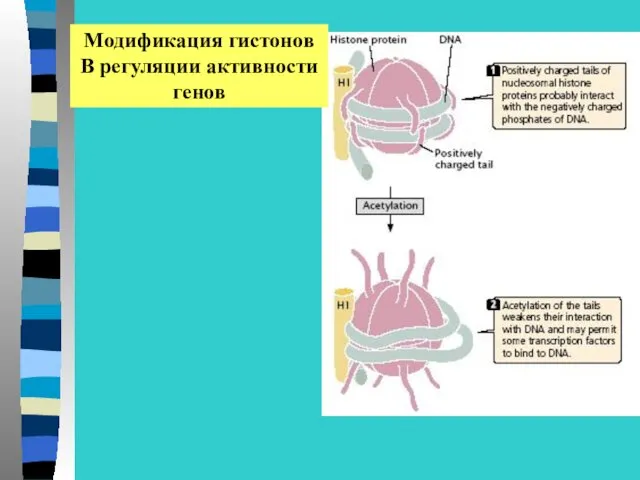
Модификация гистонов
В регуляции активности
генов
Модификация гистонов
В регуляции активности
генов

Нуклеосома – это октамер гистонов и 1,7 витка
суперспирали фрагмента ДНК
Нуклеосома – это октамер гистонов и 1,7 витка
суперспирали фрагмента ДНК

Ремоделирование нуклеосом – это изменение
их связывания с ДНК. Осуществляют белки
Перемещение
Ремоделирование нуклеосом – это изменение
их связывания с ДНК. Осуществляют белки
Перемещение

Ковалентная модификация включает –
Ацетилирование – деацетилирование
Фосфорилирование – дефосфорилирование
Метилирование
убиквитирование
Метилирование лизина по
Ковалентная модификация включает –
Ацетилирование – деацетилирование
Фосфорилирование – дефосфорилирование
Метилирование
убиквитирование
Метилирование лизина по

Гистоновый код-
Аминокислотный остаток – разные АМК могут
Подвергаться модификации –
Гистоновый код-
Аминокислотный остаток – разные АМК могут
Подвергаться модификации –

Регуляция изменением количества генов и структуры
ДНК
Утрата генетического материала –диминуция
хроматина,
Регуляция изменением количества генов и структуры
ДНК
Утрата генетического материала –диминуция
хроматина,

Активный хроматин – эухроматин
Нуклеосомная упаковка изменена или отсутствует
Длинные участки, чувствительные к
Активный хроматин – эухроматин
Нуклеосомная упаковка изменена или отсутствует
Длинные участки, чувствительные к

Метилирование ДНК
Открыто еще до открытия Уотсона и Крика, но до сих
Метилирование ДНК
Открыто еще до открытия Уотсона и Крика, но до сих

Загадки-
Отсутствует у дрозофилы и др.
Обеспечивает накопление мутаций
Загадки-
Отсутствует у дрозофилы и др.
Обеспечивает накопление мутаций

У Dr дрожжейи C.elegans метилирования нет. Нарушение метилирования у человека приводит
У Dr дрожжейи C.elegans метилирования нет. Нарушение метилирования у человека приводит

Неактивный хроматин – гетерохроматин
Конститутивный - центромера, теломера, интеркалярный
Факультативный - при гаметогенезе
Неактивный хроматин – гетерохроматин
Конститутивный - центромера, теломера, интеркалярный
Факультативный - при гаметогенезе

Транспозоны имеют рег элементы
Транспозоны ограничены инвертированными повторами. При вырезании Т они
Транспозоны имеют рег элементы
Транспозоны ограничены инвертированными повторами. При вырезании Т они

Величина генома может бстро увеличиваться за счет распространения транспозонов
4 хр-ма Др.
Величина генома может бстро увеличиваться за счет распространения транспозонов
4 хр-ма Др.

Регуляция на уровне репликации –
В ядрышках ооцитов образуются экстрахромосомные копии генов
Регуляция на уровне репликации –
В ядрышках ооцитов образуются экстрахромосомные копии генов

Регуляция генной активности на уровне транскрипции
Регуляция транскрипции у прокариот( лактозный оперон)
Позиционирование
Регуляция генной активности на уровне транскрипции
Регуляция транскрипции у прокариот( лактозный оперон)
Позиционирование

Структура факторов транскрипции
ДНК-связывающий домен
Домены активации транскрипции
Антирепрессорные домены
Домены, связывающие лиганды
Лиганды-индукторы –гормоны, ретиноевая
Структура факторов транскрипции
ДНК-связывающий домен
Домены активации транскрипции
Антирепрессорные домены
Домены, связывающие лиганды
Лиганды-индукторы –гормоны, ретиноевая
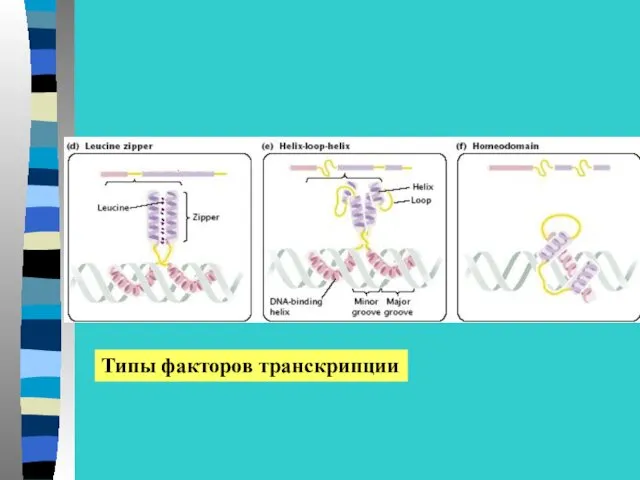
Типы факторов транскрипции
Типы факторов транскрипции
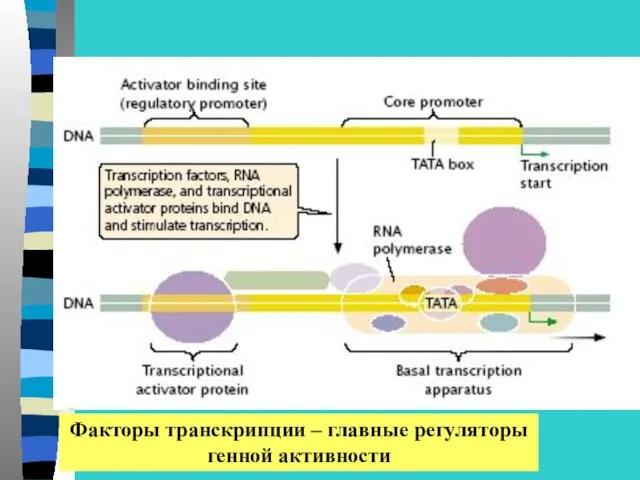
Факторы транскрипции – главные регуляторы
генной активности
Факторы транскрипции – главные регуляторы
генной активности

Транскрипция с разных промоторов
В гене коллагена 43 экзона и 2 промотора
Транскрипция с разных промоторов
В гене коллагена 43 экзона и 2 промотора

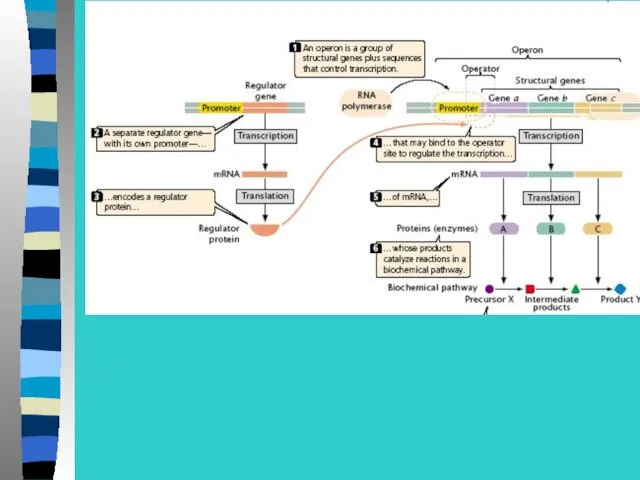
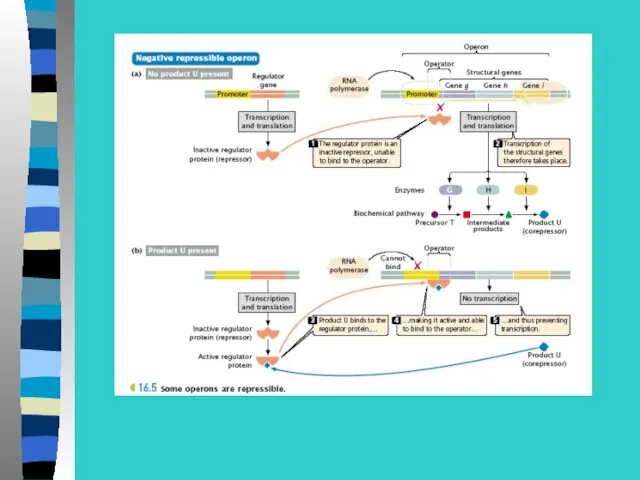
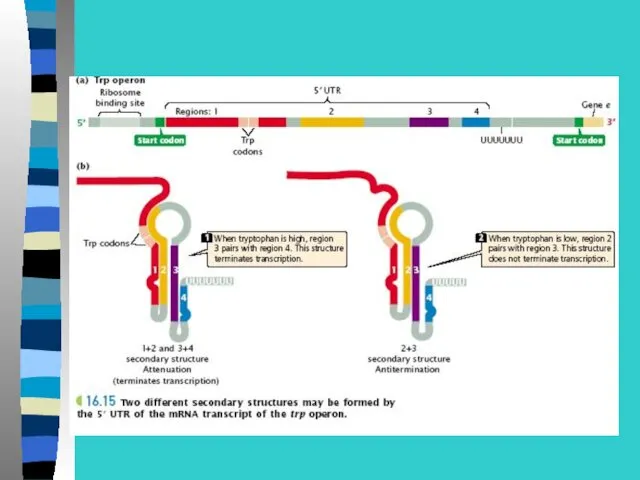
Регуляция экспрессии генов у прокариот –
на примере лактозного оперона
Регуляция экспрессии генов у прокариот –
на примере лактозного оперона
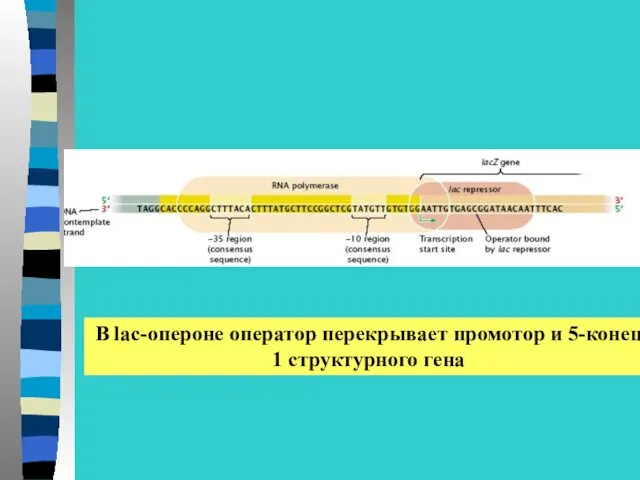
В lac-опероне оператор перекрывает промотор и 5-конец
1 структурного гена
В lac-опероне оператор перекрывает промотор и 5-конец
1 структурного гена
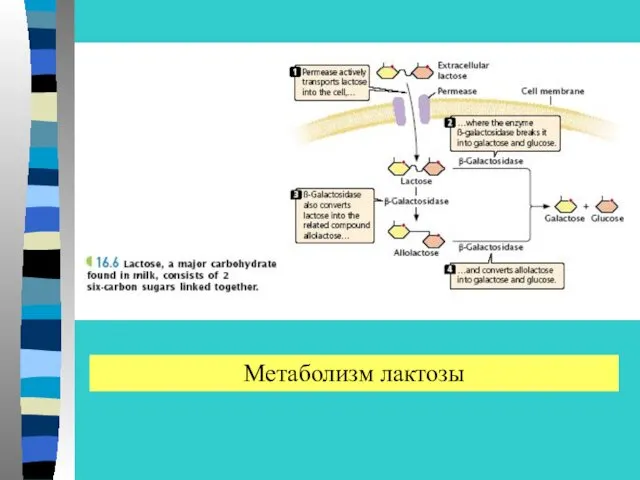
Метаболизм лактозы
Метаболизм лактозы
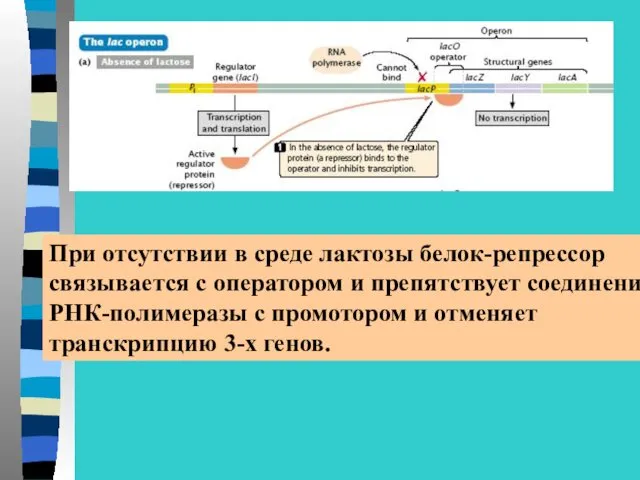
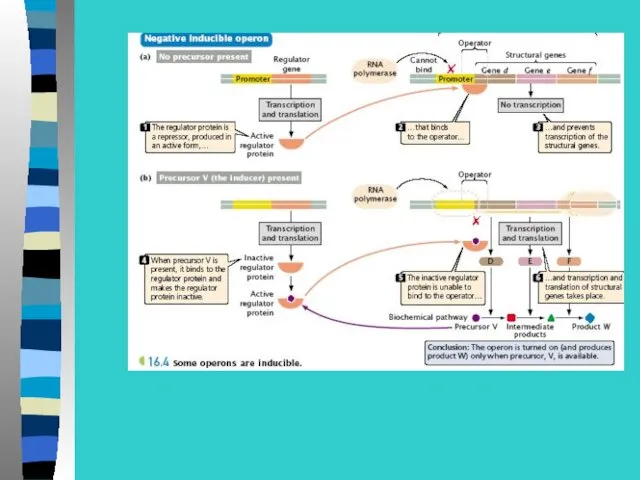
При отсутствии в среде лактозы белок-репрессор
связывается с оператором и препятствует
При отсутствии в среде лактозы белок-репрессор
связывается с оператором и препятствует
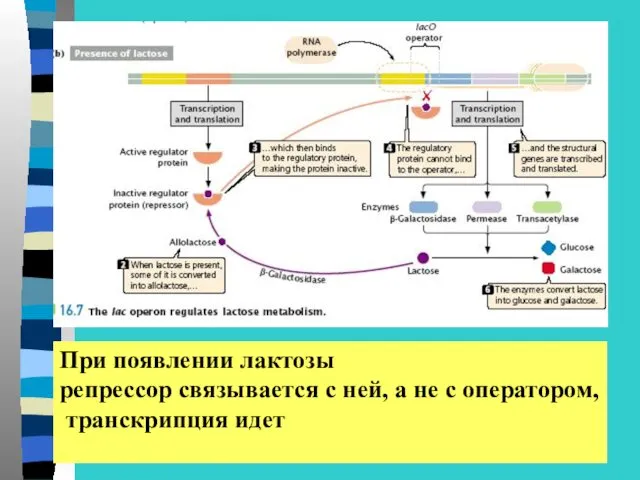
При появлении лактозы
репрессор связывается с ней, а не с оператором,
При появлении лактозы
репрессор связывается с ней, а не с оператором,
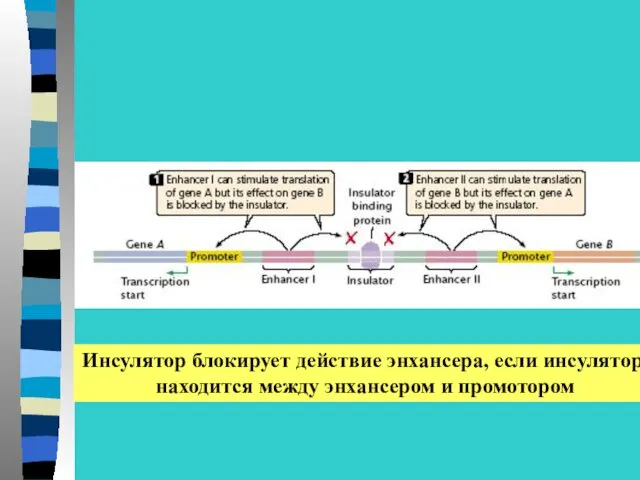
Инсулятор блокирует действие энхансера, если инсулятор
находится между энхансером и промотором
Инсулятор блокирует действие энхансера, если инсулятор
находится между энхансером и промотором

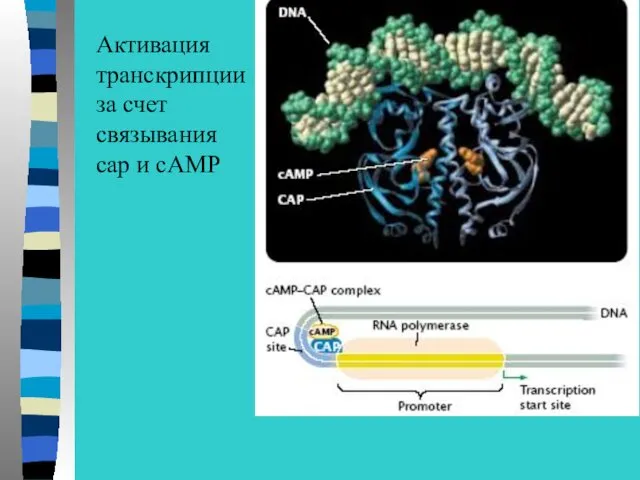
Активация
транскрипции
за счет
связывания
сap и сAMP
Активация
транскрипции
за счет
связывания
сap и сAMP

Регуляция транскрипции транспозонами (Т)
Поскольку Т несут в своем составе регуляторные сигналы
Регуляция транскрипции транспозонами (Т)
Поскольку Т несут в своем составе регуляторные сигналы

Т может изменить границы петель. Некоторые Т могут нести последовательности инсулятора,
Т может изменить границы петель. Некоторые Т могут нести последовательности инсулятора,

Эукариотические мРНК довольно стабильны (часы и сутки)До выхода в цитоплазму они
Эукариотические мРНК довольно стабильны (часы и сутки)До выхода в цитоплазму они

Посттранскрипционный уровень регуляции генной
активности
Транспорт мРНК из ядра
Процессинг мРНК
Альтернативный сплайсинг
Редактирование мРНК
Посттранскрипционный уровень регуляции генной
активности
Транспорт мРНК из ядра
Процессинг мРНК
Альтернативный сплайсинг
Редактирование мРНК
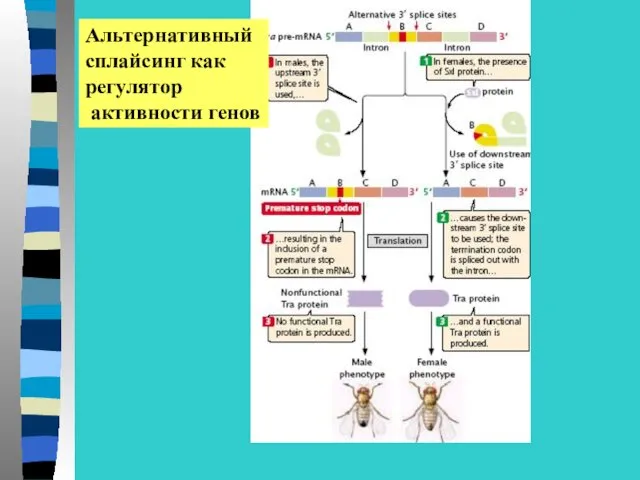
Альтернативный
сплайсинг как
регулятор
активности генов
Альтернативный
сплайсинг как
регулятор
активности генов

Редактирование РНК – термин предложен
Р. Бенни для феномена встраивания 4 У
Редактирование РНК – термин предложен
Р. Бенни для феномена встраивания 4 У

Биологические последствия редактирования РНК
Образование пригодной для трансляции РНК
И редактированная и нередактированная
Биологические последствия редактирования РНК
Образование пригодной для трансляции РНК
И редактированная и нередактированная

Регуляция генной активности на уровне трансляции
Инициация трансляции с альтернативных сайтов
Регуляция активности
Регуляция генной активности на уровне трансляции
Инициация трансляции с альтернативных сайтов
Регуляция активности

Регуляция синтеза ферритина если железа в среде мало, то соответствующая мРНК
Регуляция синтеза ферритина если железа в среде мало, то соответствующая мРНК

Дискриминация мРНК- инициирующие участки РНК имеют разное сродство к рибосомам и
Дискриминация мРНК- инициирующие участки РНК имеют разное сродство к рибосомам и

Трансляционная репрессия – белок репрессор связывается с участком инициации трансляции и
Трансляционная репрессия – белок репрессор связывается с участком инициации трансляции и

Маскирование мРНК – осуществляется белком связывающимся с 3’ НТО. Как связывание
Маскирование мРНК – осуществляется белком связывающимся с 3’ НТО. Как связывание

Рнк не выходит из ядра, пока не закончится ее процессинг.
У ВИЧ
Рнк не выходит из ядра, пока не закончится ее процессинг.
У ВИЧ

Посттрансляционный уровень регуляции
Фолдинг белка
Роль гликозилирования, фосфорилирование,
ацетилирование и др и др.
Посттрансляционный уровень регуляции
Фолдинг белка
Роль гликозилирования, фосфорилирование,
ацетилирование и др и др.

Механизмы сайленсинга
Гипотеза гистонового кода – ковалентные модификации ДНК и гистонов
гипоацетилирование, убиквитирование
Механизмы сайленсинга
Гипотеза гистонового кода – ковалентные модификации ДНК и гистонов
гипоацетилирование, убиквитирование
 Технология культивирования грибов вешанок и шампиньонов
Технология культивирования грибов вешанок и шампиньонов Нуклеиновые кислоты. История открытия
Нуклеиновые кислоты. История открытия Глаз, как орган зрения и оптическая система
Глаз, как орган зрения и оптическая система Презентация на тему Покрытосеменные (цветковые) растения
Презентация на тему Покрытосеменные (цветковые) растения Подготовка к экзамену. Биология. (11 класс)
Подготовка к экзамену. Биология. (11 класс) Царство Бактерии
Царство Бактерии Насекомые
Насекомые «Животные открытых пространств» Работа ученика 7 а класса, МОУ «СОШ №28» Филиппова Андрея
«Животные открытых пространств» Работа ученика 7 а класса, МОУ «СОШ №28» Филиппова Андрея Филогенез. Основные принципы эволюции систем органов организмов. Общие закономерности развития систем органов
Филогенез. Основные принципы эволюции систем органов организмов. Общие закономерности развития систем органов Основные виды работ по уходу за древесными растениями осенью
Основные виды работ по уходу за древесными растениями осенью Строение и деятельность внутренних органов Земноводных
Строение и деятельность внутренних органов Земноводных СИСТЕМАТИКА ВЫСШИХ РАСТЕНИЙ
СИСТЕМАТИКА ВЫСШИХ РАСТЕНИЙ  Пищеварительная система человека. Пищеварение в ротовой полости
Пищеварительная система человека. Пищеварение в ротовой полости Современные биобанки
Современные биобанки Рыбоводство. Карась
Рыбоводство. Карась Эволюция жизни и возникновение приматов
Эволюция жизни и возникновение приматов День птиц
День птиц Презентация на тему "Перша медична допомога при радіаційних ураженнях" - скачать бесплатно презентации по Биологии
Презентация на тему "Перша медична допомога при радіаційних ураженнях" - скачать бесплатно презентации по Биологии Ткани. Типы тканей и их свойства.
Ткани. Типы тканей и их свойства. Пора года - осень
Пора года - осень Наземно-повітряне середовище Температурний фактор
Наземно-повітряне середовище Температурний фактор  Презентация на тему "Кишковопорожнинні та їх значення" - скачать бесплатно презентации по Биологии
Презентация на тему "Кишковопорожнинні та їх значення" - скачать бесплатно презентации по Биологии Дыхательная система
Дыхательная система Биосинтез белка
Биосинтез белка Многообразие водорослей
Многообразие водорослей Семейство Дельфиновые, Delphinidae
Семейство Дельфиновые, Delphinidae Презентация на тему "Основы селекции растений, животных, микроорганизмов" - скачать презентации по Биологии
Презентация на тему "Основы селекции растений, животных, микроорганизмов" - скачать презентации по Биологии Стебель, его строение и значение
Стебель, его строение и значение