- Зат алмасуға кіріспе. Жасуша биоэнергетикасы және катаболизмнің ортақ жолдары

Содержание
- 2. Негізгі сұрақтары: Пирожүзім қышқылының Қышқылды декарбоксилдену. Үшкарбонды қышқыл циклі Қайықша механизмдер Макроэргиялық байланыс.Ұлпаның тыныс алуы туралы
- 3. Спецификалық және жалпы катаболизм жолдары . Катаболитті процесті екі бөлімге бөлуге болады: - катаболизмнің спецификалық жолы,
- 5. Пирожүзім қышқылының қышқылды декарбоксилденуі. Пируваттың қышқылды декорбоксилденуі матрикс жағындағы митохондрияның ішкі мембранасында мультиэнзим пируват-дегидрогеназды комплекс арқылы
- 6. Пирожүзім қышқылы (пируват) және ацетил-КоА - катаболизмнің жалпы жолы қосылған кезде көмірқышқыл газ бен суға дейін
- 7. Пируваттың қышқылдануы
- 11. Үшкарбонды қышқыл циклі
- 12. Үшкарбон қышқыл циклі
- 13. Кребс циклі
- 14. Энергияның шығуы Сент-Дьёрди – Кребс үшкарбон қышқыл циклінде субстратты сәйкес дегидрогеназамен дегидрлейтін 4 реакция орны бар:
- 18. Транспорттың қайықша механизмдері Топтық транслокацияның өзіндік нұсқасы болып протондардың (Н+) митохондриялы мембрана арқылы көшу механизмі саналады.
- 20. Малатный челнок
- 21. Тотығудың негізгі түрлері Органикалық байланыстың (субстрат)катоболизм реакциясы биосистемада бос энергияның босатылуымен байланысты, негізінде қышқылды-қалпына келгіш реакция
- 22. Маркоэргтер Жануар организмінің энергетикалық субсттратына көмірсу,липид,аминоқышқылдар жатады. көмірсудың калориясы 4,1 ккал/г, липидтікі - 9,3 ккал/г және
- 23. Клеткада энергияның трансформация механизмі Көмірсу катоболизмі кезінде бос энергияның,липид және аминқышқылдың босап шығуы, аэробты және анаэробты
- 25. Биологиялық тотығу механизмі
- 26. Бұл процесстің негізгі қызметі организмді энергиямен қамтамасыз ету. (АҮФ түрінде) Субстраттың молекулалы кислородқа ауысу процессіне мыналар
- 27. Анаэробты дегидрогеназалар Бұл ферменттер сутегі атомдарының(протондар және электрондардың) оттектебасқа кез-келген акцепторға ауысуын катализдейді Представители: альфа-кетоглутарат дегидрогеназа
- 28. N o H OH OH H CH2 O P OH O O P OH O O
- 29. АЭРОБТЫ ДЕГИДРОГЕНАЗАЛАР Бұл ферменттер сутегі атомдарының(протондар және электрондардың) кез-келген акцепторға,сонымен қоса оттекке де ауысуын катализдейді Представители:
- 30. С С С С С С С С С С С С H H H3 H3
- 31. Кофермент Q Тыныс алу тізбегінің компоненті-убихинон,бензохиноннан өндіріледі және ФАДН2 –дегидрогеназ коферменті болып табылады, протондар мен электрондардың
- 32. Электрондардың убихиноннан кислородқа тасымалдануын цитохромдар жүйесі жүзеге асырады : b, c1, c, a- a3. Олардың барлығы
- 33. Цитохромның қышқылдану-қалпына келу реакциясына әсер ету механизмі Fe3+ + e- Fe2+ Fe2+ - e - Fe3+
- 34. Тыныс алу тізбегінің құрылымы Электрондар мен протондардың митохондриальды тыныс алу тізбегіне тасымалдануы қышқылдану-қайта қалпына келу потенциалы
- 35. ТЫНЫС АЛУ ТІЗБЕГІ SH2 НАД ФАД КоQ b c1 c a a3 1/2O2 O + H
- 36. ТЫНЫС АЛУ ТІЗБЕГІ ЖӘНЕ РЕДОКС ПОТЕНЦИАЛ
- 37. ФОСФОРИЛДЕНУ ЖОЛДАРЫ Фосфорилденудің бірінші жолы НАД және ФАД ферменттерінің арасында жүзеге асады,және олардың потенциалдарының мәні 0,246
- 38. Ко-А-производные Май қышқылдары Изоцитрат Малат Пируват Глутамат НАД ФП b КоQ АТФ АДФ - Аминобарбитал, ротенон
- 39. Тотығып фосфорлану Бір субстрат молекуласының қышқылдануынан митохондрияға бір атом оттегі енгізіліп, бір молекуладан үш молекулалы фосфорлы
- 41. П. Митчелдің гипотеза ұсынысына орай, фосфорилденетін АДФ қозғалыс күші әр түрлі редокс-потенциалдар энергиясында қолданылады және ол
- 43. Скачать презентацию

Негізгі сұрақтары:
Пирожүзім қышқылының Қышқылды декарбоксилдену.
Үшкарбонды қышқыл циклі
Қайықша механизмдер
Макроэргиялық байланыс.Ұлпаның
Негізгі сұрақтары:
Пирожүзім қышқылының Қышқылды декарбоксилдену.
Үшкарбонды қышқыл циклі
Қайықша механизмдер
Макроэргиялық байланыс.Ұлпаның

Спецификалық және жалпы катаболизм жолдары
. Катаболитті процесті екі бөлімге бөлуге болады:
Спецификалық және жалпы катаболизм жолдары
. Катаболитті процесті екі бөлімге бөлуге болады:


Пирожүзім қышқылының қышқылды декарбоксилденуі.
Пируваттың қышқылды декорбоксилденуі матрикс жағындағы митохондрияның ішкі
Пирожүзім қышқылының қышқылды декарбоксилденуі.
Пируваттың қышқылды декорбоксилденуі матрикс жағындағы митохондрияның ішкі

Пирожүзім қышқылы (пируват) және ацетил-КоА - катаболизмнің жалпы жолы қосылған кезде
Пирожүзім қышқылы (пируват) және ацетил-КоА - катаболизмнің жалпы жолы қосылған кезде
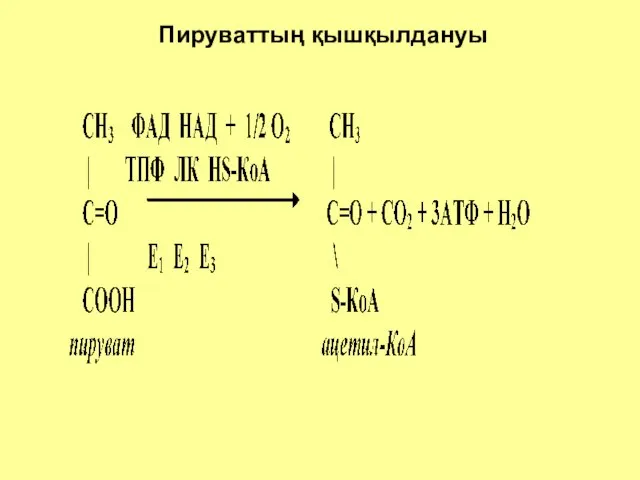
Пируваттың қышқылдануы
Пируваттың қышқылдануы




Үшкарбонды қышқыл циклі
Үшкарбонды қышқыл циклі

Үшкарбон қышқыл циклі
Үшкарбон қышқыл циклі

Кребс циклі
Кребс циклі

Энергияның шығуы
Сент-Дьёрди – Кребс үшкарбон қышқыл циклінде субстратты сәйкес дегидрогеназамен дегидрлейтін
Энергияның шығуы
Сент-Дьёрди – Кребс үшкарбон қышқыл циклінде субстратты сәйкес дегидрогеназамен дегидрлейтін



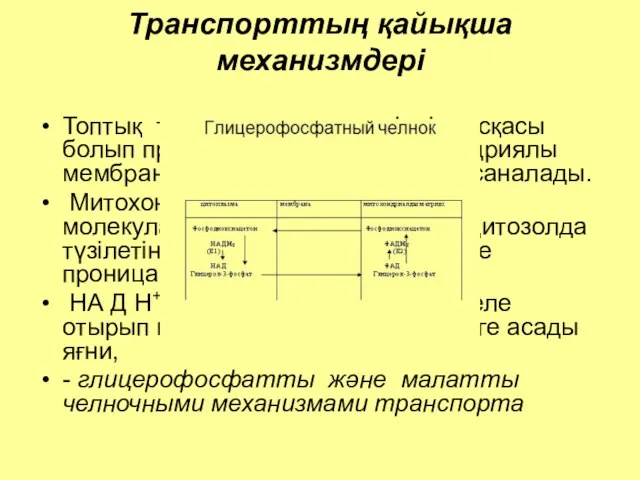
Транспорттың қайықша механизмдері
Топтық транслокацияның өзіндік нұсқасы болып протондардың (Н+) митохондриялы мембрана
Транспорттың қайықша механизмдері
Топтық транслокацияның өзіндік нұсқасы болып протондардың (Н+) митохондриялы мембрана
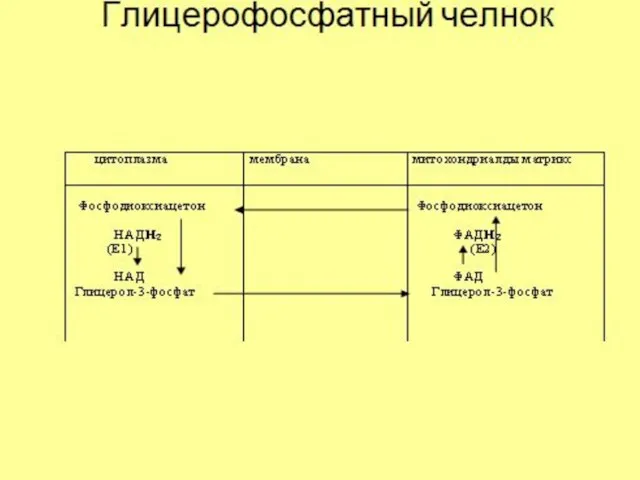

Малатный челнок
Малатный челнок

Тотығудың негізгі түрлері
Органикалық байланыстың (субстрат)катоболизм реакциясы биосистемада бос энергияның босатылуымен
Тотығудың негізгі түрлері
Органикалық байланыстың (субстрат)катоболизм реакциясы биосистемада бос энергияның босатылуымен

Маркоэргтер
Жануар организмінің энергетикалық субсттратына көмірсу,липид,аминоқышқылдар жатады. көмірсудың калориясы 4,1 ккал/г, липидтікі
Маркоэргтер
Жануар организмінің энергетикалық субсттратына көмірсу,липид,аминоқышқылдар жатады. көмірсудың калориясы 4,1 ккал/г, липидтікі

Клеткада энергияның трансформация механизмі
Көмірсу катоболизмі кезінде бос энергияның,липид және
Клеткада энергияның трансформация механизмі
Көмірсу катоболизмі кезінде бос энергияның,липид және

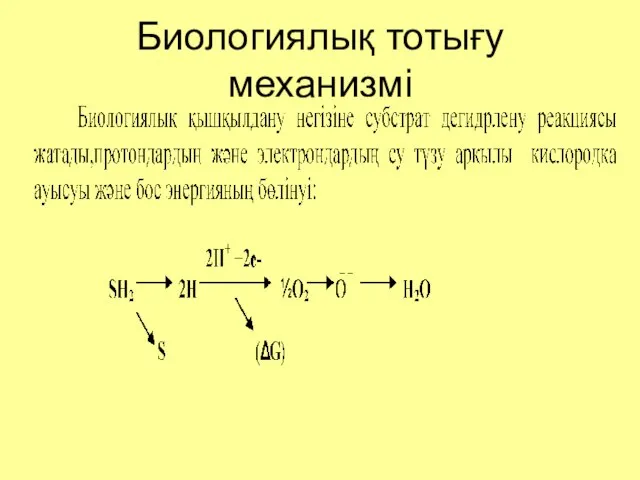
Биологиялық тотығу механизмі
Биологиялық тотығу механизмі

Бұл процесстің негізгі қызметі организмді энергиямен қамтамасыз ету. (АҮФ түрінде)
Субстраттың
Бұл процесстің негізгі қызметі организмді энергиямен қамтамасыз ету. (АҮФ түрінде)
Субстраттың

Анаэробты дегидрогеназалар
Бұл ферменттер сутегі атомдарының(протондар және электрондардың) оттектебасқа кез-келген акцепторға ауысуын
Анаэробты дегидрогеназалар
Бұл ферменттер сутегі атомдарының(протондар және электрондардың) оттектебасқа кез-келген акцепторға ауысуын
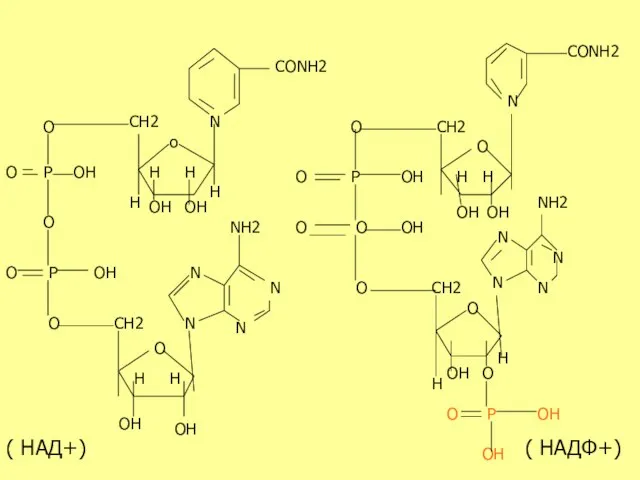
N
o
H
OH
OH
H
CH2
O
P
OH
O
O
P
OH
O
O
CH2
O
OH
OH
N
N
N
N
NH2
H
H
H
H
CONH2
( НАД+)
O
P
O
OH
O
O
OH
O
CH2
H
CH2
CONH2
N
O
NH2
N
N
N
N
H
OH
O
OH
OH
H
H
O
P
OH
O
OH
( НАДФ+)
N
o
H
OH
OH
H
CH2
O
P
OH
O
O
P
OH
O
O
CH2
O
OH
OH
N
N
N
N
NH2
H
H
H
H
CONH2
( НАД+)
O
P
O
OH
O
O
OH
O
CH2
H
CH2
CONH2
N
O
NH2
N
N
N
N
H
OH
O
OH
OH
H
H
O
P
OH
O
OH
( НАДФ+)

АЭРОБТЫ ДЕГИДРОГЕНАЗАЛАР
Бұл ферменттер сутегі атомдарының(протондар және электрондардың) кез-келген акцепторға,сонымен қоса оттекке
АЭРОБТЫ ДЕГИДРОГЕНАЗАЛАР
Бұл ферменттер сутегі атомдарының(протондар және электрондардың) кез-келген акцепторға,сонымен қоса оттекке
С
С
С
С
С
С
С
С
С
С
С
С
H
H
H3
H3
N
N
O
NH
N
CH2
H C OH
H C OH
H C OH
CH2
O
O P
С
С
С
С
С
С
С
С
С
С
С
С
H
H
H3
H3
N
N
O
NH
N
CH2
H C OH
H C OH
H C OH
CH2
O
O P

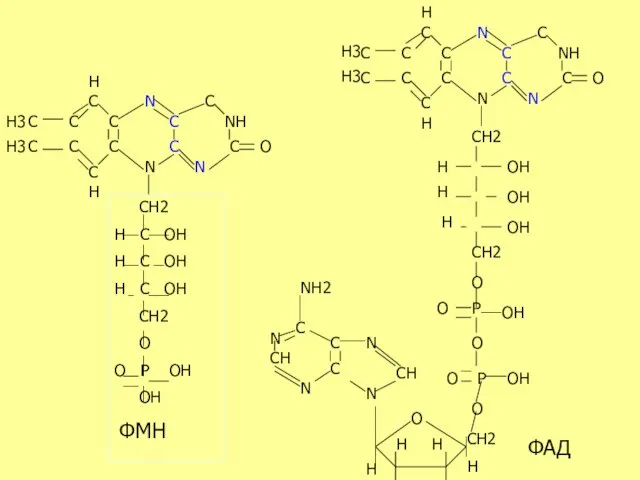
Кофермент Q
Тыныс алу тізбегінің компоненті-убихинон,бензохиноннан өндіріледі және ФАДН2 –дегидрогеназ коферменті
Кофермент Q
Тыныс алу тізбегінің компоненті-убихинон,бензохиноннан өндіріледі және ФАДН2 –дегидрогеназ коферменті

Электрондардың убихиноннан кислородқа тасымалдануын цитохромдар жүйесі жүзеге асырады :
b, c1,
Электрондардың убихиноннан кислородқа тасымалдануын цитохромдар жүйесі жүзеге асырады :
b, c1,

Цитохромның қышқылдану-қалпына келу реакциясына әсер ету механизмі
Fe3+ + e- Fe2+ Fe2+
Цитохромның қышқылдану-қалпына келу реакциясына әсер ету механизмі
Fe3+ + e- Fe2+ Fe2+

Тыныс алу тізбегінің құрылымы
Электрондар мен протондардың митохондриальды тыныс алу тізбегіне тасымалдануы
Тыныс алу тізбегінің құрылымы
Электрондар мен протондардың митохондриальды тыныс алу тізбегіне тасымалдануы
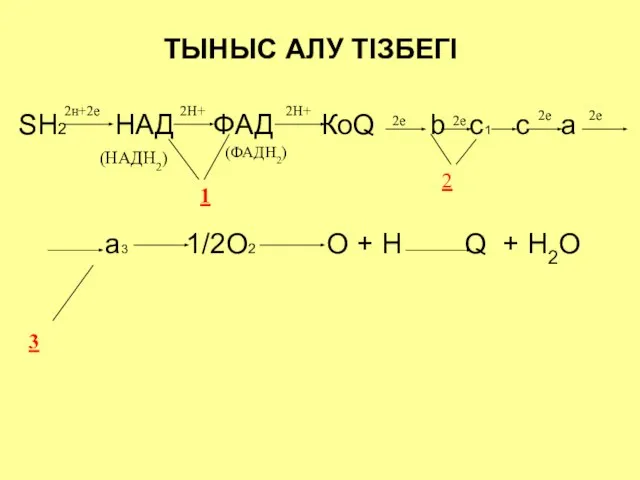
ТЫНЫС АЛУ ТІЗБЕГІ
SH2 НАД ФАД КоQ b c1 c a
a3
ТЫНЫС АЛУ ТІЗБЕГІ
SH2 НАД ФАД КоQ b c1 c a
a3

ТЫНЫС АЛУ ТІЗБЕГІ ЖӘНЕ РЕДОКС ПОТЕНЦИАЛ
ТЫНЫС АЛУ ТІЗБЕГІ ЖӘНЕ РЕДОКС ПОТЕНЦИАЛ

ФОСФОРИЛДЕНУ ЖОЛДАРЫ
Фосфорилденудің бірінші жолы НАД және ФАД ферменттерінің арасында жүзеге асады,және
ФОСФОРИЛДЕНУ ЖОЛДАРЫ
Фосфорилденудің бірінші жолы НАД және ФАД ферменттерінің арасында жүзеге асады,және

Ко-А-производные
Май қышқылдары
Изоцитрат
Малат
Пируват
Глутамат
НАД
ФП
b
КоQ
АТФ
АДФ
-
Аминобарбитал, ротенон
ФП2
ФП3
ФП4
Глицерол-3-фосфат
сукцинат
АДФ
АТФ
C1
c
a3
a
Антимицин
-
Аскорбат
О2
-
Цианид
АДФ
АТФ
1
2
3
Локализация трех пунктов фосфорилирования в цепи (1,2,3)
Ко-А-производные
Май қышқылдары
Изоцитрат
Малат
Пируват
Глутамат
НАД
ФП
b
КоQ
АТФ
АДФ
-
Аминобарбитал, ротенон
ФП2
ФП3
ФП4
Глицерол-3-фосфат
сукцинат
АДФ
АТФ
C1
c
a3
a
Антимицин
-
Аскорбат
О2
-
Цианид
АДФ
АТФ
1
2
3
Локализация трех пунктов фосфорилирования в цепи (1,2,3)

Тотығып фосфорлану
Бір субстрат молекуласының қышқылдануынан митохондрияға бір атом оттегі енгізіліп, бір
Тотығып фосфорлану
Бір субстрат молекуласының қышқылдануынан митохондрияға бір атом оттегі енгізіліп, бір

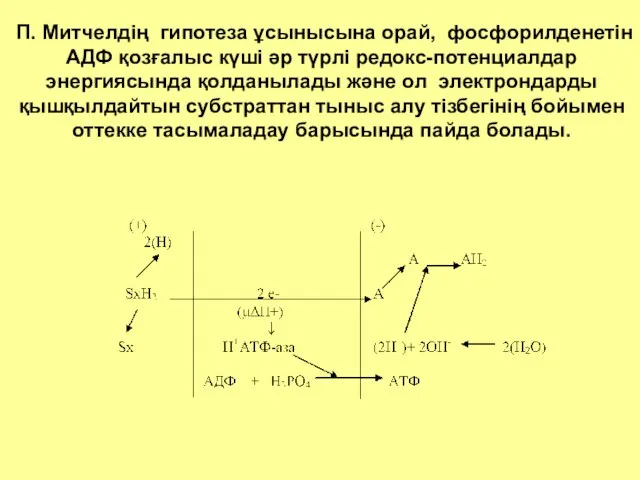
П. Митчелдің гипотеза ұсынысына орай, фосфорилденетін АДФ қозғалыс күші әр
П. Митчелдің гипотеза ұсынысына орай, фосфорилденетін АДФ қозғалыс күші әр
 Следы жизнедеятельности животных
Следы жизнедеятельности животных Характеристика классов однодольные и двудольные
Характеристика классов однодольные и двудольные Сердечно-сосудистая система
Сердечно-сосудистая система Биология с основами экологии
Биология с основами экологии Многообразие биогеоценозов (экосистем)
Многообразие биогеоценозов (экосистем) Генетика на страже здоровья человека
Генетика на страже здоровья человека Биосинтез белка
Биосинтез белка Обеспечение клеток энергией
Обеспечение клеток энергией Обмен веществ и энергии
Обмен веществ и энергии Предлагаю вашему вниманию результаты работы лаборатории «Млекопитающие, ведущие полуводный образ жизни» Во-первых, нужно было пр
Предлагаю вашему вниманию результаты работы лаборатории «Млекопитающие, ведущие полуводный образ жизни» Во-первых, нужно было пр Влаголюбивые и засухоустойчивые комнатные растения
Влаголюбивые и засухоустойчивые комнатные растения Презентация на тему "Виноград" - скачать презентации по Биологии
Презентация на тему "Виноград" - скачать презентации по Биологии Общая физиология нервной системы
Общая физиология нервной системы Хищники
Хищники Тип Моллюски
Тип Моллюски Презентация на тему Кроссворд «Происхождение человека»
Презентация на тему Кроссворд «Происхождение человека»  Ткани растений
Ткани растений Жизнь организмов на разных материках
Жизнь организмов на разных материках Подцарство Одноклеточные
Подцарство Одноклеточные Трансгенные растения и животные
Трансгенные растения и животные Физиология микроорганизмов
Физиология микроорганизмов Фитодизайн Старогородской школы
Фитодизайн Старогородской школы Радиация – проблемы и перспективы…
Радиация – проблемы и перспективы… Виды съедобных грибов. Существует множество съедобных грибов, регулярно выращиваемых и собираемых во всём мире. Грибы обладаю
Виды съедобных грибов. Существует множество съедобных грибов, регулярно выращиваемых и собираемых во всём мире. Грибы обладаю Водоплавающее млекопитающее утконос
Водоплавающее млекопитающее утконос растит ткеани355085
растит ткеани355085 Многообразие паукообразных и их значение
Многообразие паукообразных и их значение Бедренный треугольник
Бедренный треугольник