-
Биотехнология и генетическая инженерия

Содержание
- 2. Биотехнология – совокупность методов естественных и инженерных наук, использующих свойства биологических систем различного уровня организации в
- 3. Генетической инженерией называют прикладную молекулярную и клеточную генетику, разрабатывающую приемы экспериментального вмешательства, позволяющие по заранее намеченному
- 4. Разделы биотехнологии: Генетическая инженерия – технологии основаны на получении гибридных молекул ДНК и введении их в
- 5. К генетической инженерии принято относить следующие операции: 1. синтез генов вне организма; 2. выделение из клеток
- 7. Ферменты, используемые в генной инженерии. Рестриктазы (рестрикционные нуклеазы) – ферменты, способные узнавать специфические последовательности ДНК (4-6
- 8. Расщепление фрагмента ДНК рестрицирующей эндонуклеазой типа II EcoR1 с образованием липких концов
- 9. Схема действия рестриктазы EcoR 1
- 10. Расщепление фрагмента ДНК рестриктазой типа II Hind II с образованием тупых концов
- 11. Нуклеотидные последовательности, распознаваемые некоторыми ферментами рестрикции
- 12. Отжиг комплементарных липких концов фрагментов, образующихся при расщеплении двух разных образцов ДНК рестрицирующей эндонуклеазой BamH1
- 13. Синтез и выделение фрагментов ДНК
- 14. Схема ферментативного синтеза гена с помощью обратной транскрипции
- 15. Структура химически синтезированного Хораной функционально активного отрезка ДНК кишечной палочки Цифры – нумерация нуклеотидов: промотор от
- 16. Синтез кДНК Синтез кДНК. К препарату очищенной мРНК добавляют праймер oligo(dT). Для синтеза ДНК на РНК-матрице
- 17. Синтез кДНК
- 18. Выделение фрагментов ДНК, содержащих нужный ген
- 19. Векторы в генной инженерии. Для осуществления трансгеноза (введения чужеродных генов в клетку реципиента) используются вектоы (переносчики).
- 20. Генетическая карта плазмидного вектора pBR322
- 21. Генетическая карта плазмидного вектора pUC19 Плазмида состоит из 2686 пар нуклеотидов и содержит уникальные сайты узнавания
- 22. Встраивание чужеродной ДНК в плазмидный вектор Плазмидную ДНК, обработанную рестриктазой и щелочной фосфатазой, смешивают с рестрицированной
- 23. Ферментативный синтез гена и встраивание его в векторную плазмиду
- 24. Схема получения и клонирования рекомбинантной молекулы
- 25. Интеграция чужеродной ДНК в хромосому хозяина Процесс интеграции состоит в следущем: Идентификация подходящего сайта интеграции, т.
- 26. Два способа интеграции клонированного в плазмиде гена в хромосому в результате двойного (А) и одиночного (Б)
- 27. Встраивание чужеродного гена в заранее выбранный сайт в хромосоме B.subtilis 1 этап – в хромосомную ДНК
- 28. «Нокаут» гена с помощью направленной гомологичной рекомбинации. Вектор несет селективный маркерный ген (smg) и фланкирующие его
- 29. Векторы для клонирования крупных фрагментов ДНК Литический путь развития λ. А. При репликации кольцевой ДНК бактериофага
- 30. Клонирующая система на основе бактериофага λ. Фаговая ДНК имеет два BamH1-сайта, фланкирующих её I/E-сегмент. Клонируемую ДНК
- 31. Клонирующая система на основе бактериофага λ
- 32. Лигирование липких и тупых концов в молекуле ДНК фагом Т4 ДНК фага Т4 образует фосфодиэфирные связи
- 33. Клонирование с помощью космидного вектора Космида имеет точку начала репликации (ori), обеспечивающую её существование в E.coli
- 34. Клонирование с помощью космидного вектора
- 35. Перенос генов с помощью искусственных дрожжевых хромосом
- 36. Идентификация клеток, несущих трансген
- 39. Методы получения трансгенных животных: 1. Метод микроиньекции 2. Вирусный метод 3. Эмбриональный метод
- 40. Метод микроиньекции У самок вызывают гиперовуляцию после чего проводят спаривание с самцами. Из самок-доноров выделяют яйцеклетки.В
- 41. Получение линий трансгенных мышей методом микроинъекций. Яйцеклетки выделяют из самок-доноров, у которых была индуцирована гиперовуляция и
- 42. Метод микроинъекций ДНК Увеличение числа яйцеклеток, в которых будет инъецирована чужеродная ДНК, путем стимуляции гиперовуляции у
- 43. Вирусный метод Эмбрион, находящийся на стадии 8 клеток, инфицируют рекомбинантным ретровирусом, несущим трансген. Самки, которым был
- 44. Получение трансгенных мышей с использованием ретровирусных векторов Получение линии трансгенных мышей с использованием ретровирусных векторов. Эмбрион,
- 45. Эмбриональный метод Эмбриональные стволовые клетки получают из внутренней клеточной массы бластоцисты мыши. Их трансфицируют вектором, несущим
- 46. ES-клетки получают из внутренней клеточной массы бластоцисты мыши. Их трансфицируют вектором, несущим трансген, культивируют и идентифицируют
- 47. Использование модифицированных эмбриональных стволовых клеток Клетки, выделенные из мышиных эмбрионов на стадии бластоцисты, могут пролиферировать в
- 48. Трансгенные животные
- 49. Стратегия по введению чужеродных генов в клетки млекопитающих состоит в следующем: Клонированный ген вводят в ядро
- 50. Клонирование овцы методом переноса ядра Клонирование овцы методом переноса ядра. Ядро яйцеклетки удаляют с помощью микропипетки.
- 51. Клонирование овечки Долли
- 52. Трансгенный крупный рогатый скот Если предполагается использовать молочную железу в качестве «биореактора», то наиболее предпочтительным животным
- 53. Для создания трансгенных коров использовали модифицированную схему трансгеноза мышей методом микроинъекций ДНК. Процедура включала следующие основные
- 54. Получение трансгенных коров
- 55. Трансгенные овцы, козы и свиньи
- 56. Трансгенные птицы Микроинъекция ДНК в оплодотворенные яйцеклетки птиц с целью получения трансгенных линий - непростая процедура.
- 57. Получение трансгенных цыплят трансфекцией изолированных клеток бластодермы. Выделенные клетки трансфицируют трансгеном с помощью липосом и вводят
- 58. Трансгенные рыбы По мере истощения природных рыбных запасов все большую роль будет приобретать разведение рыбы в
- 59. Трансгенные мыши: методология Введение чужеродной ДНК мышам можно осуществить разными методами: С помощью ретровирусных векторов, инфицирующих
- 60. Трансгенные мыши: применение как модельного обьекта изучения болезни Альцгеймера. Болезнь Альцгеймера — это дегенеративный процесс, приводящий
- 61. Схематическое изображение нейрона коры головного мозга человека с указанием некоторых гистологических особенностей, характерных для болезни Альцгеймера.
- 62. Основным компонентом сенильных и амилоидных бляшек является белок Ар (амилоид р\ р-белок, р-амилоидный белок, (3/А4) мол.
- 63. Моделирование развития болезни Альцгеймера
- 64. Муковисцидоз — распространенная генетическая болезнь, поражающая в странах Европы одного из 2500 новорожденных. Первичный эффект дефектного
- 66. Гибридизация соматических клеток
- 67. Методика изоляции соматических гибридов (по Шапиро) ТК – фермент тимидинкиназа ГГФРТ – фермент гипоксантин-гуанин-фосфорибозилтрансфераза На селективной
- 68. Некоторые сочетания геномов, полученные слиянием соматических клеток
- 70. Скачать презентацию

Биотехнология – совокупность методов естественных и инженерных наук, использующих свойства
Биотехнология – совокупность методов естественных и инженерных наук, использующих свойства

Генетической инженерией называют прикладную молекулярную и клеточную генетику, разрабатывающую приемы
Генетической инженерией называют прикладную молекулярную и клеточную генетику, разрабатывающую приемы

Разделы биотехнологии:
Генетическая инженерия – технологии основаны на получении гибридных молекул ДНК
Разделы биотехнологии:
Генетическая инженерия – технологии основаны на получении гибридных молекул ДНК

К генетической инженерии принято относить следующие операции:
1. синтез генов вне
К генетической инженерии принято относить следующие операции:
1. синтез генов вне

Ферменты, используемые в генной инженерии.
Рестриктазы (рестрикционные нуклеазы) – ферменты, способные
Ферменты, используемые в генной инженерии.
Рестриктазы (рестрикционные нуклеазы) – ферменты, способные
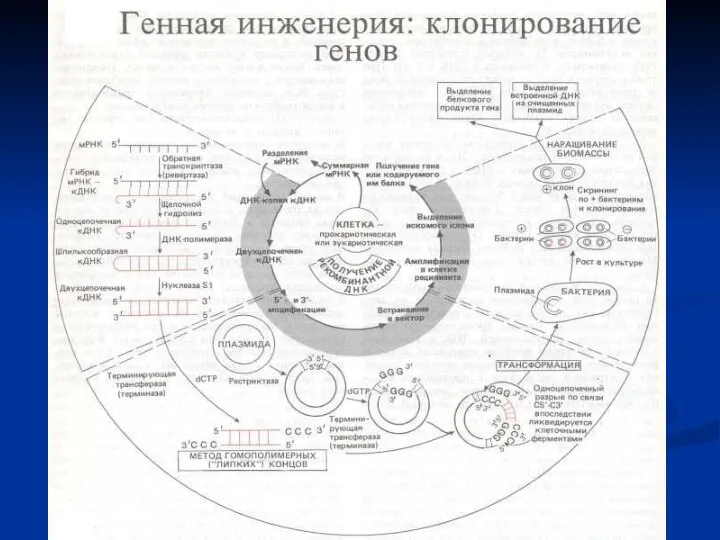
Расщепление фрагмента ДНК рестрицирующей эндонуклеазой типа II EcoR1 с образованием липких
Расщепление фрагмента ДНК рестрицирующей эндонуклеазой типа II EcoR1 с образованием липких

Схема действия рестриктазы EcoR 1
Схема действия рестриктазы EcoR 1
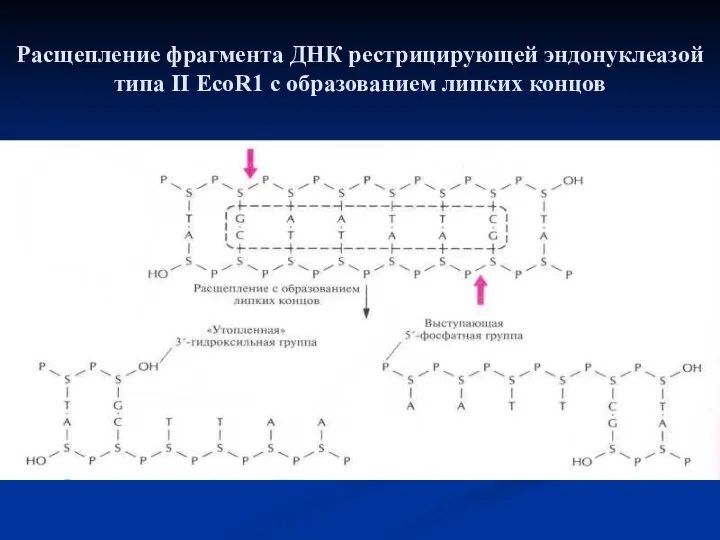
Расщепление фрагмента ДНК рестриктазой типа II Hind II с образованием тупых
Расщепление фрагмента ДНК рестриктазой типа II Hind II с образованием тупых

Нуклеотидные последовательности, распознаваемые некоторыми ферментами рестрикции
Нуклеотидные последовательности, распознаваемые некоторыми ферментами рестрикции

Отжиг комплементарных липких концов фрагментов, образующихся при расщеплении двух разных образцов
Отжиг комплементарных липких концов фрагментов, образующихся при расщеплении двух разных образцов

Синтез и выделение фрагментов ДНК
Синтез и выделение фрагментов ДНК
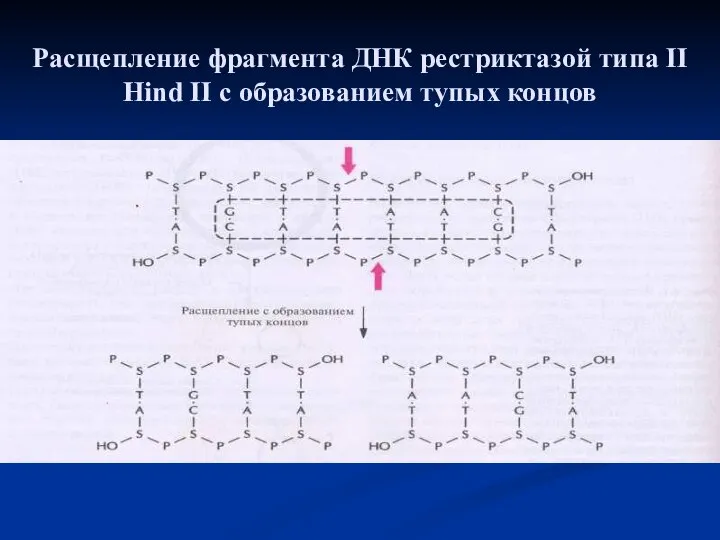
Схема ферментативного синтеза гена с помощью обратной транскрипции
Схема ферментативного синтеза гена с помощью обратной транскрипции

Структура химически синтезированного Хораной функционально активного отрезка ДНК кишечной палочки
Цифры –
Структура химически синтезированного Хораной функционально активного отрезка ДНК кишечной палочки
Цифры –

Синтез кДНК
Синтез кДНК. К препарату очищенной мРНК добавляют праймер oligo(dT). Для
Синтез кДНК
Синтез кДНК. К препарату очищенной мРНК добавляют праймер oligo(dT). Для
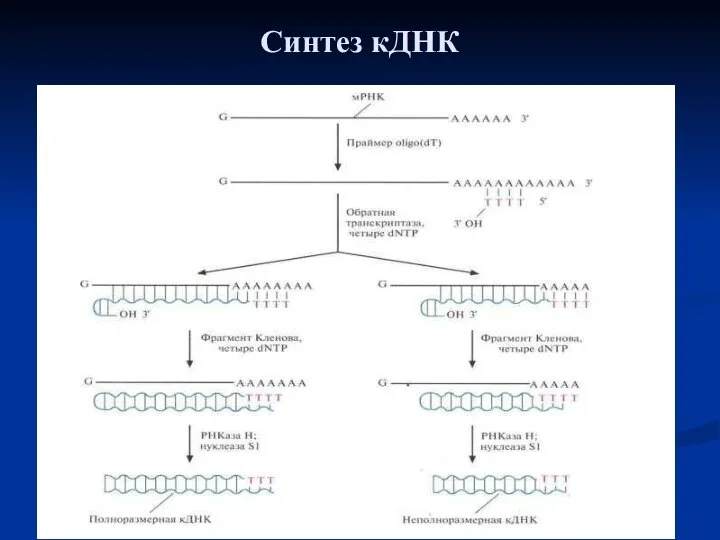
Синтез кДНК
Синтез кДНК
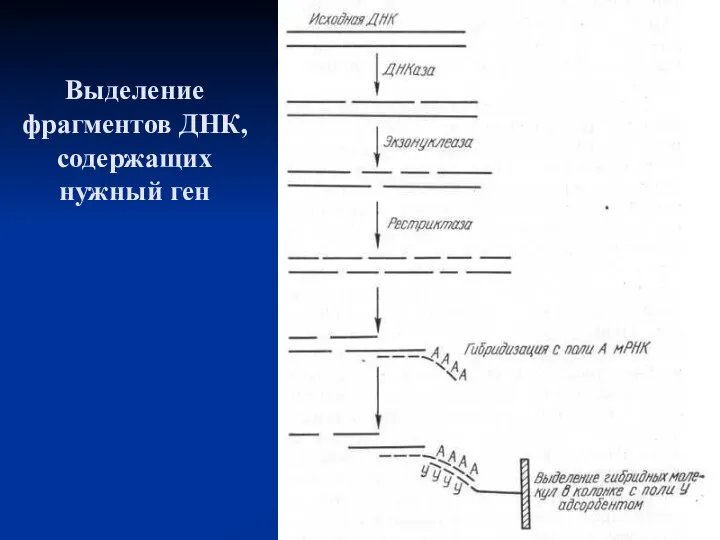
Выделение фрагментов ДНК, содержащих нужный ген
Выделение фрагментов ДНК, содержащих нужный ген

Векторы в генной инженерии.
Для осуществления трансгеноза (введения чужеродных генов в
Векторы в генной инженерии.
Для осуществления трансгеноза (введения чужеродных генов в

Генетическая карта плазмидного вектора pBR322
Генетическая карта плазмидного вектора pBR322

Генетическая карта плазмидного вектора pUC19
Плазмида состоит из 2686 пар нуклеотидов и
Генетическая карта плазмидного вектора pUC19
Плазмида состоит из 2686 пар нуклеотидов и
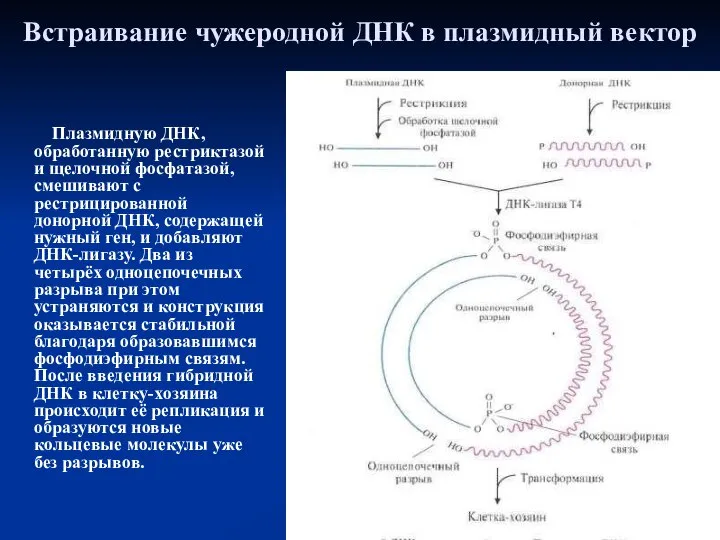
Встраивание чужеродной ДНК в плазмидный вектор
Плазмидную ДНК, обработанную рестриктазой и
Встраивание чужеродной ДНК в плазмидный вектор
Плазмидную ДНК, обработанную рестриктазой и
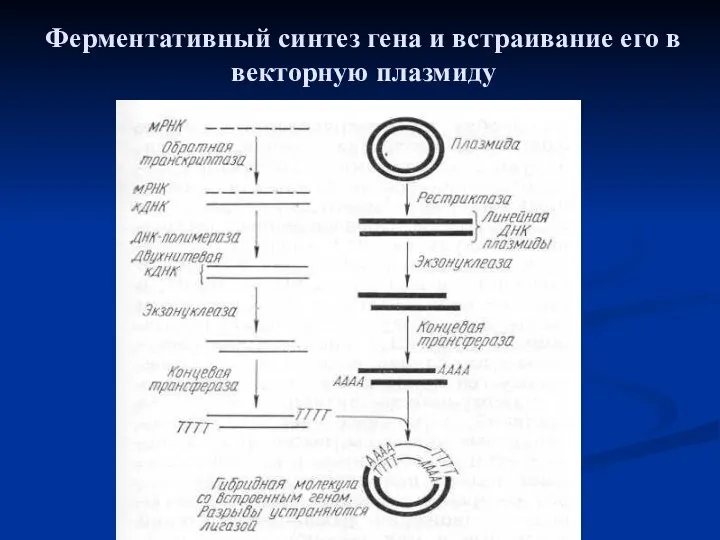
Ферментативный синтез гена и встраивание его в векторную плазмиду
Ферментативный синтез гена и встраивание его в векторную плазмиду
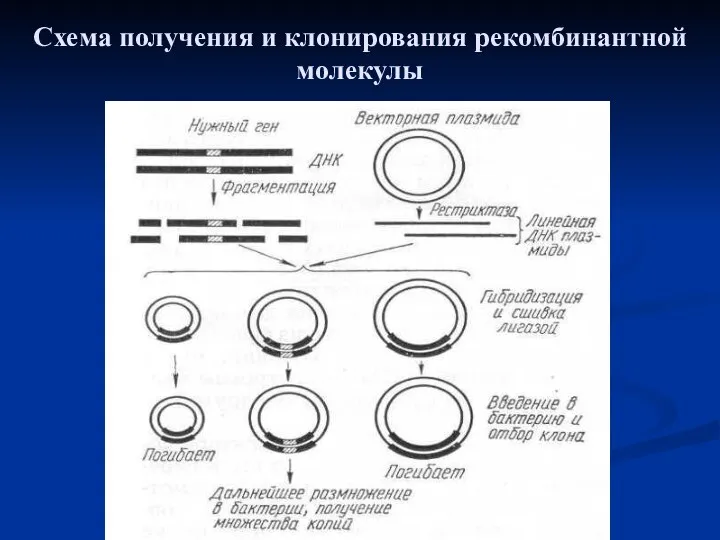
Схема получения и клонирования рекомбинантной молекулы
Схема получения и клонирования рекомбинантной молекулы

Интеграция чужеродной ДНК в хромосому хозяина
Процесс интеграции состоит в следущем:
Идентификация подходящего
Интеграция чужеродной ДНК в хромосому хозяина
Процесс интеграции состоит в следущем:
Идентификация подходящего
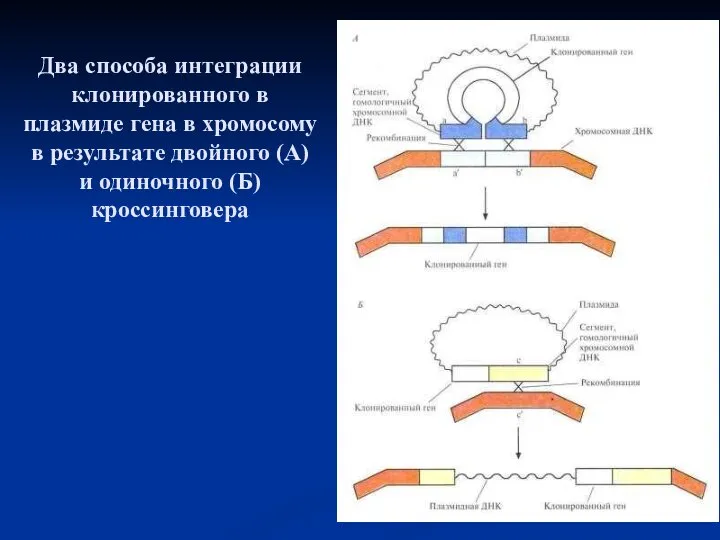
Два способа интеграции клонированного в плазмиде гена в хромосому в результате
Два способа интеграции клонированного в плазмиде гена в хромосому в результате
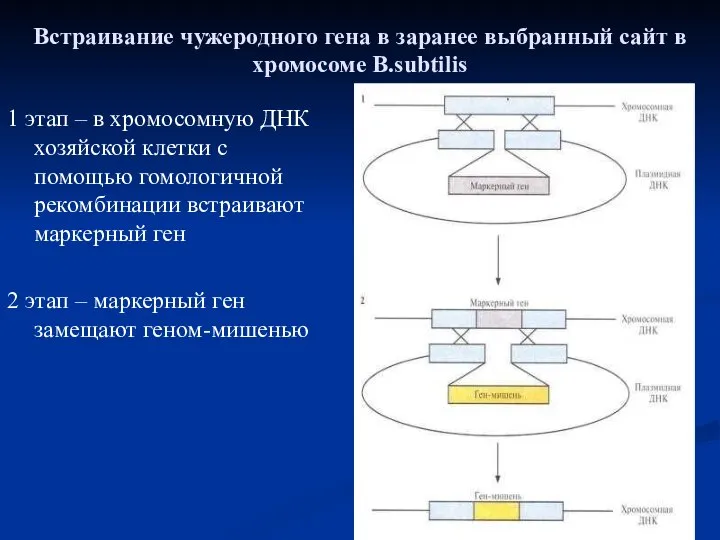
Встраивание чужеродного гена в заранее выбранный сайт в хромосоме B.subtilis
1 этап
Встраивание чужеродного гена в заранее выбранный сайт в хромосоме B.subtilis
1 этап
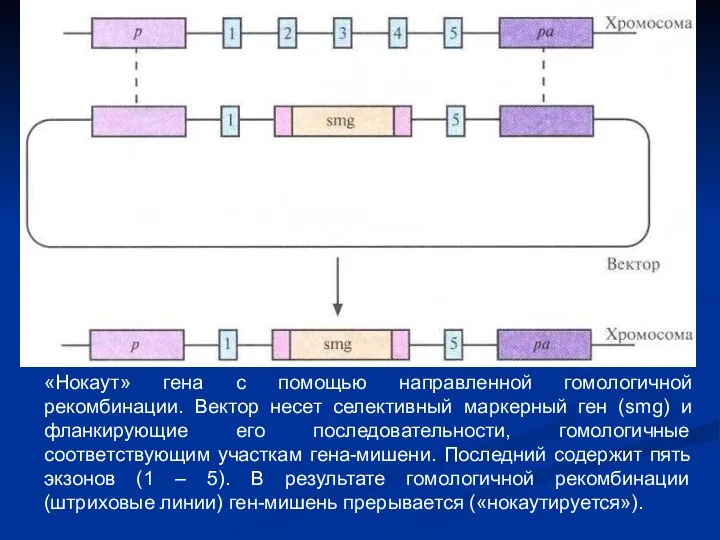
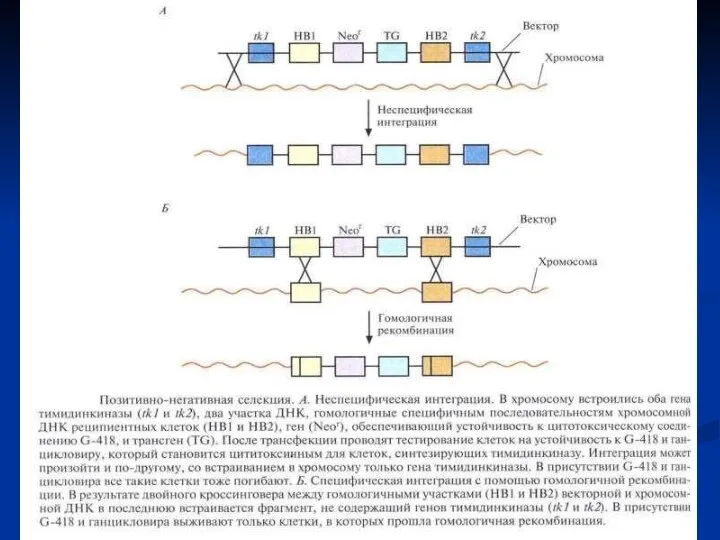
«Нокаут» гена с помощью направленной гомологичной рекомбинации. Вектор несет селективный маркерный
«Нокаут» гена с помощью направленной гомологичной рекомбинации. Вектор несет селективный маркерный

Векторы для клонирования крупных фрагментов ДНК
Литический путь развития λ.
А. При
Векторы для клонирования крупных фрагментов ДНК
Литический путь развития λ.
А. При

Клонирующая система на основе бактериофага λ. Фаговая ДНК имеет два BamH1-сайта,
Клонирующая система на основе бактериофага λ. Фаговая ДНК имеет два BamH1-сайта,
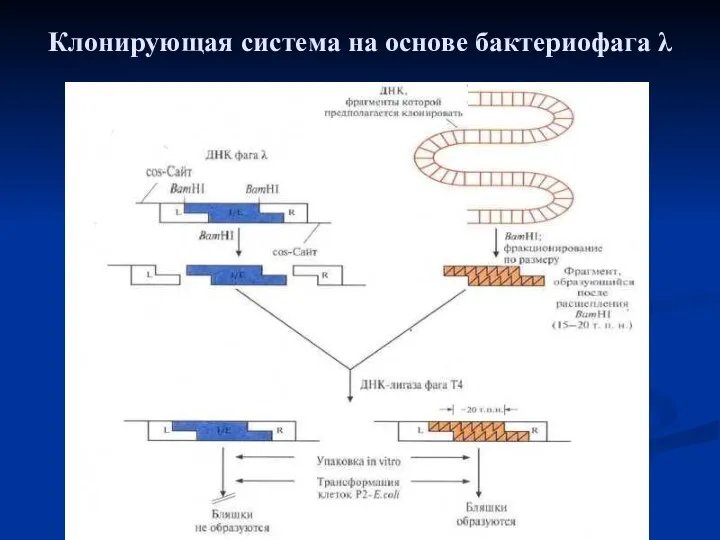
Клонирующая система на основе бактериофага λ
Клонирующая система на основе бактериофага λ

Лигирование липких и тупых концов в молекуле ДНК фагом Т4
ДНК
Лигирование липких и тупых концов в молекуле ДНК фагом Т4
ДНК

Клонирование с помощью космидного вектора
Космида имеет точку начала репликации (ori),
Клонирование с помощью космидного вектора
Космида имеет точку начала репликации (ori),
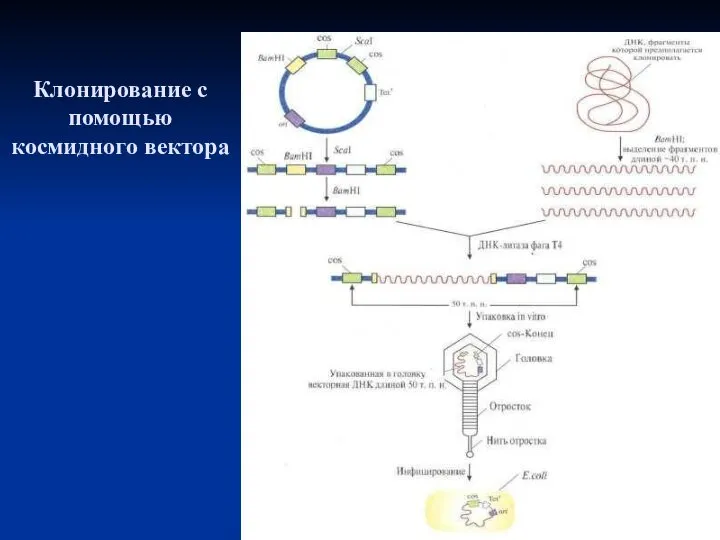
Клонирование с помощью космидного вектора
Клонирование с помощью космидного вектора

Перенос генов с помощью искусственных дрожжевых хромосом
Перенос генов с помощью искусственных дрожжевых хромосом

Идентификация клеток, несущих трансген
Идентификация клеток, несущих трансген


Методы получения трансгенных животных:
1. Метод микроиньекции
2. Вирусный метод
3. Эмбриональный метод
Методы получения трансгенных животных:
1. Метод микроиньекции
2. Вирусный метод
3. Эмбриональный метод

Метод микроиньекции
У самок вызывают гиперовуляцию после чего проводят спаривание с самцами.
Метод микроиньекции
У самок вызывают гиперовуляцию после чего проводят спаривание с самцами.
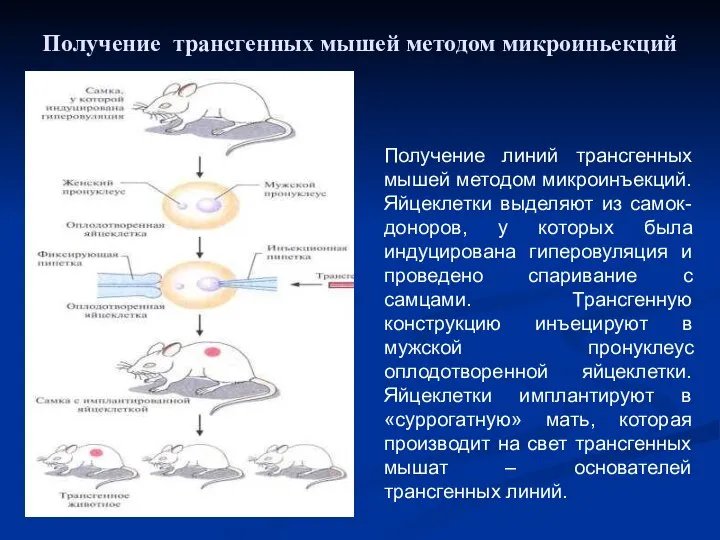
Получение линий трансгенных мышей методом микроинъекций. Яйцеклетки выделяют из самок-доноров, у
Получение линий трансгенных мышей методом микроинъекций. Яйцеклетки выделяют из самок-доноров, у

Метод микроинъекций ДНК
Увеличение числа яйцеклеток, в которых будет инъецирована чужеродная ДНК,
Метод микроинъекций ДНК
Увеличение числа яйцеклеток, в которых будет инъецирована чужеродная ДНК,

Вирусный метод
Эмбрион, находящийся на стадии 8 клеток, инфицируют рекомбинантным ретровирусом, несущим
Вирусный метод
Эмбрион, находящийся на стадии 8 клеток, инфицируют рекомбинантным ретровирусом, несущим

Получение трансгенных мышей с использованием ретровирусных векторов
Получение линии трансгенных мышей с
Получение трансгенных мышей с использованием ретровирусных векторов
Получение линии трансгенных мышей с

Эмбриональный метод
Эмбриональные стволовые клетки получают из внутренней клеточной массы бластоцисты мыши.
Эмбриональный метод
Эмбриональные стволовые клетки получают из внутренней клеточной массы бластоцисты мыши.
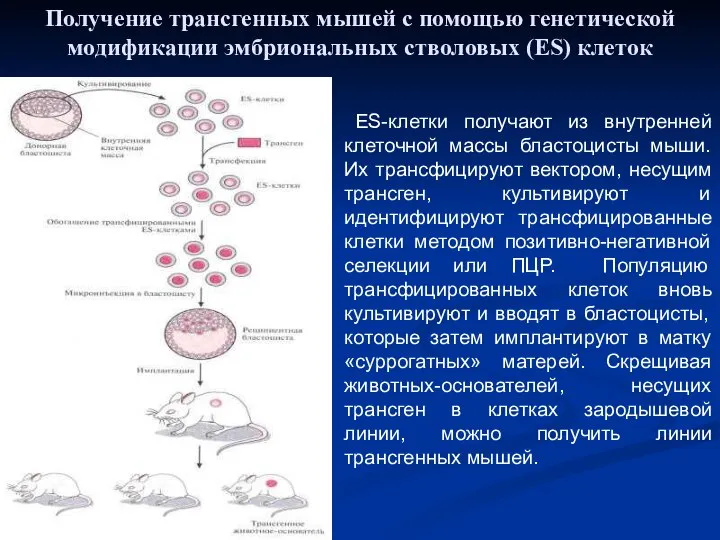
ES-клетки получают из внутренней клеточной массы бластоцисты мыши. Их трансфицируют
ES-клетки получают из внутренней клеточной массы бластоцисты мыши. Их трансфицируют

Использование модифицированных эмбриональных стволовых клеток
Клетки, выделенные из мышиных эмбрионов на стадии
Использование модифицированных эмбриональных стволовых клеток
Клетки, выделенные из мышиных эмбрионов на стадии

Трансгенные животные
Трансгенные животные

Стратегия по введению чужеродных генов в клетки млекопитающих состоит в следующем:
Клонированный
Стратегия по введению чужеродных генов в клетки млекопитающих состоит в следующем:
Клонированный

Клонирование овцы методом переноса ядра
Клонирование овцы методом переноса ядра. Ядро яйцеклетки
Клонирование овцы методом переноса ядра
Клонирование овцы методом переноса ядра. Ядро яйцеклетки
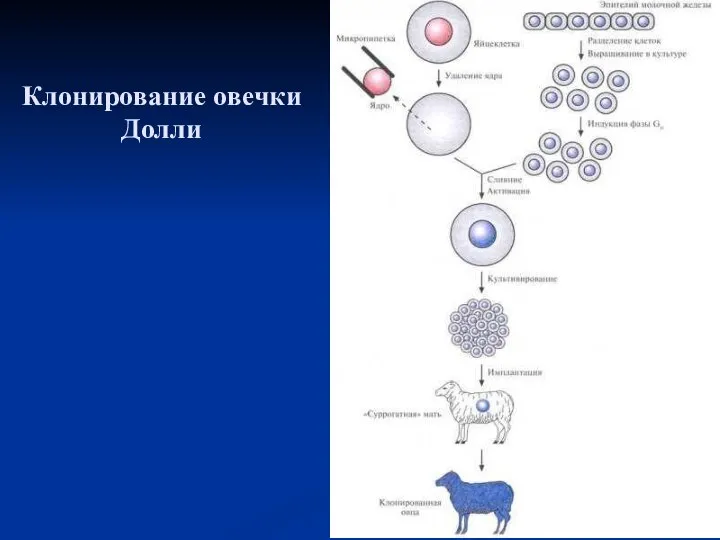
Клонирование овечки Долли
Клонирование овечки Долли

Трансгенный крупный рогатый скот
Если предполагается использовать молочную железу в качестве «биореактора»,
Трансгенный крупный рогатый скот
Если предполагается использовать молочную железу в качестве «биореактора»,

Для создания трансгенных коров использовали модифицированную схему трансгеноза мышей методом микроинъекций
Для создания трансгенных коров использовали модифицированную схему трансгеноза мышей методом микроинъекций
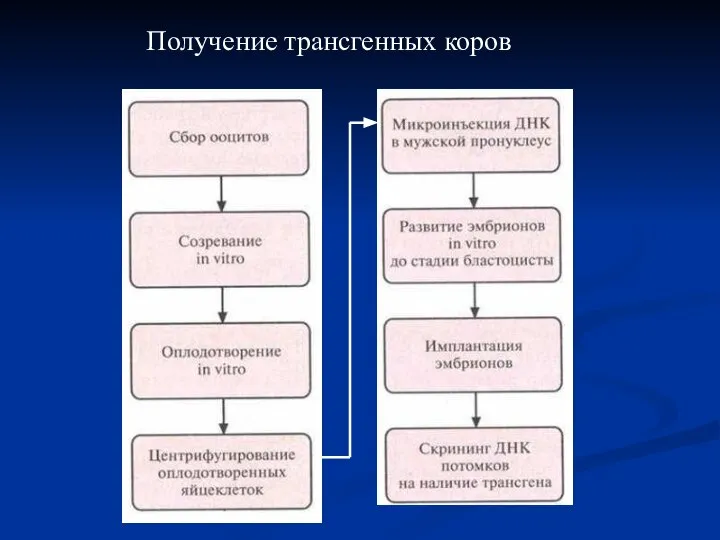
Получение трансгенных коров
Получение трансгенных коров

Трансгенные овцы, козы и свиньи
Трансгенные овцы, козы и свиньи

Трансгенные птицы
Микроинъекция ДНК в оплодотворенные яйцеклетки птиц с целью получения трансгенных
Трансгенные птицы
Микроинъекция ДНК в оплодотворенные яйцеклетки птиц с целью получения трансгенных

Получение трансгенных цыплят трансфекцией изолированных клеток бластодермы. Выделенные клетки трансфицируют трансгеном
Получение трансгенных цыплят трансфекцией изолированных клеток бластодермы. Выделенные клетки трансфицируют трансгеном

Трансгенные рыбы
По мере истощения природных рыбных запасов все большую роль будет
Трансгенные рыбы
По мере истощения природных рыбных запасов все большую роль будет

Трансгенные мыши: методология
Введение чужеродной ДНК мышам можно осуществить разными методами:
С помощью
Трансгенные мыши: методология
Введение чужеродной ДНК мышам можно осуществить разными методами:
С помощью

Трансгенные мыши: применение как модельного обьекта изучения болезни Альцгеймера.
Болезнь Альцгеймера —
Трансгенные мыши: применение как модельного обьекта изучения болезни Альцгеймера.
Болезнь Альцгеймера —

Схематическое изображение нейрона коры головного мозга человека с указанием некоторых гистологических
Схематическое изображение нейрона коры головного мозга человека с указанием некоторых гистологических

Основным компонентом сенильных и амилоидных бляшек является белок Ар (амилоид р\
Основным компонентом сенильных и амилоидных бляшек является белок Ар (амилоид р\
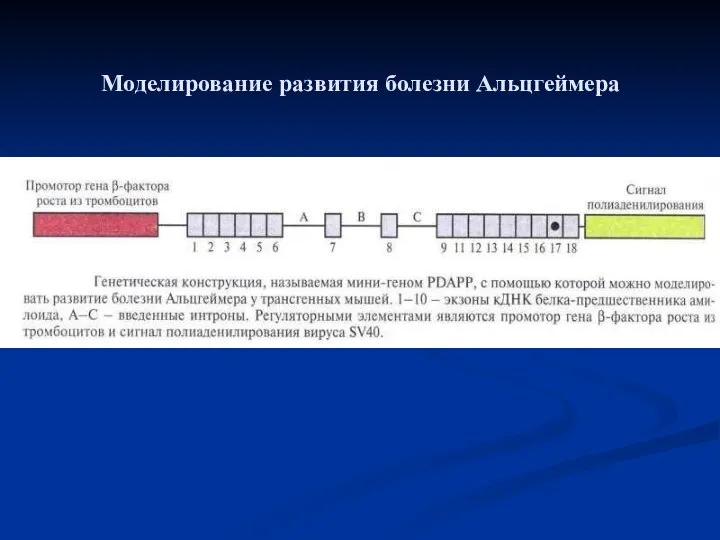
Моделирование развития болезни Альцгеймера
Моделирование развития болезни Альцгеймера

Муковисцидоз — распространенная генетическая болезнь, поражающая в странах Европы одного из
Муковисцидоз — распространенная генетическая болезнь, поражающая в странах Европы одного из


Гибридизация соматических клеток
Гибридизация соматических клеток

Методика изоляции соматических гибридов (по Шапиро)
ТК – фермент тимидинкиназа
ГГФРТ – фермент
Методика изоляции соматических гибридов (по Шапиро)
ТК – фермент тимидинкиназа
ГГФРТ – фермент

Некоторые сочетания геномов, полученные слиянием соматических клеток
Некоторые сочетания геномов, полученные слиянием соматических клеток
 Олимпиадный эксперимент – в школьный практикум. Часть 1
Олимпиадный эксперимент – в школьный практикум. Часть 1 Условия равновесия твердых тел
Условия равновесия твердых тел Рівноприскорений рух прискорення
Рівноприскорений рух прискорення Излучение источников. Интегральная показательная функция. (Лекция 5)
Излучение источников. Интегральная показательная функция. (Лекция 5) test_uravnenie_m-k-t
test_uravnenie_m-k-t Исследование возможности накопления энергии из высоковольтного разряда
Исследование возможности накопления энергии из высоковольтного разряда Надпровідність
Надпровідність Центр тяжести
Центр тяжести Двигатель. Классификация двигателей
Двигатель. Классификация двигателей Трансформаторы. Устройство трансформатора
Трансформаторы. Устройство трансформатора Раздаточные коробки передач
Раздаточные коробки передач Электр жүктемелерінің графиктері
Электр жүктемелерінің графиктері Механическое равновесие, механические колебания и волны
Механическое равновесие, механические колебания и волны Основные свойства диэлектриков. МДМ структуры и тонкие пленки. Лекция 2
Основные свойства диэлектриков. МДМ структуры и тонкие пленки. Лекция 2 Лауреат Нобелевской премии Артур Эшкин
Лауреат Нобелевской премии Артур Эшкин Испытания композитных материалов и конструкций
Испытания композитных материалов и конструкций Скачать презентацию Действие электрического тока на человека
Скачать презентацию Действие электрического тока на человека  Феноменологическая термодинамика. (Тема 11)
Феноменологическая термодинамика. (Тема 11) Наши достижения. Исследовательский проект выполнил: Воробьёв Антон, ученик 7 «А» класса МОУ СОШ №2 г.Уварово. Научный руководител
Наши достижения. Исследовательский проект выполнил: Воробьёв Антон, ученик 7 «А» класса МОУ СОШ №2 г.Уварово. Научный руководител Методы и средства измерения полосы пропускания и дисперсии оптических волокон
Методы и средства измерения полосы пропускания и дисперсии оптических волокон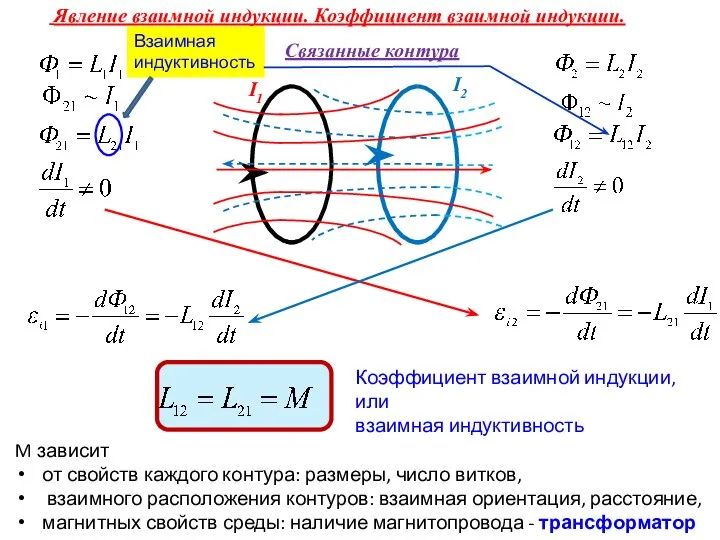 Явление взаимной индукции. Коэффициент взаимной индукции. Взаимная индуктивность
Явление взаимной индукции. Коэффициент взаимной индукции. Взаимная индуктивность Магнитное поле
Магнитное поле Диагностирование двигателя по параметрам расхода топлива. Тема 12
Диагностирование двигателя по параметрам расхода топлива. Тема 12 Механические передачи. Детали машин
Механические передачи. Детали машин Закон всемирного тяготения
Закон всемирного тяготения Лекція 6. Визначення напружень при згинанні. Умови міцності при згинанні
Лекція 6. Визначення напружень при згинанні. Умови міцності при згинанні Короткое замыкание. Предохранители
Короткое замыкание. Предохранители Комплексные частотные характеристики линейных электрических цепей
Комплексные частотные характеристики линейных электрических цепей