- Нервово-мязова передача. (Лекція 3)

Содержание
- 2. Нервово-м′язове сполучення – це спеціалізований вид синапсу між закінченнями рухового нейрона (мотонейрона) і ендомізієм м′язових волокон.
- 3. Рис. 1. Розгалуження моторного нейрона на поверхні м’язових волокон
- 4. Функції нервово-м′язових синапсів Однобічність передачі. Посилення. Адаптація, або акомодація. Гальмування. Сумація. Дискримінація. Рефрактерні фази міоневрального апарату
- 5. Розвиток нервово-м′язових сполучень у хребетних в онтогенезі.
- 6. Виділяють чотири основних механізми взаємодії між м′язом і нервом, що призводять до диференціювання нервово-м′язових синапсів. Виділення
- 7. Тонка будова нервово-м′язового синапсу скелетного м′язу.
- 8. Рис. 2. Мікрофотографія нервово-м′язового сполучення людини. Ах – закінчення аксону; * - шваннівська клітина (леммоцит); MN
- 9. Рис. 3. Схема будови нервово-м'язового синапсу
- 10. Рис. 4. Нервово-м’язовий контакт (хімічний синапс): а,в – поступове збільшення фрагмента обведено рамкою на позиції а
- 11. У 1936 р. Дейлом, Фельдбергом і Фогтом було встановлено, що передатчиком збудження у нервово-м′язових синапсах є
- 12. Синтез ацетилхоліну. АХ синтезується у холінергічних нейронах у результаті реакції між холіном і ацетил-коферментом А: Холін
- 13. Виділення ацетилхоліну. Елементарна одиниця кількості нейротрансмітера отримала назву квант. У 1956 р. Дель-Кастільо і Кац запропонували
- 14. Гіпотези виділення ацетилхоліну АХ може виділятись у синаптичну щілину і безпосередньо з позапухирцевого простору нервового закінчення
- 15. Процес виділення АХ є короткочасним і обмежується у часі такими факторами: 1) йони кальцію швидко зв′язуються
- 16. Феномени пов′язані з процесами виділення АХ: 1) Н.Є. Введенський встановив, що при зростанні частоти ритмічної стимуляції
- 17. Взаємодія нейротрансміттера (АХ) із синаптичними рецепторами. Кастільо і Катц (1957) запропонували розглядати взаємодію між АХ з
- 18. Рис. 5. Взаємодія нейротрансміттера (АХ) із синаптичними рецепторами
- 19. Рис. 6. Зображення ацетилхолінового рецептора з двома α-, а також β-, γ- і δ-субодиницями. Зліва показаний
- 20. Рис. 7. Холінорецептор електричного органа риби: тривимірне зображення рецептора збоку (а) і згори (б); в –
- 21. Види постсинаптичних потенціалів КП: Деполяризуючий потенціал кінцевої пластинки. Гіперполяризуючий постсинаптичний потенціал кінцевої пластинки.
- 22. Відмінності збуджувальних (ЗС) і гальмівних (ГС) синапсів: 1) У ЗС ширша синаптична щілина у 30 нм,
- 24. Скачать презентацию

Нервово-м′язове сполучення – це спеціалізований вид синапсу між закінченнями рухового нейрона
Нервово-м′язове сполучення – це спеціалізований вид синапсу між закінченнями рухового нейрона
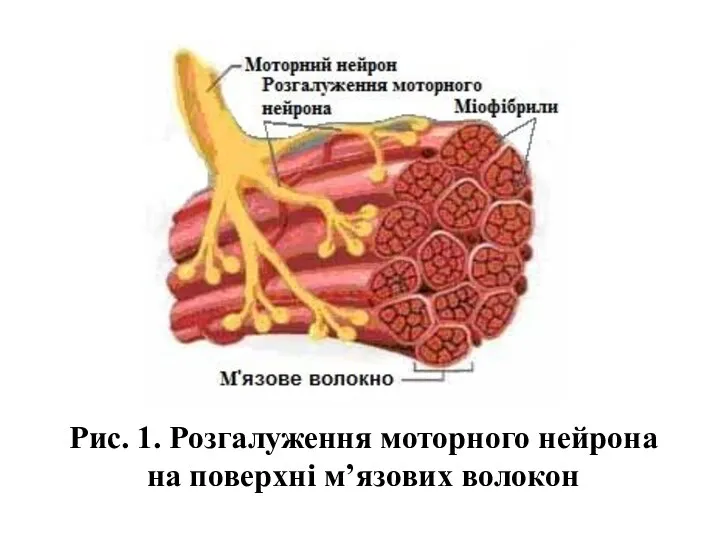
Рис. 1. Розгалуження моторного нейрона на поверхні м’язових волокон
Рис. 1. Розгалуження моторного нейрона на поверхні м’язових волокон

Функції нервово-м′язових синапсів
Однобічність передачі.
Посилення.
Адаптація, або акомодація.
Гальмування.
Сумація.
Дискримінація.
Функції нервово-м′язових синапсів
Однобічність передачі.
Посилення.
Адаптація, або акомодація.
Гальмування.
Сумація.
Дискримінація.

Розвиток нервово-м′язових сполучень у хребетних в онтогенезі.
Розвиток нервово-м′язових сполучень у хребетних в онтогенезі.

Виділяють чотири основних механізми взаємодії між м′язом і нервом, що призводять
Виділяють чотири основних механізми взаємодії між м′язом і нервом, що призводять

Тонка будова нервово-м′язового синапсу скелетного м′язу.
Тонка будова нервово-м′язового синапсу скелетного м′язу.
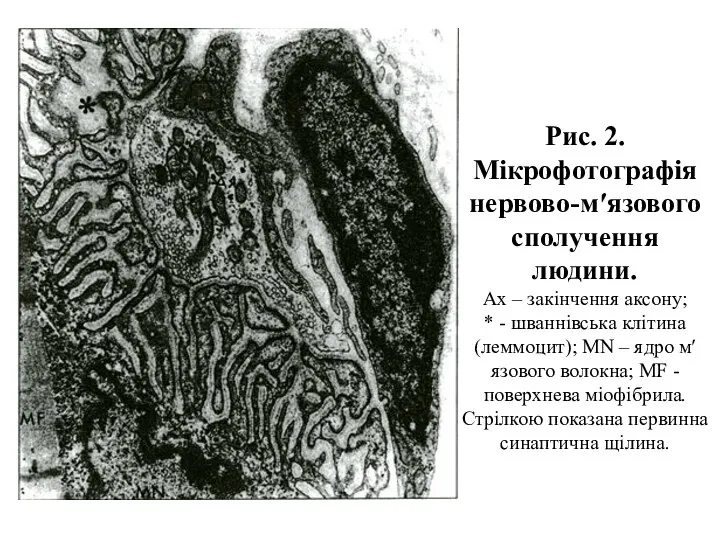
Рис. 2. Мікрофотографія нервово-м′язового сполучення людини.
Ах – закінчення аксону;
* -
Рис. 2. Мікрофотографія нервово-м′язового сполучення людини. Ах – закінчення аксону; * -
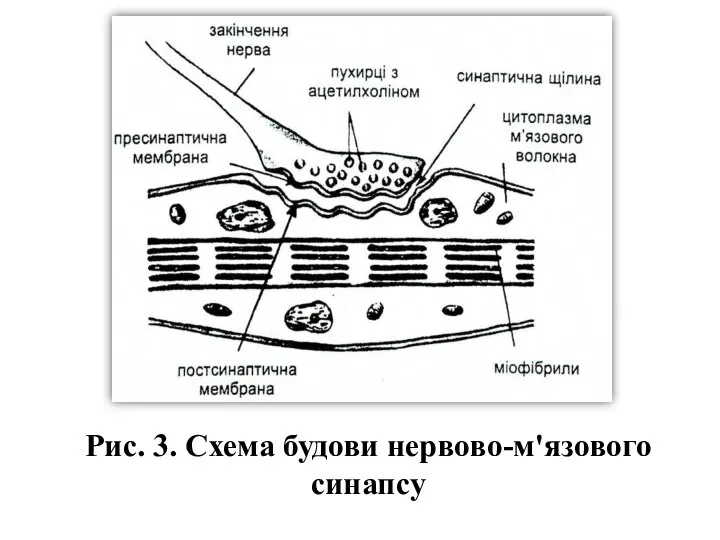
Рис. 3. Схема будови нервово-м'язового синапсу
Рис. 3. Схема будови нервово-м'язового синапсу
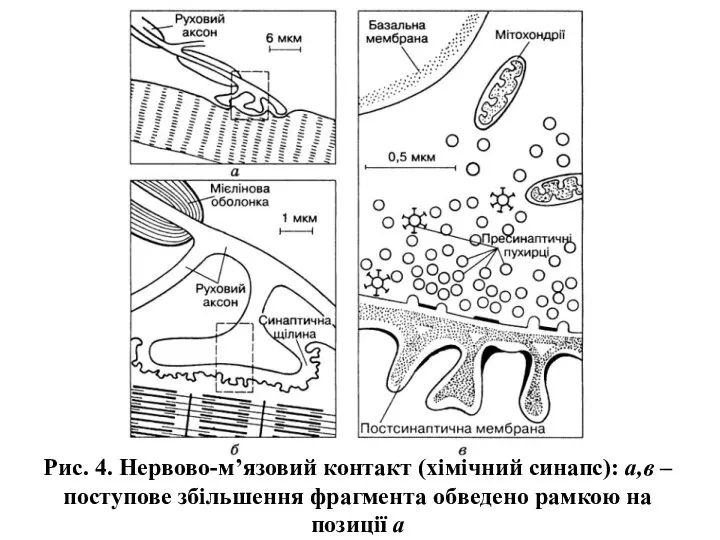
Рис. 4. Нервово-м’язовий контакт (хімічний синапс): а,в – поступове збільшення фрагмента
Рис. 4. Нервово-м’язовий контакт (хімічний синапс): а,в – поступове збільшення фрагмента

У 1936 р. Дейлом, Фельдбергом і Фогтом було встановлено, що передатчиком
У 1936 р. Дейлом, Фельдбергом і Фогтом було встановлено, що передатчиком

Синтез ацетилхоліну.
АХ синтезується у холінергічних нейронах у результаті реакції між
Синтез ацетилхоліну.
АХ синтезується у холінергічних нейронах у результаті реакції між

Виділення ацетилхоліну.
Елементарна одиниця кількості нейротрансмітера отримала назву квант.
У 1956 р. Дель-Кастільо
Виділення ацетилхоліну.
Елементарна одиниця кількості нейротрансмітера отримала назву квант.
У 1956 р. Дель-Кастільо

Гіпотези виділення ацетилхоліну
АХ може виділятись у синаптичну щілину і безпосередньо з
Гіпотези виділення ацетилхоліну
АХ може виділятись у синаптичну щілину і безпосередньо з

Процес виділення АХ є короткочасним і обмежується у часі такими факторами:
1)
Процес виділення АХ є короткочасним і обмежується у часі такими факторами:
1)

Феномени пов′язані з процесами виділення АХ:
1) Н.Є. Введенський встановив, що при
Феномени пов′язані з процесами виділення АХ:
1) Н.Є. Введенський встановив, що при

Взаємодія нейротрансміттера (АХ) із синаптичними рецепторами.
Кастільо і Катц (1957) запропонували
Взаємодія нейротрансміттера (АХ) із синаптичними рецепторами.
Кастільо і Катц (1957) запропонували
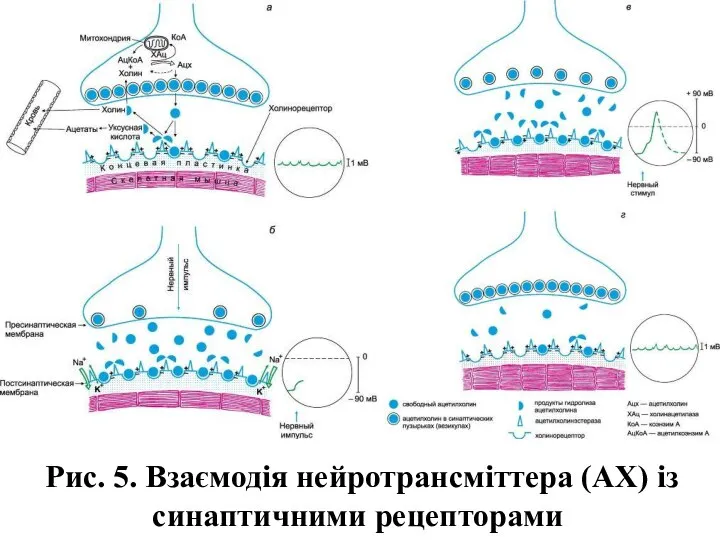
Рис. 5. Взаємодія нейротрансміттера (АХ) із синаптичними рецепторами
Рис. 5. Взаємодія нейротрансміттера (АХ) із синаптичними рецепторами
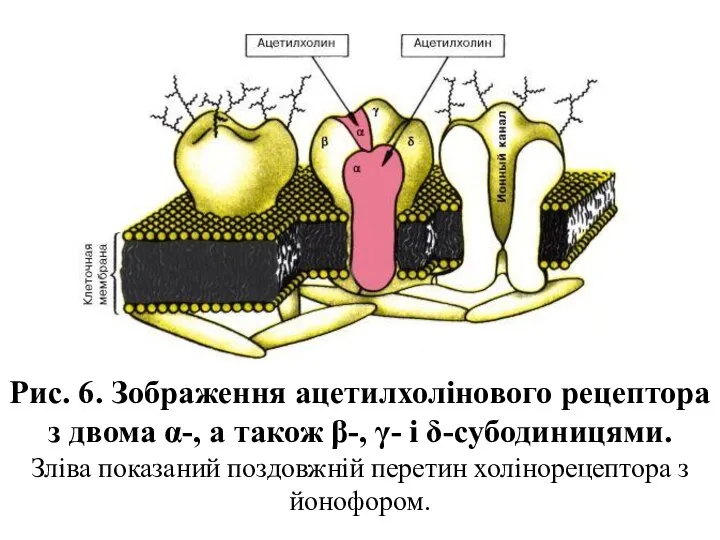
Рис. 6. Зображення ацетилхолінового рецептора з двома α-, а також β-,
Рис. 6. Зображення ацетилхолінового рецептора з двома α-, а також β-,
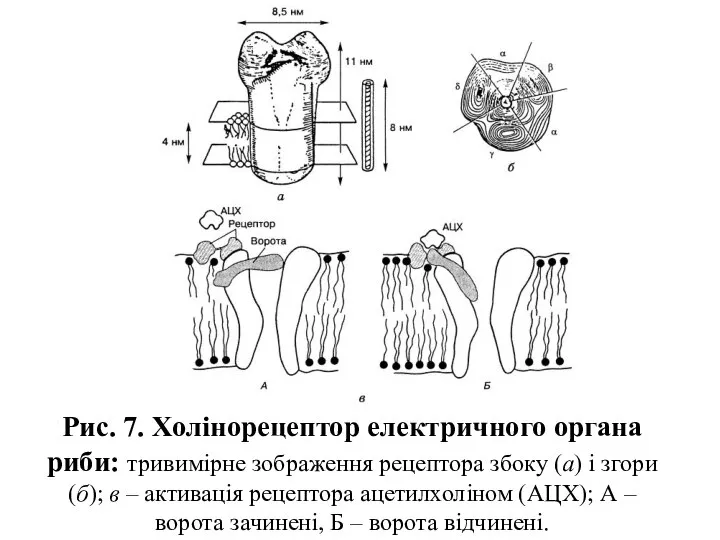
Рис. 7. Холінорецептор електричного органа риби: тривимірне зображення рецептора збоку (а)
Рис. 7. Холінорецептор електричного органа риби: тривимірне зображення рецептора збоку (а)

Види постсинаптичних
потенціалів КП:
Деполяризуючий потенціал кінцевої пластинки.
Гіперполяризуючий постсинаптичний потенціал кінцевої пластинки.
Види постсинаптичних
потенціалів КП:
Деполяризуючий потенціал кінцевої пластинки.
Гіперполяризуючий постсинаптичний потенціал кінцевої пластинки.

Відмінності збуджувальних (ЗС) і гальмівних (ГС) синапсів:
1) У ЗС ширша синаптична
Відмінності збуджувальних (ЗС) і гальмівних (ГС) синапсів:
1) У ЗС ширша синаптична
 Принцип построения МРТ
Принцип построения МРТ Основи випромінювання електромагнітних хвиль
Основи випромінювання електромагнітних хвиль Наддув ДВС. Системы Наддува. Лекция №3
Наддув ДВС. Системы Наддува. Лекция №3 Физические качества
Физические качества Электронный парамагнитный резонанс
Электронный парамагнитный резонанс Движение тела в гравитационном поле.
Движение тела в гравитационном поле. Уклон и конусность
Уклон и конусность Лекция 42. Атомная физика
Лекция 42. Атомная физика Методы исследования поверхности и/или тонких слоев
Методы исследования поверхности и/или тонких слоев УРОК ФИЗИКИ В 10 КЛАССЕ Законы Ньютона Учитель Кононов Геннадий Григорьевич СОШ № 29 Славянский район Краснод
УРОК ФИЗИКИ В 10 КЛАССЕ Законы Ньютона Учитель Кононов Геннадий Григорьевич СОШ № 29 Славянский район Краснод Определение коэффициента теплопроводности изоляционного материала методом трубы
Определение коэффициента теплопроводности изоляционного материала методом трубы Электрический ток
Электрический ток Презентация по физике "Система отсчёта" - скачать
Презентация по физике "Система отсчёта" - скачать  Второй закон термодинамики
Второй закон термодинамики Магистральды төменгі жиілікті симметриялы кабельдер
Магистральды төменгі жиілікті симметриялы кабельдер Мотоцикл
Мотоцикл Занимательные опыты по физике
Занимательные опыты по физике Прямолинейное равноускоренное движение
Прямолинейное равноускоренное движение Самоблокирующийся дифференциал
Самоблокирующийся дифференциал Квантовая природа тзлучения
Квантовая природа тзлучения Физика 10 класс 1 урок в разделе «Молекулярная физика»
Физика 10 класс 1 урок в разделе «Молекулярная физика» Презентация Гидростатическое давление
Презентация Гидростатическое давление Общая теория относительности Эйнштейна
Общая теория относительности Эйнштейна Некогерентность рассеяния. (Тема 15)
Некогерентность рассеяния. (Тема 15) Статические характеристики средств измерений. (Лекция 2)
Статические характеристики средств измерений. (Лекция 2) Трансформаторы тока
Трансформаторы тока Плавление и кристаллизация
Плавление и кристаллизация Презентация на тему "Электроёмкость. Конденсаторы"
Презентация на тему "Электроёмкость. Конденсаторы"