- Изоформы ядерных рецепторов и их функциональное значение

Содержание
- 3. Сравнение С-концевых лигандсвязывающих доменов канонических вариантов ядерных рецепторов и форм их альтернативного сплайсинга по наличию 12-й
- 4. Возможные механизмы доминант-негативной активности альтернативного варианта рецептора с измененной С-концевой последовательностью Сплайсинговый вариант лигандсвязывающего домена Коактиватор
- 5. A/B DBD LBD 66 кДа 46 кДа ядро плазматическая мембрана пальмитат Сплайсинговый вариант рецептора альфа эстрогенов
- 6. Фосфорилирование как способ регуляции активности ядерных рецепторов
- 7. Фосфорилирование как способ регуляции активности ЯР Фосфорилирование а/к остатков в домена А/В Конститутивное фосфорилирование Зависимое от
- 8. Влияние фосфорилирования N-конца PPARγ на его сумоилирование оЛНП – окисленные липопротеиды низкой плотности; Фосфорилирование как способ
- 9. Терминация рецепторного цикла
- 10. Терминация рецепторного цикла Лиганд ускоряет деградацию собственного рецептора: Свободный эстрогенный рецептор: Т1/2=5 дней Комплекс эстрогенного рецептора
- 11. Пример влияния сумоилирования Индуцируемое лигандом сумоилирование C-конца PPARγ не влияет на трансактивационную функцию PPARγ (А) Индуцируемое
- 12. Негативная регуляция транскрипции ядерными рецепторами
- 13. Варианты активирующего первичного геномного действия половых стероидов Транскрипционные факторы SP1 и AP-1 могут опосредовать взаимодействие ядерных
- 14. Негативные гормончувствительные элементы (nRE) ОСОБЕННОСТИ nRE: 1. Отличаются от позитивных гормончувствительных элементов (pRE) по последовательностям нуклеотидов,
- 15. Негативная регуляция транскрипции ядерными рецепторами (перекрытие с сайтом связывания транскрипционного фактора SP1)
- 16. Негативная регуляция транскрипции ядерными рецепторами (аллостерическое измерение конформации NR – рекрутирование корепрессора) Негативный гормончувствительный элемент (nHRE)
- 17. гОбратные агонисты стимулируют рекрутирование рецептором корепрессора вместо коактиватора Негативная регуляция транскрипции ядерными рецепторами (Действие обратных агонистов)
- 18. Негативная регуляция транскрипции ядерными рецепторами (аллостерическое измерение конформации NR- изменение конформации коактиватора)
- 19. Негативная регуляция транскрипции ядерными рецепторами (Транс-репрессия – конкуренция NR за коактиватор)
- 20. Негативная регуляция транскрипции ядерными рецепторами (Транс-репрессия – корепрессорное действие NR)
- 21. Патологии, связанные с мутациями ядерных рецепторов
- 22. Мутации ядерных рецепторов на примере андрогенного рецептора (АР) Полиморфизм АР с изменением количества триплетных повторов (CAG)n
- 23. Мутация, ведущая к дисфункции андрогенного рецептора Мутация андрогенсвязывающего кармана андрогенного рецептора Появление способности неактивных андрогенов и
- 24. Рецептор 1,25(OH)2-D3 (VDR) 1 424 ДНК Лиганд Нарушения, связанные с мутациями ядерных рецепторов Большинство мутаций (замены
- 25. Рецепторы арильных углеводородов
- 26. Поллютант агонист 2,3,7,8-тетрахлордибензо-п-диоксин (а), Растительный антагонист кампферол (б), Эндогенные агонисты: метаболиты триптофана - 3-индоксилсульфат, или индикан
- 27. Структура рецептора арильных углеводородов (AhR) AhR функционирует сходно с ядерными рецепторами, хотя структурно с ними не
- 28. Роль комплексирования рецептора арильных углеводородов с белками теплового шока В цитоплазме гетеродимеры: Неактивный AhR+ димер белка
- 29. XRE мРНК CYP1A1 AhRR и др. Hsp90 XAP2 AhR лиганд Метаболизм ксенобиотиков AhRR Arnt ядро цитоплазма
- 30. Участие AhR в активации канцерогенов Бензо[a]пирен – компонент табачного дыма, выхлопных газов
- 31. AhR и репродуктивная функция Активация AhR ведет к ускоренной инактивации половых гормонов за счет индукции метаболизирующих
- 32. Сенсор электрофильных ксенобиотиков и окислительного стресса белок Keap1
- 33. Сенсор электрофильных ксенобиотиков и окислительного стресса Keap1 в регуляции убиквитинилирования транскрипционного фактора Nrf2
- 34. 624 повторы Kelch (связывание актина и Nrf2) NES (удаление Nrf2 из ядра) Линкер (связывание убиквитинлигазы Cul3)
- 35. Мембранные рецепторы группы Notch, активируемые зависимым от лиганда протеолизом Контактный способ передачи сигнала
- 36. Лиганды рецептора Notch Интегральные трансмембранные белки с одним трансмембранным доменом: Delta-like-1, Delta-like-3, Delta-like-4, Jagged-1, Jagged-2 Основной
- 37. Принцип работы рецепторов Notch Взаимодействие с лигандом, заякоренным на соседней клетке Двухступечатое протеолитическое отщепление внутриклеточного домена
- 38. Семейство рецепторов Notch Гетеродимеры: Внеклеточная лигандсвязывающая субъединица и трансмембранная субъединица связаны дисульфидными связями После связывания с
- 39. Внутриклеточный домен Notch (NIC) индуцирует вытеснение корепрессорного белкового комплекса из связи с транскрипционным фактором CBF и
- 41. Скачать презентацию

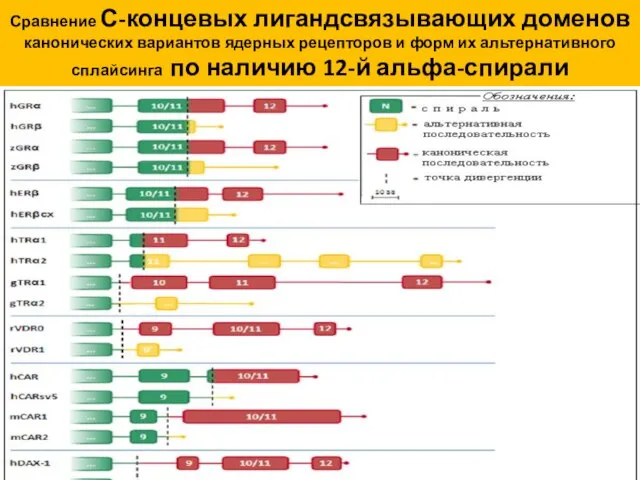
Сравнение С-концевых лигандсвязывающих доменов
канонических вариантов ядерных рецепторов и форм их
Сравнение С-концевых лигандсвязывающих доменов
канонических вариантов ядерных рецепторов и форм их
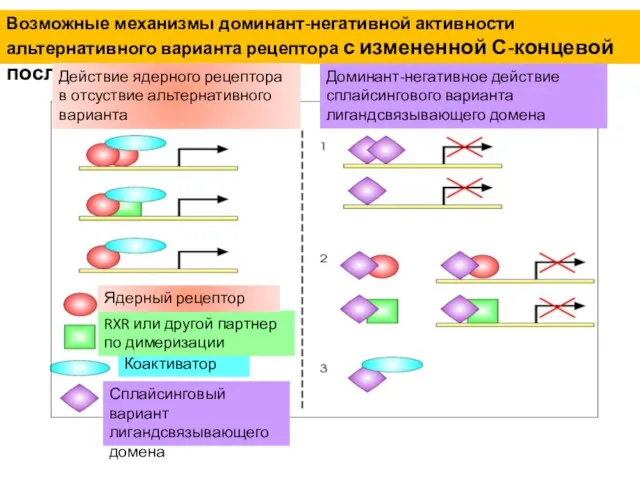
Возможные механизмы доминант-негативной активности альтернативного варианта рецептора с измененной С-концевой последовательностью
Сплайсинговый
Возможные механизмы доминант-негативной активности альтернативного варианта рецептора с измененной С-концевой последовательностью
Сплайсинговый
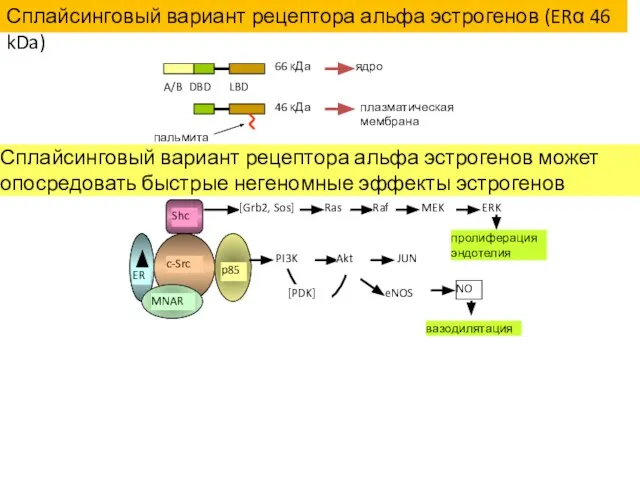
A/B
DBD
LBD
66 кДа
46 кДа
ядро
плазматическая мембрана
пальмитат
Сплайсинговый вариант рецептора альфа эстрогенов может опосредовать быстрые
A/B
DBD
LBD
66 кДа
46 кДа
ядро
плазматическая мембрана
пальмитат
Сплайсинговый вариант рецептора альфа эстрогенов может опосредовать быстрые

Фосфорилирование как способ регуляции активности ядерных рецепторов
Фосфорилирование как способ регуляции активности ядерных рецепторов

Фосфорилирование как способ регуляции активности ЯР
Фосфорилирование а/к остатков в домена А/В
Конститутивное
Фосфорилирование как способ регуляции активности ЯР
Фосфорилирование а/к остатков в домена А/В
Конститутивное
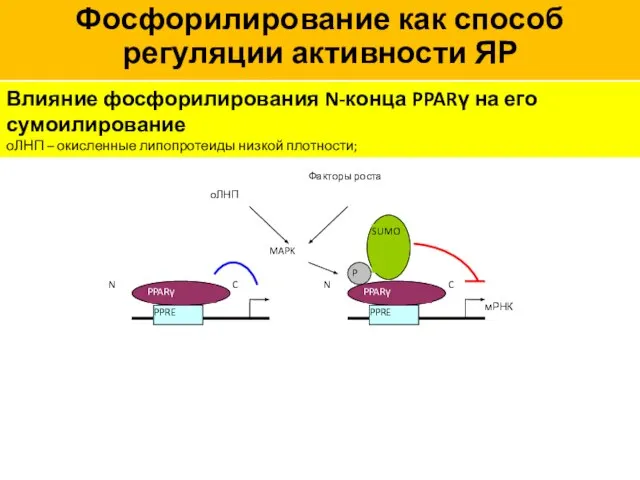
Влияние фосфорилирования N-конца PPARγ на его сумоилирование
оЛНП – окисленные липопротеиды низкой
Влияние фосфорилирования N-конца PPARγ на его сумоилирование
оЛНП – окисленные липопротеиды низкой

Терминация рецепторного цикла
Терминация рецепторного цикла

Терминация рецепторного цикла
Лиганд ускоряет деградацию собственного рецептора:
Свободный эстрогенный рецептор: Т1/2=5 дней
Комплекс
Терминация рецепторного цикла
Лиганд ускоряет деградацию собственного рецептора:
Свободный эстрогенный рецептор: Т1/2=5 дней
Комплекс
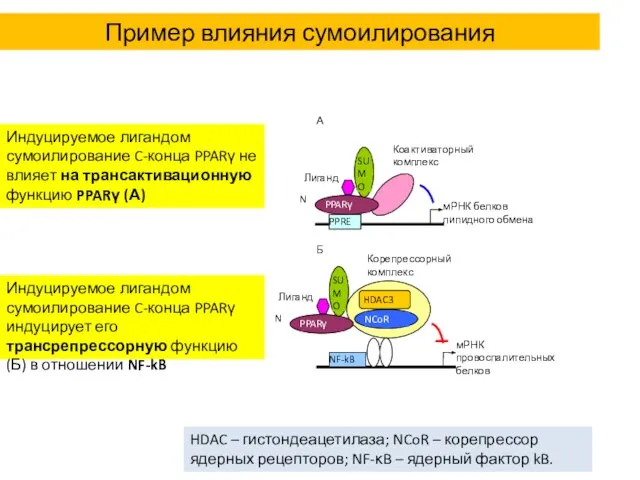
Пример влияния сумоилирования
Индуцируемое лигандом сумоилирование C-конца PPARγ не влияет на трансактивационную
Пример влияния сумоилирования
Индуцируемое лигандом сумоилирование C-конца PPARγ не влияет на трансактивационную

Негативная регуляция транскрипции
ядерными рецепторами
Негативная регуляция транскрипции
ядерными рецепторами
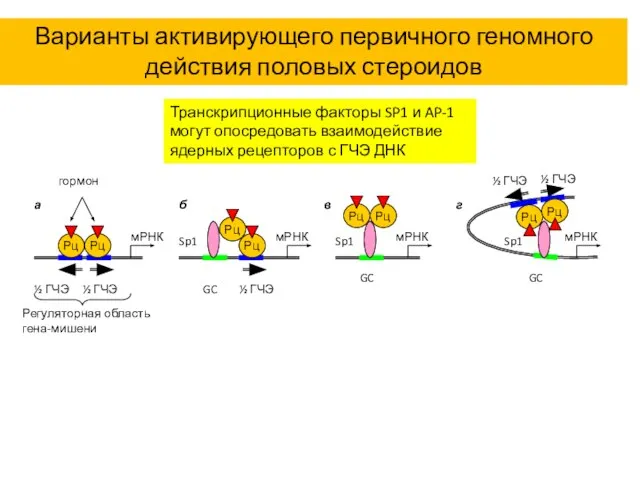
Варианты активирующего первичного геномного действия половых стероидов
Транскрипционные факторы SP1 и AP-1
Варианты активирующего первичного геномного действия половых стероидов
Транскрипционные факторы SP1 и AP-1

Негативные гормончувствительные элементы (nRE)
ОСОБЕННОСТИ nRE:
1. Отличаются от позитивных гормончувствительных элементов (pRE)
Негативные гормончувствительные элементы (nRE)
ОСОБЕННОСТИ nRE:
1. Отличаются от позитивных гормончувствительных элементов (pRE)
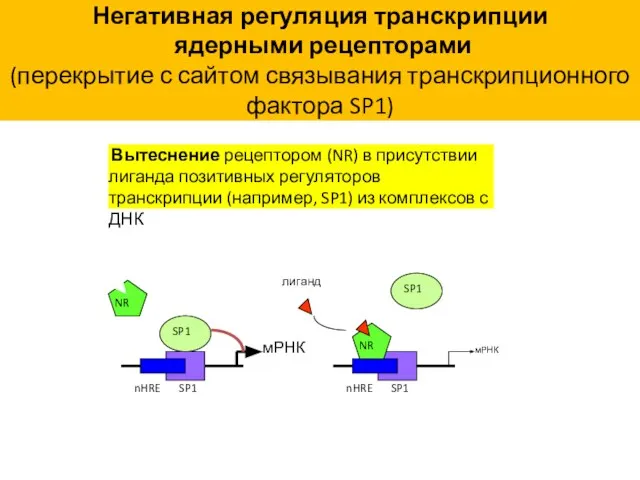
Негативная регуляция транскрипции
ядерными рецепторами
(перекрытие с сайтом связывания транскрипционного фактора
Негативная регуляция транскрипции
ядерными рецепторами
(перекрытие с сайтом связывания транскрипционного фактора
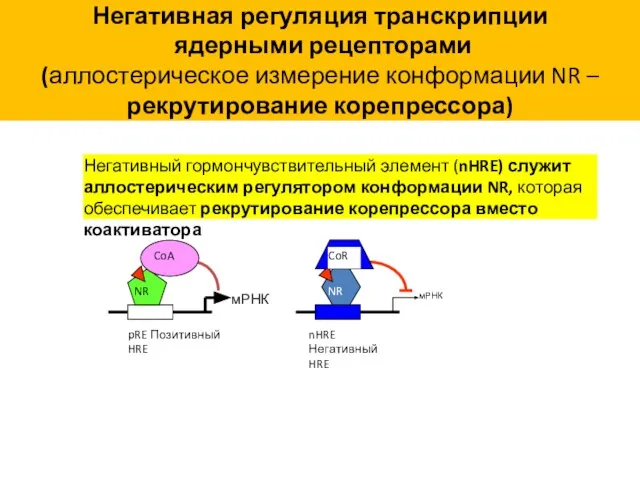
Негативная регуляция транскрипции
ядерными рецепторами
(аллостерическое измерение конформации NR – рекрутирование
Негативная регуляция транскрипции
ядерными рецепторами
(аллостерическое измерение конформации NR – рекрутирование
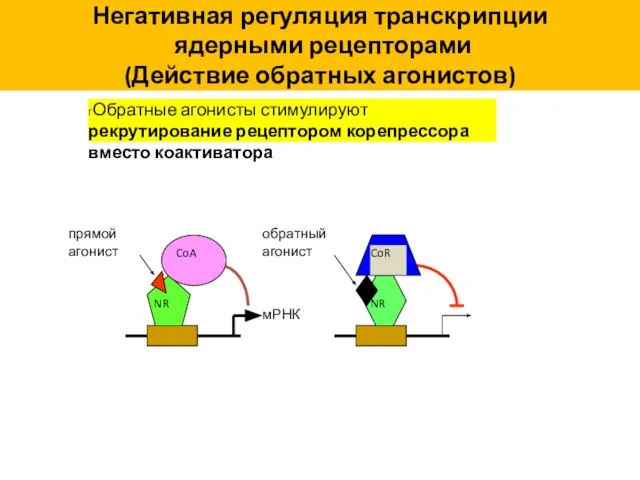
гОбратные агонисты стимулируют рекрутирование рецептором корепрессора вместо коактиватора
Негативная регуляция транскрипции
ядерными
гОбратные агонисты стимулируют рекрутирование рецептором корепрессора вместо коактиватора
Негативная регуляция транскрипции
ядерными
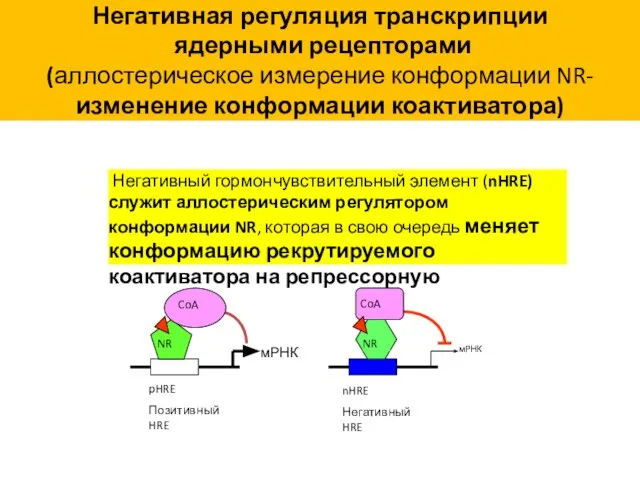
Негативная регуляция транскрипции
ядерными рецепторами
(аллостерическое измерение конформации NR-
изменение конформации коактиватора)
Негативная регуляция транскрипции
ядерными рецепторами
(аллостерическое измерение конформации NR-
изменение конформации коактиватора)
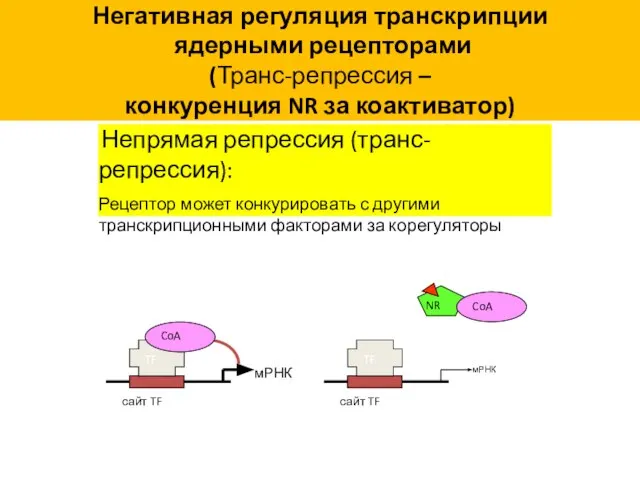
Негативная регуляция транскрипции
ядерными рецепторами
(Транс-репрессия –
конкуренция NR за коактиватор)
Негативная регуляция транскрипции
ядерными рецепторами
(Транс-репрессия –
конкуренция NR за коактиватор)
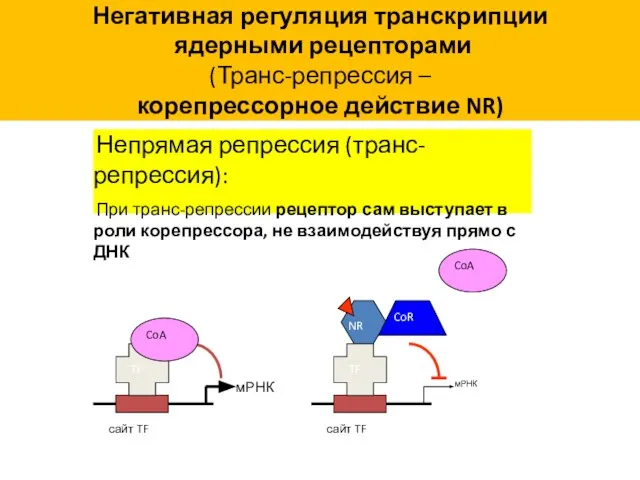
Негативная регуляция транскрипции
ядерными рецепторами
(Транс-репрессия –
корепрессорное действие NR)
Негативная регуляция транскрипции
ядерными рецепторами
(Транс-репрессия –
корепрессорное действие NR)

Патологии, связанные с мутациями ядерных рецепторов
Патологии, связанные с мутациями ядерных рецепторов

Мутации ядерных рецепторов
на примере андрогенного рецептора (АР)
Полиморфизм АР с изменением
Мутации ядерных рецепторов
на примере андрогенного рецептора (АР)
Полиморфизм АР с изменением
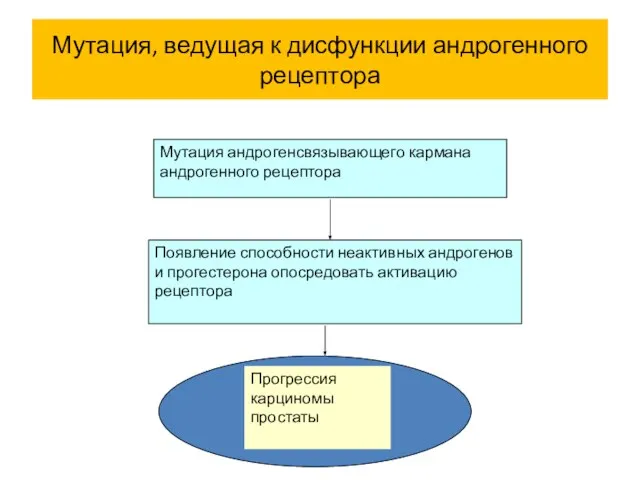
Мутация, ведущая к дисфункции андрогенного рецептора
Мутация андрогенсвязывающего кармана андрогенного рецептора
Появление способности
Мутация, ведущая к дисфункции андрогенного рецептора
Мутация андрогенсвязывающего кармана андрогенного рецептора
Появление способности
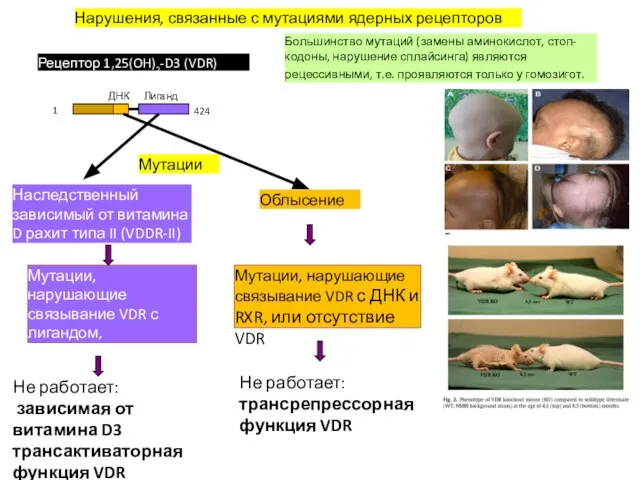
Рецептор 1,25(OH)2-D3 (VDR)
1
424
ДНК
Лиганд
Нарушения, связанные с мутациями ядерных рецепторов
Большинство мутаций (замены аминокислот,
Рецептор 1,25(OH)2-D3 (VDR)
1
424
ДНК
Лиганд
Нарушения, связанные с мутациями ядерных рецепторов
Большинство мутаций (замены аминокислот,

Рецепторы арильных углеводородов
Рецепторы арильных углеводородов
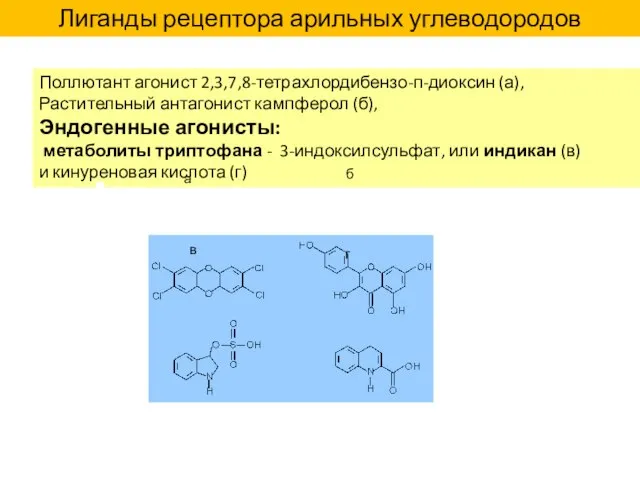
Поллютант агонист 2,3,7,8-тетрахлордибензо-п-диоксин (а),
Растительный антагонист кампферол (б),
Эндогенные агонисты:
метаболиты триптофана
Поллютант агонист 2,3,7,8-тетрахлордибензо-п-диоксин (а),
Растительный антагонист кампферол (б),
Эндогенные агонисты:
метаболиты триптофана
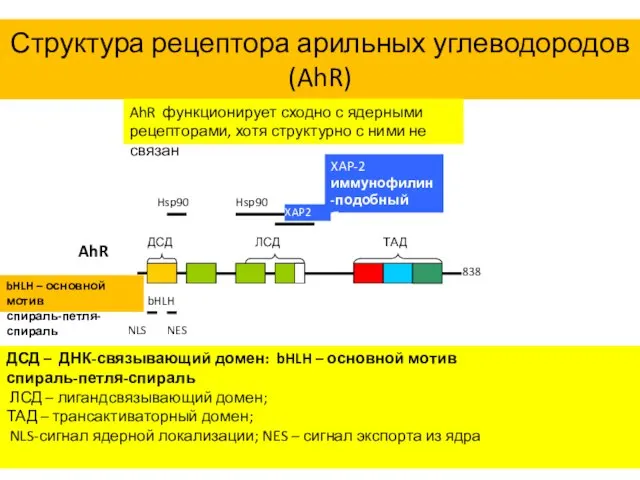
Структура рецептора арильных углеводородов (AhR)
AhR функционирует сходно с ядерными рецепторами, хотя
Структура рецептора арильных углеводородов (AhR)
AhR функционирует сходно с ядерными рецепторами, хотя

Роль комплексирования рецептора арильных углеводородов с белками теплового шока
В цитоплазме гетеродимеры:
Роль комплексирования рецептора арильных углеводородов с белками теплового шока
В цитоплазме гетеродимеры:
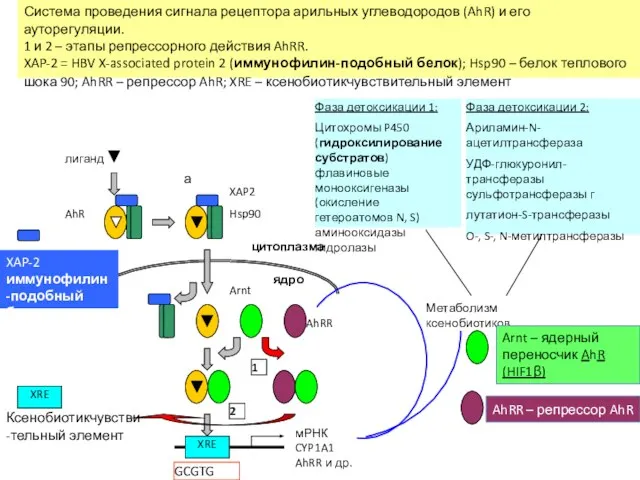
XRE
мРНК
CYP1A1
AhRR и др.
Hsp90
XAP2
AhR
лиганд
Метаболизм ксенобиотиков
AhRR
Arnt
ядро
цитоплазма
1
2
Система проведения сигнала рецептора арильных углеводородов (AhR)
XRE
мРНК
CYP1A1
AhRR и др.
Hsp90
XAP2
AhR
лиганд
Метаболизм ксенобиотиков
AhRR
Arnt
ядро
цитоплазма
1
2
Система проведения сигнала рецептора арильных углеводородов (AhR)
![Участие AhR в активации канцерогенов Бензо[a]пирен – компонент табачного дыма, выхлопных газов](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/517329/slide-29.jpg)
Участие AhR в активации канцерогенов
Бензо[a]пирен – компонент табачного дыма, выхлопных газов
Участие AhR в активации канцерогенов
Бензо[a]пирен – компонент табачного дыма, выхлопных газов

AhR и репродуктивная функция
Активация AhR ведет к ускоренной инактивации половых гормонов
AhR и репродуктивная функция
Активация AhR ведет к ускоренной инактивации половых гормонов

Сенсор электрофильных ксенобиотиков и окислительного стресса
белок Keap1
Сенсор электрофильных ксенобиотиков и окислительного стресса
белок Keap1
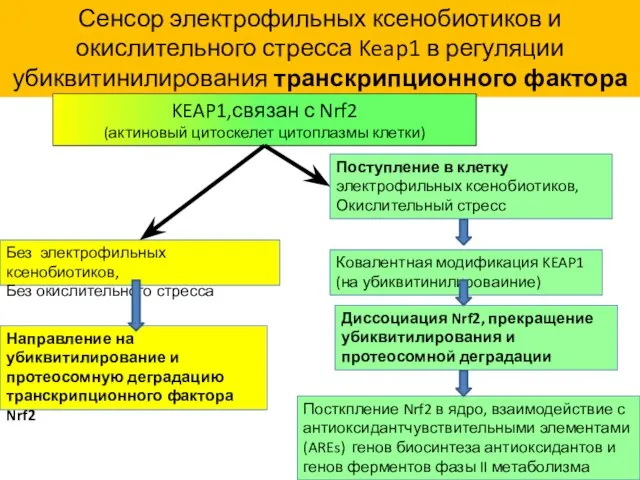
Сенсор электрофильных ксенобиотиков и окислительного стресса Keap1 в регуляции убиквитинилирования транскрипционного
Сенсор электрофильных ксенобиотиков и окислительного стресса Keap1 в регуляции убиквитинилирования транскрипционного
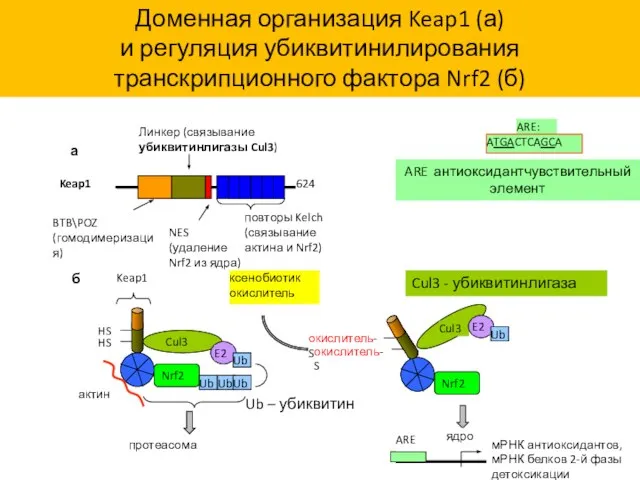
624
повторы Kelch (связывание актина и Nrf2)
NES (удаление Nrf2 из ядра)
Линкер (связывание
624
повторы Kelch (связывание актина и Nrf2)
NES (удаление Nrf2 из ядра)
Линкер (связывание

Мембранные рецепторы группы Notch,
активируемые зависимым от лиганда протеолизом
Контактный способ
Мембранные рецепторы группы Notch,
активируемые зависимым от лиганда протеолизом
Контактный способ

Лиганды рецептора Notch
Интегральные трансмембранные белки с одним трансмембранным доменом:
Delta-like-1, Delta-like-3, Delta-like-4,
Лиганды рецептора Notch
Интегральные трансмембранные белки с одним трансмембранным доменом:
Delta-like-1, Delta-like-3, Delta-like-4,
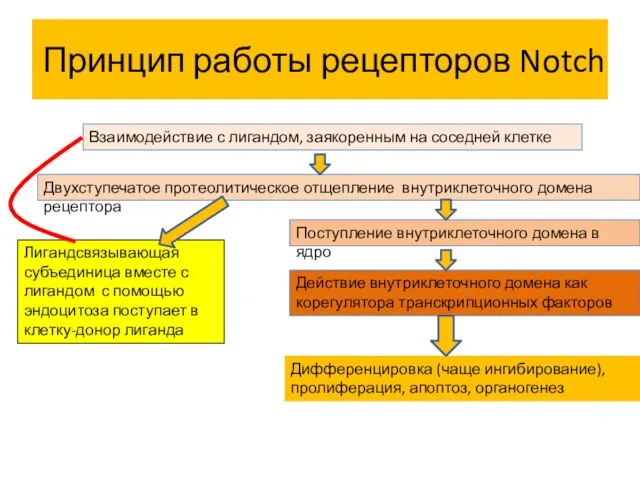
Принцип работы рецепторов Notch
Взаимодействие с лигандом, заякоренным на соседней клетке
Двухступечатое протеолитическое
Принцип работы рецепторов Notch
Взаимодействие с лигандом, заякоренным на соседней клетке
Двухступечатое протеолитическое
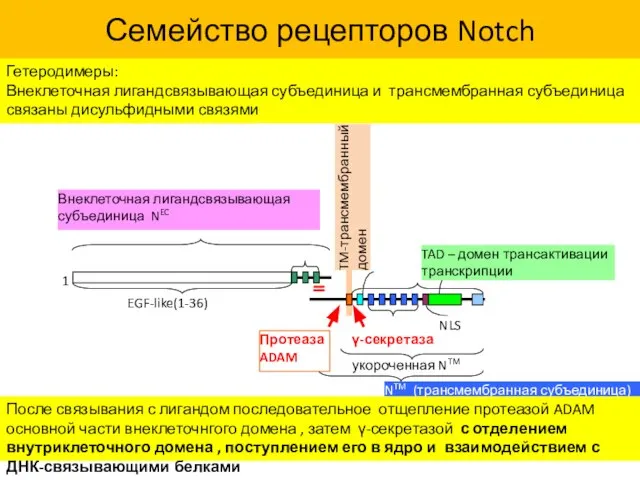
Семейство рецепторов Notch
Гетеродимеры:
Внеклеточная лигандсвязывающая субъединица и трансмембранная субъединица связаны дисульфидными связями
Семейство рецепторов Notch
Гетеродимеры:
Внеклеточная лигандсвязывающая субъединица и трансмембранная субъединица связаны дисульфидными связями
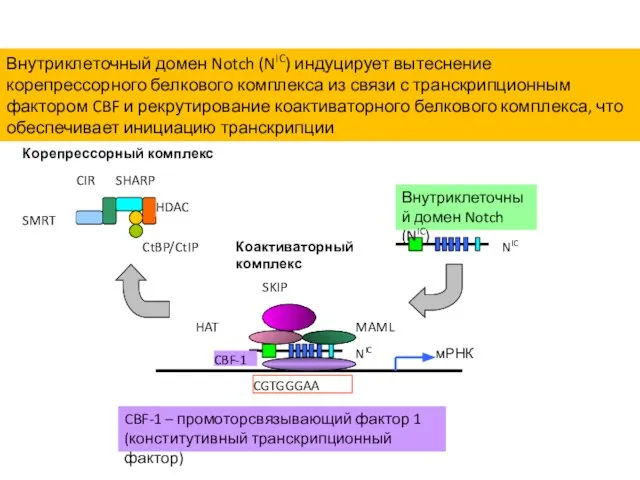
Внутриклеточный домен Notch (NIC) индуцирует вытеснение корепрессорного белкового комплекса из связи
Внутриклеточный домен Notch (NIC) индуцирует вытеснение корепрессорного белкового комплекса из связи
 Невербальное общение
Невербальное общение Дитячі інфекційні захворювання
Дитячі інфекційні захворювання Технология обработки пищевых продуктов
Технология обработки пищевых продуктов Дәлелдерге негізделген қоғамдық денсаулық сақтау аспектілеріндегі денсаулықты нығайту
Дәлелдерге негізделген қоғамдық денсаулық сақтау аспектілеріндегі денсаулықты нығайту Основы гражданской обороны
Основы гражданской обороны Аллергия (гиперчувствительность)
Аллергия (гиперчувствительность) Санатории и курорты Израиля
Санатории и курорты Израиля Понятие о травмах
Понятие о травмах 5 игр на развитие памяти
5 игр на развитие памяти Асептика и антисептика. История вопроса. Виды антисептики
Асептика и антисептика. История вопроса. Виды антисептики Уход за кожей. Профилактика опрелостей и пролежней
Уход за кожей. Профилактика опрелостей и пролежней Вспомогательные репродуктивные технологии
Вспомогательные репродуктивные технологии Фитотерапия сахарного диабета
Фитотерапия сахарного диабета Роль медицинской сестры в профилактике инфекций, связанных с оказанием медицинской помощи
Роль медицинской сестры в профилактике инфекций, связанных с оказанием медицинской помощи Внутренняя среда организма: кровь, тканевая жидкость, лимфа. Основные функции крови. Кровеносная система
Внутренняя среда организма: кровь, тканевая жидкость, лимфа. Основные функции крови. Кровеносная система Героин. Депрессант
Героин. Депрессант Психология семейных отношений
Психология семейных отношений Основы гигиены мед персонала. ВБИ. Санэпидрежим. Гигиена рук. Асептика
Основы гигиены мед персонала. ВБИ. Санэпидрежим. Гигиена рук. Асептика Draw and Label the four phases of Mitosis in a mother cell containing four chromosomes
Draw and Label the four phases of Mitosis in a mother cell containing four chromosomes Визуальная диагностика респираторного дистресс-синдрома
Визуальная диагностика респираторного дистресс-синдрома Гормональная контрацепция
Гормональная контрацепция Na+. Гипернатрийемия
Na+. Гипернатрийемия Клиническая фармакология противовирусных средств
Клиническая фармакология противовирусных средств Типы конфликтов
Типы конфликтов Поликлиника ЦКБ. Выполнение повестки
Поликлиника ЦКБ. Выполнение повестки Физические упражнения, как средство реабилитации после травм суставов
Физические упражнения, как средство реабилитации после травм суставов Особенности консультирования агрессивных клиентов
Особенности консультирования агрессивных клиентов Проектор знаков. Описание тестов
Проектор знаков. Описание тестов