- Строение мышечных веретен и сухожильных органов

Содержание
- 2. Аференты Iа – аферентные волокна первичных мышечных веретён от центральных частей, являются датчиками изменения длины мышцы,
- 3. Аференты Ib – афференты от рецепторов сухожильно-мышечного соединения (аппарат Гольджи), относятся к рецепторам растяжения, но регистрируют
- 4. γ-мотонейроны – эфферентные γ-волкна (Аγ-волокна) иннервируют интрафузальные мышечные веретёна. 2 типа волокон: – Аγ-волокна с концевой
- 6. Схема состояния и характера импульсации мышечных веретен и сухожильных органов Гольджи в покое (А), при пассивном
- 7. Рефлекторная дуга моносинаптического рефлекса растяжения
- 8. Облегчение Т–рефлексов. Если коленный и другие Т–рефлексы нижней конечности ослаблены, их можно усилить, если больной, сцепив
- 9. Дуги рефлекса растяжения и реципрокного торможения мышц–антагонистов С–мотонейроны сгибателей коленного сустава; Р–мотонейроны разгибателей коленного сустава.
- 10. Реципрокное торможение агонистов афферентами Iа. Iа-волокна образуют не только возбуждающие связи с одноимёнными α-МН в составе
- 11. Растяжение мышечных веретён разгибателя усиливают возбуждение α-МН разгибателя (первый рефлекс) и торможение α-МН сгибателя (второй рефлекс).
- 12. g–Петля (А, красный цвет) и влияние фузимоторных волокон на частоту импульсации первичного окончания мышечного веретена (Б,
- 13. γ-петля Мышечные веретёна могут вызвать сокращение экстрафузальных волокон в 2-х случаях: 1. При их растяжении. 2.
- 14. ТОНИЧЕСКАЯ ФУНКЦИЯ СПИННОГО МОЗГА. ГАММА-МОТОРНАЯ ПЕТЛЯ
- 15. Коактивация альфа– и гамма–мотонейронов при движениях. α– и γ–мотонейроны возбуждаются одновременно; однако начало импульсации афферентов мышечных
- 16. Выводы по γ-петле Главное назначение γ-иннервации состоит в том, чтобы предотвратить во время сокращения экстрафузальных волокон
- 17. Функции сухожильных органов. Рефлекторная дуга сухожильных органов служит для поддержания постоянства напряжения мышцы. У каждой мышцы
- 18. Внутрисегментарные связи волокон Ib от сухожильных органов Возбуждающая связь волокна Ib (от мышцы–сгибателя) с мотонейроном разгибателя
- 19. Афференты Ib – образуют тормозные связи с одноимёнными α-МН и возбуждающие с α-МН мышц-антагонистов, то есть
- 20. Сгибательные рефлексы Осуществляются при раздражении кожных рецепторов, рецепторов суставов, рецепторов давления и боли скелетных мышц. Например,
- 22. Возвратное торможение Реншоу и пресинаптическое торможение в спинальных двигательных рефлекторных дугах Р–мотонейрон разгибателя, С мотонейрон сгибателя
- 23. Таким образом: на уровне СМ в деятельности каждой мышцы участвуют как минимум 2 системы обратной связи:
- 24. Постуральные рефлексы (от английск. postur—поза, положение), термин, предложенный Шеррингтоном для обозначения рефлексов, обеспечивающих сохранение определенного положения.
- 25. Важнейшие постуральные рефлексы
- 26. Постуральные рефлексы, приведённые в таблице, показывают, что на уровне спинного, продолговатого и среднего мозга осуществляются сложнокоординированные
- 27. Спинальная локомоция (автоматизмы) Основные характеристики локомоции, т.е. перемещения человека или животного в окружающей среде при помощи
- 28. Предполагают, что у человека тоже есть спинальные локомоторные центры. По–видимому, их активация при раздражении кожи проявляется
- 29. СОСАТЕЛЬНЫЙ РЕФЛЕКС
- 30. Шагательный автоматизм Поднимите малыша под мышки ( стопы должны лишь прикасаться к поверхности ). Затем слегка
- 31. Автоматизм Бауэра Когда ребенок лежит на животе, подставьте свою ладонь ему под стопы. Он оттолкнется ножками
- 32. Рефлекс Бабинского Грудничок разводит пальчики на ногах веером лишь тогда, когда вы касаетесь внешней стороны стопы
- 33. Рефлекс Моро Положите малыша на спинку и хлопните ладонями по кровати рядом с его головой. Он
- 34. РЕФЛЕКС ГАЛАНТА Проведите пальцем вдоль позвоночника с правой стороны ( на расстоянии сантиметра от него ).
- 35. Хватательный автоматизм Робинсона. Угасает этот рефлекс к 4-му месяцу. А на смену ему приходит сознательное хватание
- 36. Полная параплегия возникает обычно при разрыве спинного мозга в грудном отделе– от Т2 до Т12 и
- 38. СПИНАЛЬНЫЙ ШОК Развивается в результате прерывания супраспинапьных влияний на мотонейроны спинного мозга при его травмах. Развиваются
- 39. 2. Период появления небольших рефлекторных движений пальцев ног и особенно в первую очередь большого пальца (от
- 40. Механизм гипертонуса мышц после спинального шока После исчезновения спинального шока, тонус мышц, ниже повреждения резко повышается,
- 41. СИНДРОМ БРОУН-СЕКАРА - Односторонний перерыв спинного мозга, характеризуется на стороне поражения двигательным параличем и потерей осязания,
- 42. Зоны поражения у человека при повреждении спинного мозга на уровне грудных сегментов (синдром Броун-Секара)
- 43. Неврологические нарушения после одностороннего перерыва спинного мозга (синдром Броун–Секара). А. Перерыв справа в сегменте Т8 вызывает
- 47. Вегетативные (интероцептивные ) рефлексы спинного мозга
- 48. Висцеромоторные рефлексы Проявляются в двигательных реакциях мышц грудной клетки и брюшной стенки, разгибателей спины при раздражении
- 49. рефлекс Гольца рефлекс Данини-Ашнера раздражение - удар в эпигастральную область, реакция— рефлекторная остановка сердца
- 50. Зоны Захарьина-Геда
- 52. ПАТОЛОГИЧЕСКИЕ РАЗГИБАТЕЛЬНЫЕ РЕФЛЕКСЫ СПИННОГО МОЗГА ПРИ НАРУШЕНИИ СУПРАСПИНАЛЬНЫХ ВЛИЯНИЙ У ЧЕЛОВЕКА - РЕФЛЕКС БАБИНСКОГО - при
- 53. ПАТОЛОГИЧЕСКИЕ СГИБАТЕЛЬНЫЕ РЕФЛЕКСЫ СПИННОГО МОЗГА ПРИ НАРУШЕНИИ СУПРАСПИНАЛЬНЫХ ВЛИЯНИЙ У ЧЕЛОВЕКА ХВАТАТЕЛЬНЫЙ РЕФЛЕКС ЯНУШКЕВИЧА - возникает
- 54. ПАТОЛОГИЧЕСКИЕ РЕФЛЕКСЫ, ВЫЗЫВАЕМЫЕ ПРИ НАНЕСЕНИИ РАЗДРАЖЕНИЯ НА ВЕРХНИЕ КОНЕЧНОСТИ: РЕФЛЕКС РОССОЛИНО - сгибание пальцев кисти в
- 56. Физиология ЦНС физиология ствола мозга
- 57. ФИЗИОЛОГИЯ СТВОЛА МОЗГА
- 58. Продолговатый мозг Варолиев мост Средний мозг
- 59. Функции продолговатого мозга Рефлекторная Проводниковая Тоническая
- 60. СРЕЗ ПРОДОЛГОВАТОГО МОЗГА
- 61. Расположение ядер черепных нервов в стволовой части мозга 1 — добавочный нерв, 2 — подъязычный, 3
- 62. Схема расположения двигательных центров в стволе мозга (продолговатом мозгу, мосте и среднем мозгу). Нарушая связь с
- 63. Ядра продолговатого мозга 1. ЯДРА ЧЕРЕПН0М03Г0ВЫх НЕРВОВ XII пара - ПОДЪЯЗЫЧНОГО НЕРВА - n. hypoglossus -
- 64. Ядра продолговатого мозга - 2 2. ЯДРА ЧЁРЁПНОМОЗГОВЫХ НЕРВОВ: IX пара - языкоглоточного НЕРВА: 1) двигательное
- 65. Ядра продолговатого мозга - 3 ПЕРЕКЛЮЧАЮЩИЕ ЯДРА: Голля и Бурдаха – к таламусу Ретикулярной формации -
- 66. Центры продолговатого мозга 1) Дыхательный 7) Мигания 2) Сердечно-сосудистый 8) Рвоты 3) Слюноотделения 9) Сосания 4)
- 67. РЕФЛЕКСЫ ПРОДОЛГОВАТОГО МОЗГА Жизненно-важные рефлексы Защитные рефлексы Рефлексы пищевого поведения Рефлексы поддержания позы Вегетативные рефлексы Вестибуло-вегетативные
- 68. Постуральные рефлексы СТАТИЧЕСКИЕ - от рецепторов преддверия рефлексы положения рефлексы выпрямления (установочные] СТАТОКИНЕТИЧЕСКИЕ - от рецепторов
- 69. ВЕСТИБУЛЯРНЫЕ ЯДРА ПРОДОЛГОВАТОГО МОЗГА ВЕРХНЕЕ ВЕСТИБУЛЯРНОЕ ЯДРО — ядро Бехтерева НИЖНЕЕ ВЕСТИБУЛЯРНОЕ ЯДРО — ядро Роллера
- 70. ОСНОВНЫЕ ПРОВОДЯЩИЕ ПУТИ ОТ ВЕСТИБУЛЯРНЫХ ЯДЕР ВЕСТИБУЛОСПИНАЛЬНЫЙ ПУТЬ — от ядра Дейтерса передает информацию на альфа
- 71. ОБЩАЯ СХЕМА ОБРАБОТКИ И ПЕРЕДАЧИ ИНФОРМАЦИИ ВЕСТИБУЛЯРНЫМИ ЯДРАМИ 1. От вестибулярного ганглия информация передается в верхнее
- 72. VIII пара ЧМН имеет две веточки: вестибулярная и слуховая — являющихся чувствительными (афферентными). Слуховая ветвь образована
- 73. ПУТИ ПЕРЕДАЧИ ИНФОРМАЦИИ ОТ ВЕСТИБУЛЯРНОГО АНАЛИЗАТОРА Информация от вестибулярного анализатора (VIII пара ЧМН) поступает: от волосковых
- 74. ЯДРО ДЕЙТЕРСА находится под контролем мозжечка (ядро шатра), определяя состояние активности его активности и контролирует тонус
- 75. От вестибулярных ядер продолговатого мозга информация передается к медиальному продольному пучку, который начинается от ядра Даршкевича
- 76. При патологии вестибулярного аппарата или вестибулярных ядер и вестибулярного тракта, возникают такие симптомы, как головокружение (субъективное
- 77. ЯДРА РЕТИКУЛЯРНОЙ ФОРМАЦИИ ЗАДНЕГО МОЗГА (ПМ И МОСТ), участвующих в регуляции двигательной активности: 1. Нейроны продолговатого
- 78. Роль продолговатого мозга в двигательных функциях
- 79. ФИЗИОЛОГИЯ СРЕДНЕГО И ПРОМЕЖУТОЧНОГО МОЗГА
- 80. ПРОДОЛГОВАТЫЙ МОЗГ, МОСТ И СРЕДНИЙ МОЗГ
- 81. Средний мозг
- 82. ЧЕРЕПНО-МОЗГОВЫЕ НЕРВЫ
- 83. Основные ядра среднего мозга Ядра черепномозговых нервов: --- III пара - глазодвигательного нерва --- IV пара
- 84. ДВИГАТЕЛЬНЫЕ ЯДРА СТВОЛА МОЗГА Можно условно разделить на два класса по их влиянию на мышцы: флексорные
- 85. Стволовые рефлексы тонуса мышц, позы и равновесия тела. Осуществляется с вестибулярных, проприоцентивных, тактильных и зрительных рецепторов
- 86. Медиальная разгибательная система Вестибулоспинальный путь – от латеральных вестибулярных ядер, которые получают афферентную импульсацию от рецепторов
- 87. Латеральная сгибательная система Руброспинальный путь – от красного ядра покрышки мозга, возбуждает α- и γ- мотонейроны
- 88. КРАСНОЕ ЯДРО Расположено в области среднего мозга, его нейроны получают информацию из моторной зоны коры больших
- 89. НЕЙРОНЫ КРАСНОГО ЯДРА - Через руброспинальный тракт оказывает активирующее влияние на альфа — мотонейрона мышц сгибателей
- 90. Децеребрационная ригидность. а — кошка с выключенными большими полушариями (путем перевязки сосудов); б — то же
- 91. ЧЕРНАЯ СУБСТАНЦИЯ Получает обильную информацию от базальных ганглиев и участвует в координации движений и эмоций. При
- 92. Статические рефлексы Их ролью яв-ся сохранение естественной позы человека и животных Это тонические рефлексы в покое
- 93. Положения головы, вызывающие минимум (а) и максимум (б) экстензорного тонуса (по Магнусу).
- 94. При положении головы, отметенном 360 и 180°, тонус разгибателей минимален; он доходит до .максимума при положениях
- 95. Статокинетические рефлексы Их роль – восстановление нарушенной позы Это рефлекторное перераспределение мышечного тонуса, возникающее с: Кожных
- 96. Поза лежащей на левом боку обезьяны, у которой кора мозга удалена при сохранении зрительных бугров и
- 97. Кинетические рефлексы Рефлекторное перераспределение мышечного тонуса при ходьбе и беге, вращательном движении на месте, при изменении
- 98. Рефлексы вращения – 1 Возникают с ампул полукружных каналов и включают в перераспределение тонуса мускулатуру тела
- 99. Рефлексы вращения – 2 В реализации рефлексов вращательного движения, кроме зрительного, участвуют волосковые клетки полукружных каналов
- 100. Лифтные рефлексы возникают при подъеме вверх – повышается тонус сгибателей, и животное приседает, а при опускании
- 101. Локомоторная функция ствола мозга лежит в основе совокупности согласованных движений, с помощью которых организм перемещается в
- 103. Бульбарный организм (перерезка между мостом и средним мозгом) – развивается децеребрационная регидность, хотя имеются статические позно-тонические
- 105. Характерные тонические рефлексы четырехлетнего ребенка, лишенного больших полушарий головного мозга
- 106. Сравнение реакций на пассивную (А) и активную разгрузку. Вверху: испытуемый поддерживает рукой груз; угол его локтевого
- 107. Позно-тонические рефлексы у человека ярко выражены в первое полугодие жизни, так у грудных детей: Лабиринтный тонический
- 108. В онтогенезе преобладает тонус мышц сгибателей (спинальный) Определяющий адекватную позу у плода, т.е. максимальную сгибательную активность
- 109. Сторожевой и ориентировочный рефлексы осуществляются с участием четвертохолмия, в ответ на зрительный сигнал (световой) или на
- 110. Роль ЧМН ствола С участием ядер V, IX, X, XI и XII пар осуществляется произвольные движения
- 111. Проводниковая функция ствола мозга Восходящие (афферентные) пути являются частью проводникового отдела анализаторов передающих информацию в сенсорную
- 112. Медиальная петля Преимущественно образуется из аксонов нейронов ядер: 1)Тонкого (Голля) ядра 2) Клиновидного (Бурдаха) ядра (1+2
- 113. Латеральная петля (слуховая петля) Сформирована аксонами он нейронов ядер: Трапецевидного тела Верхней оливы моста Входящих в
- 114. Неспецифическая (экстралемнисковая) – преимущественно волокна: Бокового спиноталомического пути Спиноретикулярного пути (переключаются в ядрах РФ) Часть волокон
- 115. К афферентной (проводящей) системе ствола также относятся пути передающие сенсорную информацию в кору мозжечка: Дорсальный спиномозжечковый
- 116. Нисходящие проводниковые пути ствола мозга Функционально объединяются: Пирамидный (двигательный) путь – от клеток Беца КБП прецентральной
- 117. Нейрогуморальная функция ствола мозга осуществляется за счет выработки нейронами нейромодуляторов, которые действуют как «регуляторы нейромедиаторов» Норадренергическая
- 118. Серотонинергическая система ствола мозга образована скоплением серотонинергических нейронов, образующих ядра шва, центральное серое вещество и ретикулярные
- 119. Интегративная функция ствола мозга Осуществляется его ретикулярной формацией (РФ), нейроны которой расположены диффузно или в виде
- 120. Ретикулярная формация ствола мозга
- 121. В 1946 г. американский нейрофизиолог Х.Мегоун обнаружил, что РФ ствола мозга имеет непосредственное отношение к регуляции
- 122. Х.Мегоун показал, что раздражение вентрокаудальной части гигантоклеточного ядра продолговатого мозга вызывает ослабление в одинаковой мере всех
- 123. Афферентные и эфферентные связи ретикулярной формации стволовой части мозга
- 124. Основные системы ретикулярной формации АФФЕРЕНТНЫЕ СИСТЕМЫ: от спинного мозга, от мозжечка, от четверохолмия, от коры, от
- 125. Афферентные входы РФ Преимущественно образованы от 3 источников: От температурных и болевых рецепторов по волокнам спиноретикулярного
- 126. Эфферентные выходы РФ проецируются: В спинной мозг по латеральному и медиальному ретикулоспинальным путям. К неспецифическим ядрам
- 127. Интеграция соматических функций: Проявляется в координирующем влиянии РФ, посредством которой отдельные рефлексы ствола мозга объединяются в
- 128. В 1949 г. Х.Мегоун и итальянский нейрофизиолог Дж.Моруцци показали, что раздражение РФ эффективно влияет и на
- 129. Восходящие влияния РФ на большой мозг Может быть активирующим и тормозным, реализуется 5-17% нейронов РФ, формирующими
- 130. Схема облегчающих (+) и тормозящих (-) зон РФ и ее связи с корой и подкоркой ТОРМОЗЯЩИЕ
- 131. Схема восходящих влияний РФ на кору мозга Первичный ответ специфической проекционной зоны Вторичный ответ в зонах
- 132. Активирующее влияние на КБП оказывает ростральная часть РФ Опыт Бренера (1935г.) – перерезка ствола мозга между
- 133. Бельгийский нейрофизиолог Ф.Бремер (1935), проводя перерезку головного мозга на различных уровнях и наблюдая за функциями отделенных
- 134. Первый препарат был назван Бремером cerveau isolee (изолированный передний мозг), а второй encephale isolee (изолированный головной
- 135. У животного с ceryeau isolee остаются связанными с корой головного мозга такие важнейшие афферентные системы, как
- 136. У животного же с encephale isolee реакции полностью сохраняются; такое животное открывает глаза на свет, реагирует
- 137. Однако дальнейшие исследования показали, что для поддержания бодрствующего состояния КБП важно не просто поступление к ней
- 138. У человека в спокойном дремотном состоянии такие колебания имеют частоту 8-10 в секунду и являются довольно
- 139. Когда правильные колебания большой амплитуды сменяются низковольтными, частыми колебаниями, то это, очевидно, говорит о том, что
- 140. В 1940 г. американскими исследователями А.Форбсом и Б.Мориссоном, которые исследовали вызванную электрическую активность коры при различных
- 141. Кроме этого локального ответа, афферентное раздражение вызывает длиннолатентный ответ, возникающий в обширных областях коры мозга. Этот
- 142. То обстоятельство, что вторичные ответы возникают со скрытым периодом, значительно превышающим скрытый период первичного ответа, ясно
- 143. Позже, когда было применено прямое раздражение РФ, было показано, что оно может вызвать ответ такого же
- 144. Реакция пробуждения в ЭЭГ, вызванная переферическим афферентным раздражением (А) или прямым раздражением ретикулярной формации (Б). Момент
- 145. Швейцарский физиолог В. Гесс (1929) впервые показал, что в стволе мозга можно найти точки, при раздражении
- 146. Позже Дж.Моруцци (1941) также обнаружил, что, раздражая некоторые участки РФ заднего мозга, можно у животных вызвать
- 147. Тормозные влияния РФ на КБП оказывает её каудальный отдел, стимуляция его приводит к формированию на корковых
- 148. Нисходящее влияния РФ на СМ облегчающие активность моторных систем СМ, берут начало от всех отделов РФ.
- 149. Аксоны нейронов гигантоклеточного ядра, т.е. основного ядра ретикулярной формации, очень часто делятся Т-образно, и один из
- 150. Схема нисходящих влияний РФ на мотонейроны РФ Ретикуло-спинальные пути Альфа-мотонейрон Гамма-мотонейрон Активация Торможение
- 152. Ретикулярное торможение устойчиво к действию стрихнина. Стрихнин является специфическим ядом, устраняющим постсинаптическое торможение мотонейронов, вызванное импульсацией
- 153. Восходящие влияния носят тонический характер, они легко изменяются гуморальными факторами и высокочувствительны к фармакологическим веществам. Снотворное
- 154. Ретикулярные структуры, регулирующие вегетативные функции, отличаются высокой химической чувствительностью; влияния РФ на двигательные центры также легко
- 155. Промежуточный мозг Состоит из таламической области и гипоталамуса. Таламическая область – включает таламус, коленчатые тела и
- 156. Основные ядра таламуса Специфические ядра переключающие ассоциативные моторные Неспецифические ядра срединные ядра, надколенное ядро, пограничное ядро,
- 157. Проекции специфических (а) и неспецифических (б) ядер таламуса в кору больших полушарий 1 — сенсорные афферентные
- 158. Релейные ядра (специфические) Сенсорные – переключают афферентную импульсацию в сенсорные зоны КБП, перекодируют и обрабатывают информацию
- 160. Ассоциативные системы таламуса ТАЛАМОПАРИЕТАЛЬНАЯ: ОТ ЗАДНЕЛАТЕРАЛЬНОГО ЯДРА И ПОДУШКИ ТАЛАМУСА К ТЕМЕННОЙ И ВИСОЧНОЙ КОРЕ ТАЛАМОФРОНТАЛЬНАЯ:
- 161. Основные функции ассоциативных систем таламуса ТАЛАМОПАРИЕТАЛЬНАЯ СИСТЕМА 1) Центральный аппарат анализа и синтеза обстановочной афферентации, запуска
- 162. Вентральные задние ядра являются главными реле переключения соматосенсорноафферентной системы в составе волокон медиальной петли и примыкающей
- 163. Латеральное коленчатое тело (наружное) Реле для переключения зрительной импульсации в затылочную кору (поле 17), где используется
- 164. Несенсорные релейные ядра - Передние и вентральные Передние ядра - Афферентные – поступают из мамиллярных тел
- 165. Ассоциативные ядра таламуса Относится: - подушка - медиодорсальное ядро - латеральные ядра их функция интегрирующая, в
- 166. ФИЗИОЛОГИЯ МОЗЖЕЧКА
- 167. Масса мозжечка взрослого человека 120 – 160 г. У новорожденного ≈ 20 г., в 3 месяца
- 168. Мозжечок Составляет в среднем 10 % от массы головного мозга Включает в себя более 50 %
- 170. СТРОЕНИЕ МОЗЖЕЧКА
- 172. Схема областей коры мозжечка Дольки обозначены римскими цифрами I – IX, а соответствующие части полушарий п
- 173. Анатомическое и функциональное подразделение мозжечка на: древний (архицеребеллюм) – темно-серая область, старый (палеоцеребеллюм) – заштриховано новый
- 174. Древний мозжечок (вестибулоцеребеллюм) – к нему относится клочково-узелковая доля, имеет наиболее выраженные связи с вестибулярной системой
- 175. Медиальный мозжечок – функционально включает в себя вестибуло- и спиноцеребеллюм, дает проекции к ядру шатра и
- 176. 3 пары ножек мозжечка Средние – соединяют его с варлиевым мостом (самые сильные) Верхние – соединяют
- 177. Фазы последствия удаления мозжечка 1. Фаза раздражения — связана с повреждением мозжечка, У человека — длиться
- 178. Симптомы фазы выпадения функций: 1. атония — снижение тонуса мышц 2. aстения — повышенная утомляемость при
- 179. СОМАТИЧЕСКАЯ ПРОЕКЦИЯ В КОРЕ ЧЕРВЯ И ПОЛУШАРИЯХ МОЗЖЕЧКА
- 181. Кора мозжечка содержит 3 слоя Молекулярный (самый верхний слой коры) – состоит из параллельных волокон, разветвлений
- 182. Нейронная организация коры мозжечка Вверху слева: схема поперечного среза через листок Внизу, главные синаптические контакты в
- 185. Чешский анатом Ян Пуркинье – 1837 г. описал грушевидные нейроны (neuronum piriforme) коры мозжечка, сферическая форма
- 186. Клетки Пуркинье коры мозжечка Образуют средний слой коры, являются главной функциональной единицей, обладают резко выраженной интегративной
- 187. Клетки Пуркинье достигают окончательного развития к 8 годам, поэтому дети не умеют рассчитывать движения, выглядят неуклюжими
- 189. КЛЕТОЧНЫЕ ЭЛЕМЕНТЫ КОРЫ МОЗЖЕЧКА
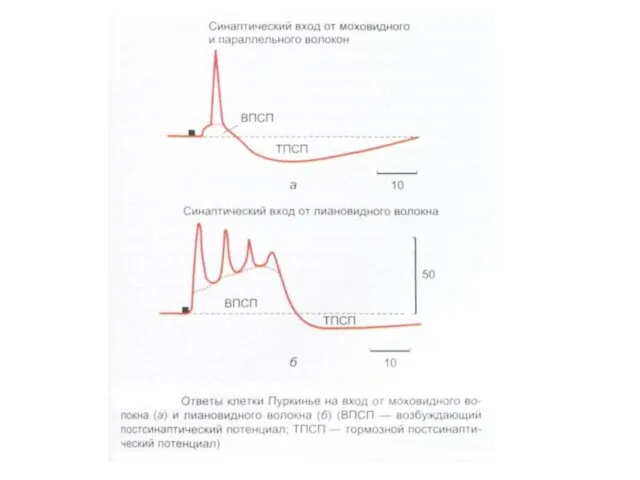
- 191. Мелкие рецептивные поля шести клеток Пуркинье, активность которых регистрировалась двумя микроэлектродами, введенными в дольки V и
- 192. СВЯЗИ КОРЫ МОЗЖЕЧКА АФФЕРЕНТНЫЕ СВЯЗИ - МОХОВИДНЫЕ ВОЛОКНА: от 1) Вестибулярных ядер - вестибулоцеребеллярные тракты 2)
- 193. Лиановидные волокна (лазающие) Почти все являются аксонами нейронов нижней оливы продолговатого мозга, к которым поступает информация
- 194. Три пары ядер мозжечка Располагаются в белом веществе: Шатра (фастигиальное) Промежуточные (пробковидное и шаровидное) Зубчатые Клетки
- 195. Связи ядер мозжечка АФФЕРЕНТНЫЕ СВЯЗИ ВСЕХ ЯДЕР - ОТ КОРЫ МОЗЖЕЧКА - ЗУБЧАТЫЕ ЯДРА: от коры
- 197. ОСНОВНЫЕ ФУНКЦИИ МОЗЖЕЧКА РЕГУЛЯЦИЯ ПОЗЫ И МЫШЕЧНОГО ТОНУСА, РАВНОВЕСИЯ, ПОДДЕРЖИВАЮЩИХ ДВИЖЕНИЙ. КОРРЕКЦИЯ МЕДЛЕННЫХ ЦЕЛЕНАПРАВ-ЛЕННЫХ ДВИЖЕНИЙ И
- 198. Двигательная функция мозжечка 1. Регуляция равновесия и формирования позы - осуществляются преимущественно древним мозжечком и частично
- 199. 4. Программирование целенаправленных движений осуществляется корой нового мозжечка, которая получает информацию из ассоциативных зон КБП через
- 200. Упрощенная функциональная схема медиальных структур мозжечка
- 201. Главные связи медиальной (слева) и латеральной (справа) частей мозжечка Проекции первой идут преимущественно в двигательные центры
- 202. Напряжение мышц разгибателей у собаки, лишенной мозжечка. СНИМКИ 3 и 4 показывают усиление тонуса при раздражении
- 203. Характер движений после удаления мозжечка У животных с удаленным или разрушенным мозжечком наблюдается: атония, астазия, астения,
- 204. Луиджи Лучиани (1840—1919) – в 1891г. показал, что основным комплексом двигательных нарушений мозжечкового происхождения является атаксия,
- 205. ЦИТАТА «Как скульптор избирательно удаляет резцом все лишнее из первоначально бес-форменного камня, так и мозжечок, подавляя
- 206. ПРИЗНАКИ ПОРАЖЕНИЯ МОЗЖЕЧКА ТРИАДА ЛЮЧИАНИ: атония, астазия, астения ТРИАДА ШАРКО: нистагм, тремор, скандированная речь АТАКСИЯ (пьяная
- 207. БАЗАЛЬНЫЕ ГАНГЛИИ
- 209. Базальные ганглии Совокупность парных образований расположенных в конечном мозге, в основании больших полушарий Полосатое тело –
- 211. Эфферентные связи базальных ганглиев Имеют 3 главных выхода: 1. от полосатого тела тормозящие пути идут к
- 213. Афферентные и эфферентные связи базальных ядер 1 — хвостатое ядро, 2 — скорлупа, 3 — черное
- 214. Главные связи базальных ганглиев. Возбуждающие влияния показаны красным, тормозные–черным. БШ – бледный шар (БШн–наружный сегмент, БШв–внутренний
- 215. Роль БГ в регуляции в ДА Отношение между полосатым телом и черным веществом Имеются двухсторонние связи,
- 216. Влияние полосатого тела В эксперименте показано тормозное влияние (медиатор АЦХ) хвостатого ядра на двигательную кору, при
- 217. Функции бледного шара Получает из полосатого тела преимущественно тормозное и частично возбуждающее влияние Бледный шар оказывает
- 218. Активность нейрона внутреннего сегмента бледного шара обезьяны А. Спонтанная, до начала движения. Б, В. Во время
- 219. Активность нейрона компактной части черного вещества обезьяны коррелирует с саккадами глаз. Вертикальной линией отмечен момент появления
- 220. Роль базальных ганглиев в регуляции двигательной активности При циркуляции возбуждения в стриопаллидарной системе (базальные ганглии →
- 221. ФУНКЦИИ БАЗАЛЬНЫХ ГАНГЛИЕВ 1. Центры координации сочетанных двигательных актов 2. Центры сложных безусловных рефлексов и инстинктов
- 222. БЛЕДНЫЙ ШАР ПОЛОСАТОЕ ТЕЛО СУБТАЛАМИЧЕСКОЕ ЯДРО СТВОЛ МОЗГА ТАЛАМУС ДВИГАТЕЛЬНАЯ КОРА АССОЦИАТИВНАЯ КОРА ЧЕРНАЯ СУБСТАНЦИЯ Формирование
- 223. ЭКСТРАПИРАМИДНАЯ СИСТЕМА
- 225. Функциональные петли, проходящие через базальные ганглии ПМК–премоторная кора; ДД К–дополнительная двигательная область коры; ДК двигательная кора;
- 226. ЭФФЕКТЫ ПОРАЖЕНИЯ СТРИОПАЛЛИДАРНОЙ СИСТЕМЫ Поражения хвостатого ядра: гиперкинезы- атетозы и хорея (пляска святого Витта) Поражения паллидум:
- 227. ЭФФЕКТЫ ПОРАЖЕНИЯ СТРИОПАЛЛИДАРНОЙ СИСТЕМЫ Базальные ядра совместно с корой больших полушарий контролируют два важных показателя —
- 228. Болезнь Хантингтона - при потери ГАМК–ергических и холинергических нейронов полосатого тела (характерна хорея — беспорядочные, неконтролируемые
- 229. ЛИМБИЧЕСКАЯ СИСТЕМА
- 230. Лимбическая система Функциональное объединение различных структур конечного, промежуточного и среднего мозга, обеспечивающее эмоционально-мотивационные компоненты поведения и
- 231. Афферентные входы От различных областей головного мозга Через гипоталамус от ретикулярной формации ствола (один из главных
- 232. Лимбическая система
- 233. ОСНОВНЫЕ ФУНКЦИИ ЛИМБИКИ 1. Организация вегетативно-соматических компонентов эмоций 2. Организация кратковременной и долговременной памяти 3. Участие
- 234. Кольцевые нейрональные связи Дают возможность длительной циркуляции (реверберации) возбуждения, что создает условия для сохранения единого функционального
- 235. 2 ЛИМБИЧЕСКИХ КРУГА БОЛЬШОЙ КРУГ ПЕЙПСА: гиппокамп - свод- мамиллярные тела - мамиллярно-таламический пучок Вик-д’Азира -
- 236. Функциональная структура лимбики по МакЛину (1970) 1. Нижний отдел - миндалина и гиппокамп - центры эмоций
- 237. Функции лимбической системы После обработки информации от внешней среды и внутренних органах запускает вегетативные, соматические и
- 238. Физиология гиппокампа
- 240. Физиология гиппокампа Гиппокамп (hippocampus) является основной структурой лимбической системы. Морфологически гиппокамп представлен стереотипно повторяющимися модулями, связанными
- 241. Физиология гиппокампа Значение тета-ритма заключается в том, что он отражает реакцию гиппокампа, а тем самым —
- 242. Физиология миндалевидного тела
- 244. Физиология миндалевидного тела Миндалевидное тело, миндалина — подкорковая структура лимбической системы, расположенная в глубине височной доли
- 245. Физиология миндалевидного тела При искусственной активации миндалины появляются реакции: принюхивания, облизывания, жевания, глотания, саливации, изменения перистальтики
- 246. Влияние миндалин на иерархические отношения в стае До операции После удаления миндалин у Дейва
- 247. Физиология гипоталамуса Гипоталамус (hypothalamus) или подбугорье — отдел головного мозга, расположенный ниже таламуса, или «зрительных бугров»,
- 248. Основные структуры гипоталамуса
- 249. ФУНКЦИИ ГИПОТАЛАМУСА Высший центр регуляции вегетативной нервной системы Высший центр регуляции эндокринных функций Регуляция мотиваций пищевого
- 250. Поведенческие функции гипоталамуса Эффекты стимуляции гипоталамуса Латеральный гипоталамус: жажда, аппетит, увеличение активности организма, ярость, агрессия. Вентромедиальное
- 251. Центры поощрения В различные области мозга обезьяны вводили электроды, и обезьяна, нажимая на рычаг, включала контакты
- 252. Центры поощрения и наказания Личные оценки могут характеризовать ощущения как приятные или неприятные (поощрительные или наказывающие,
- 253. Главные (первичные) центры поощрения располагаются по ходу медиального пучка переднего мозга и особенно в латеральном и
- 254. Центры наказания и реакции избегания расположены в центральном сером веществе, окружающем сильвиев водопровод, в среднем мозге
- 255. Ярость возникает при активации центров наказания. В этом состоянии даже легкая провокация вызывает нападение. Противоположная эмоциональная
- 256. Роль поощрения и наказания в поведении, обучении и памяти Всё или почти все, что делает человек,
- 257. Привыкание. Новые сенсорные стимулы почти всегда возбуждают значительные области коры больших полушарий. Повторение этих же стимулов
- 258. Регуляция вегетативных (висцеральных) функций Осуществляется преимущественно через гипоталамус, куда информация поступает от различных участков лимбической системы
- 259. Роль лимбической системы в обучении Связана с кругом Пейпеца, где главную роль играют гиппокамп и связанные
- 260. ЭМОЦИОНАЛЬНЫЙ МОЗГ
- 261. ЦИТАТА «Является ли эмоция продуктом волшебства или физиологическим процессом, который зависит от анатомического механизма? Я думаю
- 262. Формирование эмоций (пережеваний) Наиболее важная функция лимбической системы. В свою очередь, эмоции, являются субъективным компонентом мотиваций,
- 263. Эмоция — специфическое состояние психической сферы, обусловленная рефлекторной реакцией организма на внешние и внутренние раздражения, характеризующееся
- 264. Эмоциональное возбуждение как результат определенной мотивационной деятельности теснейшим образом связано с удовлетворением трех основных потребностей человека:
- 265. Удовлетворенная потребность вызывает эмоциональное переживание положительного характера и определяет направление поведенческой деятельности. Положительные эмоции, закрепляясь в
- 266. Эмоции, проявляются при недостатке точных сведений и путей достижения жизненных потребностей. Такое представление о природе эмоции
- 267. По Г. И. Косицкому, оценка величины эмоционального напряжения определяется по формуле: СН = Ц (Ин∙Вн∙Эн— Ис∙Вс∙Эс),
- 268. Первая стадия напряжения (CHI) — состояние внимания, мобилизация активности, повышение работоспособности. Эта стадия имеет тренирующее значение,
- 269. Первая стадия напряжения (CHI) — состояние внимания, мобилизация активности, повышение работоспособности. Эта стадия имеет тренирующее значение,
- 270. Третья стадия (СНШ) — астеническая отрицательная реакция, характеризующаяся истощением ресурсов организма и находящая свое психологическое выражение
- 271. В теории функциональной системы по П. К. Анохину нейрофизиологическая природа эмоций связывается с представлениями о функциональной
- 272. Регуляция вегетативных (висцеральных) функций Осуществляется преимущественно через гипоталамус, куда информация поступает от различных участков лимбической системы
- 273. Формирование эмоций (пережеваний) Наиболее важная функция лимбической системы. В свою очередь, эмоции, являются субъективным компонентом мотиваций,
- 274. Роль лимбической системы в обучении Связана с кругом Пейпеца, где главную роль играют гиппокамп и связанные
- 275. ФИЗИОЛОГИЯ КОРЫ БОЛЬШИХ ПОЛУШАРИЙ ГОЛОВНОГО МОЗГА
- 277. Вызванные потенциалы в коре Первичный ответ Вторичный ответ
- 279. Расположение нейронов и нервных волокон в коре больших полушарий. Номера слоёв коры I - VI
- 280. Слои коры больших полушарий 1 слой - верхний молекулярный - ветвления дендритов пирамидных нейронов, редкие горизонтальные
- 281. КБП В I – IV слоях коры происходит восприятие и обработка поступающей информации В II и
- 282. Функциональная организация коры Функциональная единица коры - вертикальная колонка диаметром около 500 мкм - макромодуль Колонка
- 283. Нейронные колонки В коре имеются функциональные объединения нейронов, расположенные в цилиндрике диаметром 0,5-1 мм, включая все
- 284. Колончатая организация корковых нейронов. А. Сагиттальный разрез через постцентральную извилину. Б. Рецептивные поля (ладонь обезьяны) пяти
- 285. Сильно упрощенная схема нейронов, их сетей, афферентных и эфферентных связей в различных слоях коры большого мозга
- 286. ФУНКЦИОНАЛЬНЫЕ ЗОНЫ КОРЫ СЕНСОРНЫЕ ( зрительные, слуховые, кожные и др.) МОТОРНЫЕ ( первичные, вторичные, комплексные) АССОЦИАТИВНЫЕ
- 287. Четыре основные доли коры головного мозга (лобная, височная, теменная и затылочная); вид сбоку. В них расположены
- 288. Локализация функций в КБП Как современная нейробиологическая концепция локализации функций базируется на принципе МНОГОФУНКЦИОНАЛЬНОСТИ корковых полей,
- 289. Цитоархитектоническая карта коры головного мозга человека по Бродману.
- 290. Основные зоны коры мозга
- 291. Локализационная карта Клейста. Наружная поверхность мозга
- 292. Локализационная карта Клейста. Внутренняя поверхность мозга
- 293. Сенсорные зоны КБП В этих зонах проецируется информация от рецепторов всех рецептивных полей (понятия – проекционная
- 294. В сенсорных зонах коры различают Первичные сенсорные поля (проекционные или центральные поля) Их структурная организация завершается
- 295. Представительство вторичных сенсорных зон Их развитие происходит в первые месяцы после рождения, что связано с образованием
- 296. Проекции чувствительности в коре постцентральной извилины Представлены: Головы и верхних отделов туловища – в нижнелатеральных участках
- 297. Проекции частей тела в соматосенсорной зоне коры больших полушарий
- 298. Слуховая сенсорная кора Расположена в верхней височной извилине и представлена: Первичным проекционным полем Вторичной слуховой корой
- 299. Зрительная сенсорная кора представлена в затылочной доли (поля 17 и 18). Первичная проекционная зона (поле 17)
- 300. Вторичная зрительная область (поля 18 и 19) в ней осуществляется тонкий анализ образов - цвета -
- 301. Ассоциативные области коры - включают участки новой КБП, которые расположены рядом с сенсорными и двигательными, но
- 302. Лобная ассоциативная кора (поля 9 – 14) имеет основной источник информации от ассоциативного медиодорсального ядра таламуса
- 303. Теменная ассоциативная кора (5, 7, 39, 40) - получает афферентные вещи от задней группы ассоциативных ядер
- 304. ФУНКЦИИ ЛОБНЫХ ДОЛЕЙ 1. Управление врожденными поведенческими реакциями при помощи накопленного опыта 2. Согласование внешних и
- 305. Двигательные области коры Выделяют первичную , вторичную и дополнительную области Пирамидные нейроны V слоя этих областей
- 306. Первичная моторная зона коры Прецентральная извилина (поле 4) Нейроны управляют функцией мотонейронов мышц лица, туловища и
- 307. Вторичная и дополнительная двигательная кора расположена на латеральной поверхности полушарий, впереди прецентральной извилины, и на медиальной
- 308. Неравномерность представления мускулатуры тела в моторной зоне коры и в гипоталамусе
- 309. Первичные речевые зоны коры Зона Брока Зона Вернике
- 310. Речевые центры
- 311. Разделение латеральной коры головного мозга на функциональные поля по Клайсту
- 312. Локализационная карта Клейста. Внутренняя поверхность мозга
- 313. Асимметрия полушарий мозга
- 314. Цитаты «Мы говорим левым полушарием» Поль Брока, 1865 « Сознание локализуется в левом полушарии, а правое
- 315. Межполушарные различия ЛЕВОЕ ПОЛУШАРИЕ ПРАВОЕ ПОЛУШАРИЕ Лучше узнаются стимулы Словесные Несловесные Легко различимые Трудно различимые Знакомые
- 316. Распознавание лиц левым и правым полушариями больного с расщепленным мозгом
- 317. Общая схема организации двигательной активности
- 318. Структура функциональной системы поведения по П.К.Анохину МОТИ ВАЦИЯ МОТИВАЦИЯ АФФЕРЕНТНЫЙ СИНТЕЗ
- 319. ФУНКЦИОНАЛЬНАЯ СИСТЕМА ЦЕЛЕНАПРАВЛЕННОГО ПОВЕДЕНИЯ АФФЕРЕНТНЫЙ СИНТЕЗ ПРИНЯТИЕ РЕШЕНИЯ ЧТО ДЕЛАТЬ? ЭФФЕРЕНТНЫЙ СИНТЕЗ ПРИНЯТИЕ РЕШЕНИЯ КАК ДЕЛАТЬ?
- 320. Схема организации двигательной системы. В иерархическом порядке представлены связи между центрами нервной системы, участвующими в регуляции
- 321. Схема связей в двигательной системе
- 323. Скачать презентацию
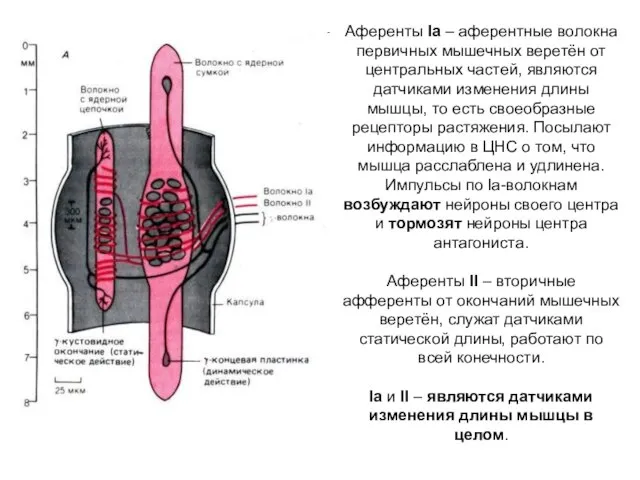
Аференты Iа – аферентные волокна первичных мышечных веретён от центральных частей,
Аференты Iа – аферентные волокна первичных мышечных веретён от центральных частей,
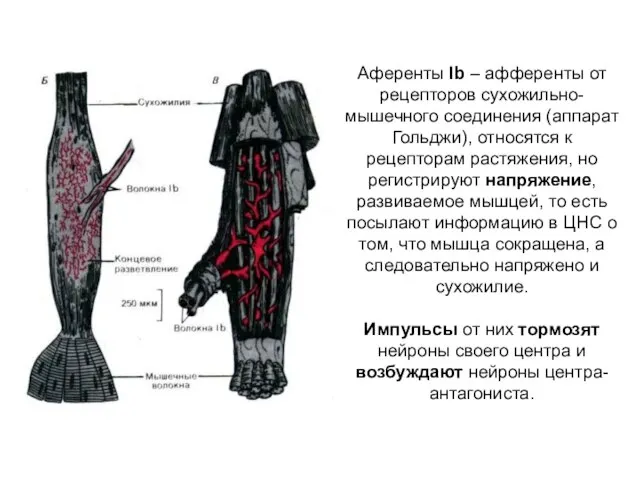
Аференты Ib – афференты от рецепторов сухожильно-мышечного соединения (аппарат Гольджи), относятся
Аференты Ib – афференты от рецепторов сухожильно-мышечного соединения (аппарат Гольджи), относятся

γ-мотонейроны – эфферентные γ-волкна (Аγ-волокна) иннервируют интрафузальные мышечные веретёна.
2 типа
γ-мотонейроны – эфферентные γ-волкна (Аγ-волокна) иннервируют интрафузальные мышечные веретёна. 2 типа

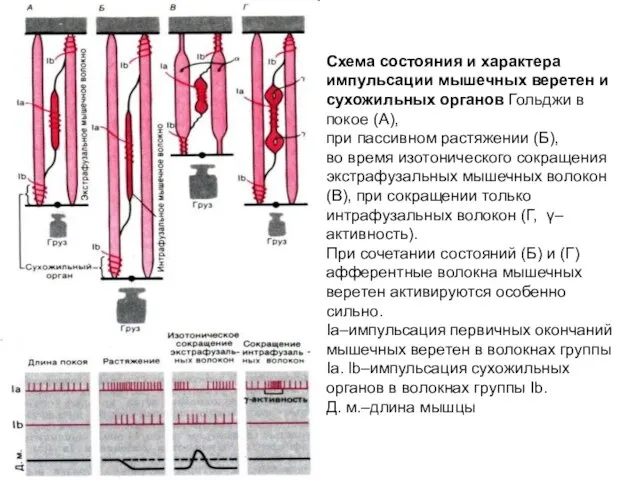
Схема состояния и характера импульсации мышечных веретен и сухожильных органов Гольджи
Схема состояния и характера импульсации мышечных веретен и сухожильных органов Гольджи
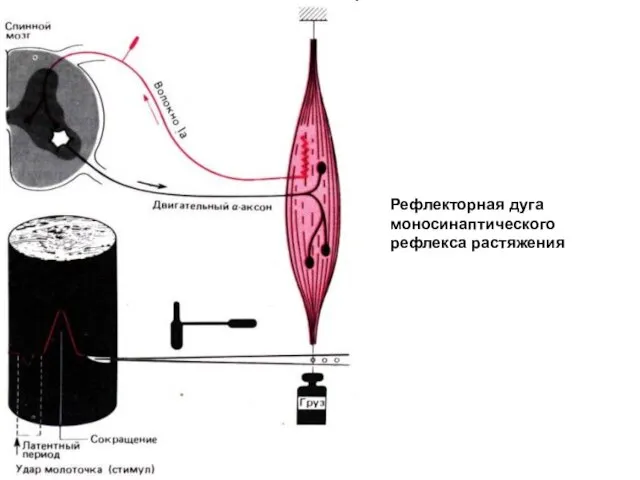
Рефлекторная дуга моносинаптического рефлекса растяжения
Рефлекторная дуга моносинаптического рефлекса растяжения

Облегчение Т–рефлексов.
Если коленный и другие Т–рефлексы нижней конечности ослаблены, их можно
Облегчение Т–рефлексов.
Если коленный и другие Т–рефлексы нижней конечности ослаблены, их можно
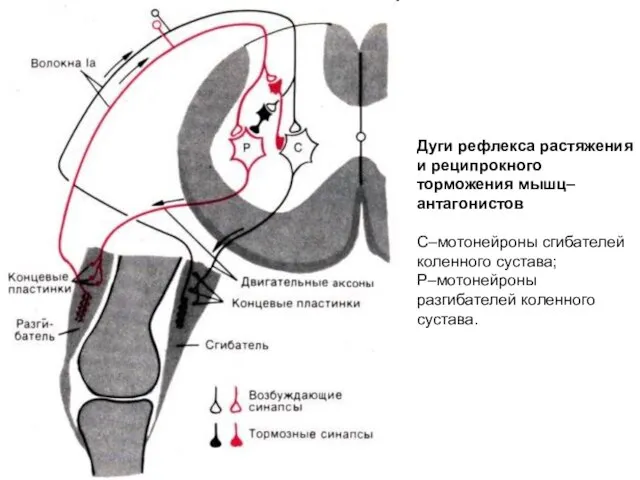
Дуги рефлекса растяжения и реципрокного торможения мышц–антагонистов
С–мотонейроны сгибателей коленного сустава;
Р–мотонейроны
Дуги рефлекса растяжения и реципрокного торможения мышц–антагонистов
С–мотонейроны сгибателей коленного сустава;
Р–мотонейроны
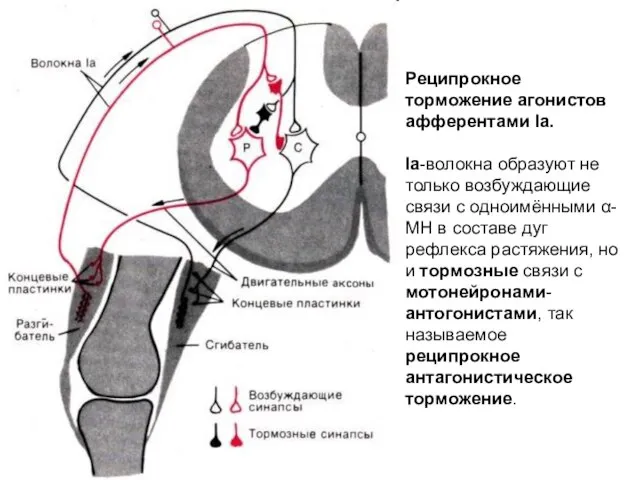
Реципрокное торможение агонистов афферентами Iа.
Iа-волокна образуют не только возбуждающие связи с
Реципрокное торможение агонистов афферентами Iа. Iа-волокна образуют не только возбуждающие связи с
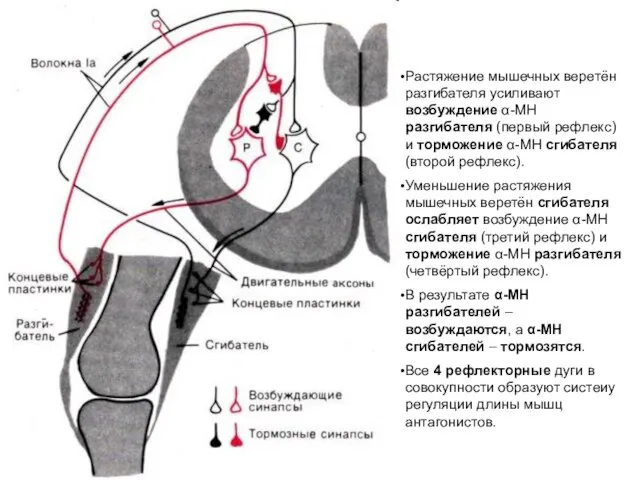
Растяжение мышечных веретён разгибателя усиливают возбуждение α-МН разгибателя (первый рефлекс) и
Растяжение мышечных веретён разгибателя усиливают возбуждение α-МН разгибателя (первый рефлекс) и
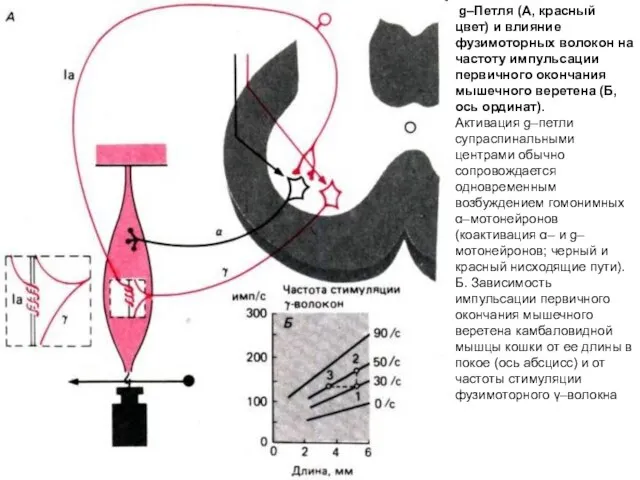
g–Петля (А, красный цвет) и влияние фузимоторных волокон на частоту
g–Петля (А, красный цвет) и влияние фузимоторных волокон на частоту

γ-петля
Мышечные веретёна могут вызвать сокращение экстрафузальных волокон в 2-х случаях:
γ-петля
Мышечные веретёна могут вызвать сокращение экстрафузальных волокон в 2-х случаях:
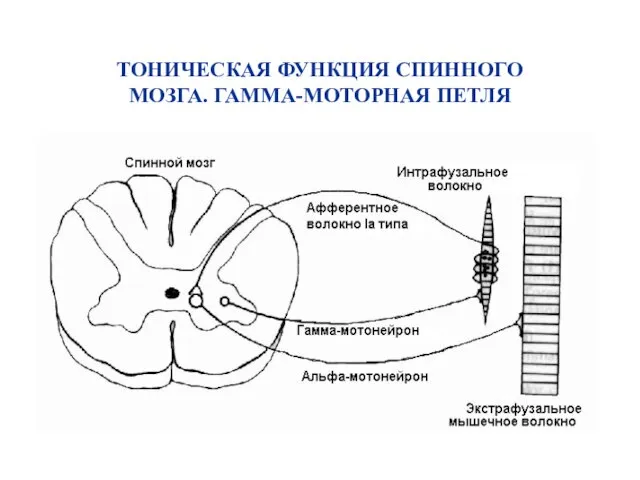
ТОНИЧЕСКАЯ ФУНКЦИЯ СПИННОГО МОЗГА. ГАММА-МОТОРНАЯ ПЕТЛЯ
ТОНИЧЕСКАЯ ФУНКЦИЯ СПИННОГО МОЗГА. ГАММА-МОТОРНАЯ ПЕТЛЯ

Коактивация альфа– и гамма–мотонейронов при движениях.
α– и γ–мотонейроны возбуждаются одновременно; однако начало импульсации афферентов мышечных веретен отстает
Коактивация альфа– и гамма–мотонейронов при движениях.
α– и γ–мотонейроны возбуждаются одновременно; однако начало импульсации афферентов мышечных веретен отстает

Выводы по γ-петле
Главное назначение γ-иннервации состоит в том, чтобы предотвратить
Выводы по γ-петле
Главное назначение γ-иннервации состоит в том, чтобы предотвратить

Функции сухожильных органов.
Рефлекторная дуга сухожильных органов служит для поддержания постоянства напряжения мышцы.
У
Функции сухожильных органов.
Рефлекторная дуга сухожильных органов служит для поддержания постоянства напряжения мышцы.
У
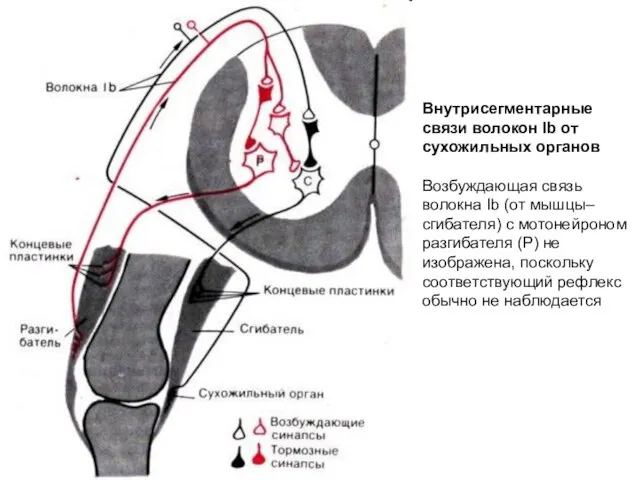
Внутрисегментарные связи волокон Ib от сухожильных органов
Возбуждающая связь волокна Ib (от
Внутрисегментарные связи волокон Ib от сухожильных органов
Возбуждающая связь волокна Ib (от
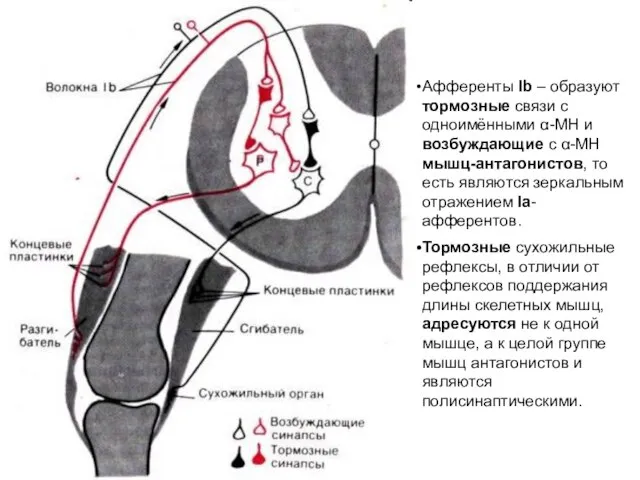
Афференты Ib – образуют тормозные связи с одноимёнными α-МН и возбуждающие
Афференты Ib – образуют тормозные связи с одноимёнными α-МН и возбуждающие

Сгибательные рефлексы
Осуществляются при раздражении кожных рецепторов, рецепторов суставов, рецепторов давления и
Сгибательные рефлексы
Осуществляются при раздражении кожных рецепторов, рецепторов суставов, рецепторов давления и


Возвратное торможение Реншоу и пресинаптическое торможение в спинальных двигательных рефлекторных дугах
Р–мотонейрон
Возвратное торможение Реншоу и пресинаптическое торможение в спинальных двигательных рефлекторных дугах
Р–мотонейрон

Таким образом: на уровне СМ в деятельности каждой мышцы участвуют как
Таким образом: на уровне СМ в деятельности каждой мышцы участвуют как

Постуральные рефлексы
(от английск. postur—поза, положение), термин, предложенный Шеррингтоном для обозначения рефлексов,
Постуральные рефлексы
(от английск. postur—поза, положение), термин, предложенный Шеррингтоном для обозначения рефлексов,

Важнейшие постуральные рефлексы
Важнейшие постуральные рефлексы

Постуральные рефлексы, приведённые в таблице, показывают, что на уровне спинного, продолговатого
Постуральные рефлексы, приведённые в таблице, показывают, что на уровне спинного, продолговатого

Спинальная локомоция
(автоматизмы)
Основные характеристики локомоции, т.е. перемещения человека или животного в окружающей
Спинальная локомоция
(автоматизмы)
Основные характеристики локомоции, т.е. перемещения человека или животного в окружающей

Предполагают, что у человека тоже есть спинальные локомоторные центры.
По–видимому,
По–видимому,

СОСАТЕЛЬНЫЙ РЕФЛЕКС
СОСАТЕЛЬНЫЙ РЕФЛЕКС

Шагательный автоматизм
Поднимите малыша под мышки ( стопы должны лишь прикасаться к
Шагательный автоматизм
Поднимите малыша под мышки ( стопы должны лишь прикасаться к

Автоматизм Бауэра
Когда ребенок лежит на животе, подставьте свою ладонь ему
Автоматизм Бауэра
Когда ребенок лежит на животе, подставьте свою ладонь ему

Рефлекс Бабинского
Грудничок разводит пальчики на ногах веером лишь тогда, когда вы
Рефлекс Бабинского
Грудничок разводит пальчики на ногах веером лишь тогда, когда вы

Рефлекс Моро
Положите малыша на спинку и хлопните ладонями по кровати
Рефлекс Моро
Положите малыша на спинку и хлопните ладонями по кровати

РЕФЛЕКС ГАЛАНТА
Проведите пальцем вдоль позвоночника с правой стороны ( на расстоянии
РЕФЛЕКС ГАЛАНТА
Проведите пальцем вдоль позвоночника с правой стороны ( на расстоянии

Хватательный автоматизм Робинсона.
Угасает этот рефлекс к 4-му месяцу. А на смену ему
Хватательный автоматизм Робинсона.
Угасает этот рефлекс к 4-му месяцу. А на смену ему

Полная параплегия
возникает обычно при разрыве спинного мозга в грудном отделе–
от
Полная параплегия возникает обычно при разрыве спинного мозга в грудном отделе– от


СПИНАЛЬНЫЙ ШОК
Развивается в результате прерывания супраспинапьных влияний на мотонейроны спинного мозга
СПИНАЛЬНЫЙ ШОК
Развивается в результате прерывания супраспинапьных влияний на мотонейроны спинного мозга

2. Период появления небольших рефлекторных движений
пальцев ног и особенно в первую очередь
2. Период появления небольших рефлекторных движений пальцев ног и особенно в первую очередь

Механизм гипертонуса мышц после спинального шока
После исчезновения спинального шока, тонус мышц,
Механизм гипертонуса мышц после спинального шока
После исчезновения спинального шока, тонус мышц,

СИНДРОМ БРОУН-СЕКАРА -
Односторонний перерыв спинного мозга, характеризуется на стороне поражения двигательным
СИНДРОМ БРОУН-СЕКАРА -
Односторонний перерыв спинного мозга, характеризуется на стороне поражения двигательным
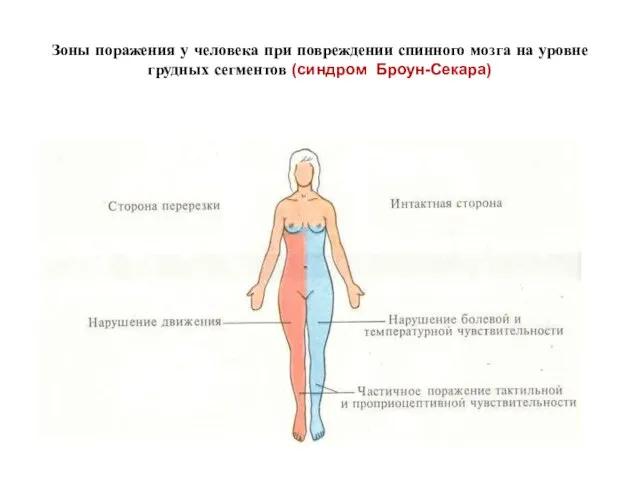
Зоны поражения у человека при повреждении спинного мозга на уровне грудных
Зоны поражения у человека при повреждении спинного мозга на уровне грудных
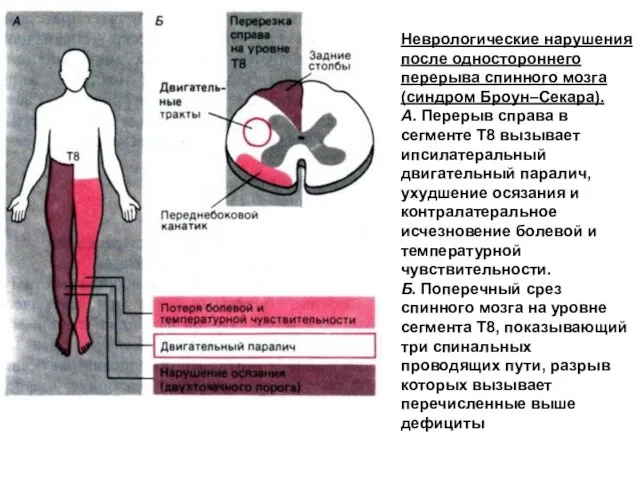
Неврологические нарушения после одностороннего перерыва спинного мозга (синдром Броун–Секара).
А. Перерыв
Неврологические нарушения после одностороннего перерыва спинного мозга (синдром Броун–Секара).
А. Перерыв



Вегетативные (интероцептивные ) рефлексы спинного мозга
Вегетативные (интероцептивные ) рефлексы спинного мозга

Висцеромоторные рефлексы
Проявляются в двигательных реакциях мышц грудной клетки и брюшной стенки,
Висцеромоторные рефлексы Проявляются в двигательных реакциях мышц грудной клетки и брюшной стенки,
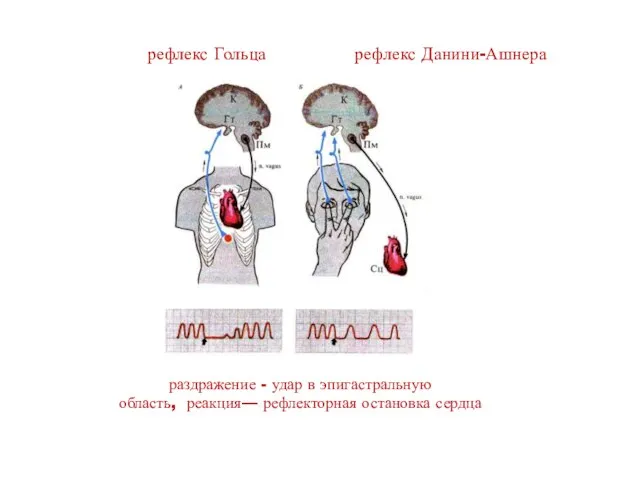
рефлекс Гольца рефлекс Данини-Ашнера
раздражение - удар в эпигастральную область, реакция— рефлекторная остановка сердца
рефлекс Гольца рефлекс Данини-Ашнера
раздражение - удар в эпигастральную область, реакция— рефлекторная остановка сердца

Зоны Захарьина-Геда
Зоны Захарьина-Геда


ПАТОЛОГИЧЕСКИЕ РАЗГИБАТЕЛЬНЫЕ РЕФЛЕКСЫ СПИННОГО МОЗГА ПРИ НАРУШЕНИИ СУПРАСПИНАЛЬНЫХ ВЛИЯНИЙ У ЧЕЛОВЕКА
- РЕФЛЕКС
ПАТОЛОГИЧЕСКИЕ РАЗГИБАТЕЛЬНЫЕ РЕФЛЕКСЫ СПИННОГО МОЗГА ПРИ НАРУШЕНИИ СУПРАСПИНАЛЬНЫХ ВЛИЯНИЙ У ЧЕЛОВЕКА
- РЕФЛЕКС

ПАТОЛОГИЧЕСКИЕ СГИБАТЕЛЬНЫЕ РЕФЛЕКСЫ
СПИННОГО МОЗГА ПРИ НАРУШЕНИИ СУПРАСПИНАЛЬНЫХ
ВЛИЯНИЙ У ЧЕЛОВЕКА
ХВАТАТЕЛЬНЫЙ РЕФЛЕКС ЯНУШКЕВИЧА
ПАТОЛОГИЧЕСКИЕ СГИБАТЕЛЬНЫЕ РЕФЛЕКСЫ
СПИННОГО МОЗГА ПРИ НАРУШЕНИИ СУПРАСПИНАЛЬНЫХ
ВЛИЯНИЙ У ЧЕЛОВЕКА
ХВАТАТЕЛЬНЫЙ РЕФЛЕКС ЯНУШКЕВИЧА

ПАТОЛОГИЧЕСКИЕ РЕФЛЕКСЫ, ВЫЗЫВАЕМЫЕ ПРИ НАНЕСЕНИИ РАЗДРАЖЕНИЯ НА ВЕРХНИЕ КОНЕЧНОСТИ:
РЕФЛЕКС РОССОЛИНО -
ПАТОЛОГИЧЕСКИЕ РЕФЛЕКСЫ, ВЫЗЫВАЕМЫЕ ПРИ НАНЕСЕНИИ РАЗДРАЖЕНИЯ НА ВЕРХНИЕ КОНЕЧНОСТИ:
РЕФЛЕКС РОССОЛИНО -


Физиология ЦНС
физиология ствола мозга
Физиология ЦНС
физиология ствола мозга

ФИЗИОЛОГИЯ СТВОЛА МОЗГА
ФИЗИОЛОГИЯ СТВОЛА МОЗГА

Продолговатый мозг
Варолиев мост
Средний мозг
Продолговатый мозг
Варолиев мост
Средний мозг

Функции продолговатого мозга
Рефлекторная
Проводниковая
Тоническая
Функции продолговатого мозга
Рефлекторная
Проводниковая
Тоническая
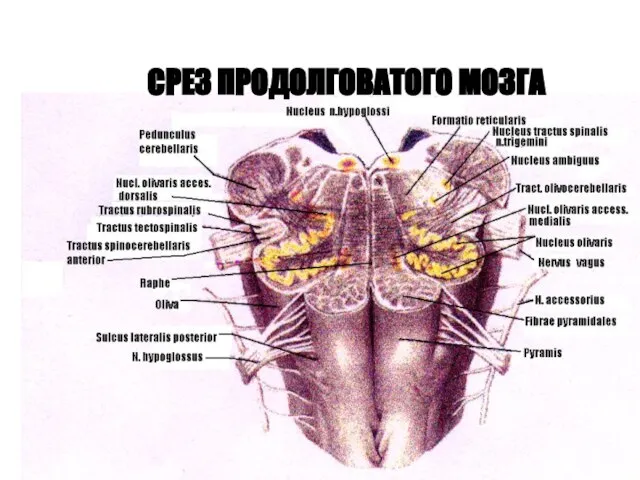
СРЕЗ ПРОДОЛГОВАТОГО МОЗГА
СРЕЗ ПРОДОЛГОВАТОГО МОЗГА
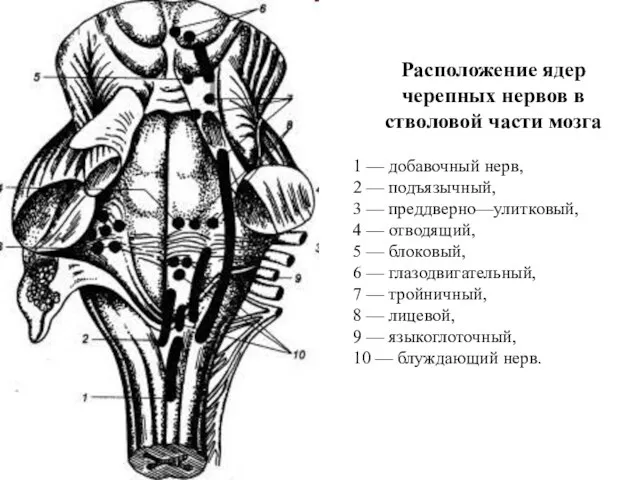
Расположение ядер черепных нервов в стволовой части мозга
1 — добавочный нерв,
Расположение ядер черепных нервов в стволовой части мозга
1 — добавочный нерв,
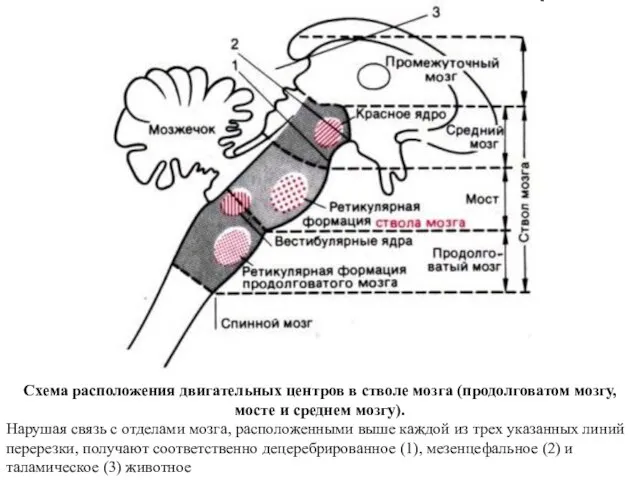
Схема расположения двигательных центров в стволе мозга (продолговатом мозгу, мосте и
Схема расположения двигательных центров в стволе мозга (продолговатом мозгу, мосте и

Ядра продолговатого мозга
1. ЯДРА ЧЕРЕПН0М03Г0ВЫх НЕРВОВ
XII пара - ПОДЪЯЗЫЧНОГО НЕРВА -
Ядра продолговатого мозга
1. ЯДРА ЧЕРЕПН0М03Г0ВЫх НЕРВОВ
XII пара - ПОДЪЯЗЫЧНОГО НЕРВА -

Ядра продолговатого мозга - 2
2. ЯДРА ЧЁРЁПНОМОЗГОВЫХ НЕРВОВ:
IX пара - языкоглоточного
Ядра продолговатого мозга - 2
2. ЯДРА ЧЁРЁПНОМОЗГОВЫХ НЕРВОВ:
IX пара - языкоглоточного

Ядра продолговатого мозга - 3
ПЕРЕКЛЮЧАЮЩИЕ ЯДРА:
Голля и Бурдаха – к таламусу
Ретикулярной
Ядра продолговатого мозга - 3
ПЕРЕКЛЮЧАЮЩИЕ ЯДРА:
Голля и Бурдаха – к таламусу
Ретикулярной

Центры продолговатого мозга
1) Дыхательный 7) Мигания
2) Сердечно-сосудистый 8) Рвоты
3) Слюноотделения 9) Сосания
4) Слезоотделения 10) Жевания
5)
Центры продолговатого мозга
1) Дыхательный 7) Мигания
2) Сердечно-сосудистый 8) Рвоты
3) Слюноотделения 9) Сосания
4) Слезоотделения 10) Жевания
5)

РЕФЛЕКСЫ ПРОДОЛГОВАТОГО МОЗГА
Жизненно-важные рефлексы
Защитные рефлексы
Рефлексы пищевого поведения
Рефлексы поддержания позы
Вегетативные рефлексы
Вестибуло-вегетативные рефлексы
РЕФЛЕКСЫ ПРОДОЛГОВАТОГО МОЗГА
Жизненно-важные рефлексы
Защитные рефлексы
Рефлексы пищевого поведения
Рефлексы поддержания позы
Вегетативные рефлексы
Вестибуло-вегетативные рефлексы

Постуральные рефлексы
СТАТИЧЕСКИЕ - от рецепторов преддверия
рефлексы положения
рефлексы выпрямления (установочные]
СТАТОКИНЕТИЧЕСКИЕ - от
Постуральные рефлексы
СТАТИЧЕСКИЕ - от рецепторов преддверия
рефлексы положения
рефлексы выпрямления (установочные]
СТАТОКИНЕТИЧЕСКИЕ - от

ВЕСТИБУЛЯРНЫЕ ЯДРА ПРОДОЛГОВАТОГО МОЗГА
ВЕРХНЕЕ ВЕСТИБУЛЯРНОЕ ЯДРО — ядро Бехтерева
НИЖНЕЕ ВЕСТИБУЛЯРНОЕ ЯДРО
ВЕСТИБУЛЯРНЫЕ ЯДРА ПРОДОЛГОВАТОГО МОЗГА
ВЕРХНЕЕ ВЕСТИБУЛЯРНОЕ ЯДРО — ядро Бехтерева
НИЖНЕЕ ВЕСТИБУЛЯРНОЕ ЯДРО

ОСНОВНЫЕ ПРОВОДЯЩИЕ ПУТИ ОТ ВЕСТИБУЛЯРНЫХ ЯДЕР
ВЕСТИБУЛОСПИНАЛЬНЫЙ ПУТЬ — от ядра Дейтерса
ОСНОВНЫЕ ПРОВОДЯЩИЕ ПУТИ ОТ ВЕСТИБУЛЯРНЫХ ЯДЕР
ВЕСТИБУЛОСПИНАЛЬНЫЙ ПУТЬ — от ядра Дейтерса

ОБЩАЯ СХЕМА ОБРАБОТКИ И ПЕРЕДАЧИ ИНФОРМАЦИИ ВЕСТИБУЛЯРНЫМИ ЯДРАМИ
1. От вестибулярного ганглия информация
ОБЩАЯ СХЕМА ОБРАБОТКИ И ПЕРЕДАЧИ ИНФОРМАЦИИ ВЕСТИБУЛЯРНЫМИ ЯДРАМИ
1. От вестибулярного ганглия информация

VIII пара ЧМН имеет две веточки: вестибулярная и слуховая — являющихся
VIII пара ЧМН имеет две веточки: вестибулярная и слуховая — являющихся

ПУТИ ПЕРЕДАЧИ ИНФОРМАЦИИ ОТ ВЕСТИБУЛЯРНОГО АНАЛИЗАТОРА
Информация от вестибулярного анализатора (VIII пара
ПУТИ ПЕРЕДАЧИ ИНФОРМАЦИИ ОТ ВЕСТИБУЛЯРНОГО АНАЛИЗАТОРА
Информация от вестибулярного анализатора (VIII пара

ЯДРО ДЕЙТЕРСА
находится под контролем мозжечка (ядро шатра), определяя состояние активности его
ЯДРО ДЕЙТЕРСА
находится под контролем мозжечка (ядро шатра), определяя состояние активности его

От вестибулярных ядер продолговатого мозга информация передается к медиальному продольному пучку,
От вестибулярных ядер продолговатого мозга информация передается к медиальному продольному пучку,

При патологии вестибулярного аппарата или
вестибулярных ядер и вестибулярного тракта,
возникают такие симптомы,
При патологии вестибулярного аппарата или вестибулярных ядер и вестибулярного тракта, возникают такие симптомы,

ЯДРА РЕТИКУЛЯРНОЙ ФОРМАЦИИ ЗАДНЕГО МОЗГА (ПМ И МОСТ),
участвующих в регуляции двигательной
ЯДРА РЕТИКУЛЯРНОЙ ФОРМАЦИИ ЗАДНЕГО МОЗГА (ПМ И МОСТ), участвующих в регуляции двигательной
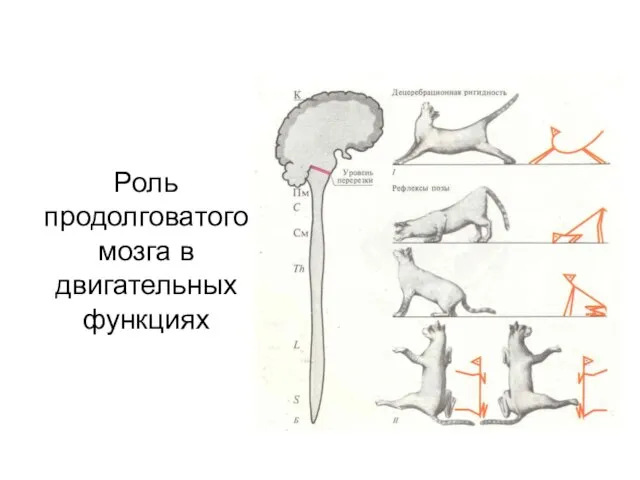
Роль продолговатого мозга в двигательных функциях
Роль продолговатого мозга в двигательных функциях
ФИЗИОЛОГИЯ СРЕДНЕГО И ПРОМЕЖУТОЧНОГО МОЗГА
ФИЗИОЛОГИЯ СРЕДНЕГО И ПРОМЕЖУТОЧНОГО МОЗГА
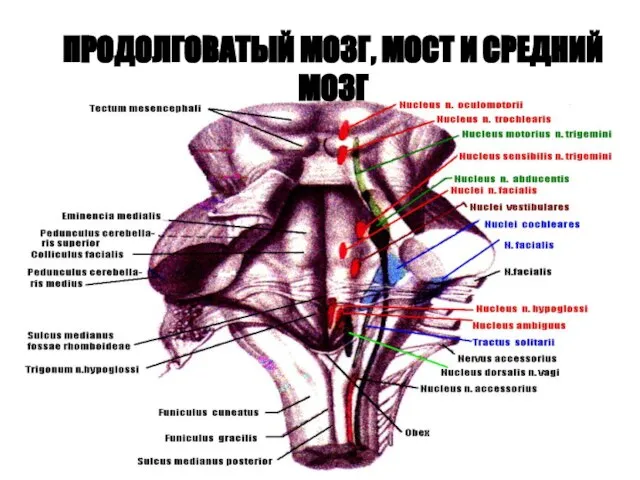
ПРОДОЛГОВАТЫЙ МОЗГ, МОСТ И СРЕДНИЙ МОЗГ
ПРОДОЛГОВАТЫЙ МОЗГ, МОСТ И СРЕДНИЙ МОЗГ
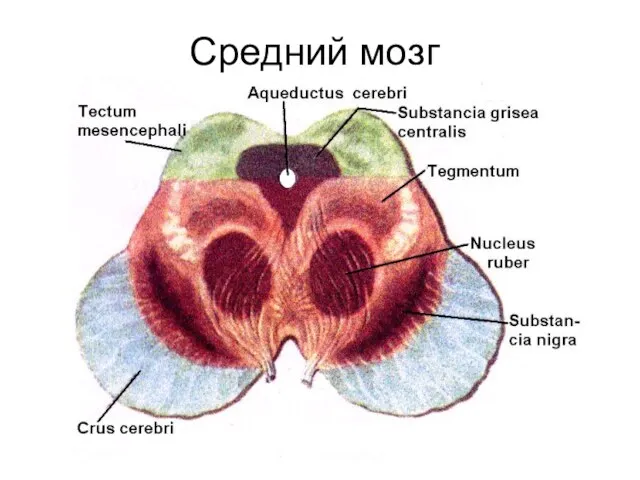
Средний мозг
Средний мозг

ЧЕРЕПНО-МОЗГОВЫЕ НЕРВЫ
ЧЕРЕПНО-МОЗГОВЫЕ НЕРВЫ

Основные ядра среднего мозга
Ядра черепномозговых нервов:
--- III пара - глазодвигательного
Основные ядра среднего мозга
Ядра черепномозговых нервов:
--- III пара - глазодвигательного

ДВИГАТЕЛЬНЫЕ ЯДРА СТВОЛА МОЗГА
Можно условно разделить на два класса по их
ДВИГАТЕЛЬНЫЕ ЯДРА СТВОЛА МОЗГА
Можно условно разделить на два класса по их

Стволовые рефлексы тонуса мышц, позы и равновесия тела.
Осуществляется с вестибулярных, проприоцентивных,
Стволовые рефлексы тонуса мышц, позы и равновесия тела.
Осуществляется с вестибулярных, проприоцентивных,

Медиальная разгибательная система
Вестибулоспинальный путь – от латеральных вестибулярных ядер, которые получают
Медиальная разгибательная система
Вестибулоспинальный путь – от латеральных вестибулярных ядер, которые получают

Латеральная сгибательная система
Руброспинальный путь – от красного ядра покрышки мозга, возбуждает
Латеральная сгибательная система
Руброспинальный путь – от красного ядра покрышки мозга, возбуждает

КРАСНОЕ ЯДРО
Расположено в области среднего мозга, его
нейроны получают информацию из моторной
зоны коры больших полушарий,
промежуточных ядер мозжечка
(пробковидного
КРАСНОЕ ЯДРО
Расположено в области среднего мозга, его
нейроны получают информацию из моторной
зоны коры больших полушарий,
промежуточных ядер мозжечка
(пробковидного

НЕЙРОНЫ КРАСНОГО ЯДРА -
Через руброспинальный тракт оказывает активирующее
влияние на альфа —
НЕЙРОНЫ КРАСНОГО ЯДРА -
Через руброспинальный тракт оказывает активирующее влияние на альфа —

Децеребрационная ригидность.
а — кошка с выключенными большими полушариями (путем перевязки сосудов);
Децеребрационная ригидность.
а — кошка с выключенными большими полушариями (путем перевязки сосудов);

ЧЕРНАЯ СУБСТАНЦИЯ
Получает обильную информацию от базальных ганглиев и участвует в координации
ЧЕРНАЯ СУБСТАНЦИЯ
Получает обильную информацию от базальных ганглиев и участвует в координации

Статические рефлексы
Их ролью яв-ся сохранение естественной позы человека и животных
Это тонические
Статические рефлексы
Их ролью яв-ся сохранение естественной позы человека и животных
Это тонические
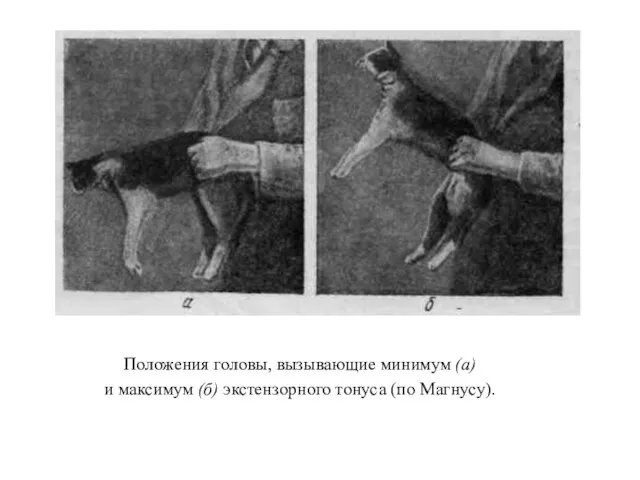
Положения головы, вызывающие минимум (а)
и максимум (б) экстензорного тонуса (по
Положения головы, вызывающие минимум (а)
и максимум (б) экстензорного тонуса (по

При положении головы, отметенном 360 и 180°, тонус разгибателей минимален; он
При положении головы, отметенном 360 и 180°, тонус разгибателей минимален; он

Статокинетические рефлексы
Их роль – восстановление нарушенной позы
Это рефлекторное перераспределение мышечного тонуса,
Статокинетические рефлексы
Их роль – восстановление нарушенной позы
Это рефлекторное перераспределение мышечного тонуса,

Поза лежащей на левом боку обезьяны, у которой кора мозга удалена
Поза лежащей на левом боку обезьяны, у которой кора мозга удалена

Кинетические рефлексы
Рефлекторное перераспределение мышечного тонуса при ходьбе и беге, вращательном движении
Кинетические рефлексы
Рефлекторное перераспределение мышечного тонуса при ходьбе и беге, вращательном движении

Рефлексы вращения – 1
Возникают с ампул полукружных каналов и включают в
Рефлексы вращения – 1
Возникают с ампул полукружных каналов и включают в

Рефлексы вращения – 2
В реализации рефлексов вращательного движения, кроме зрительного, участвуют
Рефлексы вращения – 2
В реализации рефлексов вращательного движения, кроме зрительного, участвуют

Лифтные рефлексы
возникают при подъеме вверх – повышается тонус сгибателей, и
Лифтные рефлексы
возникают при подъеме вверх – повышается тонус сгибателей, и

Локомоторная функция ствола мозга
лежит в основе совокупности согласованных движений, с помощью
Локомоторная функция ствола мозга
лежит в основе совокупности согласованных движений, с помощью


Бульбарный организм (перерезка между мостом и средним мозгом) – развивается децеребрационная
Бульбарный организм (перерезка между мостом и средним мозгом) – развивается децеребрационная

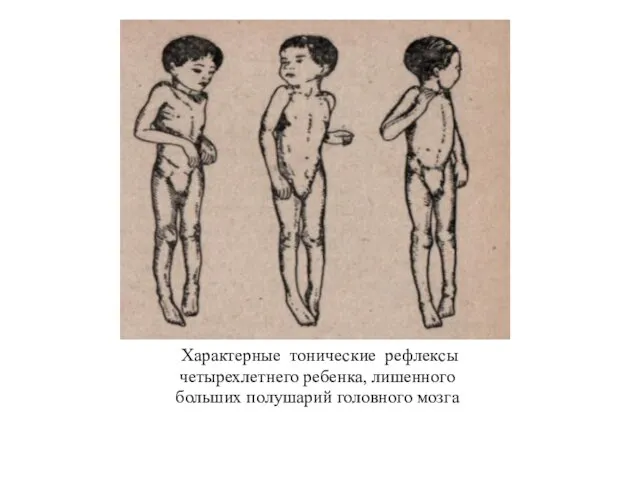
Характерные тонические рефлексы
четырехлетнего ребенка, лишенного
больших полушарий головного мозга
Характерные тонические рефлексы
четырехлетнего ребенка, лишенного
больших полушарий головного мозга

Сравнение реакций на пассивную (А) и активную разгрузку.
Вверху: испытуемый поддерживает
Сравнение реакций на пассивную (А) и активную разгрузку.
Вверху: испытуемый поддерживает

Позно-тонические рефлексы у человека
ярко выражены в первое полугодие жизни, так у
Позно-тонические рефлексы у человека
ярко выражены в первое полугодие жизни, так у

В онтогенезе
преобладает тонус мышц сгибателей (спинальный)
Определяющий адекватную позу у плода,
В онтогенезе
преобладает тонус мышц сгибателей (спинальный)
Определяющий адекватную позу у плода,

Сторожевой и ориентировочный рефлексы
осуществляются с участием четвертохолмия, в ответ на зрительный
Сторожевой и ориентировочный рефлексы
осуществляются с участием четвертохолмия, в ответ на зрительный

Роль ЧМН ствола
С участием ядер V, IX, X, XI и XII
Роль ЧМН ствола
С участием ядер V, IX, X, XI и XII

Проводниковая функция ствола мозга
Восходящие (афферентные) пути являются частью проводникового отдела анализаторов
Проводниковая функция ствола мозга
Восходящие (афферентные) пути являются частью проводникового отдела анализаторов

Медиальная петля
Преимущественно образуется из аксонов нейронов ядер:
1)Тонкого (Голля) ядра
Медиальная петля
Преимущественно образуется из аксонов нейронов ядер:
1)Тонкого (Голля) ядра

Латеральная петля (слуховая петля)
Сформирована аксонами он нейронов ядер:
Трапецевидного тела
Верхней
Латеральная петля (слуховая петля)
Сформирована аксонами он нейронов ядер:
Трапецевидного тела
Верхней

Неспецифическая (экстралемнисковая) – преимущественно волокна:
Бокового спиноталомического пути
Спиноретикулярного
Неспецифическая (экстралемнисковая) – преимущественно волокна:
Бокового спиноталомического пути
Спиноретикулярного

К афферентной (проводящей) системе ствола также относятся пути передающие сенсорную информацию
К афферентной (проводящей) системе ствола также относятся пути передающие сенсорную информацию

Нисходящие проводниковые пути ствола мозга
Функционально объединяются:
Пирамидный (двигательный) путь – от
Нисходящие проводниковые пути ствола мозга
Функционально объединяются:
Пирамидный (двигательный) путь – от

Нейрогуморальная функция ствола мозга
осуществляется за счет выработки нейронами нейромодуляторов, которые действуют
Нейрогуморальная функция ствола мозга
осуществляется за счет выработки нейронами нейромодуляторов, которые действуют

Серотонинергическая система ствола мозга
образована скоплением серотонинергических нейронов, образующих ядра шва, центральное
Серотонинергическая система ствола мозга
образована скоплением серотонинергических нейронов, образующих ядра шва, центральное

Интегративная функция ствола мозга
Осуществляется его ретикулярной формацией (РФ), нейроны которой расположены
Интегративная функция ствола мозга
Осуществляется его ретикулярной формацией (РФ), нейроны которой расположены
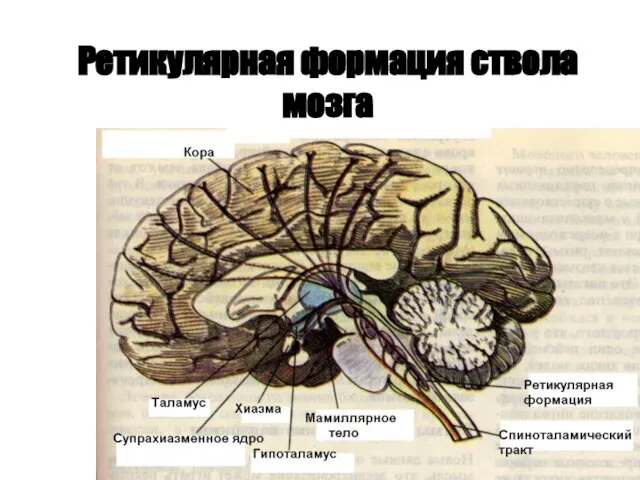
Ретикулярная формация ствола мозга
Ретикулярная формация ствола мозга

В 1946 г. американский нейрофизиолог Х.Мегоун обнаружил, что РФ ствола мозга
В 1946 г. американский нейрофизиолог Х.Мегоун обнаружил, что РФ ствола мозга

Х.Мегоун показал, что раздражение вентрокаудальной части гигантоклеточного ядра продолговатого мозга вызывает
Х.Мегоун показал, что раздражение вентрокаудальной части гигантоклеточного ядра продолговатого мозга вызывает
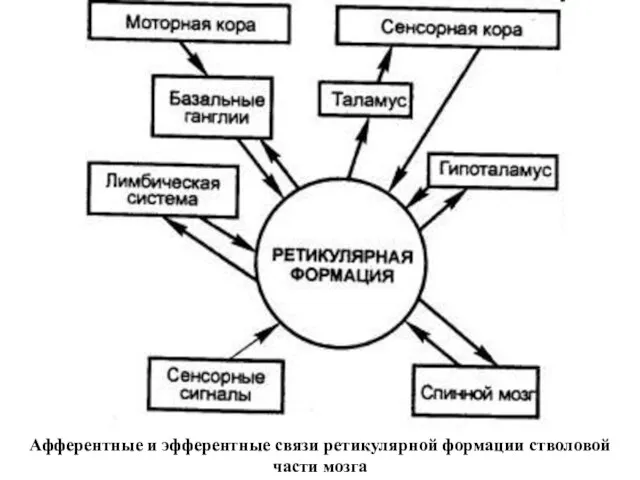
Афферентные и эфферентные связи ретикулярной формации стволовой части мозга
Афферентные и эфферентные связи ретикулярной формации стволовой части мозга

Основные системы ретикулярной формации
АФФЕРЕНТНЫЕ СИСТЕМЫ:
от спинного мозга, от мозжечка, от четверохолмия,
Основные системы ретикулярной формации
АФФЕРЕНТНЫЕ СИСТЕМЫ:
от спинного мозга, от мозжечка, от четверохолмия,

Афферентные входы РФ
Преимущественно образованы от 3 источников:
От температурных и болевых
Афферентные входы РФ
Преимущественно образованы от 3 источников:
От температурных и болевых

Эфферентные выходы РФ проецируются:
В спинной мозг по латеральному и медиальному
Эфферентные выходы РФ проецируются:
В спинной мозг по латеральному и медиальному

Интеграция соматических функций:
Проявляется в координирующем влиянии РФ, посредством которой отдельные рефлексы
Интеграция соматических функций:
Проявляется в координирующем влиянии РФ, посредством которой отдельные рефлексы

В 1949 г. Х.Мегоун и итальянский нейрофизиолог Дж.Моруцци показали, что раздражение
В 1949 г. Х.Мегоун и итальянский нейрофизиолог Дж.Моруцци показали, что раздражение

Восходящие влияния РФ на большой мозг
Может быть активирующим и тормозным, реализуется
Восходящие влияния РФ на большой мозг
Может быть активирующим и тормозным, реализуется
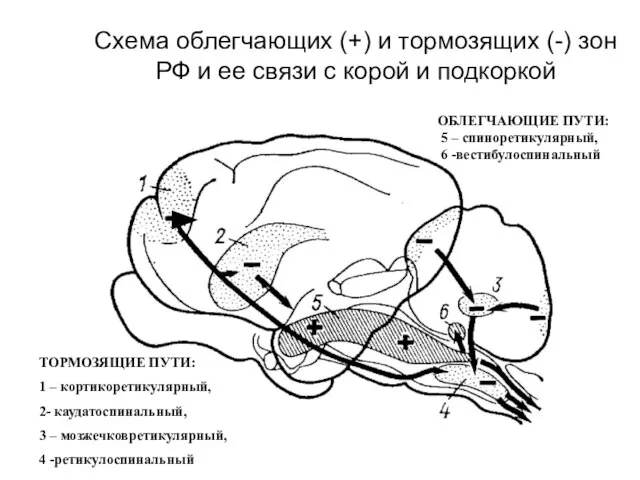
Схема облегчающих (+) и тормозящих (-) зон РФ и ее связи
Схема облегчающих (+) и тормозящих (-) зон РФ и ее связи
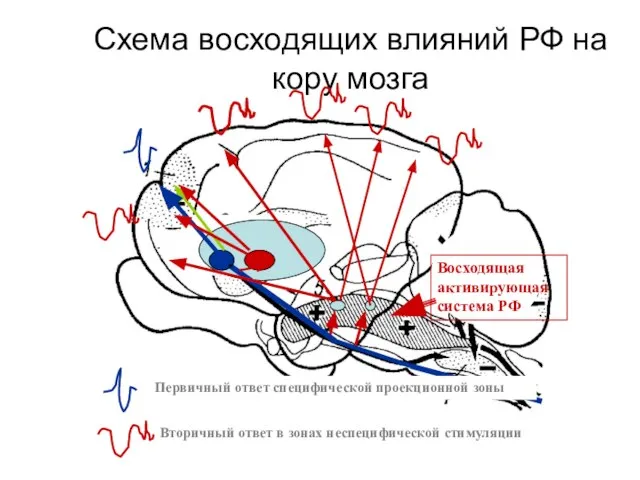
Схема восходящих влияний РФ на кору мозга
Первичный ответ специфической проекционной зоны
Вторичный
Схема восходящих влияний РФ на кору мозга
Первичный ответ специфической проекционной зоны
Вторичный

Активирующее влияние на КБП
оказывает ростральная часть РФ
Опыт Бренера (1935г.) – перерезка
Активирующее влияние на КБП
оказывает ростральная часть РФ
Опыт Бренера (1935г.) – перерезка

Бельгийский нейрофизиолог Ф.Бремер (1935), проводя перерезку головного мозга на различных уровнях
Бельгийский нейрофизиолог Ф.Бремер (1935), проводя перерезку головного мозга на различных уровнях

Первый препарат был назван Бремером cerveau isolee (изолированный передний мозг), а
Первый препарат был назван Бремером cerveau isolee (изолированный передний мозг), а

У животного с ceryeau isolee остаются связанными с корой головного мозга
У животного с ceryeau isolee остаются связанными с корой головного мозга

У животного же с encephale isolee реакции полностью сохраняются; такое животное
У животного же с encephale isolee реакции полностью сохраняются; такое животное

Однако дальнейшие исследования показали, что для поддержания бодрствующего состояния КБП важно
Однако дальнейшие исследования показали, что для поддержания бодрствующего состояния КБП важно

У человека в спокойном дремотном состоянии такие колебания имеют частоту 8-10
У человека в спокойном дремотном состоянии такие колебания имеют частоту 8-10

Когда правильные колебания большой амплитуды сменяются низковольтными, частыми колебаниями, то это,
Когда правильные колебания большой амплитуды сменяются низковольтными, частыми колебаниями, то это,

В 1940 г. американскими исследователями А.Форбсом и Б.Мориссоном, которые исследовали вызванную
В 1940 г. американскими исследователями А.Форбсом и Б.Мориссоном, которые исследовали вызванную

Кроме этого локального ответа, афферентное раздражение вызывает длиннолатентный ответ, возникающий в
Кроме этого локального ответа, афферентное раздражение вызывает длиннолатентный ответ, возникающий в

То обстоятельство, что вторичные ответы возникают со скрытым периодом, значительно превышающим
То обстоятельство, что вторичные ответы возникают со скрытым периодом, значительно превышающим

Позже, когда было применено прямое раздражение РФ, было показано, что оно
Позже, когда было применено прямое раздражение РФ, было показано, что оно
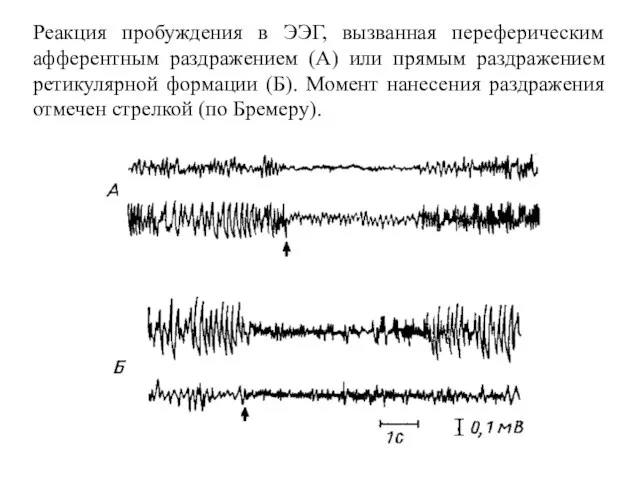
Реакция пробуждения в ЭЭГ, вызванная переферическим афферентным раздражением (А) или прямым
Реакция пробуждения в ЭЭГ, вызванная переферическим афферентным раздражением (А) или прямым

Швейцарский физиолог В. Гесс (1929) впервые показал, что в стволе мозга
Швейцарский физиолог В. Гесс (1929) впервые показал, что в стволе мозга

Позже Дж.Моруцци (1941) также обнаружил, что, раздражая некоторые участки РФ заднего
Позже Дж.Моруцци (1941) также обнаружил, что, раздражая некоторые участки РФ заднего

Тормозные влияния РФ на КБП
оказывает её каудальный отдел, стимуляция его приводит
Тормозные влияния РФ на КБП
оказывает её каудальный отдел, стимуляция его приводит

Нисходящее влияния РФ на СМ
облегчающие активность моторных систем СМ, берут начало
Нисходящее влияния РФ на СМ
облегчающие активность моторных систем СМ, берут начало

Аксоны нейронов гигантоклеточного ядра, т.е. основного ядра ретикулярной формации, очень часто
Аксоны нейронов гигантоклеточного ядра, т.е. основного ядра ретикулярной формации, очень часто
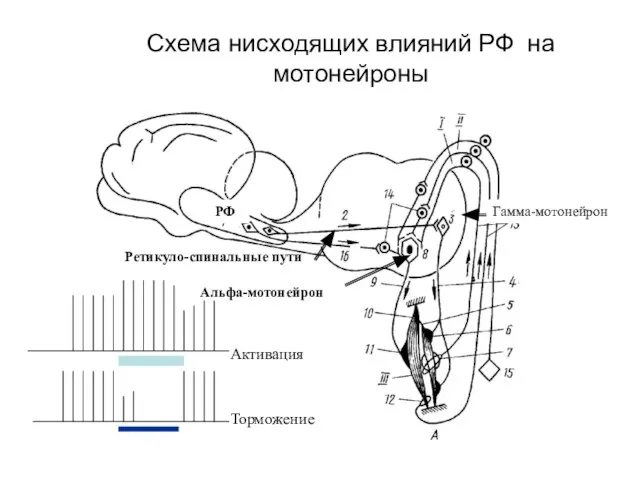
Схема нисходящих влияний РФ на мотонейроны
РФ
Ретикуло-спинальные пути
Альфа-мотонейрон
Гамма-мотонейрон
Активация
Торможение
Схема нисходящих влияний РФ на мотонейроны
РФ
Ретикуло-спинальные пути
Альфа-мотонейрон
Гамма-мотонейрон
Активация
Торможение


Ретикулярное торможение устойчиво к действию стрихнина. Стрихнин является специфическим ядом, устраняющим
Ретикулярное торможение устойчиво к действию стрихнина. Стрихнин является специфическим ядом, устраняющим

Восходящие влияния носят тонический характер, они легко изменяются гуморальными факторами и
Восходящие влияния носят тонический характер, они легко изменяются гуморальными факторами и

Ретикулярные структуры, регулирующие вегетативные функции, отличаются высокой химической чувствительностью; влияния РФ
Ретикулярные структуры, регулирующие вегетативные функции, отличаются высокой химической чувствительностью; влияния РФ

Промежуточный мозг
Состоит из таламической области и гипоталамуса.
Таламическая область – включает
Промежуточный мозг
Состоит из таламической области и гипоталамуса.
Таламическая область – включает

Основные ядра таламуса
Специфические ядра
переключающие ассоциативные моторные
Неспецифические ядра
срединные ядра, надколенное ядро,
Основные ядра таламуса
Специфические ядра
переключающие ассоциативные моторные
Неспецифические ядра
срединные ядра, надколенное ядро,
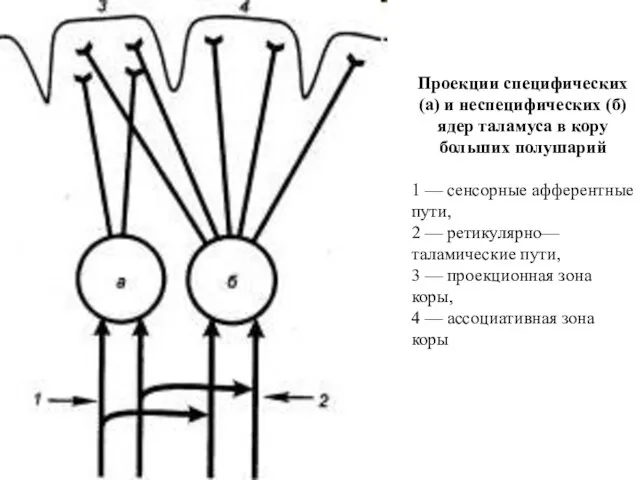
Проекции специфических (а) и неспецифических (б) ядер таламуса в кору больших
Проекции специфических (а) и неспецифических (б) ядер таламуса в кору больших

Релейные ядра (специфические)
Сенсорные – переключают афферентную импульсацию в сенсорные зоны КБП,
Релейные ядра (специфические)
Сенсорные – переключают афферентную импульсацию в сенсорные зоны КБП,


Ассоциативные системы таламуса
ТАЛАМОПАРИЕТАЛЬНАЯ: ОТ ЗАДНЕЛАТЕРАЛЬНОГО ЯДРА И ПОДУШКИ ТАЛАМУСА К ТЕМЕННОЙ
Ассоциативные системы таламуса
ТАЛАМОПАРИЕТАЛЬНАЯ: ОТ ЗАДНЕЛАТЕРАЛЬНОГО ЯДРА И ПОДУШКИ ТАЛАМУСА К ТЕМЕННОЙ

Основные функции ассоциативных систем таламуса
ТАЛАМОПАРИЕТАЛЬНАЯ СИСТЕМА
1) Центральный аппарат анализа и синтеза
Основные функции ассоциативных систем таламуса
ТАЛАМОПАРИЕТАЛЬНАЯ СИСТЕМА
1) Центральный аппарат анализа и синтеза

Вентральные задние ядра
являются главными реле переключения соматосенсорноафферентной системы в составе волокон
Вентральные задние ядра
являются главными реле переключения соматосенсорноафферентной системы в составе волокон

Латеральное коленчатое тело (наружное)
Реле для переключения зрительной импульсации в затылочную кору
Реле для переключения зрительной импульсации в затылочную кору

Несенсорные релейные ядра -
Передние и вентральные
Передние ядра
- Афферентные – поступают
Несенсорные релейные ядра -
Передние и вентральные
Передние ядра
- Афферентные – поступают

Ассоциативные ядра таламуса
Относится:
- подушка
- медиодорсальное ядро
- латеральные ядра
их функция интегрирующая, в
Ассоциативные ядра таламуса
Относится:
- подушка
- медиодорсальное ядро
- латеральные ядра
их функция интегрирующая, в

ФИЗИОЛОГИЯ МОЗЖЕЧКА
ФИЗИОЛОГИЯ МОЗЖЕЧКА

Масса мозжечка взрослого человека 120 – 160 г. У новорожденного ≈
Масса мозжечка взрослого человека 120 – 160 г. У новорожденного ≈

Мозжечок
Составляет в среднем 10 % от массы головного мозга
Включает
Мозжечок
Составляет в среднем 10 % от массы головного мозга
Включает

СТРОЕНИЕ МОЗЖЕЧКА
СТРОЕНИЕ МОЗЖЕЧКА
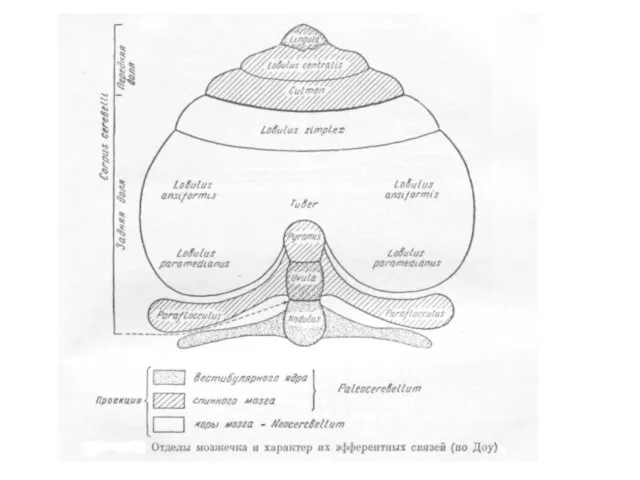
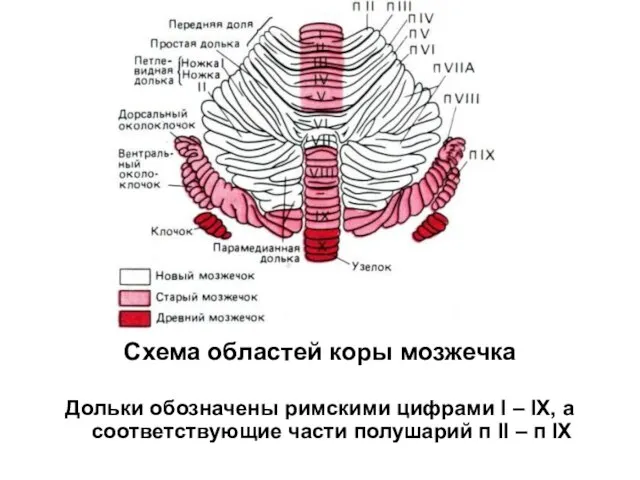
Схема областей коры мозжечка
Дольки обозначены римскими цифрами I – IX, а
Схема областей коры мозжечка
Дольки обозначены римскими цифрами I – IX, а

Анатомическое и функциональное подразделение мозжечка на:
древний (архицеребеллюм) – темно-серая область,
Анатомическое и функциональное подразделение мозжечка на: древний (архицеребеллюм) – темно-серая область,
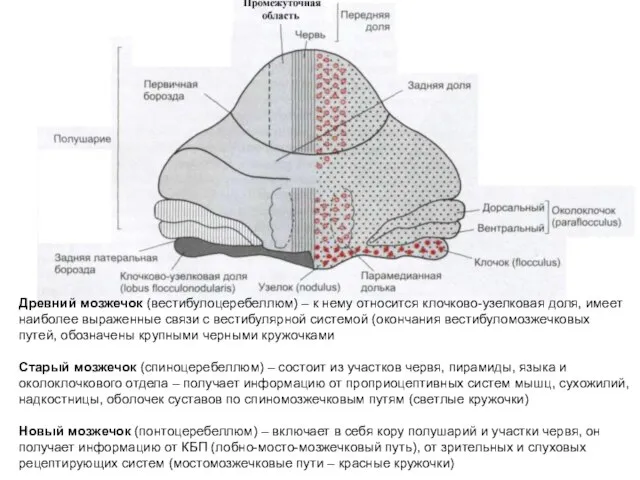
Древний мозжечок (вестибулоцеребеллюм) – к нему относится клочково-узелковая доля, имеет наиболее
Древний мозжечок (вестибулоцеребеллюм) – к нему относится клочково-узелковая доля, имеет наиболее
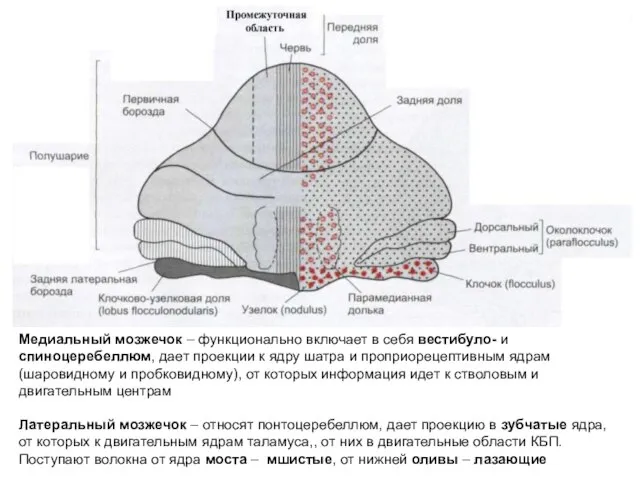
Медиальный мозжечок – функционально включает в себя вестибуло- и спиноцеребеллюм, дает
Медиальный мозжечок – функционально включает в себя вестибуло- и спиноцеребеллюм, дает

3 пары ножек мозжечка
Средние – соединяют его с варлиевым мостом (самые
3 пары ножек мозжечка
Средние – соединяют его с варлиевым мостом (самые

Фазы последствия удаления мозжечка
1. Фаза раздражения — связана с повреждением мозжечка,
У
Фазы последствия удаления мозжечка
1. Фаза раздражения — связана с повреждением мозжечка,
У

Симптомы фазы выпадения функций:
1. атония — снижение тонуса мышц
2. aстения —
Симптомы фазы выпадения функций:
1. атония — снижение тонуса мышц
2. aстения —

СОМАТИЧЕСКАЯ ПРОЕКЦИЯ В КОРЕ ЧЕРВЯ И ПОЛУШАРИЯХ МОЗЖЕЧКА
СОМАТИЧЕСКАЯ ПРОЕКЦИЯ В КОРЕ ЧЕРВЯ И ПОЛУШАРИЯХ МОЗЖЕЧКА


Кора мозжечка содержит 3 слоя
Молекулярный (самый верхний слой коры)
Кора мозжечка содержит 3 слоя Молекулярный (самый верхний слой коры)
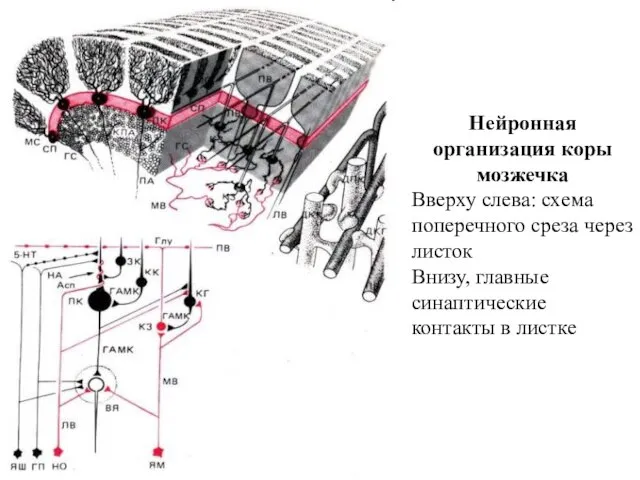
Нейронная организация коры мозжечка
Вверху слева: схема поперечного среза через листок
Внизу,
Нейронная организация коры мозжечка
Вверху слева: схема поперечного среза через листок
Внизу,



Чешский анатом Ян Пуркинье – 1837 г. описал грушевидные нейроны (neuronum
Чешский анатом Ян Пуркинье – 1837 г. описал грушевидные нейроны (neuronum

Клетки Пуркинье коры мозжечка
Образуют средний слой коры, являются главной функциональной единицей,
Клетки Пуркинье коры мозжечка
Образуют средний слой коры, являются главной функциональной единицей,

Клетки Пуркинье достигают окончательного развития к 8 годам, поэтому дети не
Клетки Пуркинье достигают окончательного развития к 8 годам, поэтому дети не
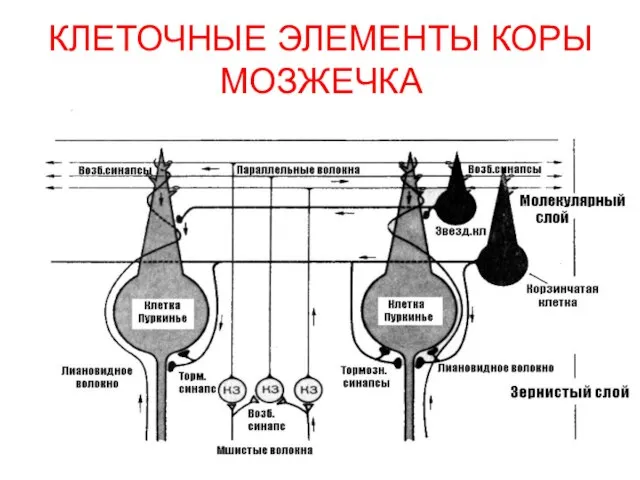
КЛЕТОЧНЫЕ ЭЛЕМЕНТЫ КОРЫ МОЗЖЕЧКА
КЛЕТОЧНЫЕ ЭЛЕМЕНТЫ КОРЫ МОЗЖЕЧКА


Мелкие рецептивные поля шести клеток Пуркинье, активность которых регистрировалась двумя микроэлектродами,
Мелкие рецептивные поля шести клеток Пуркинье, активность которых регистрировалась двумя микроэлектродами,

СВЯЗИ КОРЫ МОЗЖЕЧКА
АФФЕРЕНТНЫЕ СВЯЗИ
- МОХОВИДНЫЕ ВОЛОКНА: от
1) Вестибулярных ядер - вестибулоцеребеллярные
СВЯЗИ КОРЫ МОЗЖЕЧКА
АФФЕРЕНТНЫЕ СВЯЗИ
- МОХОВИДНЫЕ ВОЛОКНА: от
1) Вестибулярных ядер - вестибулоцеребеллярные

Лиановидные волокна (лазающие)
Почти все являются аксонами нейронов нижней оливы продолговатого мозга,
Лиановидные волокна (лазающие)
Почти все являются аксонами нейронов нижней оливы продолговатого мозга,

Три пары ядер мозжечка
Располагаются в белом веществе:
Шатра (фастигиальное)
Промежуточные (пробковидное и
Три пары ядер мозжечка
Располагаются в белом веществе:
Шатра (фастигиальное)
Промежуточные (пробковидное и

Связи ядер мозжечка
АФФЕРЕНТНЫЕ СВЯЗИ ВСЕХ ЯДЕР - ОТ КОРЫ МОЗЖЕЧКА
- ЗУБЧАТЫЕ
Связи ядер мозжечка
АФФЕРЕНТНЫЕ СВЯЗИ ВСЕХ ЯДЕР - ОТ КОРЫ МОЗЖЕЧКА
- ЗУБЧАТЫЕ


ОСНОВНЫЕ ФУНКЦИИ МОЗЖЕЧКА
РЕГУЛЯЦИЯ ПОЗЫ И МЫШЕЧНОГО ТОНУСА, РАВНОВЕСИЯ, ПОДДЕРЖИВАЮЩИХ ДВИЖЕНИЙ.
КОРРЕКЦИЯ МЕДЛЕННЫХ
ОСНОВНЫЕ ФУНКЦИИ МОЗЖЕЧКА
РЕГУЛЯЦИЯ ПОЗЫ И МЫШЕЧНОГО ТОНУСА, РАВНОВЕСИЯ, ПОДДЕРЖИВАЮЩИХ ДВИЖЕНИЙ.
КОРРЕКЦИЯ МЕДЛЕННЫХ

Двигательная функция мозжечка
1. Регуляция равновесия и формирования позы
- осуществляются преимущественно древним
Двигательная функция мозжечка
1. Регуляция равновесия и формирования позы
- осуществляются преимущественно древним

4. Программирование целенаправленных движений
осуществляется корой нового мозжечка, которая получает информацию из
осуществляется корой нового мозжечка, которая получает информацию из
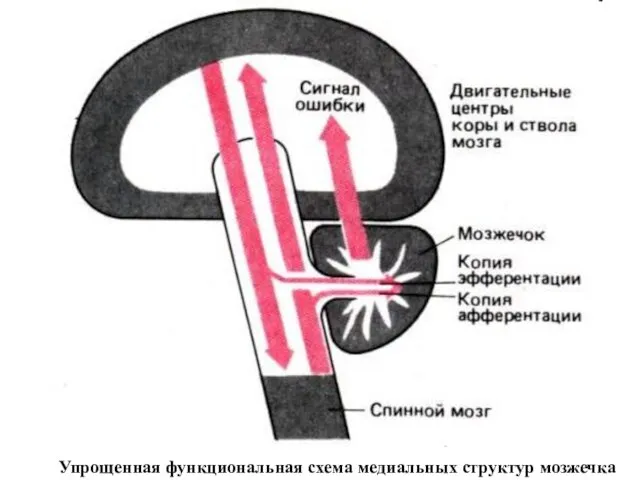
Упрощенная функциональная схема медиальных структур мозжечка
Упрощенная функциональная схема медиальных структур мозжечка
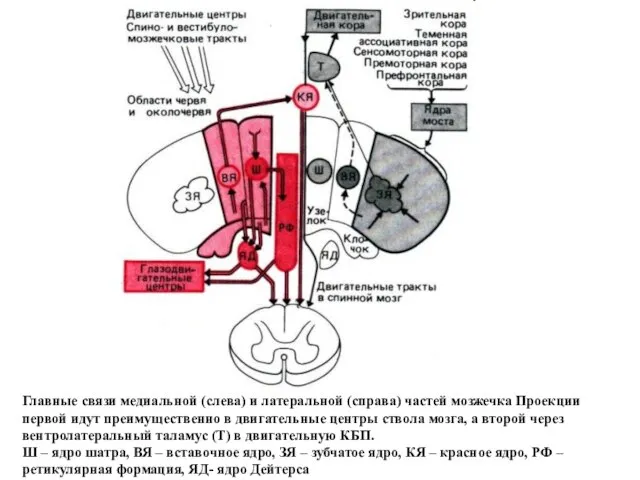
Главные связи медиальной (слева) и латеральной (справа) частей мозжечка Проекции первой
Главные связи медиальной (слева) и латеральной (справа) частей мозжечка Проекции первой

Напряжение мышц разгибателей у собаки, лишенной
мозжечка. СНИМКИ 3 и 4
Напряжение мышц разгибателей у собаки, лишенной
мозжечка. СНИМКИ 3 и 4

Характер движений после удаления мозжечка
У животных с удаленным или разрушенным мозжечком
Характер движений после удаления мозжечка
У животных с удаленным или разрушенным мозжечком

Луиджи Лучиани (1840—1919) – в 1891г. показал, что основным комплексом двигательных
Луиджи Лучиани (1840—1919) – в 1891г. показал, что основным комплексом двигательных

ЦИТАТА
«Как скульптор избирательно удаляет резцом все лишнее из первоначально бес-форменного камня,
ЦИТАТА
«Как скульптор избирательно удаляет резцом все лишнее из первоначально бес-форменного камня,

ПРИЗНАКИ ПОРАЖЕНИЯ МОЗЖЕЧКА
ТРИАДА ЛЮЧИАНИ: атония,
астазия,
астения
ТРИАДА ШАРКО: нистагм,
ПРИЗНАКИ ПОРАЖЕНИЯ МОЗЖЕЧКА
ТРИАДА ЛЮЧИАНИ: атония,
астазия,
астения
ТРИАДА ШАРКО: нистагм,
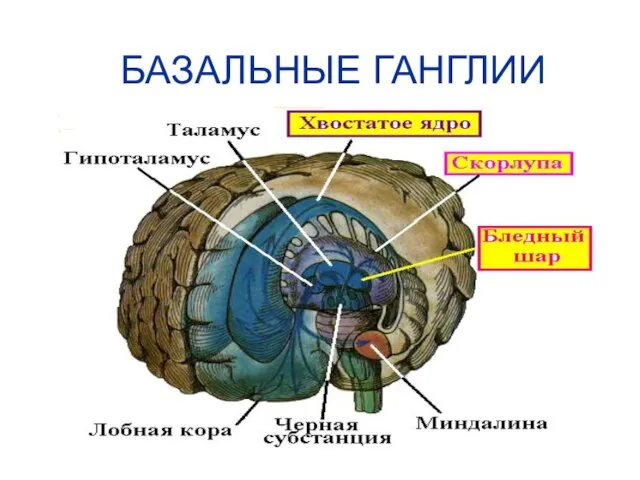
БАЗАЛЬНЫЕ ГАНГЛИИ
БАЗАЛЬНЫЕ ГАНГЛИИ


Базальные ганглии
Совокупность парных образований расположенных в конечном мозге, в основании больших
Базальные ганглии
Совокупность парных образований расположенных в конечном мозге, в основании больших


Эфферентные связи базальных ганглиев
Имеют 3 главных выхода:
1. от полосатого
Эфферентные связи базальных ганглиев
Имеют 3 главных выхода:
1. от полосатого

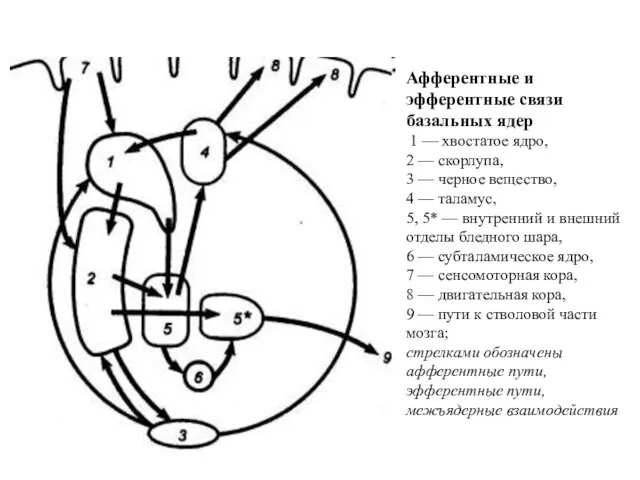
Афферентные и эфферентные связи базальных ядер
1 — хвостатое ядро,
2
Афферентные и эфферентные связи базальных ядер
1 — хвостатое ядро,
2

Главные связи базальных ганглиев.
Возбуждающие влияния показаны красным, тормозные–черным. БШ –
Главные связи базальных ганглиев.
Возбуждающие влияния показаны красным, тормозные–черным. БШ –

Роль БГ в регуляции в ДА
Отношение между полосатым телом и
Роль БГ в регуляции в ДА
Отношение между полосатым телом и

Влияние полосатого тела
В эксперименте показано тормозное влияние (медиатор АЦХ) хвостатого ядра
Влияние полосатого тела
В эксперименте показано тормозное влияние (медиатор АЦХ) хвостатого ядра

Функции бледного шара
Получает из полосатого тела преимущественно тормозное и частично
Функции бледного шара
Получает из полосатого тела преимущественно тормозное и частично
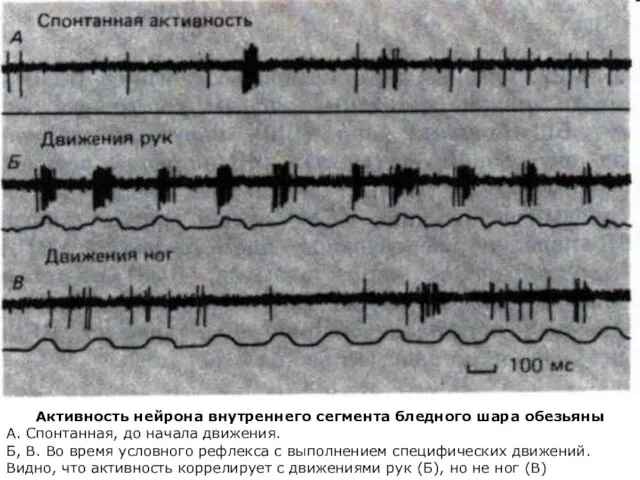
Активность нейрона внутреннего сегмента бледного шара обезьяны
А. Спонтанная, до начала движения.
Активность нейрона внутреннего сегмента бледного шара обезьяны
А. Спонтанная, до начала движения.
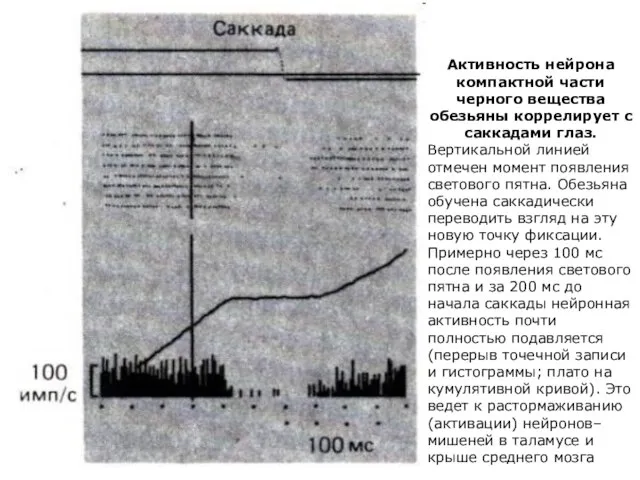
Активность нейрона компактной части черного вещества обезьяны коррелирует с саккадами глаз.
Активность нейрона компактной части черного вещества обезьяны коррелирует с саккадами глаз.

Роль базальных ганглиев в регуляции двигательной активности
При циркуляции возбуждения в стриопаллидарной
Роль базальных ганглиев в регуляции двигательной активности
При циркуляции возбуждения в стриопаллидарной

ФУНКЦИИ БАЗАЛЬНЫХ ГАНГЛИЕВ
1. Центры координации сочетанных двигательных актов
2. Центры сложных безусловных
ФУНКЦИИ БАЗАЛЬНЫХ ГАНГЛИЕВ
1. Центры координации сочетанных двигательных актов
2. Центры сложных безусловных
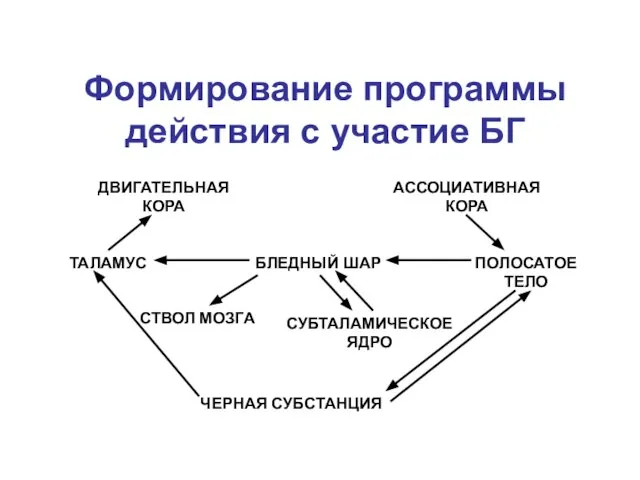
БЛЕДНЫЙ ШАР
ПОЛОСАТОЕ ТЕЛО
СУБТАЛАМИЧЕСКОЕ ЯДРО
СТВОЛ МОЗГА
ТАЛАМУС
ДВИГАТЕЛЬНАЯ КОРА
АССОЦИАТИВНАЯ КОРА
ЧЕРНАЯ СУБСТАНЦИЯ
Формирование программы действия с
БЛЕДНЫЙ ШАР
ПОЛОСАТОЕ ТЕЛО
СУБТАЛАМИЧЕСКОЕ ЯДРО
СТВОЛ МОЗГА
ТАЛАМУС
ДВИГАТЕЛЬНАЯ КОРА
АССОЦИАТИВНАЯ КОРА
ЧЕРНАЯ СУБСТАНЦИЯ
Формирование программы действия с
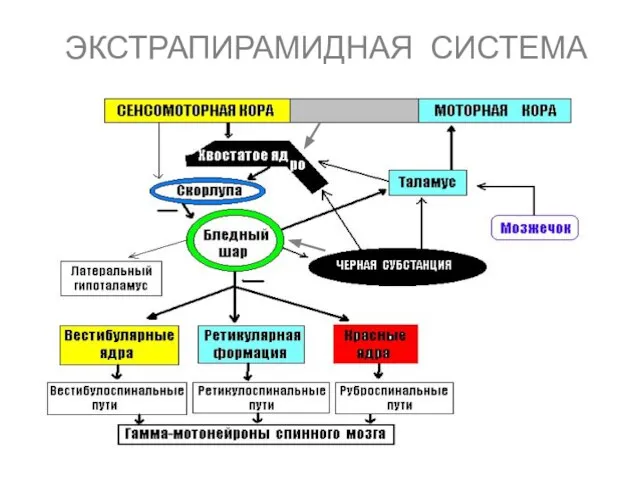
ЭКСТРАПИРАМИДНАЯ СИСТЕМА
ЭКСТРАПИРАМИДНАЯ СИСТЕМА

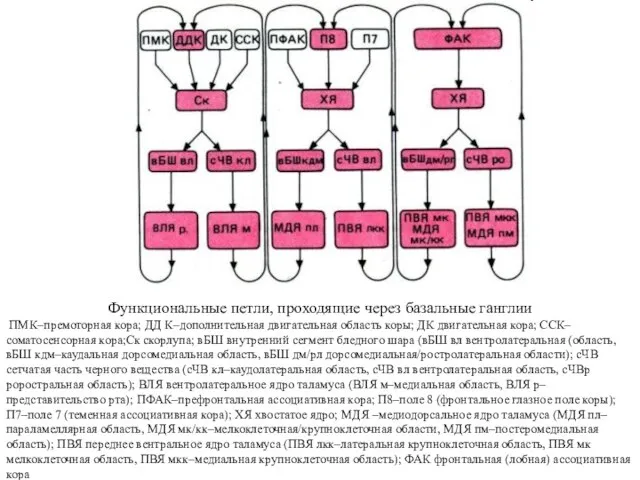
Функциональные петли, проходящие через базальные ганглии
ПМК–премоторная кора; ДД К–дополнительная
Функциональные петли, проходящие через базальные ганглии
ПМК–премоторная кора; ДД К–дополнительная

ЭФФЕКТЫ ПОРАЖЕНИЯ СТРИОПАЛЛИДАРНОЙ СИСТЕМЫ
Поражения хвостатого ядра:
гиперкинезы- атетозы и хорея (пляска
ЭФФЕКТЫ ПОРАЖЕНИЯ СТРИОПАЛЛИДАРНОЙ СИСТЕМЫ
Поражения хвостатого ядра:
гиперкинезы- атетозы и хорея (пляска

ЭФФЕКТЫ ПОРАЖЕНИЯ СТРИОПАЛЛИДАРНОЙ СИСТЕМЫ
Базальные ядра совместно с корой больших полушарий контролируют
ЭФФЕКТЫ ПОРАЖЕНИЯ СТРИОПАЛЛИДАРНОЙ СИСТЕМЫ
Базальные ядра совместно с корой больших полушарий контролируют

Болезнь Хантингтона - при потери ГАМК–ергических и холинергических нейронов полосатого тела
Болезнь Хантингтона - при потери ГАМК–ергических и холинергических нейронов полосатого тела

ЛИМБИЧЕСКАЯ СИСТЕМА
ЛИМБИЧЕСКАЯ СИСТЕМА

Лимбическая система
Функциональное объединение различных структур конечного, промежуточного и среднего мозга, обеспечивающее
Лимбическая система
Функциональное объединение различных структур конечного, промежуточного и среднего мозга, обеспечивающее

Афферентные входы
От различных областей головного мозга
Через гипоталамус от ретикулярной формации
Афферентные входы
От различных областей головного мозга
Через гипоталамус от ретикулярной формации
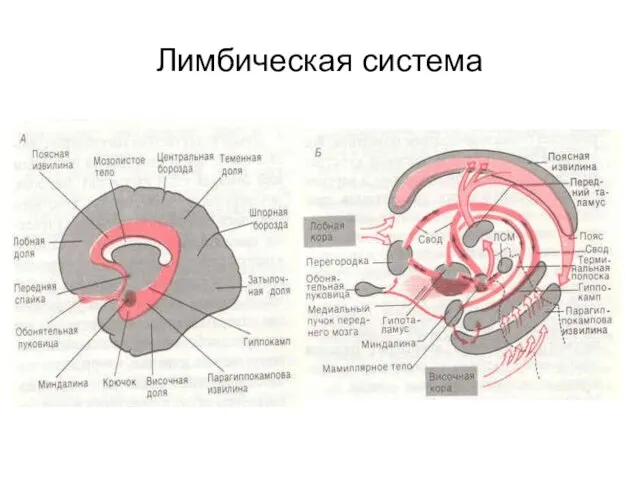
Лимбическая система
Лимбическая система

ОСНОВНЫЕ ФУНКЦИИ ЛИМБИКИ
1. Организация вегетативно-соматических компонентов эмоций
2. Организация кратковременной и долговременной
ОСНОВНЫЕ ФУНКЦИИ ЛИМБИКИ
1. Организация вегетативно-соматических компонентов эмоций
2. Организация кратковременной и долговременной

Кольцевые нейрональные связи
Дают возможность длительной циркуляции (реверберации) возбуждения, что создает условия
Кольцевые нейрональные связи
Дают возможность длительной циркуляции (реверберации) возбуждения, что создает условия
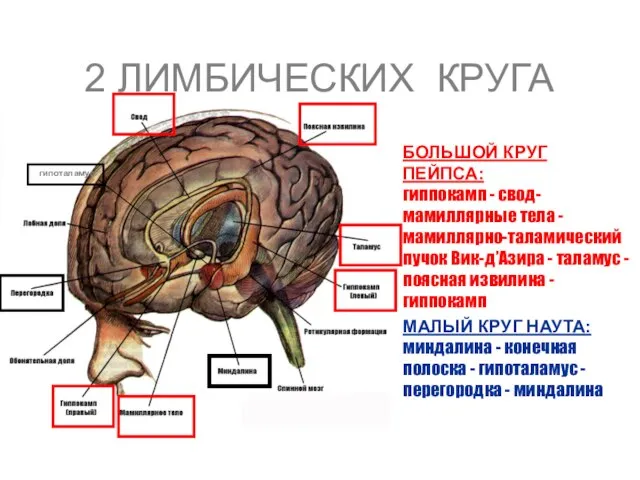
2 ЛИМБИЧЕСКИХ КРУГА
БОЛЬШОЙ КРУГ ПЕЙПСА: гиппокамп - свод- мамиллярные тела -
2 ЛИМБИЧЕСКИХ КРУГА
БОЛЬШОЙ КРУГ ПЕЙПСА: гиппокамп - свод- мамиллярные тела -

Функциональная структура лимбики по МакЛину (1970)
1. Нижний отдел - миндалина и
Функциональная структура лимбики по МакЛину (1970)
1. Нижний отдел - миндалина и

Функции
лимбической системы
После обработки информации от внешней среды и внутренних органах
Функции
лимбической системы
После обработки информации от внешней среды и внутренних органах

Физиология гиппокампа
Физиология гиппокампа


Физиология гиппокампа
Гиппокамп (hippocampus) является основной структурой лимбической системы. Морфологически гиппокамп
Физиология гиппокампа
Гиппокамп (hippocampus) является основной структурой лимбической системы. Морфологически гиппокамп

Физиология гиппокампа
Значение тета-ритма заключается в том, что он отражает реакцию гиппокампа,
Физиология гиппокампа
Значение тета-ритма заключается в том, что он отражает реакцию гиппокампа,

Физиология миндалевидного тела
Физиология миндалевидного тела


Физиология миндалевидного тела
Миндалевидное тело, миндалина — подкорковая структура лимбической системы,
Физиология миндалевидного тела
Миндалевидное тело, миндалина — подкорковая структура лимбической системы,

Физиология миндалевидного тела
При искусственной активации миндалины появляются реакции:
принюхивания,
облизывания,
жевания, глотания,
саливации,
Физиология миндалевидного тела
При искусственной активации миндалины появляются реакции:
принюхивания,
облизывания,
жевания, глотания,
саливации,

Влияние миндалин на иерархические отношения в стае
До операции
После удаления миндалин у
Влияние миндалин на иерархические отношения в стае
До операции
После удаления миндалин у
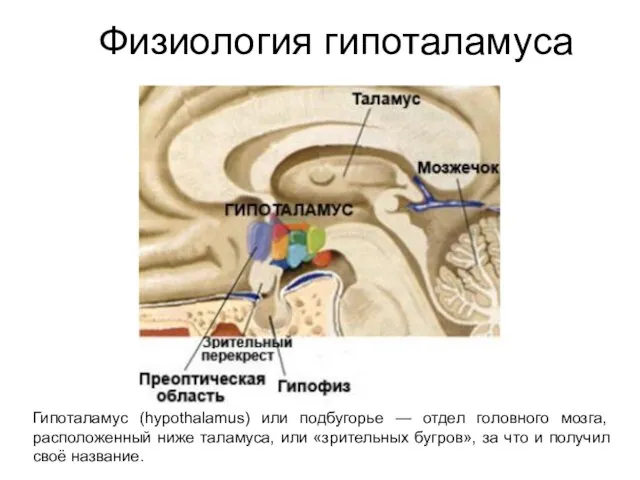
Физиология гипоталамуса
Гипоталамус (hypothalamus) или подбугорье — отдел головного мозга, расположенный
Физиология гипоталамуса
Гипоталамус (hypothalamus) или подбугорье — отдел головного мозга, расположенный

Основные структуры гипоталамуса
Основные структуры гипоталамуса

ФУНКЦИИ ГИПОТАЛАМУСА
Высший центр регуляции вегетативной нервной системы
Высший центр регуляции эндокринных функций
Регуляция
ФУНКЦИИ ГИПОТАЛАМУСА
Высший центр регуляции вегетативной нервной системы
Высший центр регуляции эндокринных функций
Регуляция

Поведенческие функции гипоталамуса
Эффекты стимуляции гипоталамуса
Латеральный гипоталамус: жажда, аппетит, увеличение активности
Поведенческие функции гипоталамуса
Эффекты стимуляции гипоталамуса
Латеральный гипоталамус: жажда, аппетит, увеличение активности

Центры поощрения
В различные области мозга обезьяны вводили электроды, и обезьяна,
Центры поощрения
В различные области мозга обезьяны вводили электроды, и обезьяна,

Центры поощрения и наказания
Личные оценки могут характеризовать ощущения как приятные
Центры поощрения и наказания
Личные оценки могут характеризовать ощущения как приятные

Главные (первичные) центры поощрения располагаются по ходу медиального пучка переднего мозга
Главные (первичные) центры поощрения располагаются по ходу медиального пучка переднего мозга

Центры наказания
и реакции избегания расположены в центральном сером веществе, окружающем
Центры наказания
и реакции избегания расположены в центральном сером веществе, окружающем

Ярость возникает при активации центров наказания. В этом состоянии даже легкая
Ярость возникает при активации центров наказания. В этом состоянии даже легкая

Роль поощрения и наказания в поведении, обучении и памяти
Всё или
Роль поощрения и наказания в поведении, обучении и памяти
Всё или

Привыкание. Новые сенсорные стимулы почти всегда возбуждают значительные области коры больших
Привыкание. Новые сенсорные стимулы почти всегда возбуждают значительные области коры больших

Регуляция вегетативных (висцеральных) функций
Осуществляется преимущественно через гипоталамус, куда информация поступает
Регуляция вегетативных (висцеральных) функций
Осуществляется преимущественно через гипоталамус, куда информация поступает

Роль лимбической системы в обучении
Связана с кругом Пейпеца, где главную роль
Роль лимбической системы в обучении
Связана с кругом Пейпеца, где главную роль

ЭМОЦИОНАЛЬНЫЙ МОЗГ
ЭМОЦИОНАЛЬНЫЙ МОЗГ

ЦИТАТА
«Является ли эмоция продуктом волшебства или физиологическим процессом, который зависит от
ЦИТАТА
«Является ли эмоция продуктом волшебства или физиологическим процессом, который зависит от

Формирование эмоций (пережеваний)
Наиболее важная функция лимбической системы. В свою очередь, эмоции,
Формирование эмоций (пережеваний)
Наиболее важная функция лимбической системы. В свою очередь, эмоции,

Эмоция — специфическое состояние психической сферы, обусловленная рефлекторной реакцией организма на
Эмоция — специфическое состояние психической сферы, обусловленная рефлекторной реакцией организма на

Эмоциональное возбуждение как результат определенной мотивационной деятельности теснейшим образом связано с
Эмоциональное возбуждение как результат определенной мотивационной деятельности теснейшим образом связано с

Удовлетворенная потребность вызывает эмоциональное переживание положительного характера и определяет направление поведенческой
Удовлетворенная потребность вызывает эмоциональное переживание положительного характера и определяет направление поведенческой

Эмоции, проявляются при недостатке точных сведений и путей достижения жизненных потребностей.
Эмоции, проявляются при недостатке точных сведений и путей достижения жизненных потребностей.

По Г. И. Косицкому, оценка величины эмоционального напряжения определяется по формуле:
СН
По Г. И. Косицкому, оценка величины эмоционального напряжения определяется по формуле:
СН

Первая стадия напряжения (CHI) — состояние внимания, мобилизация активности, повышение работоспособности.
Первая стадия напряжения (CHI) — состояние внимания, мобилизация активности, повышение работоспособности.

Первая стадия напряжения (CHI) — состояние внимания, мобилизация активности, повышение работоспособности.
Первая стадия напряжения (CHI) — состояние внимания, мобилизация активности, повышение работоспособности.

Третья стадия (СНШ) — астеническая отрицательная реакция, характеризующаяся истощением ресурсов организма
Третья стадия (СНШ) — астеническая отрицательная реакция, характеризующаяся истощением ресурсов организма

В теории функциональной системы по П. К. Анохину нейрофизиологическая природа эмоций
В теории функциональной системы по П. К. Анохину нейрофизиологическая природа эмоций

Регуляция вегетативных (висцеральных) функций
Осуществляется преимущественно через гипоталамус, куда информация поступает
Регуляция вегетативных (висцеральных) функций
Осуществляется преимущественно через гипоталамус, куда информация поступает

Формирование эмоций (пережеваний)
Наиболее важная функция лимбической системы. В свою очередь, эмоции,
Формирование эмоций (пережеваний)
Наиболее важная функция лимбической системы. В свою очередь, эмоции,

Роль лимбической системы в обучении
Связана с кругом Пейпеца, где главную роль
Роль лимбической системы в обучении
Связана с кругом Пейпеца, где главную роль

ФИЗИОЛОГИЯ
КОРЫ БОЛЬШИХ
ПОЛУШАРИЙ ГОЛОВНОГО МОЗГА
КОРЫ БОЛЬШИХ
ПОЛУШАРИЙ ГОЛОВНОГО МОЗГА


Вызванные потенциалы в коре
Первичный ответ
Вторичный ответ
Вызванные потенциалы в коре
Первичный ответ
Вторичный ответ


Расположение нейронов и нервных волокон в коре больших полушарий.
Номера слоёв коры
Расположение нейронов и нервных волокон в коре больших полушарий. Номера слоёв коры

Слои коры больших полушарий
1 слой - верхний молекулярный - ветвления дендритов
Слои коры больших полушарий
1 слой - верхний молекулярный - ветвления дендритов

КБП
В I – IV слоях коры происходит восприятие и обработка поступающей
КБП
В I – IV слоях коры происходит восприятие и обработка поступающей

Функциональная организация коры
Функциональная единица коры - вертикальная колонка диаметром около 500
Функциональная организация коры
Функциональная единица коры - вертикальная колонка диаметром около 500

Нейронные колонки
В коре имеются функциональные объединения нейронов, расположенные в цилиндрике диаметром
Нейронные колонки
В коре имеются функциональные объединения нейронов, расположенные в цилиндрике диаметром
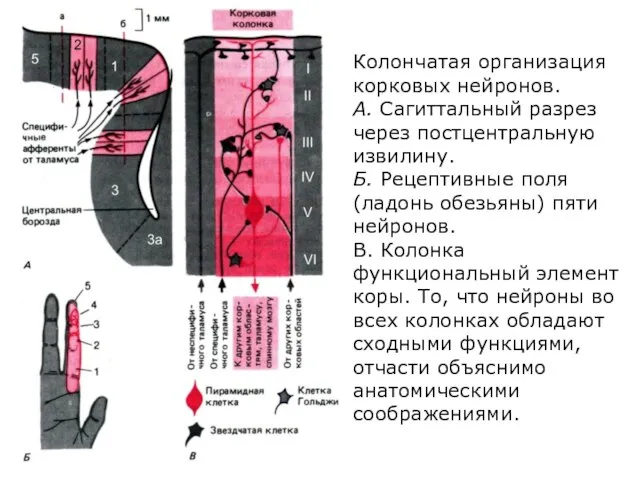
Колончатая организация корковых нейронов.
А. Сагиттальный разрез через постцентральную извилину.
Б.
Колончатая организация корковых нейронов.
А. Сагиттальный разрез через постцентральную извилину.
Б.
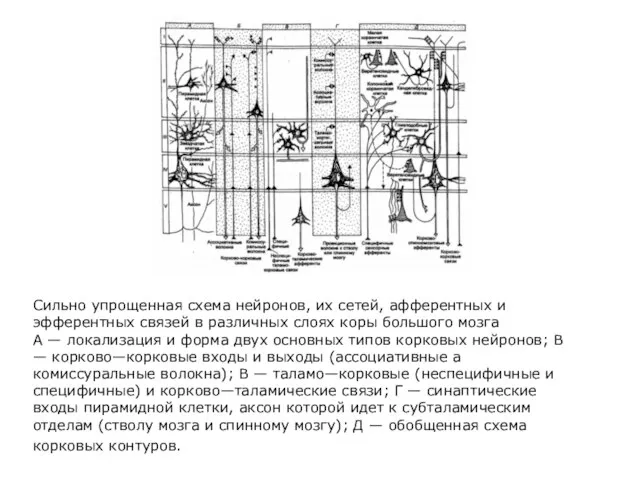
Сильно упрощенная схема нейронов, их сетей, афферентных и эфферентных связей в
Сильно упрощенная схема нейронов, их сетей, афферентных и эфферентных связей в

ФУНКЦИОНАЛЬНЫЕ ЗОНЫ КОРЫ
СЕНСОРНЫЕ ( зрительные, слуховые, кожные и др.)
МОТОРНЫЕ ( первичные,
ФУНКЦИОНАЛЬНЫЕ ЗОНЫ КОРЫ
СЕНСОРНЫЕ ( зрительные, слуховые, кожные и др.)
МОТОРНЫЕ ( первичные,
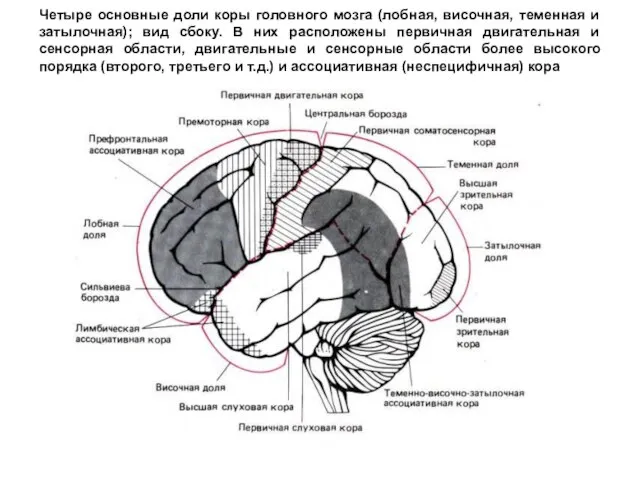
Четыре основные доли коры головного мозга (лобная, височная, теменная и затылочная);
Четыре основные доли коры головного мозга (лобная, височная, теменная и затылочная);

Локализация функций в КБП
Как современная нейробиологическая концепция локализации функций базируется на
Локализация функций в КБП
Как современная нейробиологическая концепция локализации функций базируется на
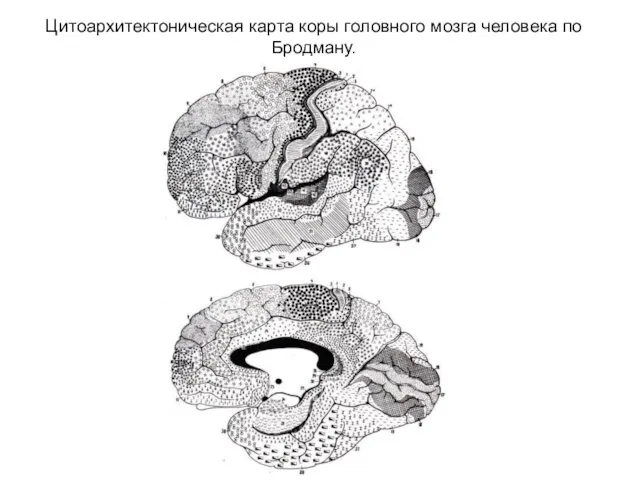
Цитоархитектоническая карта коры головного мозга человека по Бродману.
Цитоархитектоническая карта коры головного мозга человека по Бродману.
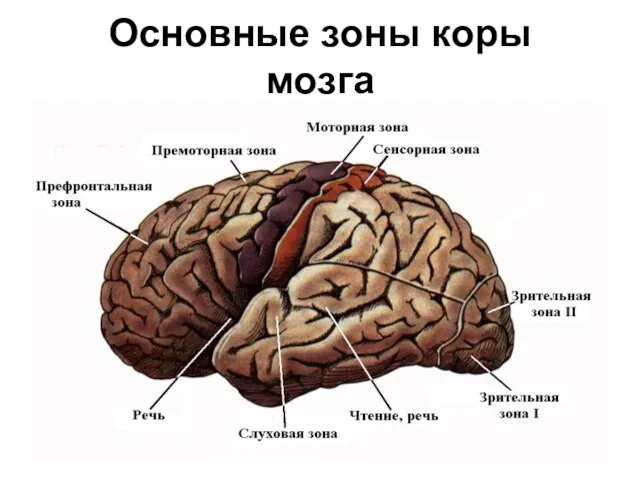
Основные зоны коры мозга
Основные зоны коры мозга
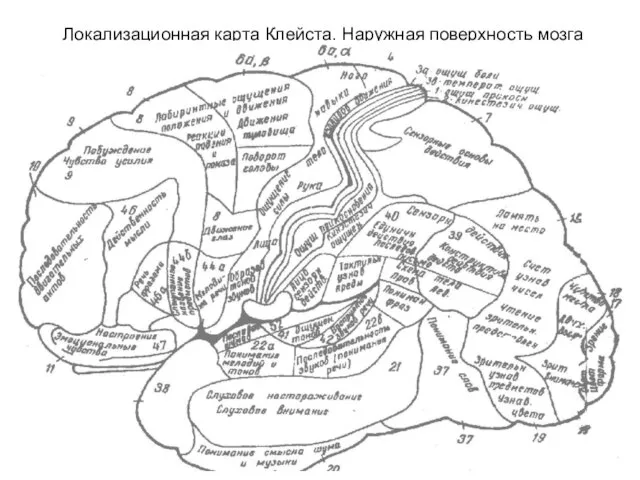
Локализационная карта Клейста. Наружная поверхность мозга
Локализационная карта Клейста. Наружная поверхность мозга
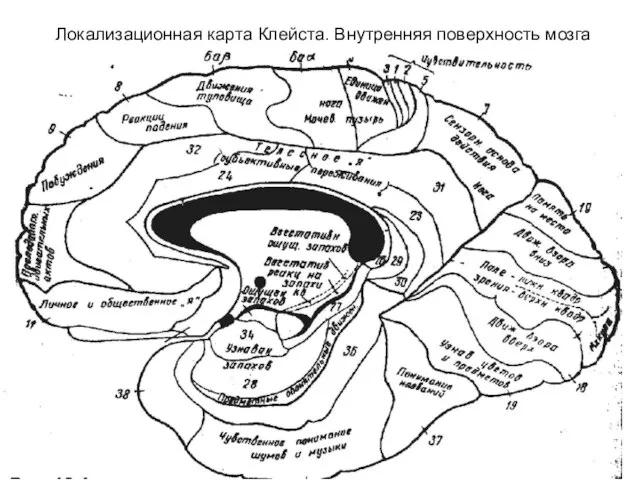
Локализационная карта Клейста. Внутренняя поверхность мозга
Локализационная карта Клейста. Внутренняя поверхность мозга

Сенсорные зоны КБП
В этих зонах проецируется информация от рецепторов всех рецептивных
Сенсорные зоны КБП
В этих зонах проецируется информация от рецепторов всех рецептивных

В сенсорных зонах коры различают
Первичные сенсорные поля (проекционные или центральные поля)
В сенсорных зонах коры различают
Первичные сенсорные поля (проекционные или центральные поля)

Представительство вторичных сенсорных зон
Их развитие происходит в первые месяцы после рождения,
Представительство вторичных сенсорных зон
Их развитие происходит в первые месяцы после рождения,

Проекции чувствительности в коре постцентральной извилины
Представлены:
Головы и верхних отделов
Проекции чувствительности в коре постцентральной извилины
Представлены:
Головы и верхних отделов
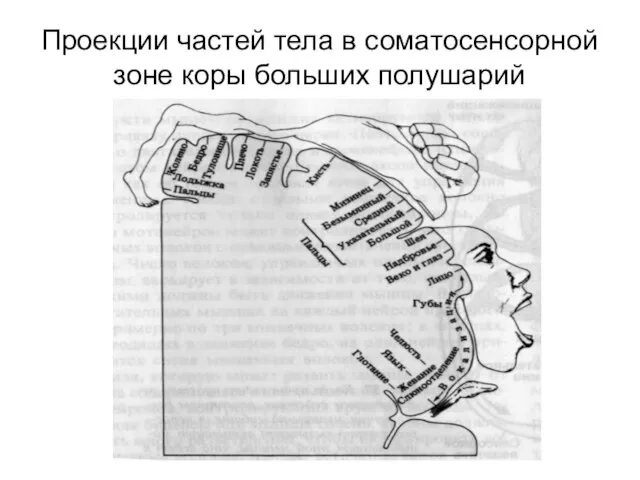
Проекции частей тела в соматосенсорной зоне коры больших полушарий
Проекции частей тела в соматосенсорной зоне коры больших полушарий

Слуховая сенсорная кора
Расположена в верхней височной извилине и представлена:
Первичным
Слуховая сенсорная кора
Расположена в верхней височной извилине и представлена:
Первичным

Зрительная сенсорная кора
представлена в затылочной доли (поля 17 и 18).
Зрительная сенсорная кора
представлена в затылочной доли (поля 17 и 18).

Вторичная зрительная область (поля 18 и 19)
в ней осуществляется тонкий
Вторичная зрительная область (поля 18 и 19)
в ней осуществляется тонкий

Ассоциативные области коры
- включают участки новой КБП, которые расположены рядом
Ассоциативные области коры
- включают участки новой КБП, которые расположены рядом

Лобная ассоциативная кора (поля 9 – 14)
имеет основной источник информации
Лобная ассоциативная кора (поля 9 – 14)
имеет основной источник информации

Теменная ассоциативная кора (5, 7, 39, 40)
- получает афферентные вещи от
Теменная ассоциативная кора (5, 7, 39, 40)
- получает афферентные вещи от

ФУНКЦИИ ЛОБНЫХ ДОЛЕЙ
1. Управление врожденными поведенческими реакциями при помощи накопленного опыта
2.
ФУНКЦИИ ЛОБНЫХ ДОЛЕЙ
1. Управление врожденными поведенческими реакциями при помощи накопленного опыта
2.

Двигательные области коры
Выделяют первичную , вторичную и дополнительную области
Пирамидные нейроны V
Двигательные области коры
Выделяют первичную , вторичную и дополнительную области
Пирамидные нейроны V

Первичная моторная зона коры
Прецентральная извилина (поле 4)
Нейроны управляют функцией мотонейронов мышц
Первичная моторная зона коры
Прецентральная извилина (поле 4)
Нейроны управляют функцией мотонейронов мышц

Вторичная и дополнительная двигательная кора
расположена на латеральной поверхности полушарий, впереди
Вторичная и дополнительная двигательная кора
расположена на латеральной поверхности полушарий, впереди
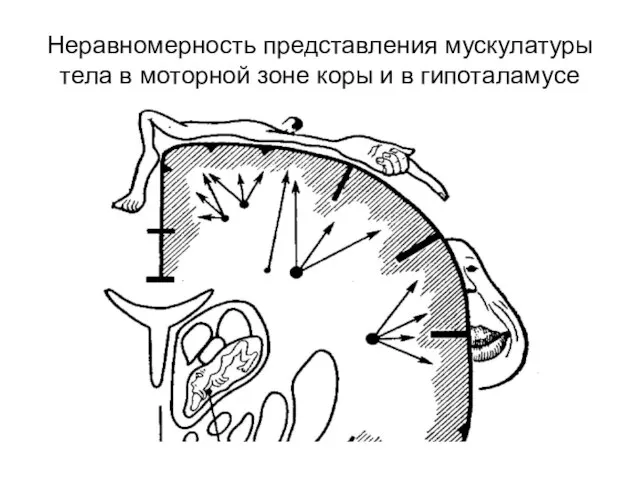
Неравномерность представления мускулатуры тела в моторной зоне коры и в гипоталамусе
Неравномерность представления мускулатуры тела в моторной зоне коры и в гипоталамусе
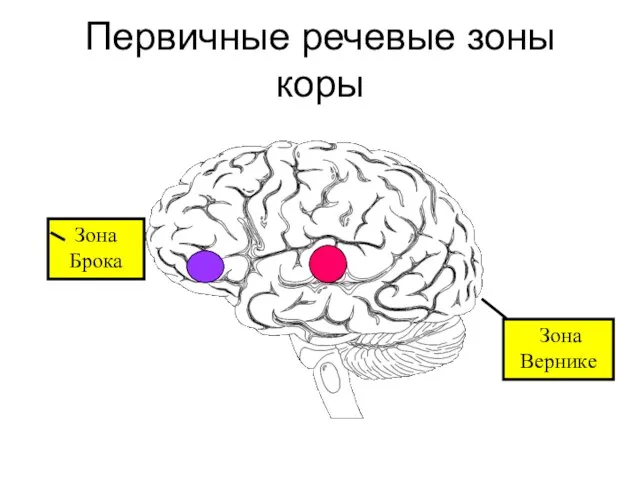
Первичные речевые зоны коры
Зона
Брока
Зона
Вернике
Первичные речевые зоны коры
Зона
Брока
Зона
Вернике
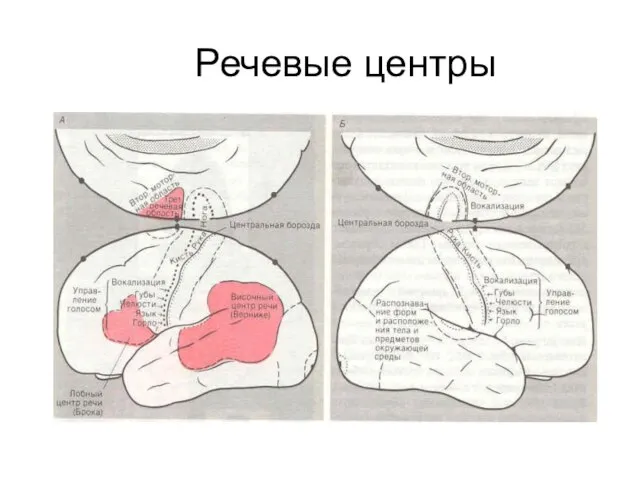
Речевые центры
Речевые центры
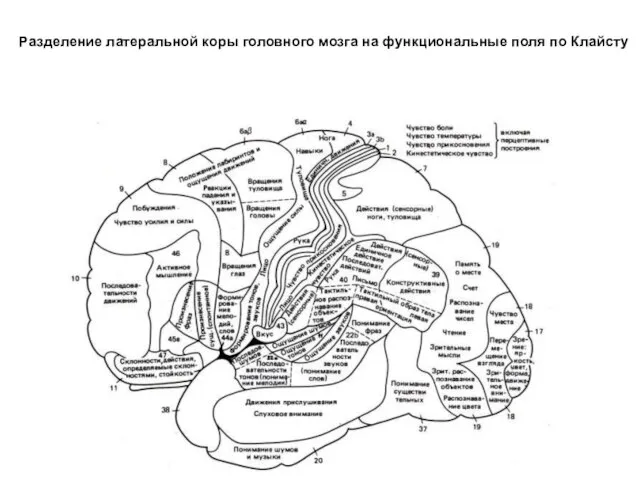
Разделение латеральной коры головного мозга на функциональные поля по Клайсту
Разделение латеральной коры головного мозга на функциональные поля по Клайсту
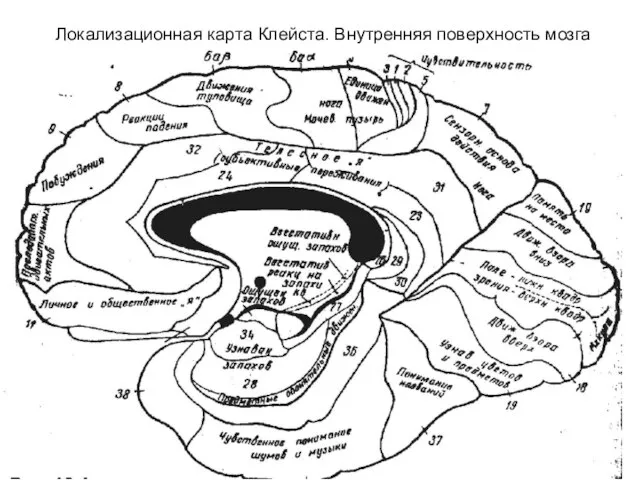
Локализационная карта Клейста. Внутренняя поверхность мозга
Локализационная карта Клейста. Внутренняя поверхность мозга
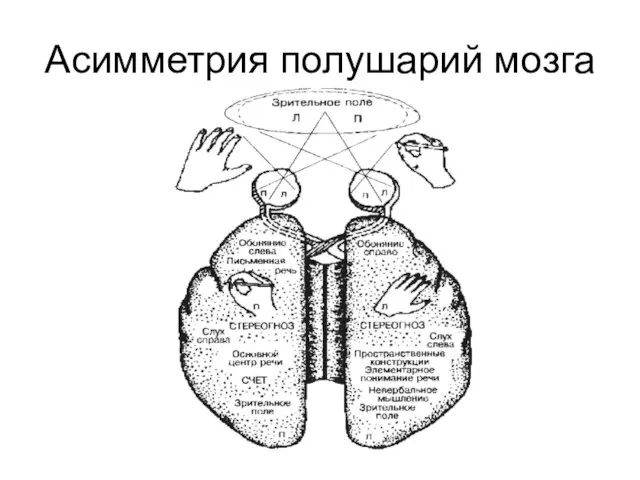
Асимметрия полушарий мозга
Асимметрия полушарий мозга

Цитаты
«Мы говорим левым полушарием»
Поль Брока, 1865
« Сознание локализуется в
Цитаты
«Мы говорим левым полушарием»
Поль Брока, 1865
« Сознание локализуется в

Межполушарные различия
ЛЕВОЕ ПОЛУШАРИЕ ПРАВОЕ ПОЛУШАРИЕ
Лучше узнаются стимулы
Словесные Несловесные
Легко
Межполушарные различия
ЛЕВОЕ ПОЛУШАРИЕ ПРАВОЕ ПОЛУШАРИЕ
Лучше узнаются стимулы
Словесные Несловесные
Легко

Распознавание лиц левым и правым полушариями больного с расщепленным мозгом
Распознавание лиц левым и правым полушариями больного с расщепленным мозгом

Общая схема организации двигательной активности
Общая схема организации двигательной активности
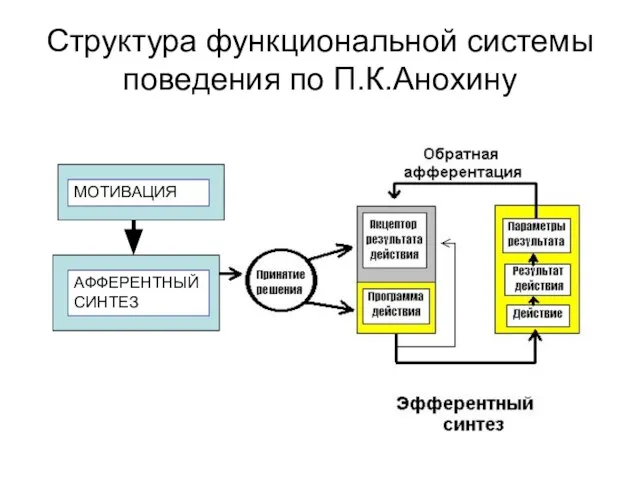
Структура функциональной системы поведения по П.К.Анохину
МОТИ ВАЦИЯ
МОТИВАЦИЯ
АФФЕРЕНТНЫЙ СИНТЕЗ
Структура функциональной системы поведения по П.К.Анохину
МОТИ ВАЦИЯ
МОТИВАЦИЯ
АФФЕРЕНТНЫЙ СИНТЕЗ
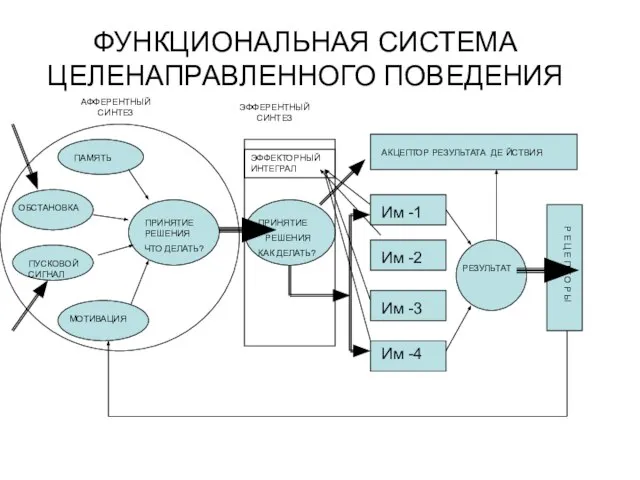
ФУНКЦИОНАЛЬНАЯ СИСТЕМА ЦЕЛЕНАПРАВЛЕННОГО ПОВЕДЕНИЯ
АФФЕРЕНТНЫЙ СИНТЕЗ
ПРИНЯТИЕ РЕШЕНИЯ
ЧТО ДЕЛАТЬ?
ЭФФЕРЕНТНЫЙ СИНТЕЗ
ПРИНЯТИЕ
ФУНКЦИОНАЛЬНАЯ СИСТЕМА ЦЕЛЕНАПРАВЛЕННОГО ПОВЕДЕНИЯ
АФФЕРЕНТНЫЙ СИНТЕЗ
ПРИНЯТИЕ РЕШЕНИЯ
ЧТО ДЕЛАТЬ?
ЭФФЕРЕНТНЫЙ СИНТЕЗ
ПРИНЯТИЕ
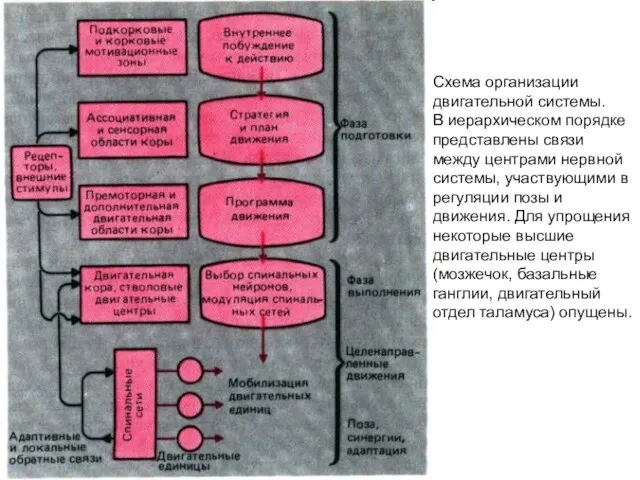
Схема организации двигательной системы.
В иерархическом порядке представлены связи между центрами
Схема организации двигательной системы.
В иерархическом порядке представлены связи между центрами
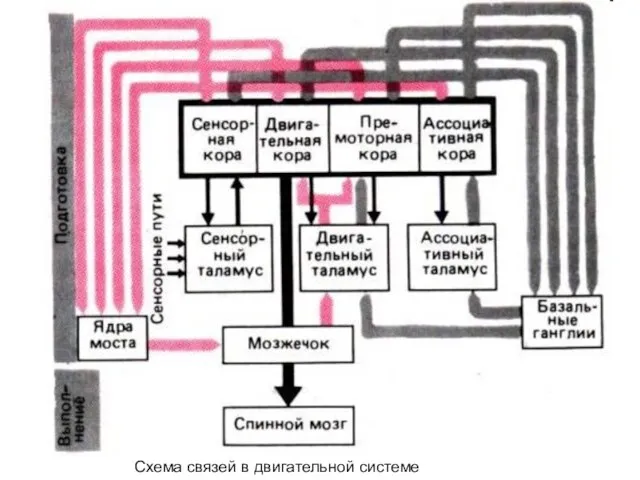
Схема связей в двигательной системе
Схема связей в двигательной системе
 Менингококковая инфекция
Менингококковая инфекция Нервная система
Нервная система Ожирение среди подростков. Ожирение - хроническое заболевание
Ожирение среди подростков. Ожирение - хроническое заболевание Танымдық психикалық процестер: түйсік, қабылдау
Танымдық психикалық процестер: түйсік, қабылдау Лфк при заболеваниях центральной нервной системы
Лфк при заболеваниях центральной нервной системы Органы кроветворения
Органы кроветворения Спирометрия. Внешнее дыхание
Спирометрия. Внешнее дыхание Буллезные дерматозы
Буллезные дерматозы Права пациента в аптечных организациях
Права пациента в аптечных организациях Вакцинопрофилактика менингококковой инфекции у детей и перспективы ее включения
Вакцинопрофилактика менингококковой инфекции у детей и перспективы ее включения Описторхоз у детей
Описторхоз у детей Создание благоприятного социально-психологического климата в классном коллективе учащихся и родителей
Создание благоприятного социально-психологического климата в классном коллективе учащихся и родителей Информация по компании Интертич для ТОО Болашак Атырау
Информация по компании Интертич для ТОО Болашак Атырау Туберкулездің емі
Туберкулездің емі Цитологическая диагностика рака молочной железы
Цитологическая диагностика рака молочной железы Флоатинг. Что такое флоатинг?
Флоатинг. Что такое флоатинг? Опиоиды как адъюванты для регионарной анестезии
Опиоиды как адъюванты для регионарной анестезии Миодистрофия Дюшенна. Обзор лечения. Стероиды
Миодистрофия Дюшенна. Обзор лечения. Стероиды Анализ заболеваемости, диагностики и профилактики туберкулёза в республике Крым
Анализ заболеваемости, диагностики и профилактики туберкулёза в республике Крым Леонардо да Винчи – анатом
Леонардо да Винчи – анатом Бүйрекке тас зілуінің рентгенологиялық зерттеу әдістері
Бүйрекке тас зілуінің рентгенологиялық зерттеу әдістері Заболевания женской и мужской половой системы. Патология беременности и последа
Заболевания женской и мужской половой системы. Патология беременности и последа Сенсорные процессы у человека. Психологическая сущность ощущений. Тема 2
Сенсорные процессы у человека. Психологическая сущность ощущений. Тема 2 Особенности организма детей среднего и старшего возраста
Особенности организма детей среднего и старшего возраста Дерматовенерология. Анатомия, гистология и функции кожи
Дерматовенерология. Анатомия, гистология и функции кожи Швы хирургические
Швы хирургические Дисциркуляторная энцефалопатия
Дисциркуляторная энцефалопатия Реабилитационный процесс восстановления речи при афазии
Реабилитационный процесс восстановления речи при афазии