- Барометрическая гипоксия Нырятельная гипоксия

Содержание
- 2. Внешняя среда ↓ Органы дыхания (легкие, жабры) ↓ Кровеносная система + Кровь (О2–переносящий белок) ↓ Внутриклеточные
- 3. Таблица 18 Состав сухого атмосферного воздуха, %, (Otis, 1964), по: [71] Азот 78.09 Кислород 20.95 Аргон
- 4. Таблица 19 Парциальное давление (р) и концентрация (с) кислорода и двуокиси углерода в крови и альвеолярном
- 5. Гипоксия возникает, когда давление кислорода (рО2) во внешней среде снижается: при падении барометрического давления, 2. при
- 6. 1-высота, на которой большинство неакклиматизированных людей теряет сознание от недостатка кислорода, 2-наибольшая высота постоянных поселений человека,
- 7. У спортсменов максимальное время бега со скоростью 12 км/ч: Уровень моря – 30 мин Высота 2270
- 8. Рис. 46. Концентрация молочной кислоты в крови крыс, находящихся 1 ч в воздушной среде с разным
- 9. Рис. 47. Динамика концентрации молочной кислоты в тканях белых крыс во время пребывания в горах Тянь-Шаня
- 10. Рис. 48`. Физиологические изменения, протекающие в организме в процессе адаптации к гипоксии (Барбашова, 1964), по: [54].
- 12. У человека МОК, л/мин, при дыхании воздушной смесью с концентрацией кислорода 8.5% до 5 мин 10
- 14. Рис. 49. Минутный объем кровообращения у спокойных собак во время месячного пребывания на высоте 3200 м
- 15. Таблица 22. Органная скорость кровотока (мл/мин-1 • кг-1) у спокойных собак во время пребывания на высоте
- 17. Рис. 50. Содержание оксигемоглобина в артериальной крови лам (1), грызунов - вискунов (2) и овец (3),
- 18. Таблица 24 Гематологические показатели у жителей разных высот (Hurtado, 1964), по: [32]
- 19. Таблица 25. Эритроциты, гемоглобин крови и миоглобин в мышцах у собак, выросших на разных высотах (Hurtado
- 20. Рис. 51. Кривые диссоциации оксигемоглобина крови ламы и викуньи (слева) по сравнению с теми же кривыми
- 21. Рис. 52. Кривые диссоциации оксигемоглобина в крови беременной козы и ее плода (Barcroft, 1935); пунктиром показаны
- 22. При адаптации к гипоксии не всегда находят сдвиг кривой диссоциации НbО2 влево, при котором сродство О2
- 23. Таблица 27 Параметры легких у 24 мужчин – аборигенов различных высот (Gareila-Oyola, Saldana, 1974), по: [32]
- 26. Основное различие этих двух групп рабочих касается крови: при длительном пребывании на большой высоте концентрация в
- 27. Почему снижается работоспособность неадаптированного человека на высоте? На высоте для обеспечения кислородом работы скелетных мышц учащается
- 28. У аборигенов исходные показатели работы легких и сердца ниже, а поверхность легочных альвеол выше. Клеточные системы
- 29. Рис. 53. Нарушение очерка и появление грубых ошибок после 3 мин в барокамере на высоте 7500
- 30. При острой дыхательной гипоксии увеличиваются минутные объемы дыхания и кровообращения, доля анаэробной энергетики в клетках растет
- 31. При длительной дыхательной гипоксии МОК снижается даже ниже исходного уровня, МОД остается повышенным, альвеолярная поверхность легких
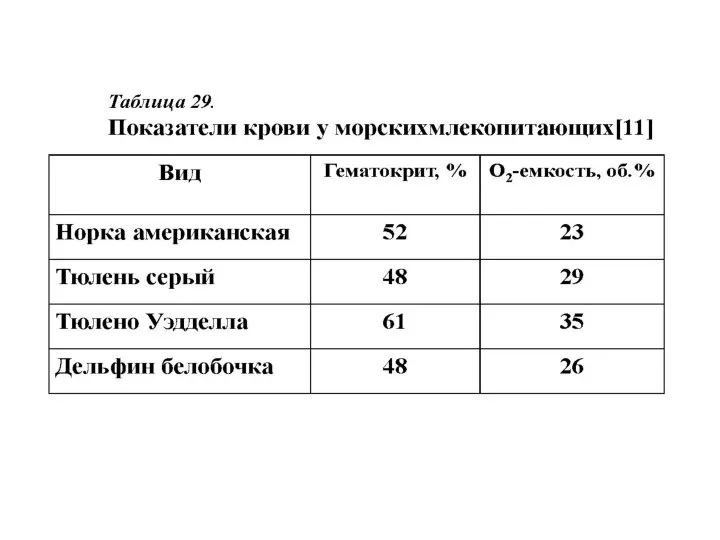
- 33. Таблица 28 Содержание кислорода в организме водных млекопитающих, мл О2/кг Мт [41, 95]
- 35. Таблица 30. Запас кислорода в организме байкальского тюленя [37, 38] При подсчетах было принято: 1) для
- 36. Рис. 55. Рентгенограмма печеночного венозного синуса и печеночных вен у морского котика [11]
- 37. Рис. 56. Кривые диссоциации НbО2 обыкновенного тюленя (1), хохлоча (2), серого тюленя (3) (Scholander, 1940; Lenfant
- 38. У морских млекопитающих энергетика коротких пищевых ныряний аэробна и обеспечена депонированным кислородом.
- 39. Рис. 57. Органные доли МОК у щенков байкальской нерпы при нырянии [5, 36, 76]. Внизу -
- 40. Морские млекопитающие после исчерпания своих кислородных депо используют энергию анаэробного гликолиза. Поэтому органы этих животных, даже
- 41. Таблица 31. Количество углеводов в органах байкальской нерпы с Мт 20 кг до, в конце 19-минутного
- 42. У водных млекопитающих (по большей части это морские животные) Энергетика коротких пищевых ныряний аэробна и обеспечена
- 44. Скачать презентацию

Внешняя среда
↓
Органы дыхания (легкие, жабры)
↓
Кровеносная система + Кровь (О2–переносящий белок)
Внешняя среда
↓
Органы дыхания (легкие, жабры)
↓
Кровеносная система + Кровь (О2–переносящий белок)
![Таблица 18 Состав сухого атмосферного воздуха, %, (Otis, 1964), по: [71]](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1298731/slide-2.jpg)
Таблица 18
Состав сухого атмосферного воздуха,
%, (Otis, 1964), по: [71]
Азот 78.09
Кислород 20.95
Аргон
Таблица 18
Состав сухого атмосферного воздуха,
%, (Otis, 1964), по: [71]
Азот 78.09
Кислород 20.95
Аргон

Таблица 19
Парциальное давление (р) и концентрация (с)
кислорода и двуокиси углерода
Таблица 19
Парциальное давление (р) и концентрация (с)
кислорода и двуокиси углерода

Гипоксия возникает,
когда давление кислорода (рО2)
во внешней среде снижается:
при
Гипоксия возникает,
когда давление кислорода (рО2)
во внешней среде снижается:
при
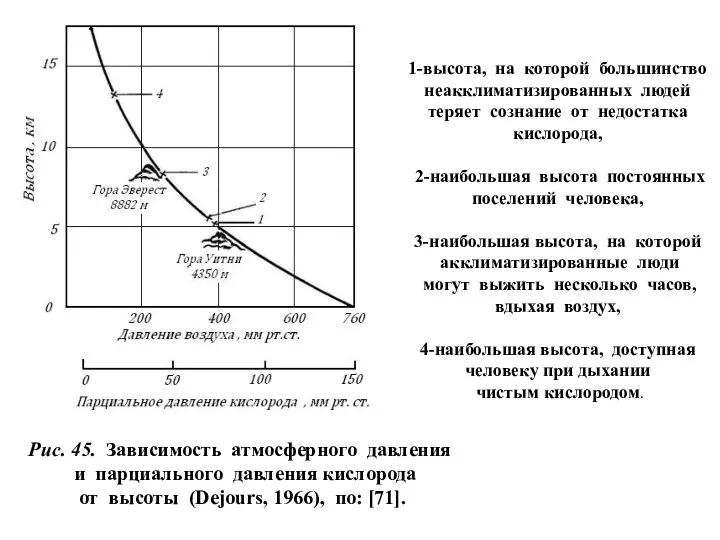
1-высота, на которой большинство
неакклиматизированных людей
теряет сознание от недостатка
1-высота, на которой большинство
неакклиматизированных людей
теряет сознание от недостатка

У спортсменов максимальное время
бега со скоростью 12 км/ч:
Уровень моря –
У спортсменов максимальное время
бега со скоростью 12 км/ч:
Уровень моря –
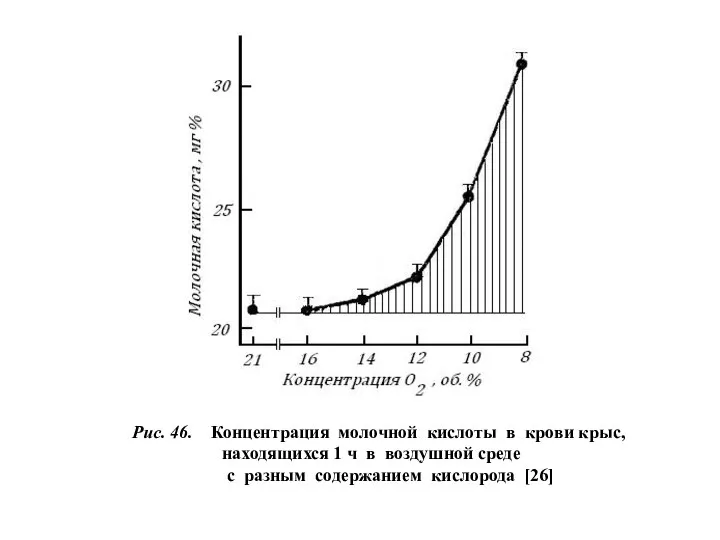
Рис. 46. Концентрация молочной кислоты в крови крыс,
находящихся 1
Рис. 46. Концентрация молочной кислоты в крови крыс,
находящихся 1
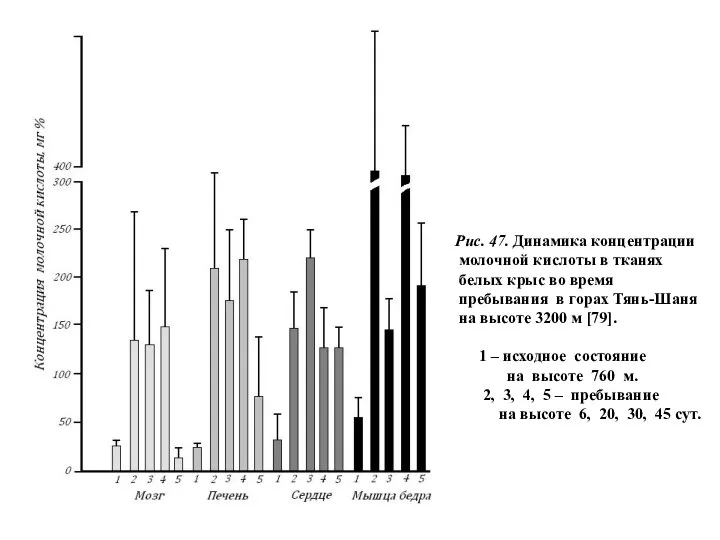
Рис. 47. Динамика концентрации
молочной кислоты в тканях
белых крыс во
Рис. 47. Динамика концентрации
молочной кислоты в тканях
белых крыс во
![Рис. 48`. Физиологические изменения, протекающие в организме в процессе адаптации к гипоксии (Барбашова, 1964), по: [54].](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1298731/slide-9.jpg)
Рис. 48`. Физиологические изменения, протекающие в организме
в процессе адаптации к
Рис. 48`. Физиологические изменения, протекающие в организме
в процессе адаптации к
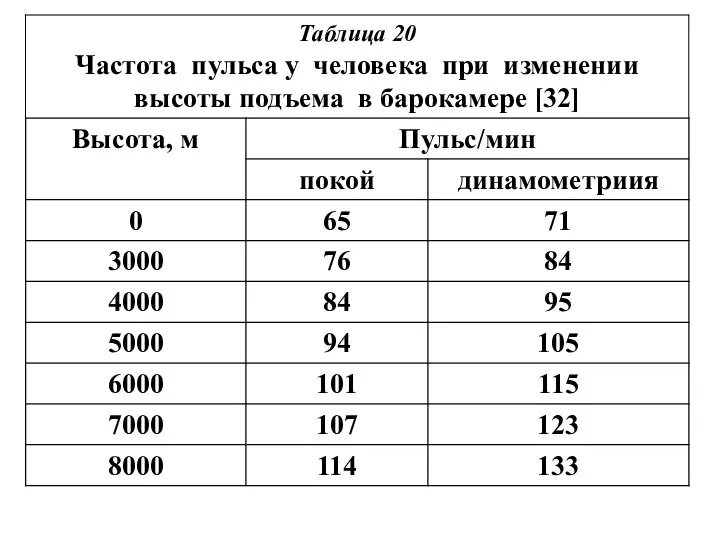

У человека МОК, л/мин,
при дыхании воздушной смесью
с концентрацией кислорода
У человека МОК, л/мин,
при дыхании воздушной смесью
с концентрацией кислорода
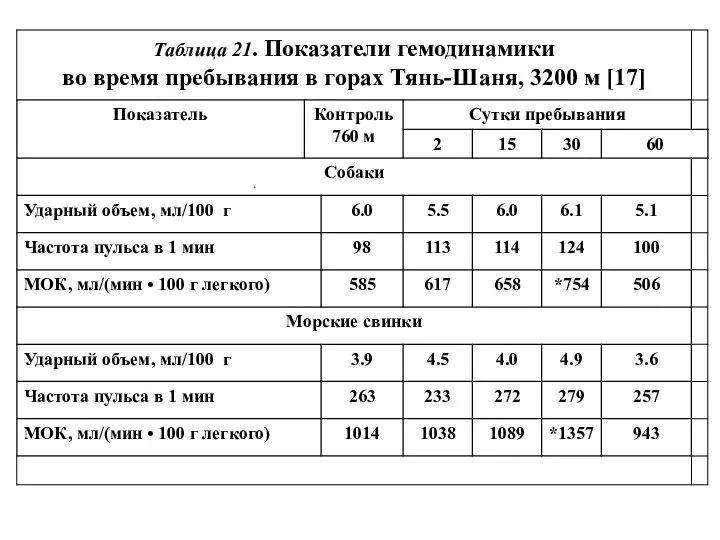
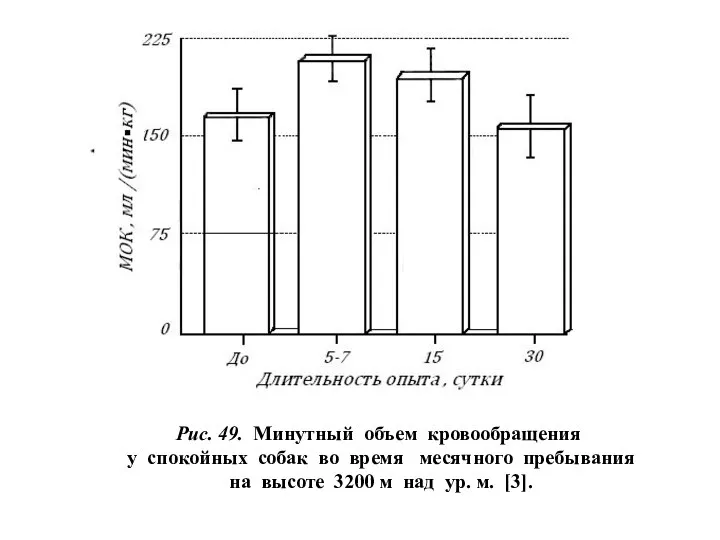
Рис. 49. Минутный объем кровообращения
у спокойных собак во время
Рис. 49. Минутный объем кровообращения
у спокойных собак во время
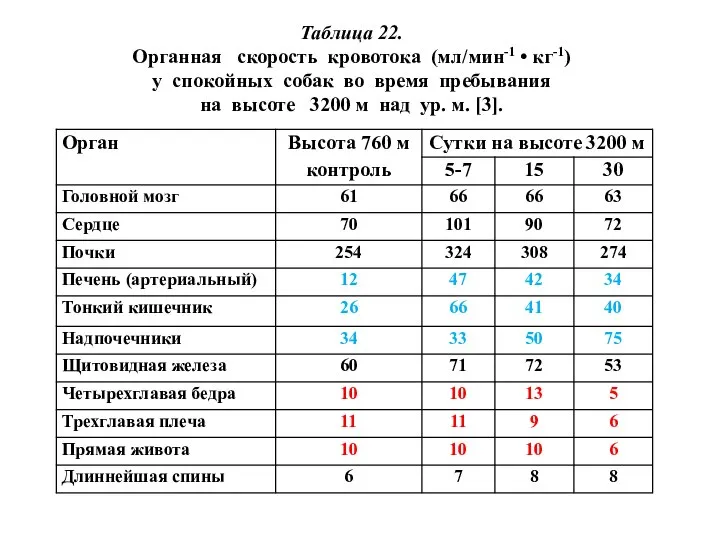
Таблица 22.
Органная скорость кровотока (мл/мин-1 • кг-1)
у спокойных собак
Таблица 22.
Органная скорость кровотока (мл/мин-1 • кг-1)
у спокойных собак
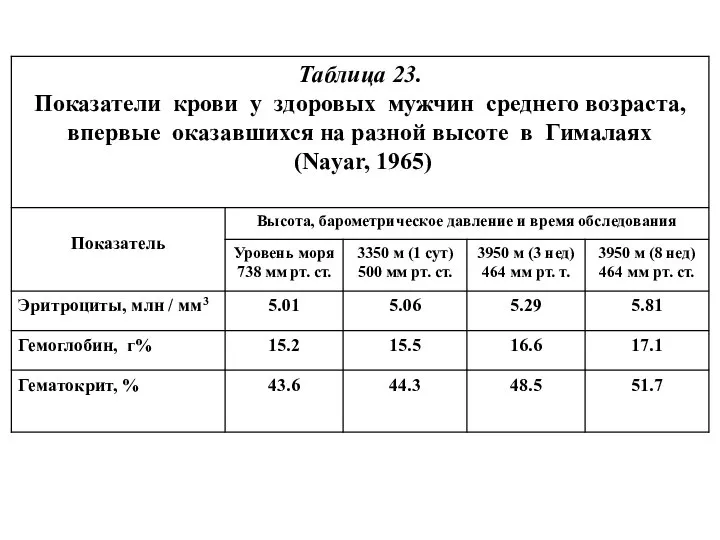
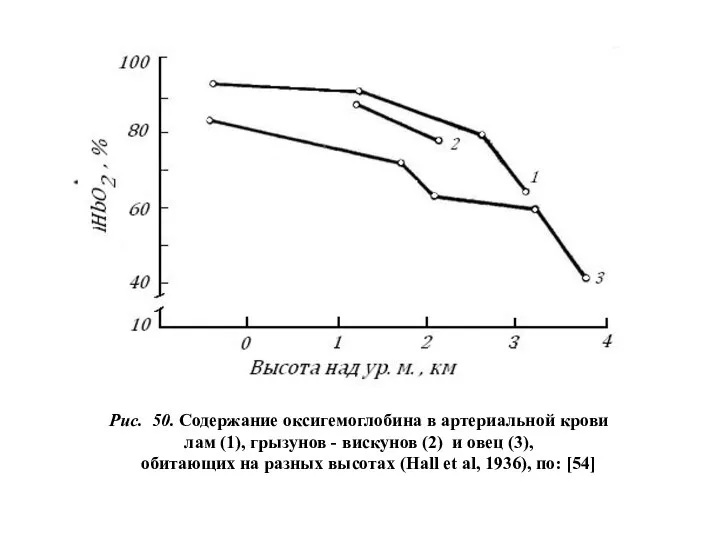
Рис. 50. Содержание оксигемоглобина в артериальной крови
лам (1), грызунов - вискунов
Рис. 50. Содержание оксигемоглобина в артериальной крови
лам (1), грызунов - вискунов
![Таблица 24 Гематологические показатели у жителей разных высот (Hurtado, 1964), по: [32]](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1298731/slide-17.jpg)
Таблица 24
Гематологические показатели у жителей разных высот (Hurtado, 1964), по: [32]
Таблица 24
Гематологические показатели у жителей разных высот (Hurtado, 1964), по: [32]
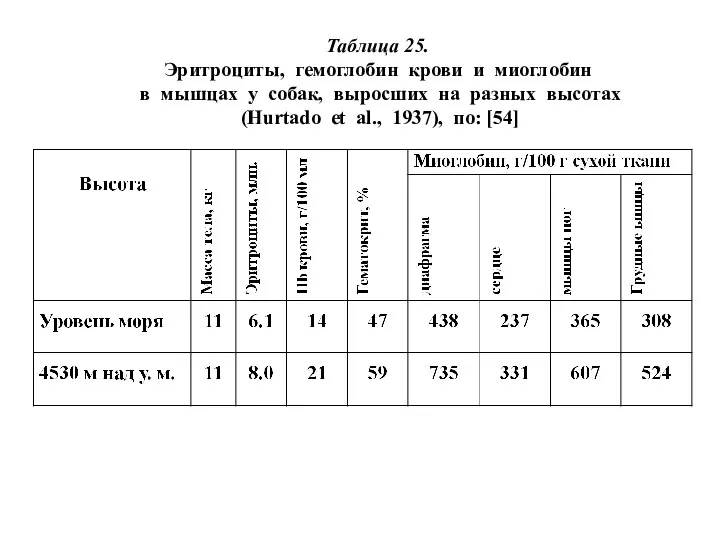
Таблица 25.
Эритроциты, гемоглобин крови и миоглобин
в мышцах у
Таблица 25.
Эритроциты, гемоглобин крови и миоглобин
в мышцах у
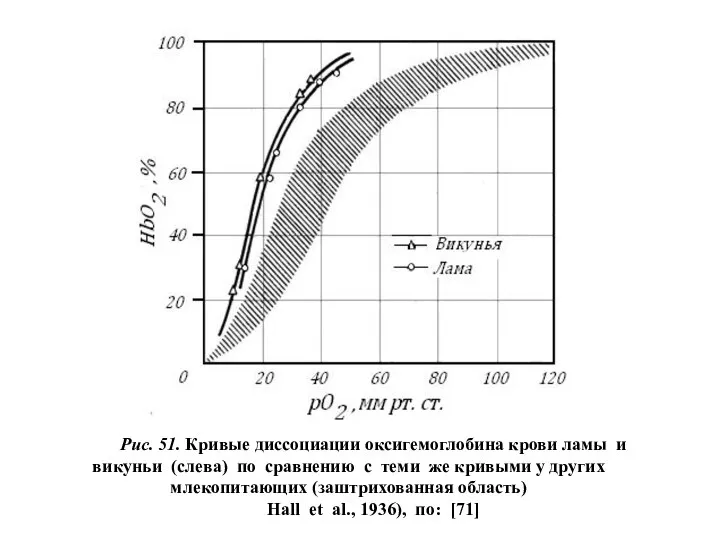
Рис. 51. Кривые диссоциации оксигемоглобина крови ламы и викуньи (слева) по
Рис. 51. Кривые диссоциации оксигемоглобина крови ламы и викуньи (слева) по
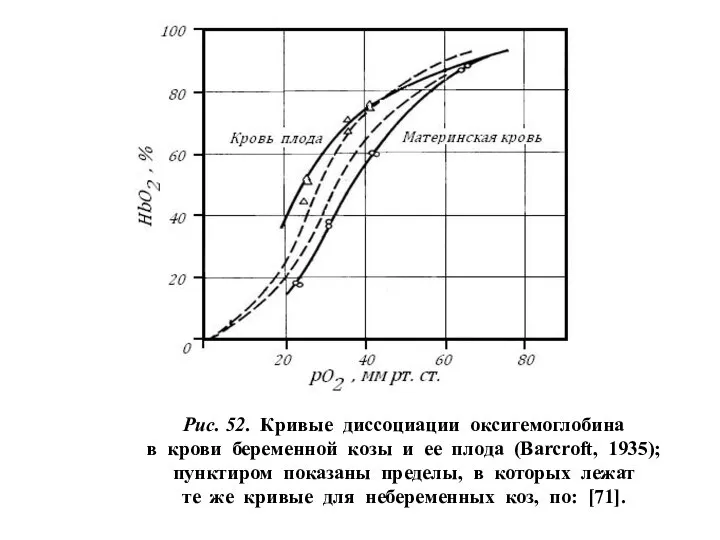
Рис. 52. Кривые диссоциации оксигемоглобина
в крови беременной козы и ее
Рис. 52. Кривые диссоциации оксигемоглобина
в крови беременной козы и ее

При адаптации к гипоксии не всегда находят
сдвиг кривой диссоциации НbО2
При адаптации к гипоксии не всегда находят
сдвиг кривой диссоциации НbО2
![Таблица 27 Параметры легких у 24 мужчин – аборигенов различных высот (Gareila-Oyola, Saldana, 1974), по: [32]](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1298731/slide-22.jpg)
Таблица 27
Параметры легких у 24 мужчин
– аборигенов различных высот
Таблица 27
Параметры легких у 24 мужчин
– аборигенов различных высот
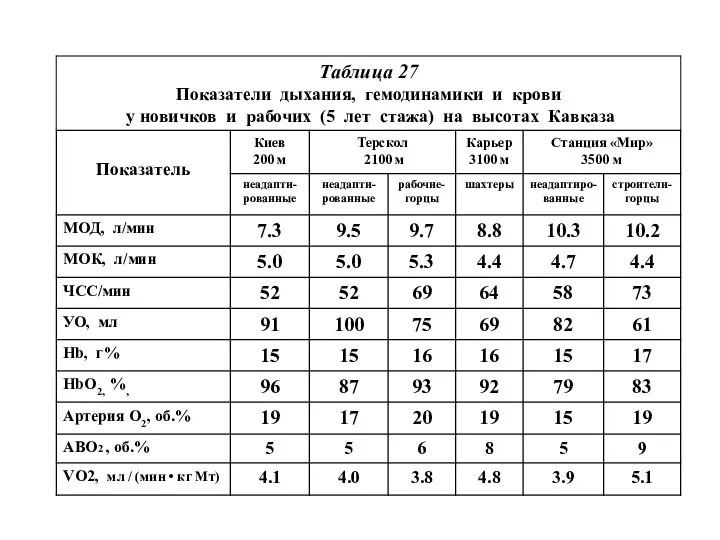


Основное различие этих двух групп рабочих
касается крови:
при длительном пребывании
Основное различие этих двух групп рабочих
касается крови:
при длительном пребывании

Почему снижается работоспособность
неадаптированного человека на высоте?
На высоте для обеспечения
Почему снижается работоспособность
неадаптированного человека на высоте?
На высоте для обеспечения

У аборигенов
исходные показатели работы легких и сердца ниже,
а поверхность
У аборигенов
исходные показатели работы легких и сердца ниже,
а поверхность

Рис. 53. Нарушение очерка
и появление грубых ошибок
после 3 мин
Рис. 53. Нарушение очерка
и появление грубых ошибок
после 3 мин

При острой дыхательной гипоксии
увеличиваются минутные объемы
дыхания и кровообращения,
доля анаэробной
При острой дыхательной гипоксии
увеличиваются минутные объемы
дыхания и кровообращения,
доля анаэробной

При длительной дыхательной гипоксии
МОК снижается даже ниже исходного уровня,
МОД
При длительной дыхательной гипоксии
МОК снижается даже ниже исходного уровня,
МОД

![Таблица 28 Содержание кислорода в организме водных млекопитающих, мл О2/кг Мт [41, 95]](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1298731/slide-32.jpg)
Таблица 28
Содержание кислорода в организме
водных млекопитающих, мл О2/кг Мт
Таблица 28
Содержание кислорода в организме
водных млекопитающих, мл О2/кг Мт
![Таблица 30. Запас кислорода в организме байкальского тюленя [37, 38] При](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1298731/slide-34.jpg)
Таблица 30.
Запас кислорода в организме байкальского тюленя [37, 38]
При подсчетах
Таблица 30.
Запас кислорода в организме байкальского тюленя [37, 38]
При подсчетах
![Рис. 55. Рентгенограмма печеночного венозного синуса и печеночных вен у морского котика [11]](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1298731/slide-35.jpg)
Рис. 55. Рентгенограмма печеночного венозного синуса
и печеночных вен у морского
Рис. 55. Рентгенограмма печеночного венозного синуса
и печеночных вен у морского
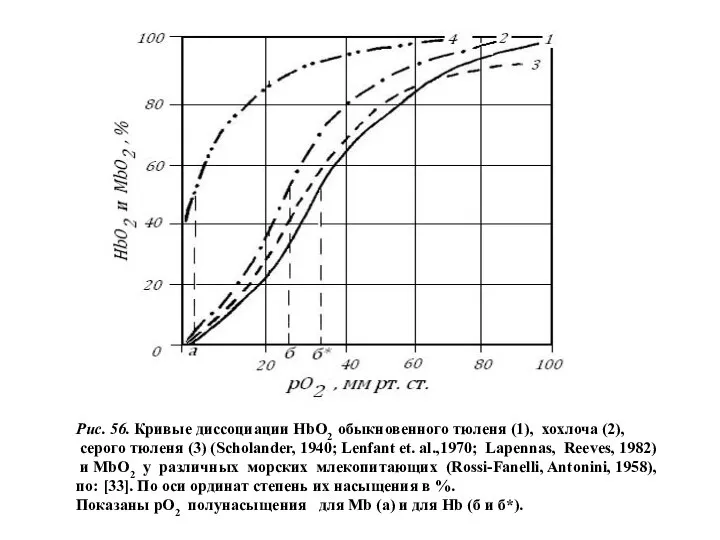
Рис. 56. Кривые диссоциации НbО2 обыкновенного тюленя (1), хохлоча (2),
серого
Рис. 56. Кривые диссоциации НbО2 обыкновенного тюленя (1), хохлоча (2),
серого

У морских млекопитающих
энергетика коротких пищевых ныряний
аэробна и обеспечена
депонированным
У морских млекопитающих
энергетика коротких пищевых ныряний
аэробна и обеспечена
депонированным
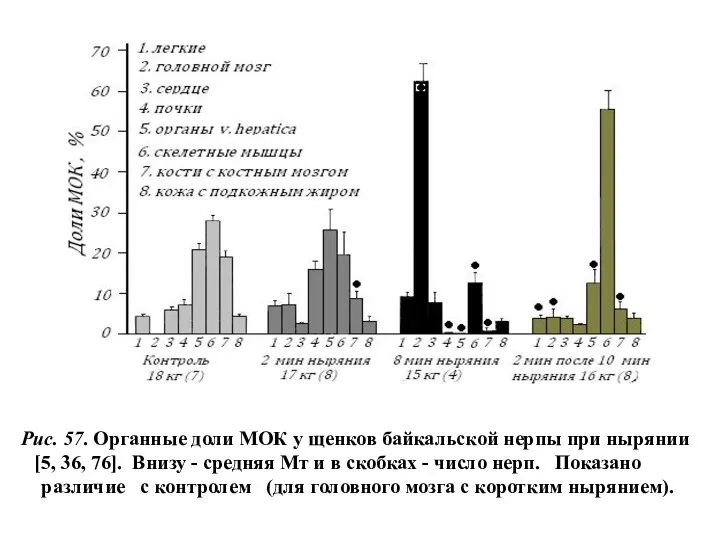
Рис. 57. Органные доли МОК у щенков байкальской нерпы при нырянии
Рис. 57. Органные доли МОК у щенков байкальской нерпы при нырянии

Морские млекопитающие после исчерпания своих кислородных депо
используют энергию анаэробного гликолиза.
Морские млекопитающие после исчерпания своих кислородных депо
используют энергию анаэробного гликолиза.
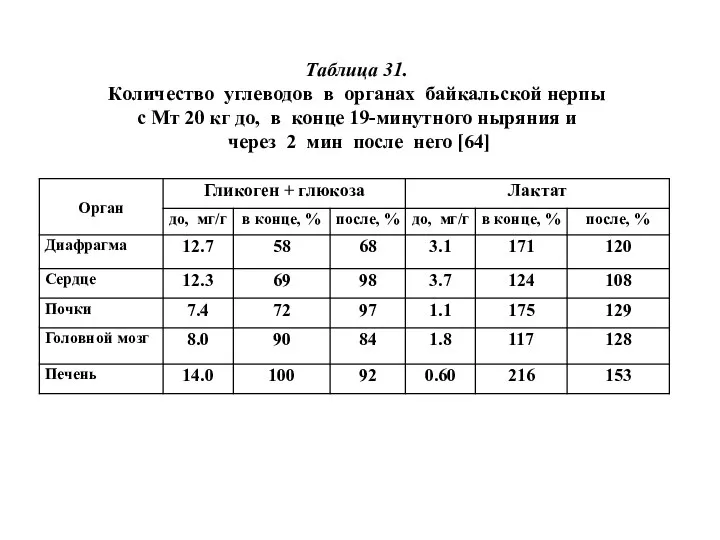
Таблица 31.
Количество углеводов в органах байкальской нерпы
с Мт 20
Таблица 31.
Количество углеводов в органах байкальской нерпы
с Мт 20

У водных млекопитающих
(по большей части это морские животные)
Энергетика
У водных млекопитающих
(по большей части это морские животные)
Энергетика
 Статистикалық ақпарат
Статистикалық ақпарат Пионерская организация
Пионерская организация Проектирования системы пожарной сигнализации и оповещения о пожаре
Проектирования системы пожарной сигнализации и оповещения о пожаре Автор: Кулакова Наталья Ивановна, учитель начальных классов 2015 год
Автор: Кулакова Наталья Ивановна, учитель начальных классов 2015 год  Особые экономические зоны России (ОЭЗ России) Подготовила студентка Житкова Екатерина 1 курса экономического фак
Особые экономические зоны России (ОЭЗ России) Подготовила студентка Житкова Екатерина 1 курса экономического фак Информационно-правовой портал Гарант. РУ
Информационно-правовой портал Гарант. РУ Связь времен в народном искусстве
Связь времен в народном искусстве МУНИЦИПАЛЬНОЕ БЮДЖЕТНОЕ ОБЩЕОБРАЗОВАТЕЛЬНОЕ УЧРЕЖДЕНИЕ «ГИМНАЗИЯ №2 г. ВЛАДИВОСТОКА»
МУНИЦИПАЛЬНОЕ БЮДЖЕТНОЕ ОБЩЕОБРАЗОВАТЕЛЬНОЕ УЧРЕЖДЕНИЕ «ГИМНАЗИЯ №2 г. ВЛАДИВОСТОКА» Бизнес-план по развитию крестьянского (фермерского) хозяйства начинающего фермера
Бизнес-план по развитию крестьянского (фермерского) хозяйства начинающего фермера ЛЕКЦИЯ № 3 АБСОЛЮТНЫЕ И ОТНОСИТЕЛЬНЫЕ ВЕЛИЧИНЫ В СТАТИСТИКЕ
ЛЕКЦИЯ № 3 АБСОЛЮТНЫЕ И ОТНОСИТЕЛЬНЫЕ ВЕЛИЧИНЫ В СТАТИСТИКЕ  Родительское собрание Воспитаем вместе Первое родительское собрание Учитель: Моисеева Ирина Александровна
Родительское собрание Воспитаем вместе Первое родительское собрание Учитель: Моисеева Ирина Александровна Проводы Масленицы
Проводы Масленицы Борьба греко-римская
Борьба греко-римская Православный компонент при изучении предмета "Основы духовно-нравственной культуры народов России"
Православный компонент при изучении предмета "Основы духовно-нравственной культуры народов России" Стандарты телефонного общения. Билайн
Стандарты телефонного общения. Билайн Керечан Ганна Іванівна - презентация для начальной школы
Керечан Ганна Іванівна - презентация для начальной школы Латинский язык 8
Латинский язык 8 СТО-ның міндеттері
СТО-ның міндеттері Парные согласные 2 класс - презентация_
Парные согласные 2 класс - презентация_ Отказ от использования гермоконтейнера
Отказ от использования гермоконтейнера Строение вируса гепатита В
Строение вируса гепатита В ДЕЛОВАЯ ПЕРЕПИСКА ЯЗЫК И СТИЛЬ
ДЕЛОВАЯ ПЕРЕПИСКА ЯЗЫК И СТИЛЬ «Стресс в подростковом возрасте»
«Стресс в подростковом возрасте» Книга и творчество Презентация к библиотечному уроку по теме «Художники-иллюстраторы» для 3 класса
Книга и творчество Презентация к библиотечному уроку по теме «Художники-иллюстраторы» для 3 класса План работы молодежного общественного совета при избирательной комиссии Краснодарского края
План работы молодежного общественного совета при избирательной комиссии Краснодарского края Технология несъемной опалубки
Технология несъемной опалубки Средства наблюдения
Средства наблюдения Контроллер клавиатуры
Контроллер клавиатуры